Effect of Rice Flour Fermentation with Lactobacillus spicheri DSM 15429 on the Nutritional Features of Gluten-Free Muffins

 , ,
, ,  , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
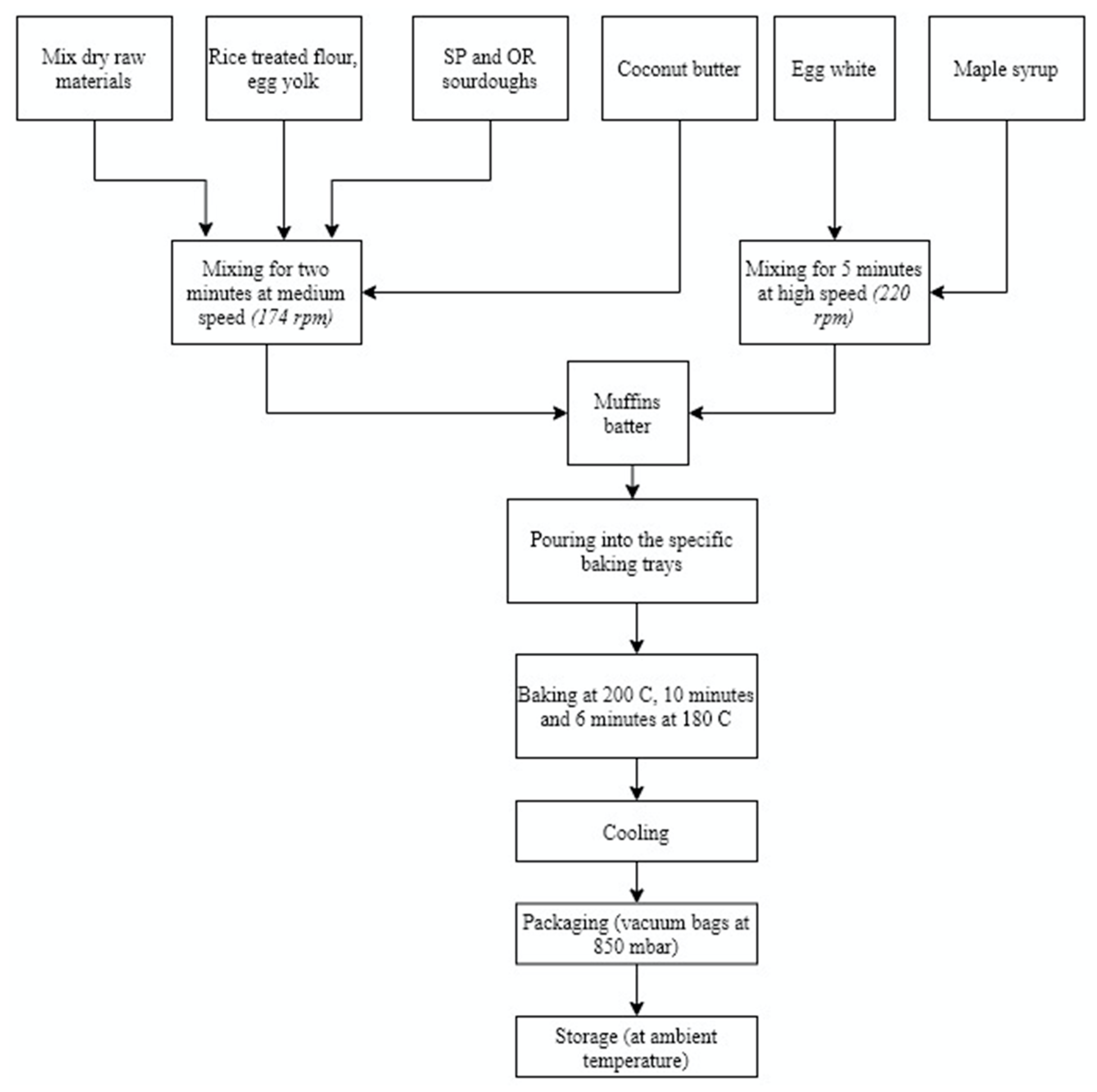
2.2. Microbial Starter Culture Preparation, Sourdough Preparation, and Muffins Formulation
2.3. Amino-Acid Determination
2.4. Minerals Content
2.5. Lactic Acid Determination
2.6. Total Phenols Assay by Folin–Ciocalteau Reagent
2.7. Antioxidant Capacity by DPPH Assay
2.8. pH Determination
2.9. Microbiological Analysis of Gluten-Free Muffins
Total Number of Yeast and Molds Determination
2.10. Statistical Analysis
3. Results
3.1. Amino Acids, Mineral, Total Phenols Content, and Radical Scavenging Activity of RWF
3.1.1. Amino-Acids Content of the RWF
3.1.2. Mineral Content of RWF
3.1.3. Total Phenols and Radical Scavenging Activity of RWF
3.2. Amino-Acids, Mineral, Total Phenols Content, and Radical Scavenging Activity of OR and SP Sourdoughs
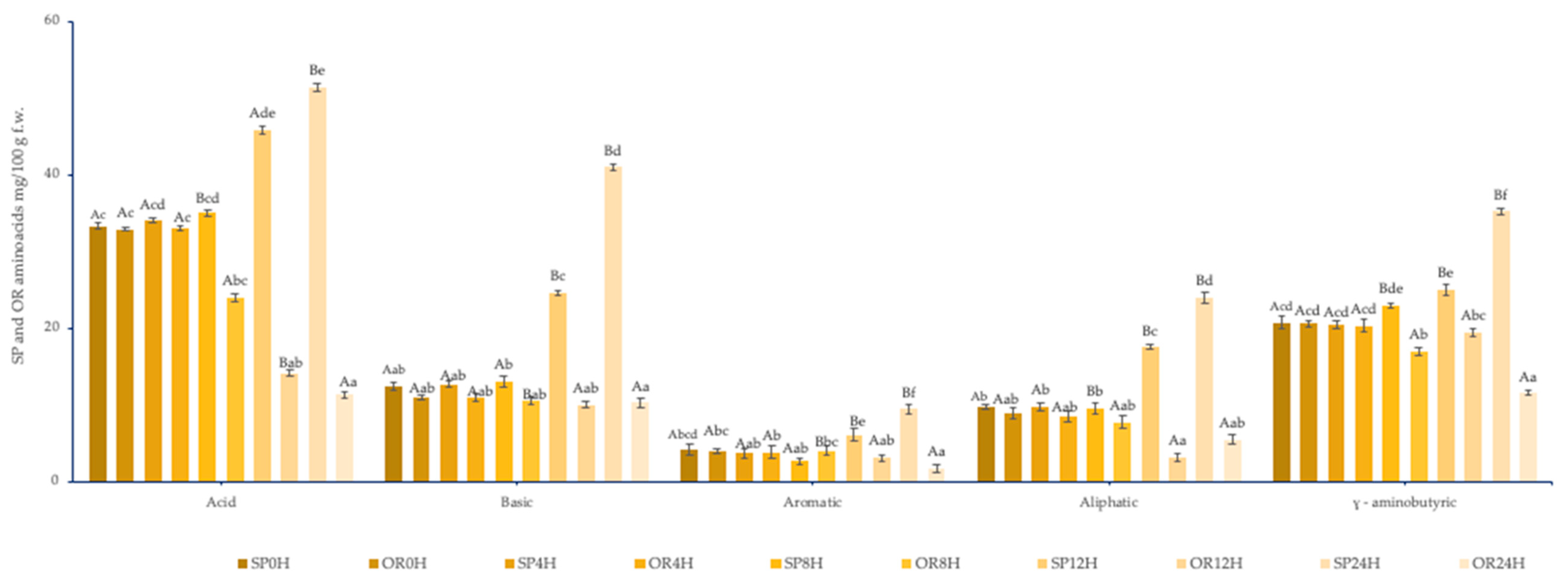
3.2.1. Amino-Acids Content of SP and OR Sourdoughs
3.2.2. Mineral Content of SP and OR Sourdoughs
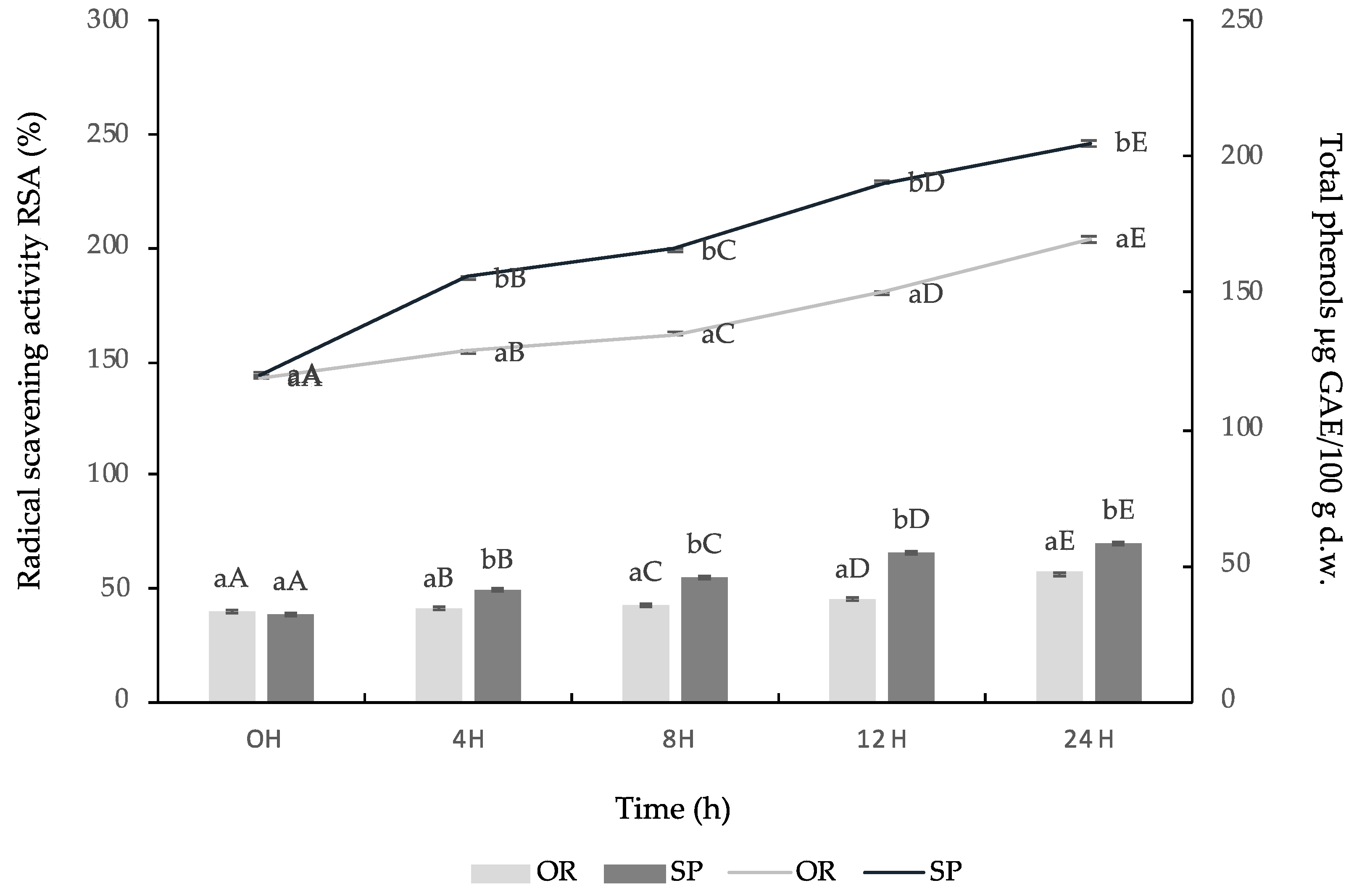
3.2.3. Total Phenolic Content and Antioxidant Activity of OR and SP Sourdough
3.3. Amino-Acids, Mineral, Total Phenols, Radical Scavenging Activity, and Lactic Acid Content of Final Baked Muffins
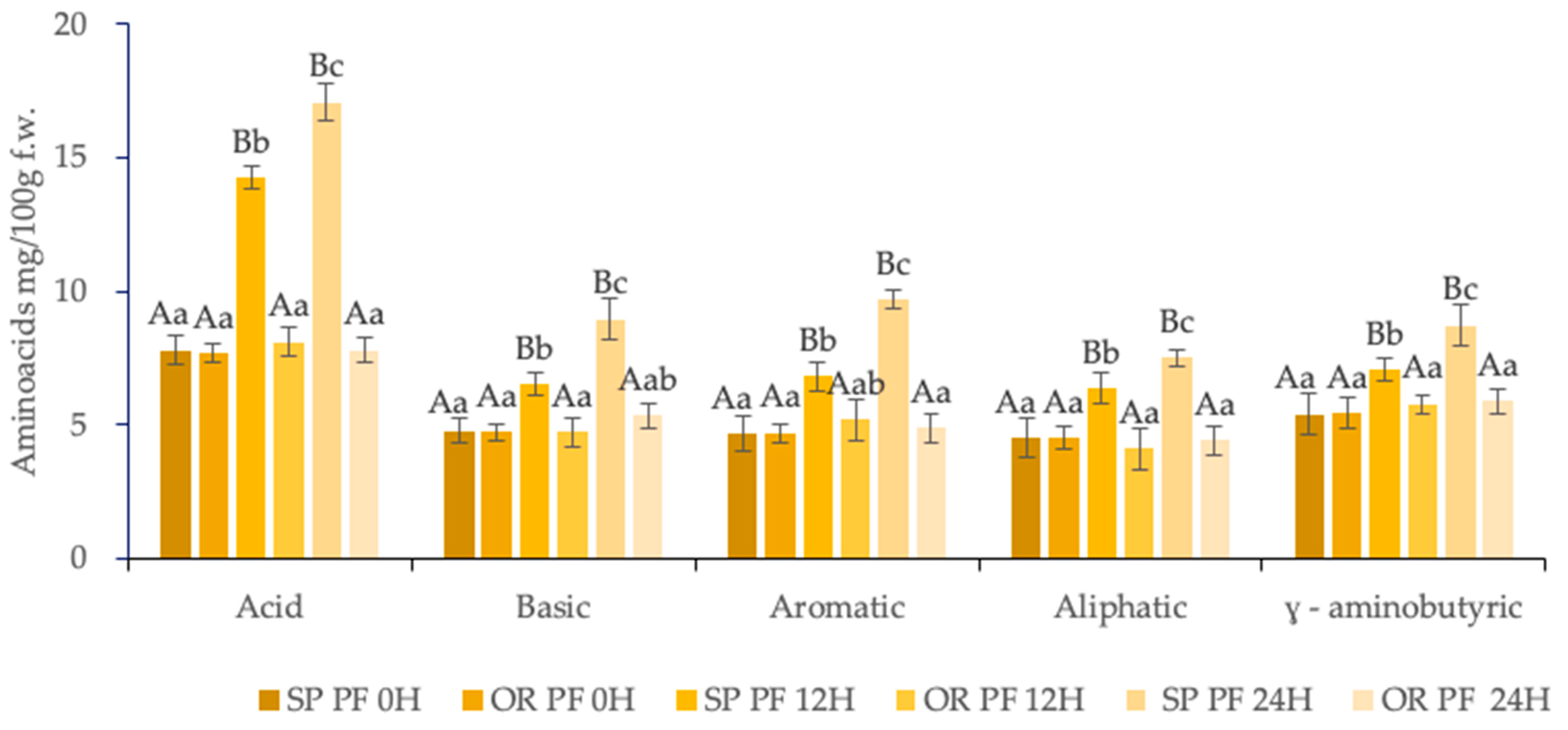
3.3.1. Amino-Acids Content of Final Baked Muffins
3.3.2. Minerals Final Baked Muffins Content
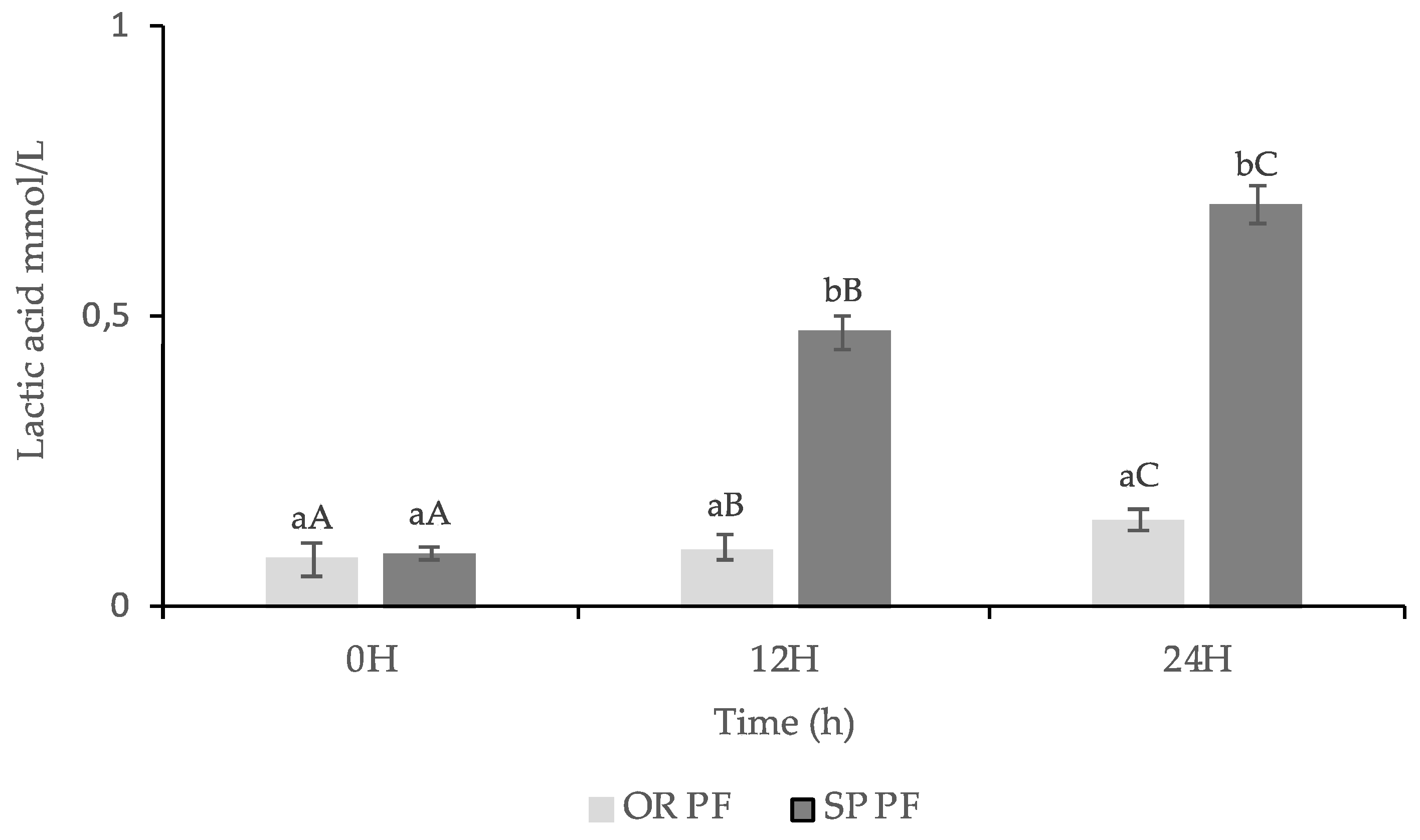
3.3.3. Lactic Acid Content
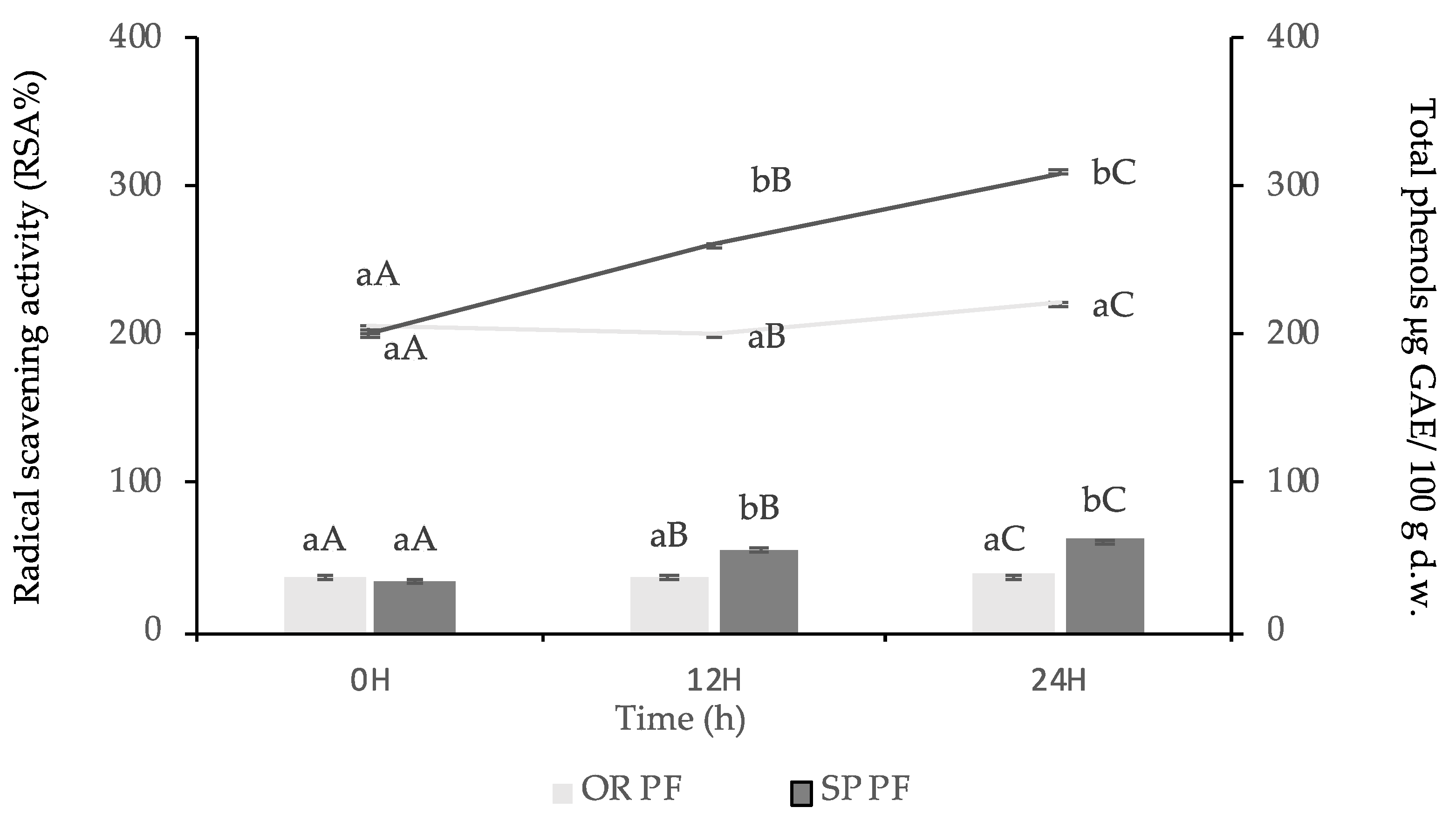
3.3.4. Total Phenolic Content and Antioxidant Activity of Final Baked Goods
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Codex Alimentarius. CODEX STAN 118-1979: Standard for Foods for Special Dietary Use for Persons Intolerant to Gluten. Available online: http://www.fao.org/fao-who-codexalimentarius/standards/en/ (accessed on 15 March 2020).
- Aponte, M.; Boscaino, F.; Sorrentino, A.; Coppola, R.; Masi, P.; Romano, A. Volatile compounds and bacterial community dynamics of chestnut-flour-based sourdoughs. Food Chem. 2013, 141, 2394–2404. [Google Scholar] [CrossRef] [PubMed]
- Ziobro, R.; Juszczak, L.; Witczak, M.; Korus, J. Non-gluten proteins as structure forming agents in gluten free bread. J. Food Sci. Technol. 2015, 53, 571–580. [Google Scholar] [CrossRef] [Green Version]
- Theethira, T.G.; Dennis, M.; Leffler, D.A.; Daniel, A. Nutritional consequences of celiac disease and the gluten-free diet. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Wen, H.; Li, C.; Gu, Z. Differences in functional properties and biochemical characteristics of congenetic rice proteins. J. Cereal Sci. 2009, 50, 184–189. [Google Scholar] [CrossRef]
- Singh, A.; Matta, N. Disulphide linkages occur in many polypeptides of rice protein fractions: A two-dimensional gel electrophoretic study. Rice Sci. 2011, 18, 86–94. [Google Scholar] [CrossRef]
- Chan, Y.J.; Lu, W.C.; Lin, H.Y.; Wu, Z.R.; Liou, C.W.; Li, P.H. Effect of Rice Protein Hydrolysates as an Egg Replacement on the Physicochemical Properties of Flaky Egg Rolls. Foods 2020, 9, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilowefah, M.; Chinma, C.; Bakar, J.; Hasanah, M.; Ghazali, H.M.; Muhammad, K.; Makeri, M. Fermented Brown Rice Flour as Functional Food Ingredient. Foods 2014, 3, 149–159. [Google Scholar] [CrossRef]
- Chan, K.W.; Khong, N.M.H.; Iqbal, S.; Ismail, M. Simulated Gastrointestinal pH Condition Improves Antioxidant Properties of Wheat and Rice Flours. Int. J. Mol. Sci. 2012, 13, 7496–7507. [Google Scholar] [CrossRef] [Green Version]
- Gujral, H.S.; Guardiola, I.; Carbonell, J.V.; Rosell, C.M. Effect of cyclodextrinase on dough rheology and bread quality from rice flour. J. Agric. Food Chem. 2003, 51, 3814–3818. [Google Scholar] [CrossRef]
- Mert, S.; Sahin, S.; Sumnu, G. Development of Gluten-Free Wafer Sheet Formulations. Food Sci. Technol. 2015, 63, 1121–1127. [Google Scholar] [CrossRef]
- Monk, J.L.M.; Vanier, N.L.; Casaril, J.; Berto, R.M.; De Oliveira, M.; Gomes, C.B.; De Carvalho, M.P.; Guerra Dias, A.R.; Elias, M.C. Effects of milling on proximate composition, folic acid, fatty acids and technological properties of rice. J. Food Compos. Anal. 2013, 30, 73–79. [Google Scholar] [CrossRef]
- Păucean, A.; Man, S.; Muste, S.; Pop, A. Effect of quinoa flour addition on quality characteristics of rice gluten-free cookies. J. Agroaliment. Process. Technol. 2015, 21, 371–378. [Google Scholar]
- Perera, I.; Seneweera, S.; Hirotsu, N. Manipulating the Phytic Acid Content of Rice Grain Toward Improving Micronutrient Bioavailability. Rice 2018, 11, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogelmann, S.A.; Seitter, M.; Singer, U.; Brandt, M.J.; Hertel, C. Adaptability of lactic acid bacteria and yeasts to sourdoughs prepared from cereals, pseudocereals and cassava and use of competitive strains as starters. Int. J. Food Microbiol. 2009, 130, 205–212. [Google Scholar] [CrossRef]
- Coda, R.; Di Cagno, R.; Gobbetti, M.; Rizzello, C.G. Sourdough lactic acid bacteria: Exploration of non-wheat cereal-based fermentation. Food Microbiol. 2014, 37, 51–58. [Google Scholar] [CrossRef]
- Lee, S.M.; Hwang, Y.R.; Kim, M.S.; Chung, M.S.; Kim, Y.S. Comparison of Volatile and Nonvolatile Compounds in Rice Fermented by Different Lactic Acid Bacteria. Molecules 2019, 24, 1183. [Google Scholar] [CrossRef] [Green Version]
- Chis, M.S.; Păucean, A.; Man, S.M.; Muresan, V.; Socaci, S.A.; Pop, A.; Stan, L.; Rusu, B.; Muste, S. Textural and Sensory Features Changes of Gluten Free Muffins Based on Rice Sourdough Fermented with Lactobacillus spicheri DSM 15429. Foods 2020, 9, 363. [Google Scholar] [CrossRef] [Green Version]
- Balli, D.; Bellumori, M.; Paoli, P.; Pieraccini, G.; Di Paola, M.; De Filippo, C.; Di Gioia, D.; Mulinacci, N.; Innocenti, M. Study on a Fermented Whole Wheat: Phenolic Content, Activity on PTP1B Enzyme and In Vitro Prebiotic Properties. Molecules 2019, 24, 1120. [Google Scholar] [CrossRef] [Green Version]
- Ilowefah, M.; Bakar, J.; Ghazali, H.M.; Mediani, A.; Muhammad, K. Physicochemical and functional properties of yeast fermented brown rice flour. J. Food Sci. Technol. 2014, 52, 5534–5545. [Google Scholar] [CrossRef] [Green Version]
- Teleky, B.E.; Martău, A.G.; Ranga, F.; Chețan, F.; Vodnar, D.C. Exploitation of Lactic Acid Bacteria and Baker’s Yeast as Single or Multiple Starter Cultures of Wheat Flour Dough Enriched with Soy Flour. Biomolecules 2020, 10, 778. [Google Scholar] [CrossRef]
- Biscola, V.; Albuquerque, M.A.C.; Nunes, T.P.; Vieira, A.D.S.; Franco, D.G.M. Lactic Acid Bacteria: A Functional Approach, 1st ed.; Albuquerque, M.A.C., LeBlanc, A.M., LeBlanc, J.G., Bedani, R., Eds.; Crc Press (Taylor and Francis Group): Boca Raton, FL, USA, 2020; p. 292. [Google Scholar]
- Maidana, D.S.; Finch, S.; Garro, M.; Savoy, G.; Ganzle, M.; Vignolo, G. Development of gluten-free breads started with chia and flaxseed sourdoughs fermented by selected lactic acid bacteria. LWT Food Sci. Technol. 2020, 109189. [Google Scholar] [CrossRef]
- Meroth, C.B.; Hammes, W.P.; Hertel, C. Characterisation of the microbiota of rice sourdoughs and description of Lactobacillus spicheri sp. nov. Syst. Appl. Microbiol. 2004, 27, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Bourekoua, H.; Benatallah, L.; Zidoune, M.N.; Rosell, C.M. Developing gluten free bakery improvers by hydrothermal treatment of rice and corn flours. Lwt Food Sci. Technol. 2016, 73, 342–350. [Google Scholar] [CrossRef] [Green Version]
- Bobis, O.; Dezmirean, D.S.; Mărghitaș, L.A.; Bonta, V.; Urcan, A.; Pașca, C.; Moise, A.R. Morus sp. for revigorating silkworm breeding in Romania and promoting health benefits of leaves and fruits. Sci. Papers. Series B Hortic. 2018, LXII, 211–216. [Google Scholar]
- Chiş, M.S.; Păucean, A.; Stan, L.; Suharoschi, R.; Man, S.M.; Muste, S. Protein metabolic conversion of nutritional features during quinoa sourdough fermentation and its impact on baked goods. Cyta J. Food 2018, 280, 744–753. [Google Scholar] [CrossRef]
- Păucean, A.; Moldovan, O.P.; Mureşan, V.; Socaci, S.A.; Dulf, F.; Man, M.S.; Mureşan, A.E.; Muste, S. Folic acid, minerals, amino-acids, fatty acids and volatile compounds of green and red lentils. Folic acid content optimization in wheat-lentils composite flours. Chem. Cent. J. 2018, 12, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mihoc, M.; Pop, G.; Alexa, E.; Radulov, I. Nutritive quality of romanian hemp varieties (Cannabis sativa L.) with special focus on oil and metal contents of seeds. Chem. Cent. J. 2012, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Păucean, A.; Vodnar, D.C.; Socaci, S.A.; Socaciu, C. Carbohydrate metabolic conversions to lactic acid and volatile derivatives, as influenced by Lactobacillus plantarum ATCC 8014 and Lactobacillus casei ATCC 393 efficiency during in vitro and sourdough fermentation. Eur. Food Res. Technol. 2013, 237, 679–689. [Google Scholar] [CrossRef]
- Bunea, A.; Ruginǎ, D.O.; Pintea, A.M.; Sconţa, Z.; Bunea, C.I.; Socaciu, C. Comparative polyphenolic content and antioxidant activities of some wild and cultivated blueberries from Romania. Not. Bot. Horti Agrobot. 2011, 39, 70–76. [Google Scholar] [CrossRef] [Green Version]
- Păucean, A.; Man, S.M.; Chis, M.S.; Mureşan, V.; Pop, C.R.; Socaci, S.A.; Mureşan, C.C.; Muste, S. Use of pseudocereals preferment made with aromatic yeast strains for enhancing wheat bread quality. Foods 2019, 8, 443. [Google Scholar] [CrossRef] [Green Version]
- Dordević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of fermentation on antioxidant properties of some cereals and pseudo cereals. Food Chem. 2010, 119, 957–963. [Google Scholar] [CrossRef]
- Chiş, M.S.; Păucean, A.; Stan, L.; Mureşan, V.; Vlaic, R.A.; Man, S.; Biriş-Dorhoi, S.E.; Muste, S. Lactobacillus plantarum ATCC 8014 in quinoa sourdough adaptability and antioxidant potential. Rom. Biotech. Lett. 2018, 23, 13581–13591. [Google Scholar]
- International Organization for Standardization. International Standard ISO 21527-2, 1st ed.; International Organization for Standardization: Geneva, Switzerland, 2008. [Google Scholar]
- Antoniewska, A.; Rutkowska, J.; Pineda, M.M.; Adamska, A. Antioxidative, nutritional and sensory properties of muffins with buckwheat flakes and amaranth flour blend partially substituting for wheat flour. Lwt Food Sci. Technol. 2018, 89, 217–223. [Google Scholar] [CrossRef]
- Nagy, M.; Semeniuc, C.A.; Socaci, A.M.; Pop, C.R.; Rotar, A.M.; Sălagean, C.D.; Tofană, M. Utilization of brewer’s spent grain and mushrooms in fortification of smoked sausages. Food Sci. Technol. 2017, 37, 315–320. [Google Scholar] [CrossRef] [Green Version]
- Kati, K.; Kaisa, P.; Karin, A. Influence and Interactions of Processing Conditions and Starter Culture on Formation of Acids, Volatile Compounds, and Amino Acids in Wheat Sourdoughs. Cereal Chem. 2004, 81, 598–610. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. Composition and protein profile analysis of rice protein ingredients. J. Food Compos. Anal. 2017, 59, 18–26. [Google Scholar] [CrossRef]
- Liang, J.; Han, B.; Han, L.; Nout, M.J.R.; Hamer, R.J. Iron, zinc and phytic acid content of selected rice varieties from China. J. Sci. Food Agric. 2007, 510, 504–510. [Google Scholar] [CrossRef]
- Oko, A.O.; Ubi, B.E.; Efisue, A.A.; Dambaba, N. Comparative Analysis of the Chemical Nutrient Composition of Selected Local and Newly Introduced Rice Varieties Grown in Ebonyi State of Nigeria. Int. J. Agric. For. 2012, 2, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Verma, D.K.; Srivastav, P.P. Proximate Composition, Mineral Content and Fatty Acids Analyses of Aromatic and Non-Aromatic Indian Rice. Rice Sci. 2017, 24, 21–31. [Google Scholar] [CrossRef]
- Mbatchou, V.C.; Dawda, S. The Nutritional Composition of Four Rice Varieties Grown and Used in Different Food Preparations in Kassena-Nankana District, Ghana. Int. J. Res. Chem. Environ. 2013, 3, 308–315. [Google Scholar]
- Heinemann, R.J.B.; Fagundes, P.L.; Pinto, E.A.; Penteado, M.V.C. Comparative study of nutrient composition of commercial brown, parboiled and milled rice from Brazil. J. Food Compos. Anal. 2015, 18, 287–296. [Google Scholar] [CrossRef]
- Pang, Y.; Ahmed, S.; Xu, Y.; Beta, T.; Zhu, Z.; Shao, Y. Bound phenolic compounds and antioxidant properties of whole grain and bran of white, red and black rice. Food Chem. 2018, 240, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Sakač, M.; Pestorić, M.; Mišan, A.; Nedeljković, N.; Jambrec, D.; Jovanov, P.; Banjac, V.; Torbica, A.; Hadnađev, M.; Mandić, A. Antioxidant capacity, mineral content and sensory properties of gluten-free rice and buckwheat cookies. Food Technol. Biotechnol. 2015, 53, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Butsat, S.; Siriamornpun, S. Antioxidant capacities and phenolic compounds of the husk, bran and endosperm of Thai rice. Food Chem. 2010, 119, 606–613. [Google Scholar] [CrossRef]
- Gorinstein, S.; Vargas, O.J.M.; Jaramillo, N.O.; Sallas, I.A.; Ayala, A.L.M.; Arancibia-Avila, P.; Toledo, F.; Katrich, E.; Trakhtenberg, S. The total polyphenols and the antioxidant potentials of some selected cereals and pseudocereals. Eur. Food Res. Technol. 2007, 225, 321–328. [Google Scholar] [CrossRef]
- Fitriani, D.R.; Rumpagaporn, P. Antioxidant Activity of Enzymatically Treated Extracts from Commercially Defatted Rice Bran. Agricultural Sciences: Leading Thailand to World Class Standards. In Proceedings of the 52nd Kasetsart University Annual Conference, Bangkok, Thailand, 4–7 February 2014. [Google Scholar]
- Lloyd, B.J.; Siebenmorgen, T.J.; Beers, K.W. Effects of commercial processing on antioxidants in rice bran. Cereal Chem. 2000, 77, 551–555. [Google Scholar] [CrossRef] [Green Version]
- Thiele, C.; Gänzle, M.G.; Vogel, R.F. Contribution of sourdough lactobacilli, yeast, and cereal enzymes to the generation of amino acids in dough relevant for bread flavor. Cereal Chem. 2002, 79, 45–51. [Google Scholar] [CrossRef]
- Dallagnol, A.M.; Pescuma, M.; De Valdez, G.E.; Rollán, G. Fermentation of quinoa and wheat slurries by Lactobacillus plantarum CRL 778: Proteolytic activity. Appl. Microbiol. Biotechnol. 2013, 97, 3129–3140. [Google Scholar] [CrossRef]
- Ruiz-Rodríguez, L.; Bleckwedel, J.; Ortiz, M.E.; Pescuma, M.; Mozzi, F. Lactic Acid Bacteria, Industrial Biotechnology: Microorganisms, 1st ed.; Wittmann, C., Liao, J.C., Eds.; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2016; Volume 3, pp. 395–451. [Google Scholar] [CrossRef]
- Papadimitriou, K.; Zoumpopoulou, G.; Georgalaki, M.; Alexandraki, V.; Kazou, M.; Anastasiou, R.; Tsakalidou, E. Sourdough Bread, Innovations in Traditional Foods; Galanakis, C.M., Ed.; Woodhead Publishing: Cambridge, UK, 2019; pp. 127–158. [Google Scholar] [CrossRef]
- Mamhoud, A.; Nionelli, L.; Bouzaine, T.; Hamdi, M.; Gobbetti, M.; Rizzello, C.G. Selection of lactic acid bacteria isolated from tunisian cereals and exploitation of the use as starters for sourdough fermentation. Int. J. Food Microbiol. 2016, 225, 9–19. [Google Scholar] [CrossRef]
- Coda, R.; Rizzello, C.G.; Gobbetti, M. Use of sourdough fermentation and pseudo-cereals and leguminous flours for the making of a functional bread enriched of -aminobutyric acid (GABA). Int. J. Food Microbiol. 2010, 137, 236–245. [Google Scholar] [CrossRef]
- Gänzle, M.G. Enzymatic and bacterial conversions during sourdough fermentation. Food Microbiol. 2014, 37, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; Rizzello, C.G.; De Angelis, M.; Cassone, A.; Giuliani, G.; Benedusi, A.; Limitone, A.; Surico, M.F.; Gobbetti, M. Use of selected sourdough strains of Lactobacillus for removing gluten and enhancing the nutritional properties of gluten-free bread. J. Food Prot. 2008, 71, 1491–1495. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Pontonio, E.; Waters, D.M.; Arendt, E.K. Applications of microbial fermentations for production of gluten-free products and perspectives. Appl. Microbiol. Biotechnol. 2012, 93, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.; Zúñiga, M. Amino acid catabolic pathways of lactic acid bacteria. Crit. Rev. Microbiol. 2006, 32, 155–183. [Google Scholar] [CrossRef]
- Blandino, A.; Al-Aseeri, M.E.; Pandiella, S.S.; Cantero, D.; Webb, C. Cereal-based fermented foods and beverages. Food Res. Int. 2003, 36, 527–543. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Corsetti, A.; Di Cagno, R.; Calasso, R.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- Carrizo, S.L.; Montes de Oca, C.E.; Laiño, J.E.; Suarez, N.E.; Vignolo, G.; LeBlanc, J.G.; Rollán, G. Ancestral Andean grain quinoa as source of lactic acid bacteria capable to degrade phytate and produce B-group vitamins. Food Res. Int. 2016, 89, 488–494. [Google Scholar] [CrossRef]
- Kumari, S.; Bhinder, S.; Singh, B.; Kaur, A.; Singh, N. Effect of buckwheat incorporation on batter fermentation, rheology, phenolic, amino acid composition and textural properties of idli. LWT Food Sci. Technol. 2020, 122, 1–8. [Google Scholar] [CrossRef]
- Montemurro, M.; Coda, R.; Rizzello, C.G. Recent Advances in the Use of SourdoughBiotechnology in Pasta Making. Foods 2019, 8, 129. [Google Scholar] [CrossRef] [Green Version]
- Sharma, N.; Angural, S.; Rana, M.; Puri, N.; Kondepudi, K.K.; Gupta, N. Phytase producing lactic acid bacteria: Cell factories for enhancing micronutrient bioavailability of phytate rich foods. Trends Food Sci. Technol. 2020, 96, 1–12. [Google Scholar] [CrossRef]
- Nuobariene, L.; Cizeikiene, D.; Gradzeviciute, E.; Hansen, A.E.; Rasmussen, S.K.; Juodeikiene, G.; Vogensen, F.K. Phytase-active lactic acid bacteria from sourdoughs: Isolation and identification. LWT Food Sci. Technol. 2015, 63, 766–772. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Nionelli, L.; Coda, R.; De Angelis, M.; Gobbetti, M. Effect of sourdough fermentation on stabilisation, and chemical and nutritional characteristics of wheat germ. Food Chem. 2010, 119, 1079–1089. [Google Scholar] [CrossRef]
- Qiu, Y.; Liu, Q.; Beta, T. Antioxidant properties of commercial wild rice and analysis of soluble and insoluble phenolic acids. Food Chem. 2010, 121, 140–147. [Google Scholar] [CrossRef]
- Antognoni, F.; Mandrioli, R.; Potente, G.; Taneyo Saa, D.L.; Gianotti, A. Changes in carotenoids, phenolic acids and antioxidant capacity in bread wheat doughs fermented with different lactic acid bacteria strains. Food Chem. 2019, 292, 211–216. [Google Scholar] [CrossRef]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y. Enhanced Antioxidant Activity for Apple Juice. Molecules 2019, 24, 51. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Lorusso, A.; Russo, V.; Pinto, D.; Marzani, B.; Gobbetti, M. Improving the antioxidant properties of quinoa flour through fermentation with selected autochthonous lactic acid bacteria. Int. J. Food Microbiol. 2017, 241, 252–261. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Bhanwar, S.; Bamnia, M.; Ghosh, M.; Ganguli, A. Use of Lactococcus lactis to enrich sourdough bread with γ-aminobutyric acid. Int. J. Food Sci. Nutr. 2013, 64, 77–81. [Google Scholar] [CrossRef]
- Venturi, M.; Galli, V.; Pini, N.; Guerrini, S.; Granchi, L. Use of selected lactobacilli to increase γ-Aminobutyric acid (GABA) content in sourdough bread enriched with amaranth flour. Foods 2019, 8, 218. [Google Scholar] [CrossRef] [Green Version]
- Ashaolu, T.J. Safety and quality of bacterially fermented functional foods and beverages: A mini review. Food Qual. Saf. 2020, 20, 1–5. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food. Res. Int. 2016, 89, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Rebellato, A.P.; Pacheco, B.C.; Prado, J.P.; Lima Pallone, J.A. Iron in fortified biscuits: A simple method for its quantification, bioaccessibility study and physicochemical quality. Food Res. Int. 2015, 77, 385–391. [Google Scholar] [CrossRef] [Green Version]
- Abosede, A.M.; Ifesan, B.O.T.; Enujiugha, V.N.; Adefisola, B.A. Microbiological and Physicochemical Properties of Wholegrain Millet Sourdough Breads. Int. J. Food Sci. Nutr. 2019, 4, 74–82. [Google Scholar]
- Lopez, H.W.; Duclos, V.; Coudray, C.; Krespine, V.; Feillet-Coudray, C.; Messager, A.; Demigne’, C.; Remesy, C. Making bread with sourdough improves mineral bioavailability from reconstituted whole wheat flour in rats. Nutrition 2013, 19, 524–530. [Google Scholar] [CrossRef]
- Katina, K. Sourdough: A Tool for the Improved Flavour, Texture and Shelf-Life of Wheat Bread, Academic Dissertation; University of Helsinki: Helsinki, Finland, 2005. [Google Scholar]
- Ross, R.P.; Morgan, S.; Hill, C. Preservation and fermentation: Past, present and future. Int. J. Food Microbiol. 2002, 79, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Axel, C.; Roker, B.; Brosnan, B.; Zannini, E.; Furey, A.; Coffey, A.; Arendt, E.K. Application of Lactobacillus amylovorus DSM19280 in gluten-free sourdough bread to improve the microbial shelf life. Food Microbiol. 2015, 47, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Alkay, Z.; Kilmanoğlu, H.; Durak, M.Z. Prevention of Sourdough Bread Mould Spoliage by antifungal Lactic Acid Bacteria Fermentation. Eur. J. Sci. Technol. 2020, 18, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Rizzello, C.G.; Lorusso, A.; Montemurro, M.; Gobbetti, M. Use of sourdough made with quinoa (Chenopodium quinoa) flour and autochthonous selected lactic acid bacteria for enhancing the nutritional, textural and sensory features of white bread. Food Microbiol. 2016, 56, 1–13. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef]
- Verni, M.; Verardo, V.; Rizzello, C.G. How Fermentation Affects the Antioxidant Properties of Cereals and Legumes. Foods 2019, 8, 362. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acids | RWF (mg/100 g f.w.) | |
|---|---|---|
| Acid group | Glutamine | 14.11 ± 0.12 |
| Asparagine | 25.53 ± 0.31 | |
| Glutamic acid | 32.22 ± 0.11 | |
| Aspartic acid | 37.05 ± 0.21 | |
| Basic group | Histidine | 1.92 ± 0.10 |
| Lysine | 5.50 ± 0.13 | |
| Ornithine | 0.74 ± 0.09 | |
| Arginine | 11.12 ± 0.13 | |
| Aromatic group | Tryptophan | 1.74 ± 0.17 |
| Tyrosine | 2.63 ± 0.14 | |
| Phenylalanine | 1.50 ± 0.21 | |
| Aliphatic group | Prolamine | 3.10 ± 0.13 |
| Isoleucine | 0.88 ± 0.07 | |
| Leucine | 1.84 ± 0.05 | |
| Valine | 1.95 ± 0.03 | |
| Alanine | 7.61 ± 0.04 | |
| Glycine | 2.28 ± 0.02 | |
| γ-aminobutyric group | Proline | 3.10 ± 0.03 |
| Threonine | 2.61 ± 0.02 | |
| Serine | 6.20 ± 0.02 | |
| γ-aminobutyric acid | 12.55 ± 0.5 | |
| ∑Acid group | 108.91 ± 0.75 | |
| ∑Basic group | 19.38 ± 0.45 | |
| ∑Aromatic group | 5.87 ± 0.52 | |
| ∑Aliphatic group | 17.66 ± 0.34 | |
| ∑γ-aminobutyric group | 24.46 ± 0.87 | |
| Parameters | RWF |
|---|---|
| Minerals, mg/100 g f.w. | |
| Ca | 15.02 ± 0.20 |
| Mg | 231.36 ± 0.47 |
| K | 437.18 ± 0.26 |
| Fe | 1.21 ± 0.14 |
| Cu | 0.35 ± 0.01 |
| Zn | 0.97 ± 0.04 |
| Mn | 1.14 ±0.02 |
| Cr | 0.016 ± 0.01 |
| Samples | Ca | Mg | K | Fe | Cu | Zn | Mn |
|---|---|---|---|---|---|---|---|
| OR 0H | 7.92 ± 0.04 Aa | 50.00 ± 0.23 Aa | 200.32 ± 0.56 Aa | 0.42 ± 0.03 Aa | 0.09 ± 0.02 Abc | 0.39 ± 0.02 Aa | 0.79 ± 0.03 Ad |
| SP 0H | 7.90 ± 0.31 Aa | 50.01 ± 0.12 Aa | 199.32 ± 0.19 Aa | 0.42 ± 0.02 Aa | 0.08 ± 0.01 Aa | 0.39 ± 0.01 Aa | 0.79 ± 0.02 Ad |
| OR 4H | 8.21 ± 0.23 Aab | 55.80 ± 0.32 Ab | 205.50 ± 0.45 Aa | 0.44 ± 0.01 Aab | 0.12 ± 0.01 Abc | 0.43 ± 0.02 Abc | 0.29 ± 0.02 Ab |
| SP 4H | 8.40 ± 0.12 Aabc | 70.00 ± 0.25 Bc | 220.31 ± 0.37 Bbc | 0.45 ± 0.11 Aab | 0.09 ± 0.03 Aab | 0.42 ± 0.02 Ab | 0.80 ± 0.03 Bd |
| OR 8H | 8.50 ± 0.11 Aabc | 57.31± 0.67 Ab | 210.10 ± 0.45 Aab | 0.45 ± 0.10 Aab | 0.11 ± 0.02 Acd | 0.45 ± 0.03 Ac | 0.35 ± 0.04 Ab |
| SP 8H | 9.01 ± 0.54 Abc | 80.00 ± 0.89 Bd | 240.90 ± 0.78 Bd | 0.56 ± 0.03 Ac | 0.13 ± 0.01 Aef | 0.49 ± 0.03 Bd | 0.81 ± 0.06 Bd |
| OR 12H | 8.92 ± 0.32 Abc | 60.91 ± 0.45 Ac | 226.80 ± 0.38 Ac | 0.47 ± 0.03 Ab | 0.12d ± 0.03 Ae | 0.47 ± 0.02 Ad | 0.59 ± 0.03 Ac |
| SP 12H | 9.31 ± 0.51 Bc | 90.48 ± 0.37 Be | 299.63 ± 0.59 Bf | 0.76 ± 0.06 Be | 0.17 ± 0.01 Bg | 0.62 ± 0.01 Bf | 0.81 ± 0.07 Bd |
| OR 24H | 9.15 ± 0.25 Ac | 68.32 ± 0.52 Ad | 255.95 ± 0.16 Ae | 0.64 ± 0.01 Ad | 0.13 ± 0.01 Af | 0.51 ± 0.02 Ae | 1.16 ± 0.07 Ae |
| SP 24H | 18.19 ± 0.51 Bd | 97.49 ± 0.71 Bf | 395.11 ± 0.32 Bg | 0.96 ± 0.03 Bf | 0.25 ± 0.02 Bh | 0.75 ± 0.04 Bg | 1.33 ± 0.08 Bf |
| Samples | K | Mg | Ca | Mn | Fe | Zn | Cu |
|---|---|---|---|---|---|---|---|
| OR SP 0H | 389.23 ± 1.20 Aa | 142.56 ± 0.67 Aa | 11.32 ± 0.56 Aa | 0.85 ± 0.3 Aa | 0.91 ± 0.32 Aa | 0.85 ± 0.31 Aa | 0.34 ± 0.01 Aa |
| SP PF 0H | 390.23 ± 1.01 Aa | 140.32 ± 0.99 Aa | 11.32a ± 0.34 Aa | 0.87 ± 0.01 Aa | 0.90 ± 0.02 Aa | 0.84 ± 0.03Aa | 0.35 ± 0.01 Aa |
| OR PF 12H | 400.66 ± 0.67 Aa | 158.84 ± 1.20 Ab | 12.02 ± 0.12 Aa | 0.91 ± 0.05 Aa | 0.98 ± 0.22 Aab | 0.90 ± 0.01 Aab | 0.36 ± 0.01 Aa |
| SP PF 12H | 437.70 ± 0.55 Bb | 175.70 ± 0.34 Bc | 14.66 ± 0.22 Bbc | 1.14 ± 0.04 Bb | 1.20 ± 0.21 Bb | 1.19 ± 0.03 Bc | 0.44 ± 0.01 Ab |
| OR PF 24H | 410.10 ± 0.34 Aab | 177.81 ± 0.32 Ac | 13.06a ± 0.35 Ab | 0.95 ± 0.01 Aa | 1.01 ± 0.02 Aab | 0.95 ± 0.04Aab | 0.39 ± 0.02Aab |
| SP PF 24H | 481.11 ± 0.12 Bc | 190.99 ± 0.22 Bd | 16.27 ± 1.01 Bc | 1.35 ± 0.04 Bc | 1.49 ± 0.03 Bc | 1.47 ± 0.02 Bd | 0.53 ± 0.04 Bc |
| Storage (Days) | Parameter | OR PF 0H | SP PF 0H | OR PF 12H | SP PF 12H | OR PF 24H | SP PF 24H |
|---|---|---|---|---|---|---|---|
| Day 1 | TYMC, cfu/g | 5.6 ± 0.02 Aa | 5.7 ± 0.25 Aa | 7.4 ± 0.49 Bc | 6.7 ± 0.49 Aab | 12.5 ± 0.56 Ae | 10.4 ± 0.64Ad |
| Day 3 | TYMC, cfu/g | 15.6 ± 0.04 Aa | 15.4 ± 0.89 Aa | 14.7 ± 0.78 Bc | 12.9 ± 0.55 Ab | 15.3 ± 0.55 Bbc | 12.6 ± 0.67Ab |
| Day 5 | TYMC, cfu/g | 40.3 ± 0.12 Ae | 40.5 ± 0.56 Ae | 38 ± 0.89 Bd | 28.57 ± 0.89 Aa | 35.7 ± 0.69 Bc | 30 ± 0.89 Aab |
| Day 7 | TYMC, cfu/g | 60.3 ± 1.00 Ae | 60.5 ± 0.28 Ae | 45 ± 0.95 Bc | 32 ± 0.77 Aa | 50.9 ± 0.42Bd | 40 ± 1.00 Ab |
| Day 9 | TYMC, cfu/g | 110.3 ± 0.98 Bef | 109 ± 0.45 Ae | 90.8 ± 1.5 Bc | 70.6 ± 1.2 Ab | 100.8 ± 1.4 Bd | 60.6 ± 0.99 Aa |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiş, M.S.; Păucean, A.; Man, S.M.; Bonta, V.; Pop, A.; Stan, L.; Beldean, B.V.; Pop, C.R.; Mureşan, V.; Muste, S. Effect of Rice Flour Fermentation with Lactobacillus spicheri DSM 15429 on the Nutritional Features of Gluten-Free Muffins. Foods 2020, 9, 822. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060822
Chiş MS, Păucean A, Man SM, Bonta V, Pop A, Stan L, Beldean BV, Pop CR, Mureşan V, Muste S. Effect of Rice Flour Fermentation with Lactobacillus spicheri DSM 15429 on the Nutritional Features of Gluten-Free Muffins. Foods. 2020; 9(6):822. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060822
Chicago/Turabian StyleChiş, Maria Simona, Adriana Păucean, Simona Maria Man, Victorița Bonta, Anamaria Pop, Laura Stan, Bianca Vasilica Beldean (Tătar), Carmen Rodica Pop, Vlad Mureşan, and Sevastiţa Muste. 2020. "Effect of Rice Flour Fermentation with Lactobacillus spicheri DSM 15429 on the Nutritional Features of Gluten-Free Muffins" Foods 9, no. 6: 822. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060822