Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao

Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of NAC Family Genes in Theobroma cacao
2.2. Sequence Analysis and Structural Characterization
2.3. Phylogenetic Analysis and Classification of the TcNAC Gene Family
2.4. Analysis of Cis-Acting Elements in the TcNAC Gene Promoter
2.5. Chromosomal Location and Evolutionary Analysis of TcNAC Genes
3. Results
3.1. Identification of TcNAC Genes in Theobroma cacao
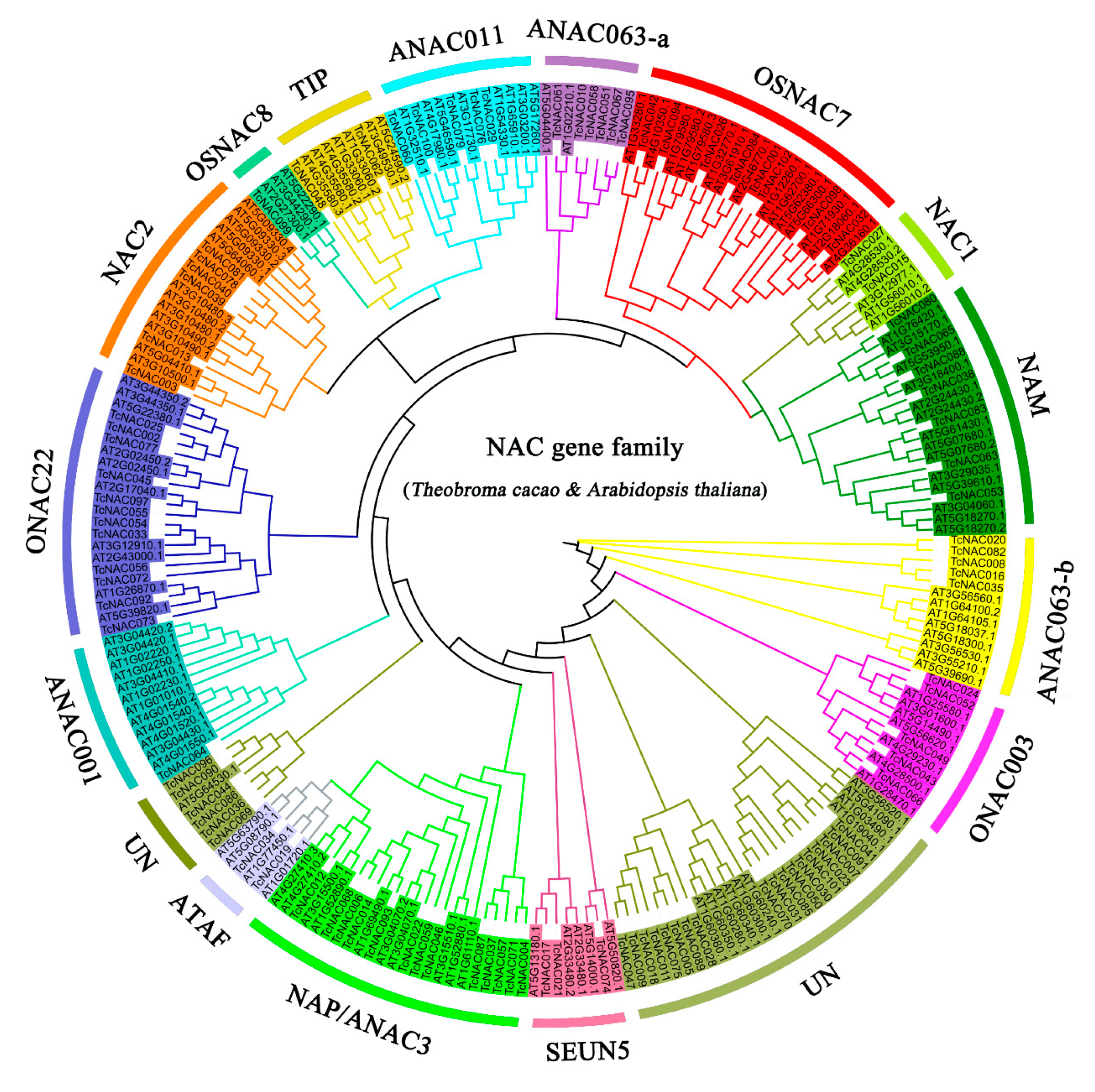
3.2. Phylogenetic Analysis and Classification of the NAC Gene Family in Theobroma cacao and Arabidopsis
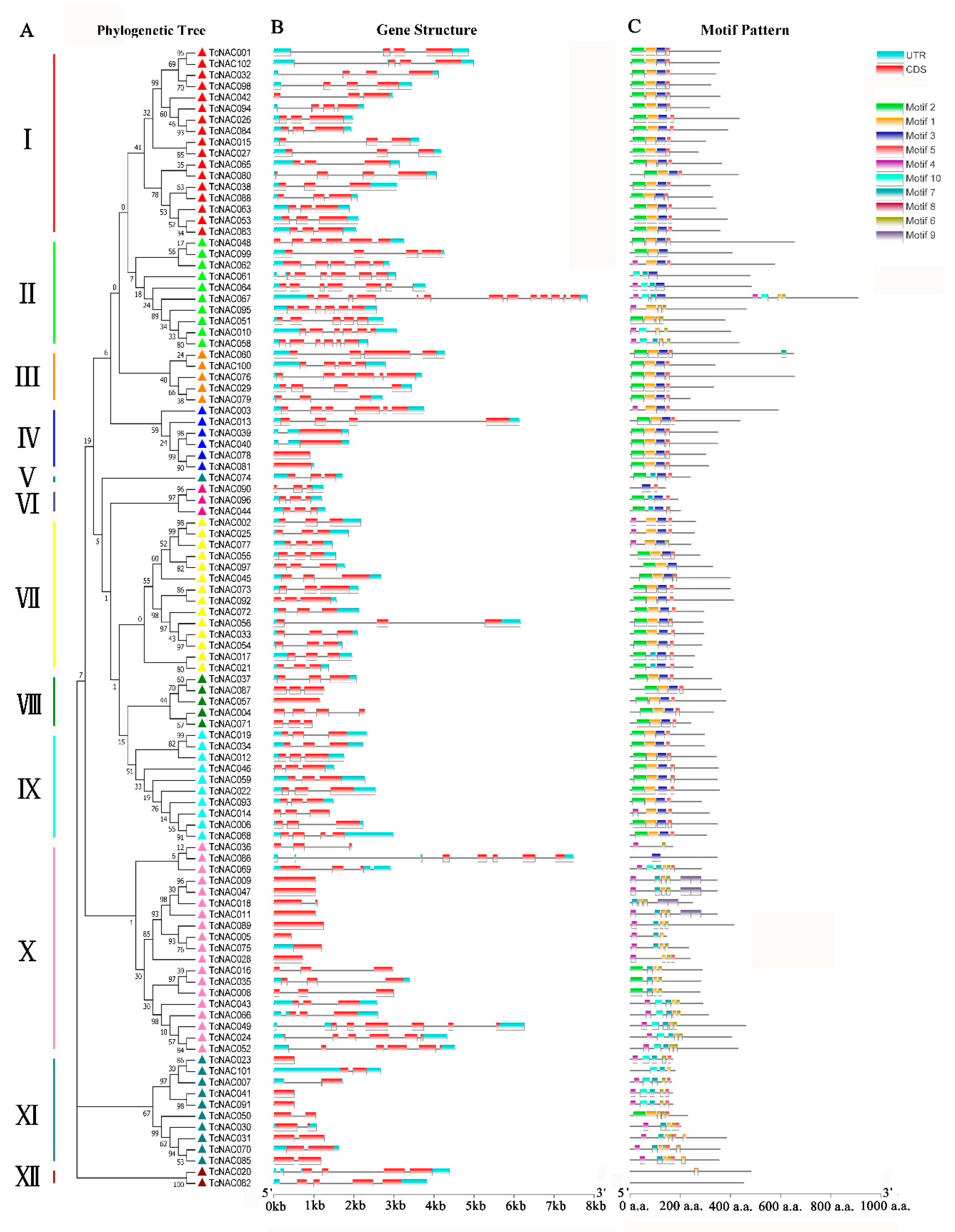
3.3. TcNAC Gene Structure and Conserved Motifs
3.4. Responsive Elements in TcNAC Promoters
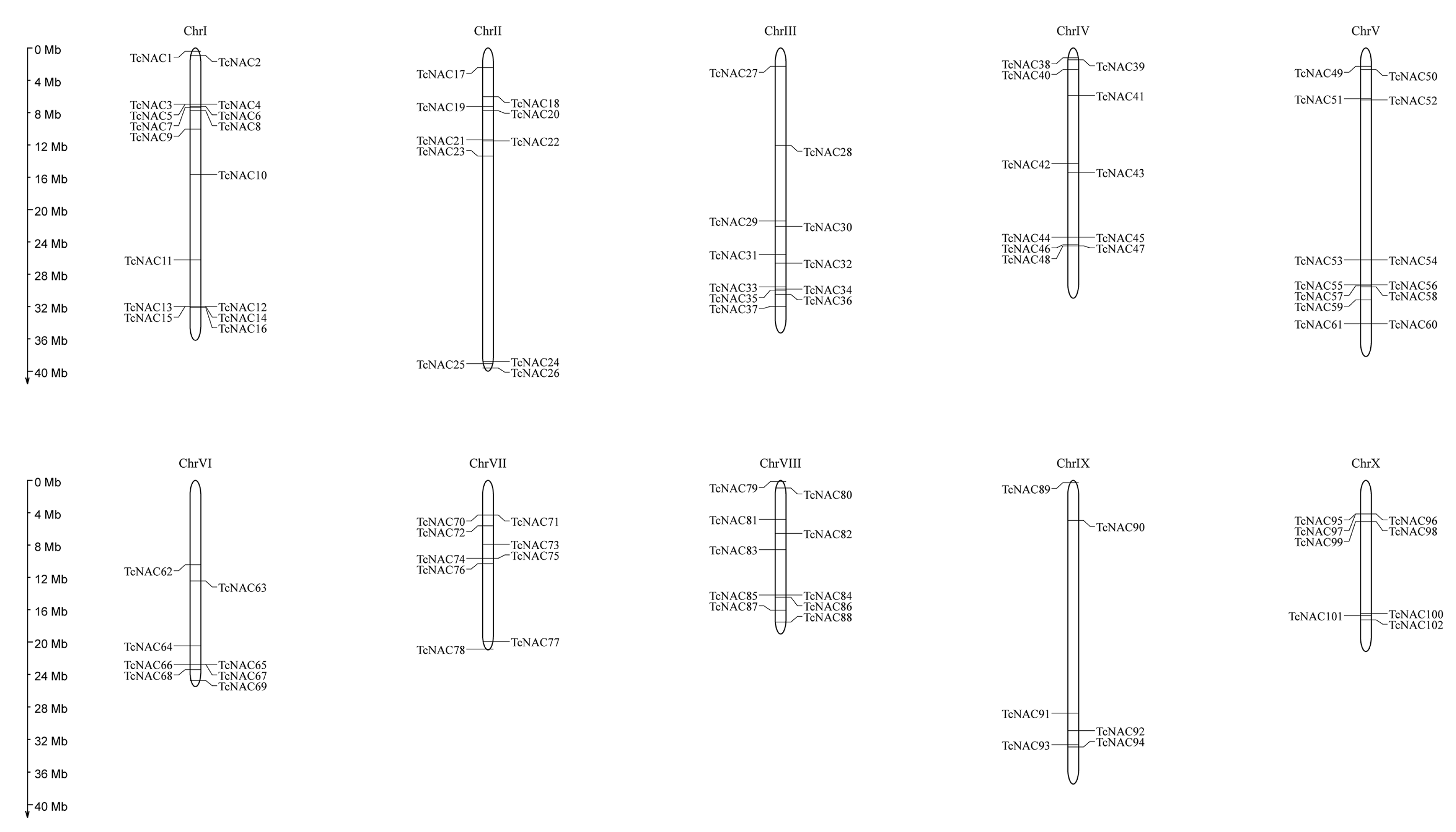
3.5. Chromosomal Location of Theobroma cacao NAC genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Latchman, D.S. Transcription factors: An overview. Int. J. Exp. Pathol. 1993, 74, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Birkenbihl, R.P.; Jach, G.; Saedler, H.; Huijser, P. Functional dissection of the plant-specific SBP-domain: overlap of the DNA-binding and nuclear localization domains. J. Mol. Biol. 2005, 352, 585–596. [Google Scholar] [CrossRef] [Green Version]
- Riaño-Pachón, D.M.; Ruzicic, S.; Dreyer, I.; Mueller-Roeber, B. PlnTFDB: An integrative plant transcription factor database. BMC Bioinform. 2007, 8, 42. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Jin, P.J.; Liang, T.; Yi, Z.; Xiao, C.G.; Ge, G.; Jing, C.L. PlantTFDB 2.0: update and improvement of the comprehensive plant transcription factor database. Nucleic Acids Res. 2011, 39, D1114–D1117. [Google Scholar]
- Souer, E.; Houwelingen, A.V.; Kloos, D.; Mol, J.; Koes, R. The no apical meristem gene of Petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [Green Version]
- Ernst, H.A.; Olsen, A.N.; Larsen, S.; Lo, L.L. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.K.; Rung, J.H.; Gregersen, P.L.; Gjetting, T.; Fuglsang, A.T.; Hansen, M.; Joehnk, N.; Lyngkjaer, M.F.; Collinge, D.B. The HvNAC6 transcription factor: a positive regulator of penetration resistance in barley and Arabidopsis. Plant Mol. Biol. 2007, 65, 137. [Google Scholar] [CrossRef]
- Duval, M.; Hsieh, T.F.; Kim, S.Y.; Thomas, T.L. Molecular characterization of AtNAM: A member of the Arabidopsis NAC domain superfamily. Plant Mol. Biol. 2002, 50, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Kusano, H.; Asano, T.; Shimada, H.; Kadowaki, K.I. Molecular characterization of ONAC300, a novel NAC gene specifically expressed at early stages in various developing tissues of rice. Mol. Genet. Genomics 2005, 272, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Jae-Heung, K.; Seung Hwan, Y.; Park, A.H.; Olivier, L.; Kyung-Hwan, H. ANAC012, a member of the plant-specific NAC transcription factor family, negatively regulates xylary fiber development in Arabidopsis thaliana. Plant. J. 2010, 50, 1035–1048. [Google Scholar]
- Cohen, L.S.; Gosenfeld, L.; Wilkins, J.; Kammerer, R.C.; Tachiki, K. Demonstration of an amino acid metabolite of phencyclidine. N. Engl. J. Med. 1982, 306, 1427–1428. [Google Scholar] [PubMed]
- Cristobal, U.; Assaf, D.; Tzion, F.; Ann, B.; Jorge, D. A NAC Gene regulating senescence improves grain protein, zinc, and iron content in wheat. Science 2006, 314, 1298–1301. [Google Scholar]
- Youn-Sung, K.; Sang-Gyu, K.; Jung-Eun, P.; Hye-Young, P.; Mi-Hye, L.; Nam-Hai, C.; Chung-Mo, P. A membrane-bound NAC transcription factor regulates cell division in Arabidopsis. Plant Cell 2006, 18, 3132–3144. [Google Scholar]
- Liu, Y.Z.; Baig, M.N.R.; Fan, R.; Ye, J.L.; Cao, Y.C.; Deng, X.X. Identification and expression pattern of a novel NAM, ATAF, and CUC-like gene from Citrus sinensis osbeck. Plant Mol. Biol. Rep. 2009, 27, 292. [Google Scholar] [CrossRef]
- Willemsen, V.; Bauch, M.; Bennett, T.; Campilho, A.; Wolkenfelt, H.; Jian, X.; Haseloff, J.; Scheres, B. The NAC domain transcription factors FEZ and SOMBRERO control the orientation of cell division plane in Arabidopsis root Stem Cells. Dev. Cell 2009, 15, 913–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiroaki, K.; Taizo, M.; Yoshibumi, K.; Tamao, S.; Atsushi, K. Overexpression of the NAC transcription factor family gene ANAC036 results in a dwarf phenotype in Arabidopsis thaliana. J. Plant Physiol. 2010, 167, 571–577. [Google Scholar]
- Yong, F.G.; Su, S.G. AtNAP, a NAC family transcription factor, has an important role in leaf senescence. Plant. J. 2010, 46, 601–612. [Google Scholar]
- Jung, K.M.; Mi-Jeong, P.; Pil Joon, S.; Jin-Su, S.; Hie-Joon, K.; Chung-Mo, P. Controlled nuclear import of the transcription factor NTL6 reveals a cytoplasmic role of SnRK2.8 in the drought-stress response. Biochem. J. 2012, 448, 353–363. [Google Scholar]
- Lee, S.; Seo, P.J.; Lee, H.J.; Park, C.M. A NAC transcription factor NTL4 promotes reactive oxygen species production during drought-induced leaf senescence in Arabidopsis. Plant. J. 2012, 70, 831–844. [Google Scholar] [CrossRef]
- Negi, S.; Tak, H.; Ganapathi, T.R. Expression analysis of MusaNAC68 transcription factor and its functional analysis by overexpression in transgenic banana plants. Plant Cell Tissue Organ Cult. 2016, 125, 59–70. [Google Scholar] [CrossRef]
- Nobutaka, M.; Akira, I.; Hiroyuki, Y.; Masato, Y.; Motoaki, S.; Kazuo, S.; Masaru, O.T. NAC transcription factors, NST1 and NST3, are key regulators of the formation of secondary walls in woody tissues of Arabidopsis. Plant Cell 2007, 19, 270–280. [Google Scholar]
- Mangrauthia, S.K.; Agarwal, S.; Sailaja, B.; Sarla, N.; Voleti, S.R. Transcriptome analysis of Oryza sativa (Rice) seed germination at high temperature shows dynamics of genome expression associated with hormones signalling and abiotic stress pathways. Trop. Plant Biol. 2016, 9, 215–228. [Google Scholar] [CrossRef]
- Jensen, M.K.; Grant, M.R.; Rung, J.H.; Collinge, D.B.; Lyngkjaer, M.F. Transcriptional regulation by an NAC (NAM-ATAF1,2-CUC2) transcription factor attenuates ABA signalling for efficient basal defence towards Blumeria graminis f. sp hordei in Arabidopsis. Plant. J. 2010, 56, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Lv, B.; Luo, L.; He, J.; Ming, F. Corrigendum: The NAC-type transcription factor OsNAC2 regulates ABA-dependent genes and abiotic stress tolerance in rice. Sci. Rep. 2017, 7, 46890. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M.; Satoh, K.; Kondoh, H.; Ooka, H.; Kikuchi, S. Genome-wide analysis of NAC transcription factor family in rice. Gene 2010, 465, 30–44. [Google Scholar] [CrossRef]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [Green Version]
- Zhuo, X.; Zheng, T.; Zhang, Z.; Zhang, Y.; Jiang, L.; Ahmad, S.; Sun, L.; Wang, J.; Cheng, T.; Zhang, Q. Genome-wide analysis of the NAC transcription factor gene family reveals differential expression patterns and cold-stress responses in the woody plant Prunus mume. Genes 2018, 9, 494. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.D.; Yuan, J.; Yan, A.Y.; Zi, W.X.; Xue, H.B.; Jing, G.; Shi, R.T.; Yoon, K.H.; Shu, M.H.; Fei, F.H. Identification and expression analysis of the NAC gene family in Coffea canephora. Agronomy 2019, 9, 670. [Google Scholar]
- Liu, M.; Ma, Z.; Sun, W. Genome-wide analysis of the NAC transcription factor family in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2019, 20, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, K.A.; Donovan, J.L.; Waterhouse, A.L.; Gary, W. Cocoa and health: a decade of research. Br. J. Nutr. 2008, 99, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corti, R.; Flammer, A.J.; Hollenberg, N.K.; Lüscher, T.F. Cocoa and cardiovascular health. Circulation 2009, 119, 1433–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, Q.R.D.; Gattward, J.N.; Almoosawi, S.; Dantas, P.A.D.S.; Júnior, Q.R.D.A. Cocoa and human health: from head to foot—A review. Crit. Rev. Food. Sci. 2016, 56, 1–12. [Google Scholar] [CrossRef]
- Xavier, A.; Jerome, S.; Jean-Marc, A.; Guiltinan, M.J.; Gaetan, D.; Jerome, G.; Mathilde, A.; Cristian, C.; Thierry, L.; Maximova, S.N. The genome of Theobroma cacao. Nat. Genet. 2011, 43, 101–108. [Google Scholar]
- Li, S.; Gao, J.; Yao, L.; Ren, G.; Zhu, X.; Gao, S.; Qiu, K.; Zhou, X.; Kuai, B. The role of ANAC072 in the regulation of chlorophyll degradation during age- and dark-induced leaf senescence. Plant Cell Rep. 2016, 35, 1729–1741. [Google Scholar] [CrossRef]
- Liu, H.; Wu, M.; Zhu, D.; Pan, F.; Wang, Y.; Wang, Y.; Xiang, Y. Genome-Wide analysis of the AAAP gene family in moso bamboo (Phyllostachys edulis). BMC Plant Biol. 2017, 17, 29. [Google Scholar] [CrossRef] [Green Version]
- Thirumalaikumar, V.P.; Devkar, V.; Mehterov, N.; Ali, S.; Ozgur, R.; Turkan, I.; Mueller-Roeber, B.; Balazadeh, S. NAC transcription factor JUNGBRUNNEN1 enhances drought tolerance in tomato. Plant Biotechnol. J. 2017, 16, 354–366. [Google Scholar] [CrossRef]
- Wu, H.; Fu, B.; Sun, P.; Xiao, C.; Liu, J.H. A NAC transcription factor represses putrescine biosynthesis and affects drought tolerance. Plant Physiol. 2016, 172, 1532–1547. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J. Pfam: the protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Wang, B.; Li, Z.; Peng, Z.; Zhang, J. TsNAC1 is a key transcription factor in abiotic stress resistance and growth. Plant Physiol. 2017, 176, 742–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, H.; Zhu, D.; Gao, Y.; Yan, H.; Xiang, Y. Genome-wide analysis of VQ motif-containing proteins in Moso bamboo (Phyllostachys edulis). Planta 2017, 246, 165. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 1, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Felsenstein, J. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Cavalcanti, A.; Chen, F.C.; Bouman, P.; Li, W.H. Extent of gene duplication in the genomes of Drosophila, nematode, and yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Zhang, X.; Yue, J.X.; Tian, D.; Chen, J.Q. Recent duplications dominate NBS-encoding gene expansion in two woody species. Mol. Genet. Genomics. 2008, 280, 187–198. [Google Scholar] [CrossRef]
- Aida, M.; Ishida, T.; Fukaki, H.; Fujisawa, H.; Tasaka, M. Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell 1997, 9, 841–857. [Google Scholar] [CrossRef] [Green Version]
- Collinge, M.; Boller, T. Differential induction of two potato genes, Stprx2 and StNAC, in response to infection by Phytophthora infestans and to wounding. Plant Mol. Biol. 2001, 46, 521–529. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Tandem Repeat Sequence | Ka | Ks | Ka/Ks |
|---|---|---|---|
| TcNAC048/TcNAC047 | 0.069652 | 0.204593 | 0.34044 |
| TcNAC055/TcNAC057 | 0.022392 | 0.046771 | 0.47876 |
| TcNAC055/TcNAC056 | 0.031369 | 0.066627 | 0.47082 |
| TcNAC056/TcNAC057 | 0.020842 | 0.045468 | 0.45839 |
| TcNAC047/TcNAC046 | 0.071078 | 0.056378 | 1.26074 |
| TcNAC085/TcNAC084 | 0.057197 | 0.128216 | 0.44609 |
| TcNAC085/TcNAC086 | 0.051726 | 0.110318 | 0.46888 |
| TcNAC003/TcNAC004 | 0.028539 | 0.129133 | 0.22101 |
| TcNAC100/TcNAC101 | 0.014445 | 0.046379 | 0.33115 |
| TcNAC084/TcNAC086 | 0.053422 | 0.090511 | 0.59023 |
| TcNAC063/TcNAC005 | 0.111896 | 0.215804 | 0.51851 |
| TcNAC048/TcNAC046 | 0.077084 | 0.210968 | 0.36538 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, S.; Zhang, Q.; Shi, Y.; Sun, Z.; Zhang, Q.; Hou, S.; Wu, R.; Jiang, L.; Zhao, X.; Guo, Y. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes 2020, 11, 35. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010035
Shen S, Zhang Q, Shi Y, Sun Z, Zhang Q, Hou S, Wu R, Jiang L, Zhao X, Guo Y. Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao. Genes. 2020; 11(1):35. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010035
Chicago/Turabian StyleShen, Shiya, Qianru Zhang, Yu Shi, Zhenmei Sun, Qianqian Zhang, Sijia Hou, Rongling Wu, Libo Jiang, Xiyang Zhao, and Yunqian Guo. 2020. "Genome-Wide Analysis of the NAC Domain Transcription Factor Gene Family in Theobroma cacao" Genes 11, no. 1: 35. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010035