Application of Plant Extracts to Control Postharvest Gray Mold and Susceptibility of Apple Fruits to B. cinerea from Different Plant Hosts



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Material and Apple Fruits
2.2. Antifungal Activity In Vitro
2.3. Antifungal Activity of the Extracts against Postharvest Gray Mold
2.4. Susceptibility of Apple Fruits to B. cinerea
2.5. Plant Extracts and Their Composition
2.6. Statistical Analysis
3. Results
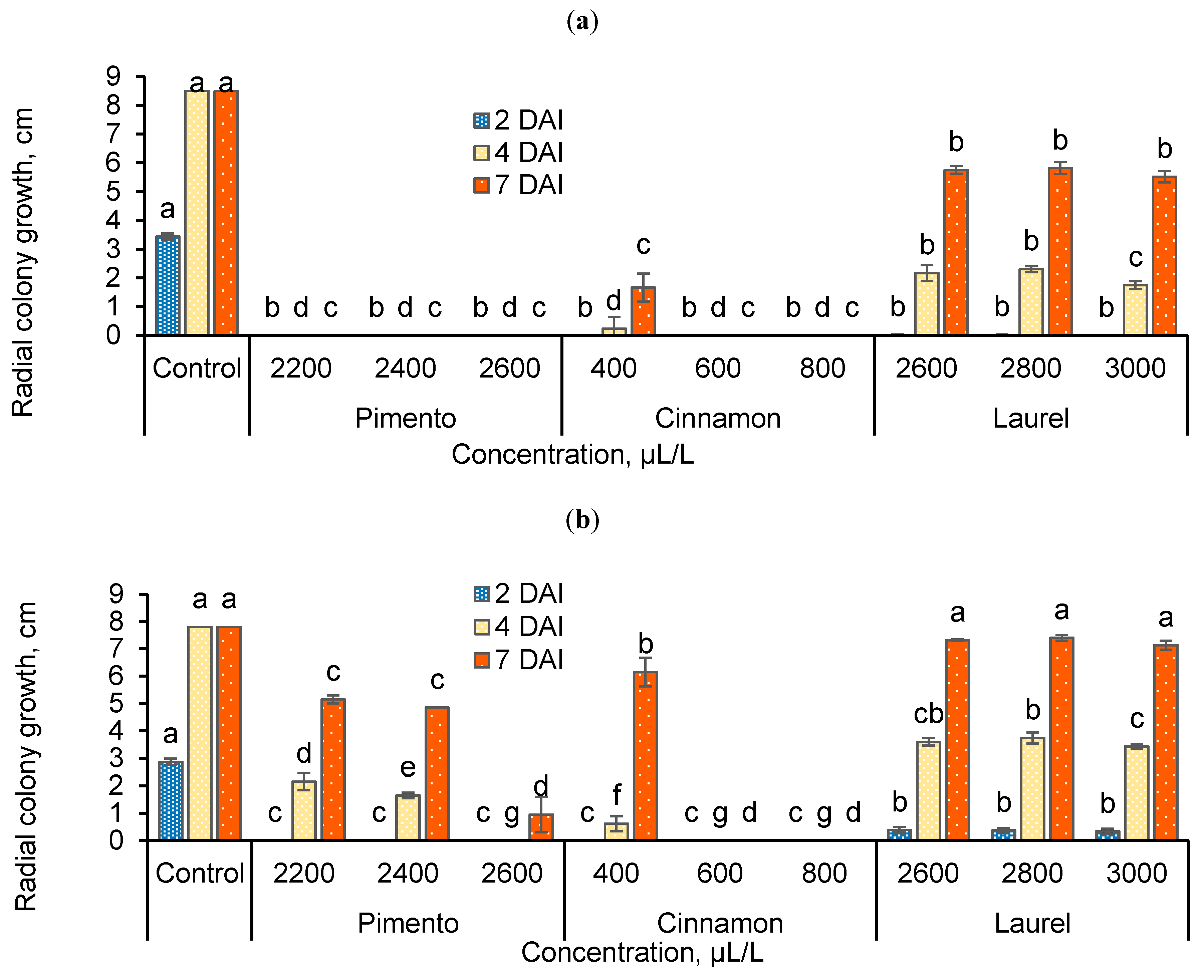
3.1. Antifungal Activity of the Extracts In Vitro
3.2. Antifungal Activity of the Extracts against Postharvest B. cinerea
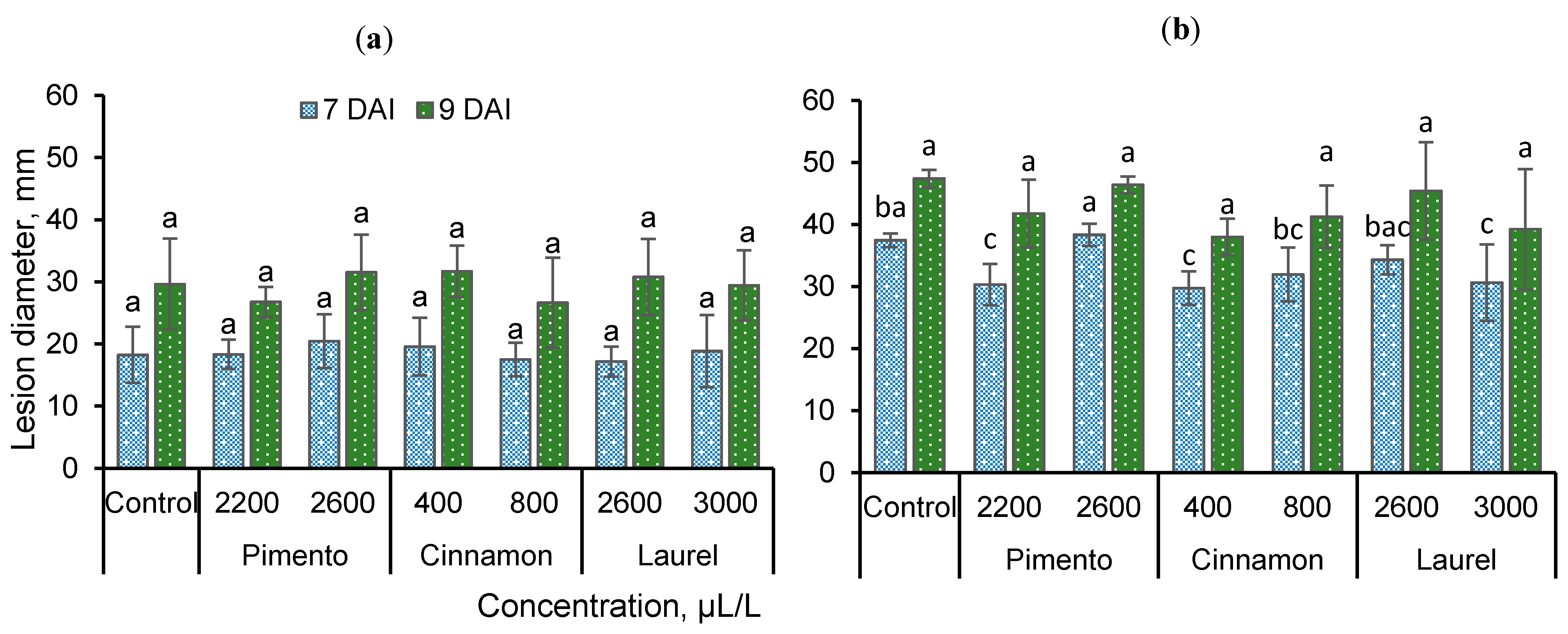
3.3. Susceptibility of Apple Fruits to B. cinerea
3.4. Composition of Plant Extracts
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Dewey, F.M.; Grant-Downton, R. Botrytis—Biology, Detection and Quantification. In Botrytis—The Fungus, the Pathogen and its Management in Agricultural Systems; Fillinger, S., Elad, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 17–34. ISBN 9783319233710. [Google Scholar] [CrossRef]
- Hua, L.; Yong, C.; Zhanquan, Z.; Boqiang, L.; Guozheng, Q.; Shiping, T. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef] [Green Version]
- Petrasch, S.; Silva, C.J.; Mesquida-Pesci, S.D.; Gallegos, K.; van den Abeele, C.; Papin, V.; Fernandez-Acero, F.J.; Knapp, S.J.; Blanco-Ulate, B. Infection strategies deployed by Botrytis cinerea, Fusarium acuminatum, and Rhizopus stolonifer as a function of tomato fruit ripening stage. Front. Plant Sci. 2019, 10, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [Green Version]
- Romanazzi, G.; Smilanick, J.L.; Feliziani, E.; Droby, S. Integrated management of postharvest gray mold on fruit crops. Postharvest Biol. Technol. 2016, 113, 69–76. [Google Scholar] [CrossRef]
- Sundström, J.F.; Albihn, A.; Boqvist, S.; Ljungvall, K.; Marstorp, H.; Martiin, C.; Nyberg, K.; Vågsholm, I.; Yuen, J.; Magnusson, U. Future threats to agricultural food production posed by environmental degradation, climate change, and animal and plant diseases—A risk analysis in three economic and climate settings. Food Secur. 2014, 6, 201–215. [Google Scholar] [CrossRef] [Green Version]
- Sehajpal, P.K.; Singh, P.J. Effect of temperature on growth, sporulation and sclerotial formation of the fungus Botrytis gladiolorum Timm. in different culture media and standardization of inoculum load of the fungus for generation of disease. Int. J. Res. 2014, 1, 772–779. [Google Scholar]
- Fininsa, H.T.C.; Tesfaye, S.S.K.; Terefe, H.; Fininsa, C.; Sahile, S.; Tesfaye, K. Effect of Temperature on Growth and Sporulation of Botrytis fabae, and Resistance Reactions of Faba Bean against the Pathogen. J. Plant Pathol. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef]
- Elad, Y.; Pertot, I.; Prado, A.M.C.; Stewart, A. Plant Hosts of Botrytis spp. In Botrytis—The Fungus, the Pathogen and Its Management in Agricultural Systems; Springer International Publishing: New York, NY, USA, 2016; pp. 413–486. ISBN 9783319233710. [Google Scholar] [CrossRef]
- Bui, T.T.A.; Al Wright, S.I.; Falk, A.B.; Vanwalleghem, T.; Van Hemelrijck, W.; Hertog, M.L.A.T.M.; Keulemans, J.; Davey, M.W. Botrytis cinerea differentially induces postharvest antioxidant responses in ‘Braeburn’ and ‘Golden Delicious’ apple fruit. J. Sci. Food Agric. 2019, 99, 5662–5670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choquer, M.; Fournier, E.; Kunz, C.; Levis, C.; Pradier, J.-M.; Simon, A.; Viaud, M. Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett. 2007, 277, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Fournier, E.; Giraud, T. Sympatric genetic differentiation of a generalist pathogenic fungus, Botrytis cinerea, on two different host plants, grapevine and bramble. J. Evol. Biol. 2008, 21, 122–132. [Google Scholar] [CrossRef]
- Cotoras, M.; Silva, E. Differences in the initial events of infection of Botrytis cinerea strains isolated from tomato and grape. Mycologia 2005, 97, 485–492. [Google Scholar] [CrossRef] [PubMed]
- Droby, S.; Lichter, A. Post-Harvest Botrytis Infection: Etiology, Development and Management. In Botrytis: Biology, Pathology and Control; Elad, Y., Williamson, B., Tudzynski, P., Delen, N., Eds.; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-2624-9. [Google Scholar] [CrossRef]
- Gholamnezhad, J. Effect of plant extracts on activity of some defense enzymes of apple fruit in interaction with Botrytis cinerea. J. Integr. Agric. 2019, 18, 115–123. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Faust, J.E.; Schnabel, G. Characterization of Botrytis cinerea from Commercial Cut Flower Roses. Plant Dis. 2019, 103, 1577–1583. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C. Phytotoxicity: An Overview of the Physiological Responses of Plants Exposed to Fungicides. J. Bot. 2012, 2012, 135479. [Google Scholar] [CrossRef] [Green Version]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea—Prospects and challenges. Biocontrol Sci. Technol. 2018, 29, 207–228. [Google Scholar] [CrossRef]
- Valiuškaitė, A.; Uselis, N.; Kviklys, D.; Lanauskas, J.; Rasiukevičiūtė, N. The effect of sustainable plant protection and apple tree management on fruit quality and yield. Zemdirbyste Agric. 2017, 104, 353–358. [Google Scholar] [CrossRef] [Green Version]
- Jurick, W.M.; Macarisin, O.; Gaskins, V.L.; Park, E.; Yu, J.; Janisiewicz, W.; Peter, K.A. Characterization of Postharvest Fungicide-Resistant Botrytis cinerea Isolates from Commercially Stored Apple Fruit. Phytopathology 2017, 107, 362–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority (EFSA). The 2017 European Union Report on Pesticide Residues in Food. EFSA J. 2019, 17, 5743. [Google Scholar] [CrossRef] [Green Version]
- Shuping, D.; Eloff, J. The use of plants to protect plants and food against fungal pathogens: A review. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Zheng, J.; Liu, T.; Guo, Z.; Zhang, L.; Mao, L.; Zhang, Y.; Jiang, H. Fumigation and contact activities of 18 plant essential oils on Villosiclava virens, the pathogenic fungus of rice false smut. Sci. Rep. 2019, 9, 7330. [Google Scholar] [CrossRef] [Green Version]
- Safaei-Ghomi, J.; Ahd, A.A. Antimicrobial and antifungal properties of the essential oil and methanol extracts of Eucalyptus largiflorens and Eucalyptus intertexta. Pharmacogn. Mag. 2010, 6, 172–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Andrade, C.M.M.; da Rosa, M.F.; Boufleuer, É.; Ferreira, F.B.P.; Iwanaga, C.C.; Gonçalves, J.E.; Cortez, D.A.G.; Martins, C.V.B.; Linde, G.A.; Simões, M.R.; et al. Chemical composition and antifungal activity of essential oil and fractions extracted from the leaves of Laurus nobilis L. cultivated in Southern Brazil. J. Med. Plants Res. 2016, 10, 865–871. [Google Scholar] [CrossRef] [Green Version]
- Morcia, C.; Malnati, M.S.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2012, 29, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Huang, Q.; Wang, Z.; Cao, H.; Zhang, D. Structure-activity relationships of cinnamaldehyde and eugenol derivatives against plant pathogenic fungi. Ind. Crop. Prod. 2017, 97, 388–394. [Google Scholar] [CrossRef]
- Olea, A.F.; Bravo, A.; Martínez, R.; Thomas, M.; Sedan, C.; Catalán, L.E.; Zambrano, E.; Carvajal, D.; Silva-Moreno, E.; Carrasco, H. Antifungal Activity of Eugenol Derivatives against Botrytis cinerea. Molecules 2019, 24, 1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Amrani, S.; Lalami, A.E.O.; Zoubi, Y.E.; Moukhafi, K.; Bouslamti, R.; Lairini, S.; Bouslmati, R. Evaluation of antibacterial and antioxidant effects of cinnamon and clove essential oils from Madagascar. Mater. Today Proc. 2019, 13, 762–770. [Google Scholar] [CrossRef]
- Xing, Y.; Li, X.; Xu, Q.; Yun, J.; Lu, Y. Antifungal activities of cinnamon oil against Rhizopus nigricans, Aspergillus flavus and Penicillium expansum in vitro and in vivo fruit test. Int. J. Food Sci. Technol. 2010, 45, 1837–1842. [Google Scholar] [CrossRef]
- Moghadam, Z.A.; Hosseini, H.; Hadian, Z.; Asgari, B.; Mirmoghtadaie, L.; Mohammadi, A.; Shamloo, E.; Javadi, N.H.S. Evaluation of the Antifungal Activity of Cinnamon, Clove, Thymes, Zataria Multiflora, Cumin and Caraway Essential Oils against Ochratoxigenic Aspergillus ochraceus. J. Pharm. Res. Int. 2019, 26, 1–16. [Google Scholar] [CrossRef]
- Somrani, M.; Inglés, M.-C.; Debbabi, H.; Abidi, F.; Palop, A. Garlic, Onion, and Cinnamon Essential Oil Anti-Biofilms’ Effect against Listeria monocytogenes. Foods 2020, 9, 567. [Google Scholar] [CrossRef]
- Mitropoulou, G.; Nikolaou, A.; Santarmaki, V.; Sgouros, G.; Kourkoutas, Y. Citrus medica and Cinnamomum zeylanicum Essential Oils as Potential Biopreservatives against Spoilage in Low Alcohol Wine Products. Foods 2020, 9, 577. [Google Scholar] [CrossRef]
- Nowotarska, S.W.; Nowotarski, K.; Grant, I.R.; Elliott, C.T.; Friedman, M.; Situ, C. Mechanisms of Antimicrobial Action of Cinnamon and Oregano Oils, Cinnamaldehyde, Carvacrol, 2,5-Dihydroxybenzaldehyde, and 2-Hydroxy-5-Methoxybenzaldehyde against Mycobacterium avium subsp. paratuberculosis (Map). Foods 2017, 6, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asha, M.M.; Chaithra, M.; Yashoda, K.; Vivek, M.N.; Prashith Kekuda, T.R. Antibacterial activity of leaf and bark extracts of Pimenta dioica (Linn.) Merill against clinical isolates of Staphylococcus aureus and Streptococcus mutants. World J. Pharm. Pharm. Sci. 2013, 2, 3207–3215. [Google Scholar]
- Šernaitė, L.; Valiuškaitė, A.; Rasiukevičiūtė, N.; Dambrauskienė, E.; Viškelis, P. Biocontrol of strawberry grey mold using pepper extracts. In Proceedings of the X International Scientific Agricultural Symposium “Agrosym 2019”, Jahorina, Bosnia and Herzegovina, 3–6 October 2019; pp. 893–898. [Google Scholar]
- Chaudhari, A.K.; Singh, V.K.; Dwivedy, A.K.; Das, S.; Upadhyay, N.; Singh, A.; Dkhar, M.S.; Kayang, H.; Prakash, B.; Dubey, N.K. Chemically characterised Pimenta dioica (L.) Merr. essential oil as a novel plant based antimicrobial against fungal and aflatoxin B1 contamination of stored maize and its possible mode of action. Nat. Prod. Res. 2018, 34, 745–749. [Google Scholar] [CrossRef] [PubMed]
- De Corato, U.; Maccioni, O.; Trupo, M.; Di Sanzo, G. Use of essential oil of Laurus nobilis obtained by means of a supercritical carbon dioxide technique against post harvest spoilage fungi. Crop. Prot. 2010, 29, 142–147. [Google Scholar] [CrossRef]
- Dammak, I.; Hamdi, Z.; El Euch, S.K.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia officinalis, Lavandula dentata and Laurus nobilis essential oils and a major monoterpene constituent 1,8-cineole against Aspergillus carbonarius. Ind. Crop. Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Özcan, M.M.; Al Juhaimi, F.Y. Antioxidant and antifungal activity of some aromatic plant extracts. J. Med. Plants Res. 2011, 5, 1361–1366. [Google Scholar]
- Šernaitė, L.; Rasiukevičiūtė, N.; Dambrauskienė, E.; Viškelis, P.; Valiuškaitė, A. Biocontrol of strawberry pathogen Botrytis cinerea using plant extracts and essential oils. Zemdirbyste Agric. 2020, 107, 147–152. [Google Scholar] [CrossRef]
- Rafiq, R.; Hayek, S.A.; Anyanwu, U.; Hardy, B.I.; Giddings, V.L.; Ibrahim, S.A.; Tahergorabi, R.; Kang, H.W. Antibacterial and Antioxidant Activities of Essential Oils from Artemisia herba-alba Asso., Pelargonium capitatum × radens and Laurus nobilis L. Foods 2016, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Dogu, D.M.; Zobar, D. Effects of Some Plant Essential Oils Against Botrytis cinerea and Tetranychus urticae on Grapevine. Turkish J. Agric. Nat. Sci. 2014, 1, 1268–1273. [Google Scholar]
- Mbili, N.C.; Opara, U.L.; Lennox, C.L.; Vries, F.A. Citrus and lemongrass essential oils inhibit Botrytis cinerea on ‘Golden Delicious’, ‘Pink Lady’ and ‘Granny Smith’ apples. J. Plant Dis. Prot. 2017, 124, 499–511. [Google Scholar] [CrossRef]
- Daniel, C.K.; Lennox, C.L.; Vries, F.A. In vivo application of garlic extracts in combination with clove oil to prevent postharvest decay caused by Botrytis cinerea, Penicillium expansum and Neofabraea alba on apples. Postharvest Biol. Technol. 2015, 99, 88–92. [Google Scholar] [CrossRef]
- Banani, H.; Olivieri, L.; Santoro, K.; Garibaldi, A.; Gullino, M.L.; Spadaro, D. Thyme and Savory Essential Oil Efficacy and Induction of Resistance against Botrytis cinerea through Priming of Defense Responses in Apple. Foods 2018, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasiukevičiūtė, N.; Rugienius, R.; Šikšnianienė, J.B. Genetic diversity of Botrytis cinerea from strawberry in Lithuania. Zemdirbyste Agric. 2018, 105, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Youssef, K.; Roberto, S.R. Premature Apple Fruit Drop: Associated Fungal Species and Attempted Management Solutions. Horticulturae 2020, 6, 31. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. The Extracts of Cinnamon and Clove as Potential Biofungicides against Strawberry Grey Mould. Plants 2020, 9, 613. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Extract | Concentration, µL/L | Inhibition of Strawberry B. cinerea, % | Inhibition of Apple B. cinerea, % |
|---|---|---|---|
| Pimento | 2200 | −0.55 | 19.09 |
| 2600 | −12.21 | −2.36 | |
| Cinnamon | 400 | −7.41 | 20.63 |
| 800 | 3.98 | 14.69 | |
| Laurel | 2600 | 5.76 | 8.41 |
| 3000 | −3.43 | 18.22 |
| Extract | Concentration, µL/L | Inoculated with Strawberry B. cinerea, % | Inoculated with Apple B. cinerea, % |
|---|---|---|---|
| Pimento | 2200 | 91.67 | 100 |
| 2600 | 100 | 83.33 | |
| Cinnamon | 400 | 100 | 100 |
| 800 | 83.33 | 100 | |
| Laurel | 2600 | 91.67 | 100 |
| 3000 | 100 | 100 |
| Pimenta dioica | Laurus nobilis | |||
|---|---|---|---|---|
| Component | PA 1 (%) | RT 2 | PA (%) | RT |
| α-Pinene | 0.69 | 6.694 | 1.04 | 6.668 |
| Sabinene | 1.45 | 7.718 | 2.10 | 7.692 |
| β-Pinene | 0.73 | 7.810 | 1.24 | 7.782 |
| Myrcene | 5.13 | 8.185 | ||
| p-Cymene | 0.28 | 9.122 | 0.56 | 9.091 |
| Limonene | 0.35 | 9.253 | 0.37 | 9.197 |
| Eucalyptol | 11.81 | 9.346 | 29.10 | 9.328 |
| cis-Sabinene hydrate | 0.20 | 10.335 | ||
| trans-β-Ocimene | 0.59 | 9.752 | ||
| Linalool | 0.83 | 11.265 | 3.40 | 11.242 |
| Delta-terpineol | 0.35 | 13.192 | ||
| Terpinen-4-ol | 0.47 | 13.505 | 2.17 | 13.473 |
| α-terpineol | 0.58 | 13.905 | 1.54 | 13.867 |
| Estragole | 0.31 | 14.064 | ||
| Linalyl acetate | 0.35 | 15.528 | ||
| Trans-cinnamaldehyde | 2.04 | 16.087 | ||
| δ-Terpinyl acetate | 1.21 | 17.236 | ||
| α-Terpinyl acetate | 0.91 | 18.139 | 18.25 | 18.158 |
| Eugenol | 10.72 | 18.546 | 3.25 | 18.394 |
| α-Ylangene | 0.24 | 18.721 | ||
| β-Elemene | 0.94 | 19.297 | 0.86 | 19.242 |
| Methyleugenol | 44.39 | 19.861 | 2.04 | 19.532 |
| trans-Caryophyllene | 6.67 | 20.114 | 1.97 | 19.983 |
| 1-Methyl-4-(1-acetoxy-1-methylethyl)-cyclol | 0.38 | 20.279 | ||
| 6,9-Guaiadiene + coumarin | 0.31 | 20.531 | ||
| Isogermacrene D | 0.22 | 20.686 | ||
| α-Humulene | 1.21 | 20.925 | 0.34 | 20.835 |
| α-Neoclovene | 0.37 | 20.891 | ||
| Germacrene D | 0.57 | 21.583 | ||
| β-selinene | 0.24 | 21.721 | 0.31 | 21.650 |
| Trans-methyl isoeugenol + viridiflorene | 0.30 | 21.921 | ||
| γ-cadinene | 0.34 | 22.300 | ||
| Eugenyl acetate | 0.47 | 22.524 | ||
| Trans-α-bisabolene | 0.74 | 22.904 | ||
| Spathulenol | 0.28 | 23.918 | ||
| Caryophyllene oxide | 0.48 | 24.108 | 0.84 | 24.042 |
| Caryophylla-4(12),8(13)-dien-5-α-ol | 0.33 | 25.586 | ||
| β-Eudesmol | 0.40 | 26.002 | ||
| Neointermedeol | 0.43 | 26.128 | ||
| Dehydrosaussurea lactone | 1.20 | 29.403 | ||
| m-Camphorene | 0.30 | 30.907 | ||
| Hexadecenoic acid | 0.47 | 31.072 | ||
| p-Camphorene | 0.42 | 31.328 | ||
| Methyl linoleate | 0.26 | 32.468 | ||
| Phytol | 0.38 | 32.674 | ||
| Linoleic acid | 3.26 | 33.025 | 1.95 | 32.985 |
| Stearic acid | 1.77 | 33.242 | ||
| Other 3 | 3.37 | 5.1 | ||
| Total identified | 99.50% | 87.99% | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. Application of Plant Extracts to Control Postharvest Gray Mold and Susceptibility of Apple Fruits to B. cinerea from Different Plant Hosts. Foods 2020, 9, 1430. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9101430
Šernaitė L, Rasiukevičiūtė N, Valiuškaitė A. Application of Plant Extracts to Control Postharvest Gray Mold and Susceptibility of Apple Fruits to B. cinerea from Different Plant Hosts. Foods. 2020; 9(10):1430. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9101430
Chicago/Turabian StyleŠernaitė, Lina, Neringa Rasiukevičiūtė, and Alma Valiuškaitė. 2020. "Application of Plant Extracts to Control Postharvest Gray Mold and Susceptibility of Apple Fruits to B. cinerea from Different Plant Hosts" Foods 9, no. 10: 1430. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9101430