Shedding Light on the Antimicrobial Peptide Arsenal of Terrestrial Isopods: Focus on Armadillidins, a New Crustacean AMP Family

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Transcriptome Datasets and Preparation of Query Sets
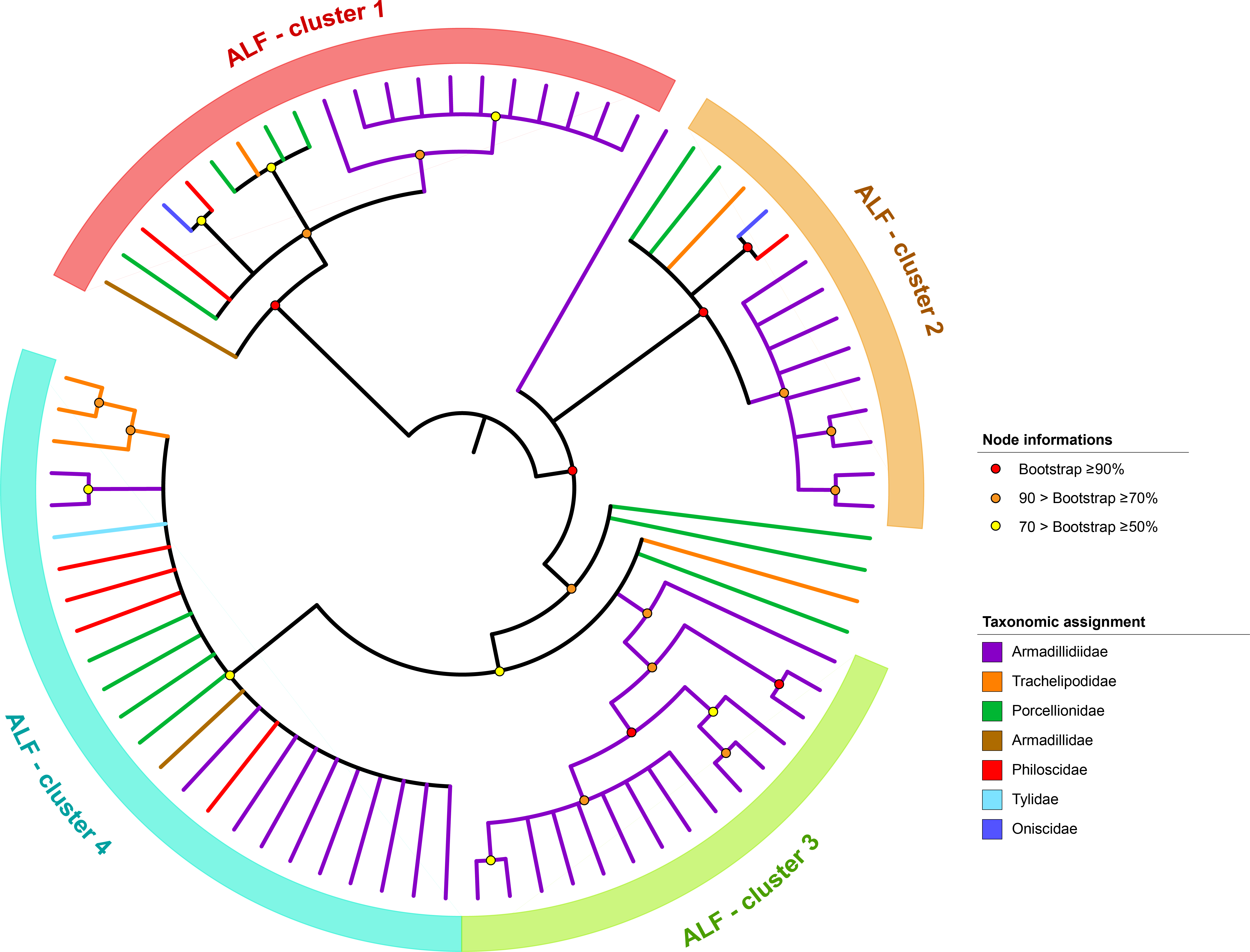
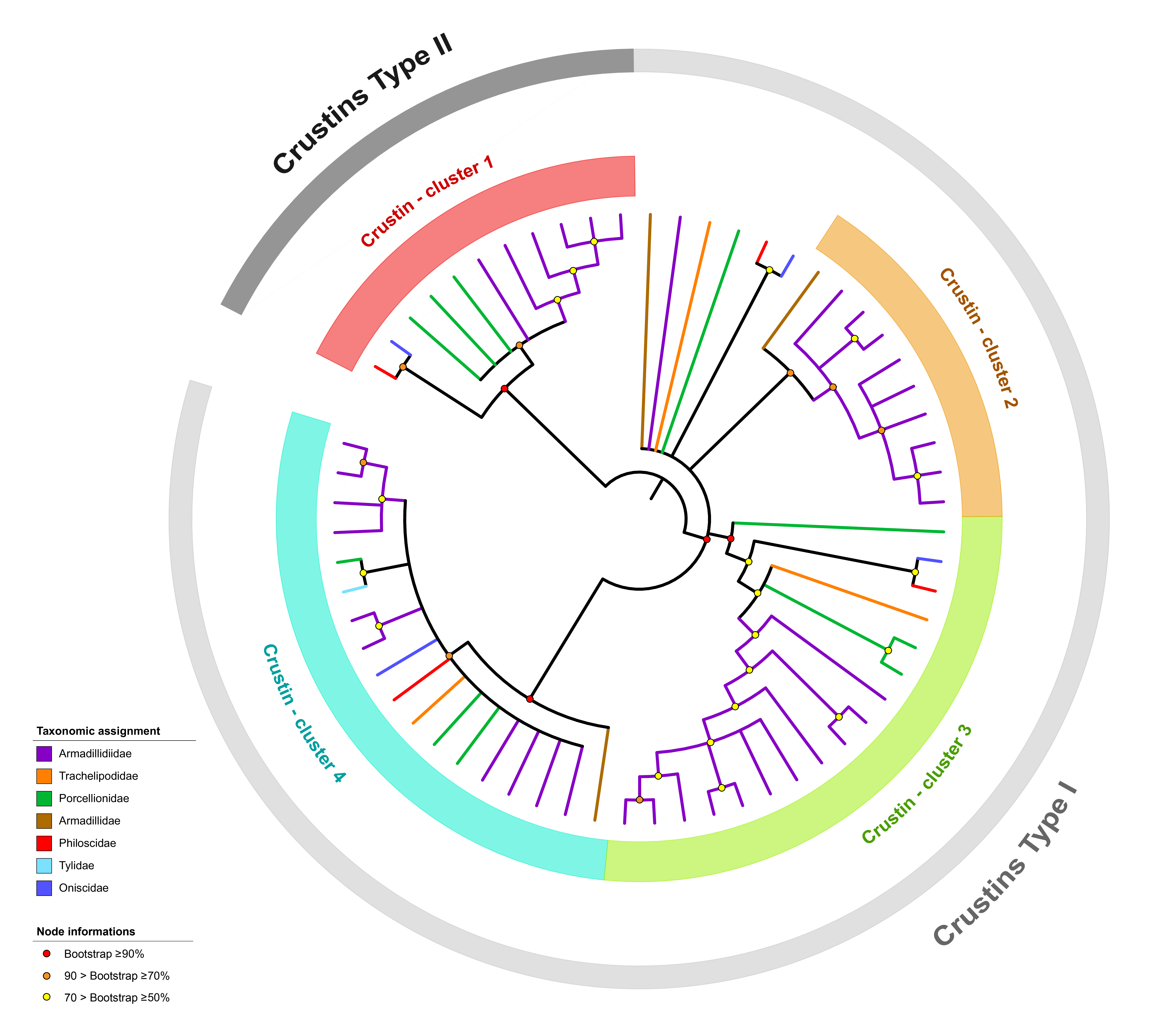
2.2. In Silico Characterization and Phylogenetic Analyses of Crustins and ALFs
2.3. Identification and Characterization of Armadillidins
2.4. Bacterial Growth Inhibition Assays
3. Results and Discussion
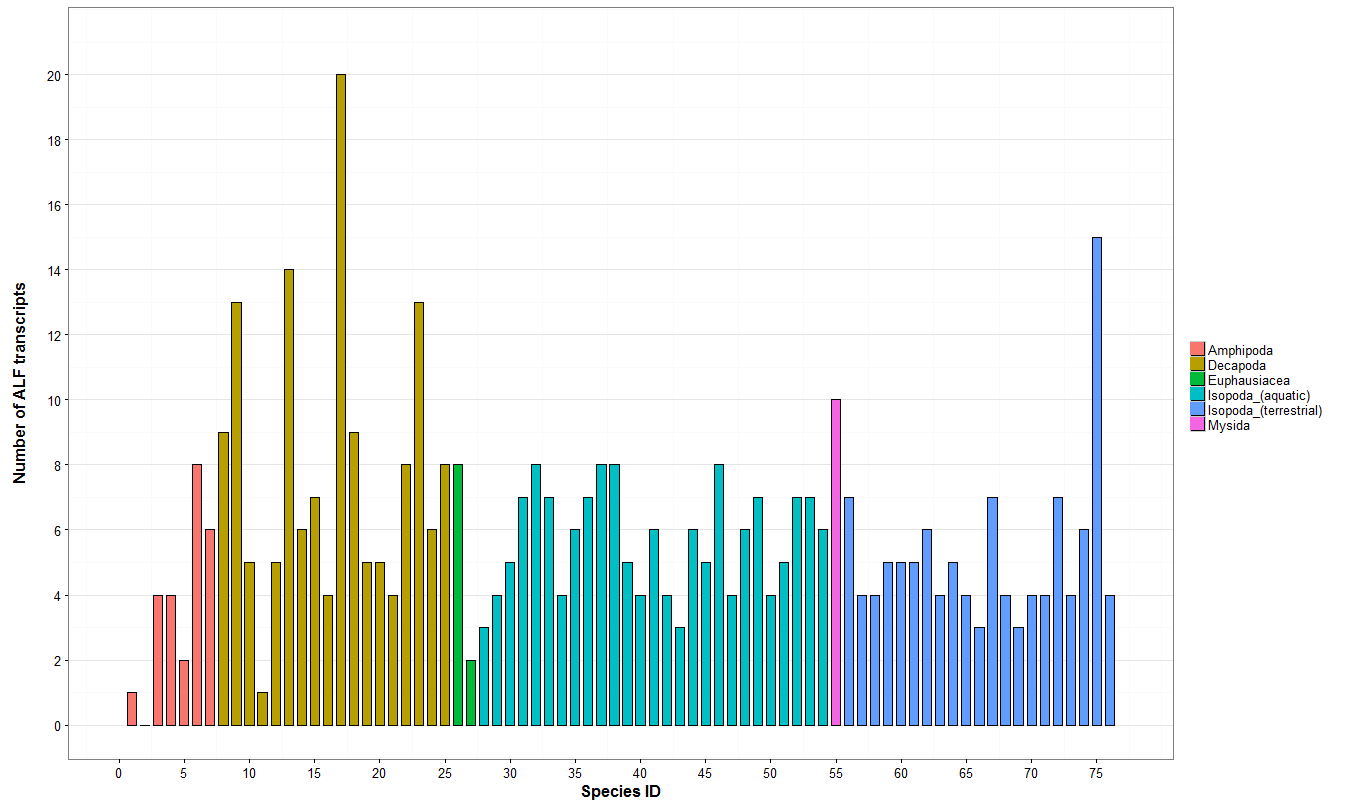
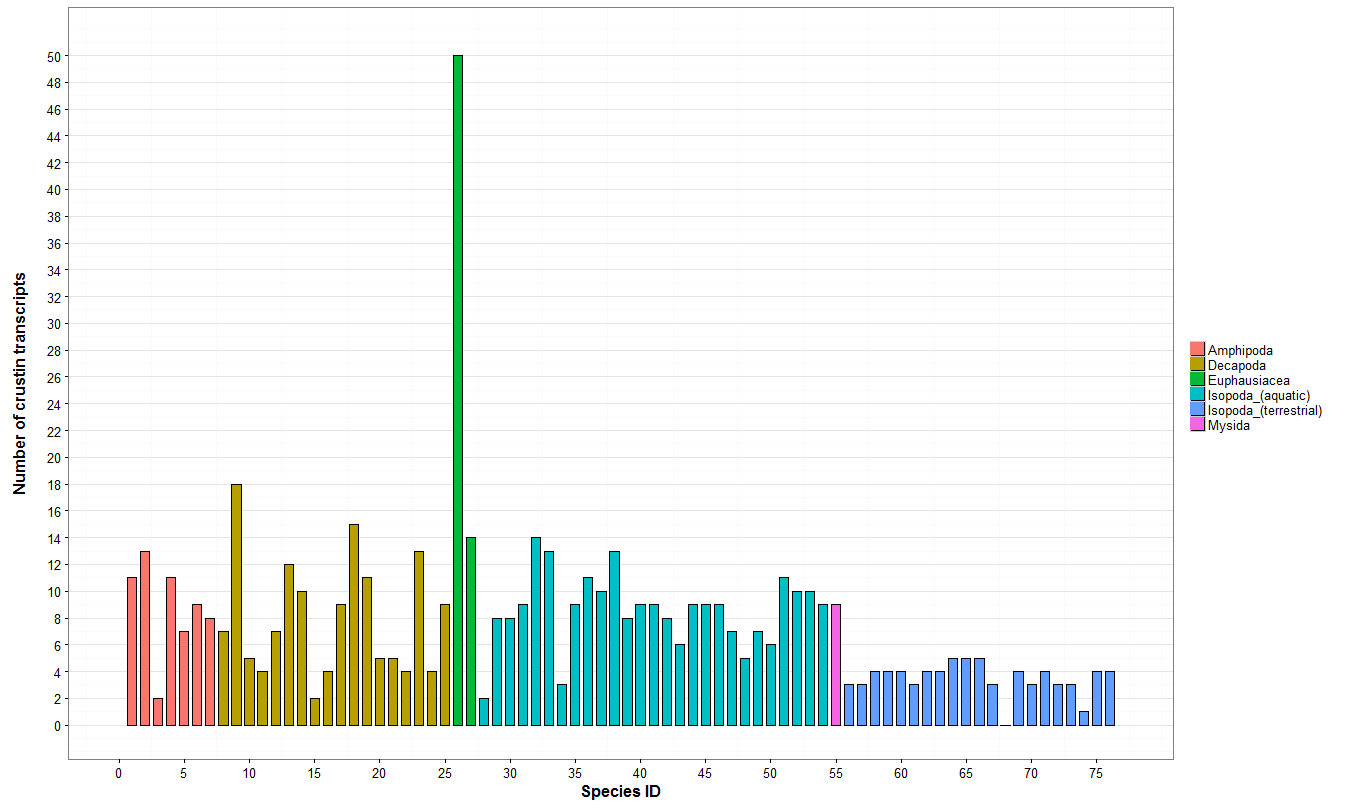
3.1. Description of the ALF and Crustin Repertoire of Terrestrial Isopods
3.2. Armadillidins, a New and Unique Crustacean AMP Family from Terrestrial Isopods
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Giribet, G.; Edgecombe, G.D. Reevaluating the Arthropod Tree of Life. Annu. Rev. Entomol. 2012, 57, 167–186. [Google Scholar] [CrossRef] [Green Version]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invertebr. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Cerenius, L.; Kawabata, S.; Lee, B.L.; Nonaka, M.; Söderhäll, K. Proteolytic cascades and their involvement in invertebrate immunity. Trends Biochem. Sci. 2010, 35, 575–583. [Google Scholar] [CrossRef]
- Balandin, S.V.; Ovchinnikova, T.V. Antimicrobial peptides of invertebrates. Part 1. Structure, biosynthesis, and evolution. Russ. J. Bioorg. Chem. 2016, 42, 229–248. [Google Scholar] [CrossRef]
- Ageitos, J.M.; Sánchez-Pérez, A.; Calo-Mata, P.; Villa, T.G. Antimicrobial peptides (AMPs): Ancient compounds that represent novel weapons in the fight against bacteria. Biochem. Pharmacol. 2017, 133, 117–138. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Dyrynda, E.A. Antimicrobial proteins: From old proteins, new tricks. Mol. Immunol. 2015, 68, 383–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornung, E. Evolutionary adaptation of oniscidean isopods to terrestrial life: Structure, physiology and behavior. Terr. Arthropod Rev. 2011, 4, 95–130. [Google Scholar] [CrossRef]
- Jeffery, S.; Gardi, C.; Jones, A. Institute for Environment and Sustainability Atlas Européen de la Biodiversité des Sols; European Commission Joint Research Centre: Luxembourg, 2013; ISBN 978-92-79-29726-7. [Google Scholar]
- Bouchon, D.; Zimmer, M.; Dittmer, J. The Terrestrial Isopod Microbiome: An All-in-One Toolbox for Animal–Microbe Interactions of Ecological Relevance. Front. Microbiol. 2016, 7, 1472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbinière, J.; Braquart-Varnier, C.; Grève, P.; Strub, J.-M.; Frère, J.; Van Dorsselaer, A.; Martin, G. Armadillidin: A novel glycine-rich antibacterial peptide directed against gram-positive bacteria in the woodlouse Armadillidium vulgare (Terrestrial Isopod, Crustacean). Dev. Comp. Immunol. 2005, 29, 489–499. [Google Scholar] [CrossRef]
- Verdon, J.; Coutos-Thevenot, P.; Rodier, M.-H.; Landon, C.; Depayras, S.; Noel, C.; La Camera, S.; Moumen, B.; Greve, P.; Bouchon, D.; et al. Armadillidin H, a Glycine-Rich Peptide from the Terrestrial Crustacean Armadillidium vulgare, Displays an Unexpected Wide Antimicrobial Spectrum with Membranolytic Activity. Front. Microbiol. 2016, 7, 1484. [Google Scholar] [CrossRef] [Green Version]
- Becking, T.; Giraud, I.; Raimond, M.; Moumen, B.; Chandler, C.; Cordaux, R.; Gilbert, C. Diversity and evolution of sex determination systems in terrestrial isopods. Sci. Rep. 2017, 7, 1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific White Shrimp, Litopenaeus vannamei, and the Atlantic White Shrimp, L. setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Sruthy, K.S.; Chaithanya, E.R.; Sathyan, N.; Nair, A.; Antony, S.P.; Bright Singh, I.S.; Philip, R. Molecular Characterization and Phylogenetic Analysis of Novel Isoform of Anti-lipopolysaccharide Factor from the Mantis Shrimp, Miyakea nepa. Probiot. Antimicrob. Proteins 2015, 7, 275–283. [Google Scholar] [CrossRef]
- Gu, H.; Sun, Q.; Jiang, S.; Zhang, J.; Sun, L. First characterization of an anti-lipopolysaccharide factor (ALF) from hydrothermal vent shrimp: Insights into the immune function of deep-sea crustacean ALF. Dev. Comp. Immunol. 2018, 84, 382–395. [Google Scholar] [CrossRef]
- Relf, J.M.; Chisholm, J.R.S.; Kemp, G.D.; Smith, V.J. Purification and characterization of a cysteine-rich 11.5-kDa antibacterial protein from the granular haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1999, 264, 350–357. [Google Scholar] [CrossRef] [Green Version]
- Bartlett, T.C.; Cuthbertson, B.J.; Shepard, E.F.; Chapman, R.W.; Gross, P.S.; Warr, G.W. Crustins, Homologues of an 11.5-kDa Antibacterial Peptide, from Two Species of Penaeid Shrimp, Litopenaeus vannamei and Litopenaeus setiferus. Mar. Biotechnol. 2002, 4, 278–293. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Talavera, G.; Castresana, J. Improvement of phylogenies after removing divergent and ambiguously aligned blocks from protein sequence alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics (Oxf. Engl.) 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics (Oxf. Engl.) 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive tree of life (iTOL) v3: An online tool for the display and annotation of phylogenetic and other trees. Nucleic Acids Res. 2016, 44, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of combined transmembrane topology and signal peptide prediction—The Phobius web server. Nucleic Acids Res. 2007, 35, W429–W432. [Google Scholar] [CrossRef] [Green Version]
- Lai, A.G.; Aboobaker, A.A. Comparative genomic analysis of innate immunity reveals novel and conserved components in crustacean food crop species. BMC Genom. 2017, 18, 389. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Albores, F.; Martínez-Porchas, M. Crustins are distinctive members of the WAP-containing protein superfamily: An improved classification approach. Dev. Comp. Immunol. 2017, 76, 9–17. [Google Scholar] [CrossRef]
- Bals, R.; Wilson, J.M. Cathelicidins—A family of multifunctional antimicrobial peptides. Cell. Mol. Life Sci. CMLS 2003, 60, 711–720. [Google Scholar] [CrossRef]
- Vanhoye, D.; Bruston, F.; Nicolas, P.; Amiche, M. Antimicrobial peptides from hylid and ranin frogs originated from a 150-million-year-old ancestral precursor with a conserved signal peptide but a hypermutable antimicrobial domain. Eur. J. Biochem. 2003, 270, 2068–2081. [Google Scholar] [CrossRef]
- Lorenzini, D.M.; Fukuzawa, A.H.; da Silva, P.I.; Machado-Santelli, G.; Bijovsky, A.T.; Daffre, S. Molecular cloning, expression analysis and cellular localization of gomesin, an anti-microbial peptide from hemocytes of the spider Acanthoscurria gomesiana. Insect Biochem. Mol. Biol. 2003, 33, 1011–1016. [Google Scholar] [CrossRef]
- Baumann, T.; Kämpfer, U.; Schürch, S.; Schaller, J.; Largiadèr, C.; Nentwig, W.; Kuhn-Nentwig, L. Ctenidins: Antimicrobial glycine-rich peptides from the hemocytes of the spider Cupiennius salei. Cell. Mol. Life Sci. 2010, 67, 2787–2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.C.; VijayRaghavan, K.; Celniker, S.E.; Tanouye, M.A. Identification of a Drosophila muscle development gene with structural homology to mammalian early growth response transcription factors. Proc. Natl. Acad. Sci. USA 1995, 92, 10344–10348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, J.C.; Berto, R.F.; Gois, E.A.; Fontenele-Cardi, N.C.; Honório-Júnior, J.E.R.; Konno, K.; Richardson, M.; Rocha, M.F.G.; Camargo, A.A.C.M.; Pimenta, D.C.; et al. Leptoglycin: A new Glycine/Leucine-rich antimicrobial peptide isolated from the skin secretion of the South American frog Leptodactylus pentadactylus (Leptodactylidae). Toxicon 2009, 54, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Chung, T.J.; Park, C.W.; Hahn, Y.; Chung, J.H.; Lee, B.L.; Han, D.M.; Jung, Y.H.; Kim, S.; Lee, Y. Structure and Expression of the Tenecin 3 Gene inTenebrio molitor. Biochem. Biophys. Res. Commun. 1996, 218, 6–11. [Google Scholar] [CrossRef]
- Park, C.J.; Park, C.B.; Hong, S.-S.; Lee, H.-S.; Lee, S.Y.; Kim, S.C. Characterization and cDNA cloning of two glycine- and histidine-rich antimicrobial peptides from the roots of shepherd’s purse, Capsella bursa-pastoris. Plant Mol. Biol. 2000, 44, 187–197. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
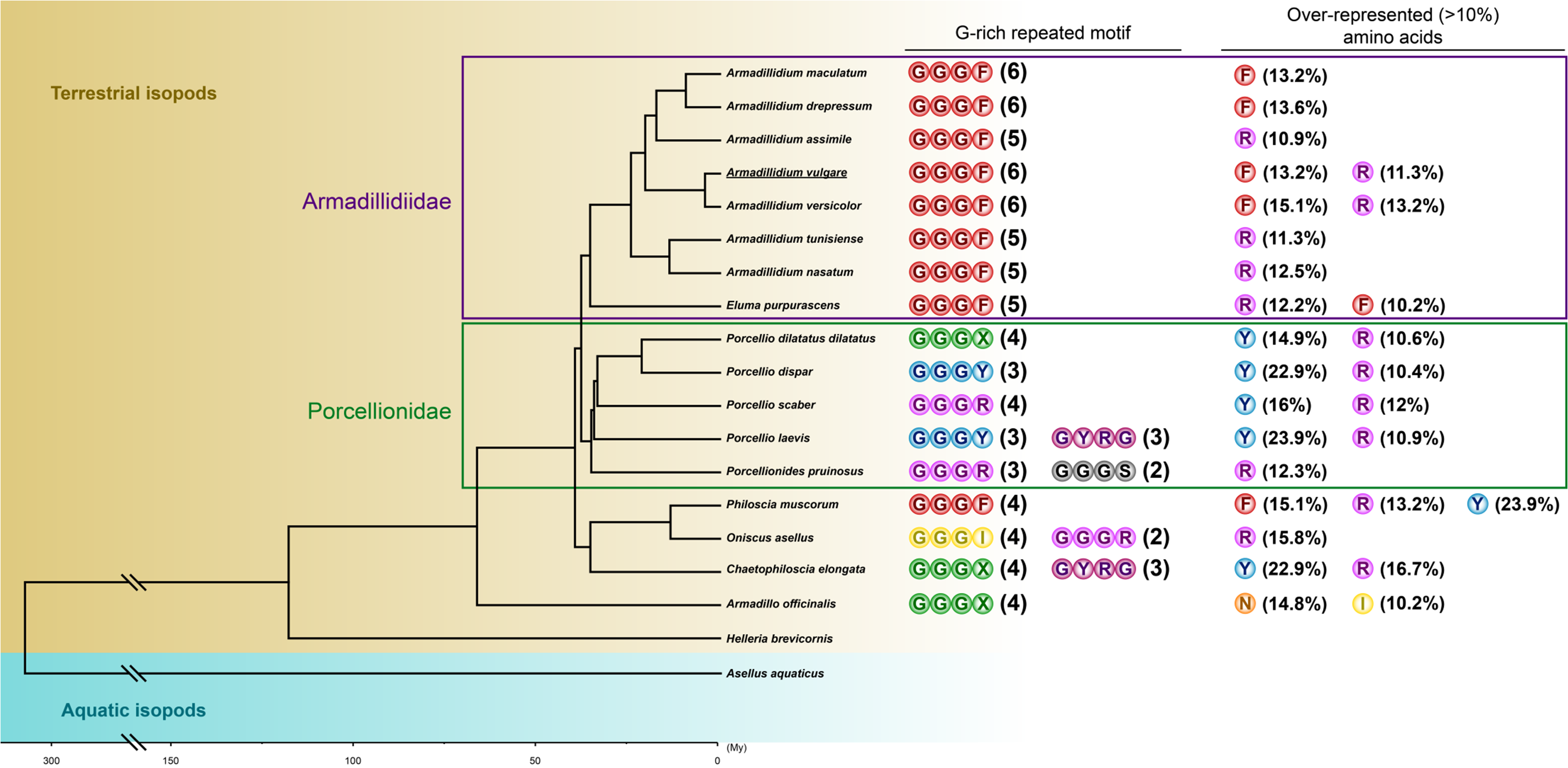{kind=link}
{kind=link}
| Crustin Cluster | Parameter | Mean ± Std | Range | Type (According to Vargas-Albores and Porchas, 2017) | Type (According to Tassanakajon et al., 2015) |
|---|---|---|---|---|---|
| 1 | Size | 153.3 ± 15.8 | 130–166 | II | II |
| Glycine content (%) | 30.1 ± 2.3 | 24.6–32.3 | |||
| Repetitive Tetrapeptide | 8.5 ± 2.2 | 3–11 | |||
| 2 | Size | 38.8 ± 1.4 | 38–41 | I | I |
| Glycine content (%) | 8.0 ± 0.7 | 7.3–9.8 | |||
| Repetitive Tetrapeptide | 0.0 ± 0.0 | 0–0 | |||
| 3 | Size | 61.9 ± 0.2 | 61–62 | I | I |
| Glycine content (%) | 7.2 ± 1.2 | 6.5–9.8 | |||
| Repetitive Tetrapeptide | 0.0 ± 0.0 | 0–0 | |||
| 4 | Size | 39.2 ± 1.6 | 37–41 | I | I |
| Glycine content (%) | 4.8 ± 2.5 | 2.5–9.3 | |||
| Repetitive Tetrapeptide | 0.0 ± 0.0 | 0–0 |
| Organism | Mature Peptide Sequence | Length (aa) | Molecular Weight 1 (Da) | Glycine (%) | pI 1 |
|---|---|---|---|---|---|
| A. assimile | GHIRRPYIGGGGLYGGGGGFHRGGGFHRGGGGFIGGGGFHRGGGFNRGGSYGYNG | 55 | 5354.8 | 50.9 | 11.4 |
| A. depressum | GGFGRPYIGGGGFNRGGGLHRGGGFNSGGGFNRGGGFNRGGGFNRGGGFHRGGSFGYNG | 59 | 5726.1 | 49.2 | 12.1 |
| A. maculatum | GGFGRPYIGGGGFNRGGGFHRGGGFRSGGGFHRGGGFNRGGGFHRGGSYGYNG | 53 | 5252.6 | 49.1 | 11.9 |
| A. nasatum | GHIGRPYIGGGGGGIYRGGGFRTGGGFRTGGGFHRGGGGFHRGGGFHRGGSYGYNG | 56 | 5485.9 | 50 | 11.6 |
| A. tunisiense | GHIGRPYIGGGGIYRGGGFRTGGGFHRGGGFHRGGGFQRGGGFYGGGSYGYNG | 53 | 5268.7 | 49.1 | 11 |
| A. versicolor | GGFGRPYIGGGGFNRGGGFHRGGGFNRGGGFHRGGGFNRGGGFHRGGSFGYNG | 53 | 5263.6 | 49.1 | 12.1 |
| A. vulgare | GHLGRPYIGGGGGFNRGGGFHRGGGFHRGGGFHSGGGFHRGGGFHSGGSFGYR | 53 | 5259.6 | 47.2 | 12 |
| GHLGRPYIGGGGGFNRGGGFHRGGGFHRGGGFQSGGGFHRGGGFHSGGSFGYR | 53 | 5250.6 | 47.2 | 12 | |
| A. officinalis | TFKPCGRSSGGSRCNRGYNRGIIGISGGNNKINGGGDFDDDDYESDYEDYNNGIIGIRGGTNTVNGGGSNNPKDSALKDYNNGIIGIGGGTNTVNGGGSNNPKDSAFKDPRGNRGIIGISGGRNVVQRG | 129 | 13,190.1 | 24.8 | 8.6 |
| C. elongata | TPGRPYYGGGYNGGYRGGYRRGGGFYGGGRFYGGGEGYRGGYYRGYRG | 48 | 5150.5 | 45.8 | 10.1 |
| E. purpurascens | SYVRRPYIGGGGGGFHRGGGFHRGGGFISGGGFHRGGGFNRGGGYGYNG | 49 | 4892.3 | 46.9 | 11.4 |
| P. dilatatusdilatatus | GHHGYGGSYGGRRYGHGGGRFGGIRGGGYGGGGHIGGGYGGYGGYRG | 47 | 4490.7 | 57.4 | 10.1 |
| P. dispar | GYIRKPYIGRGYGGGGYHRGGGFGYGGGYYRGGVGYGGGGYGGYGYRG | 48 | 4855.2 | 52.1 | 9.8 |
| P. laevis | SFIRKPYIGGGYGGYRGGGGYGGYRGGYYRGGGHYGGGYGGYGYRG | 46 | 4729.1 | 50 | 9.8 |
| P. muscorum | TFGRPYYGGGFNRGFGGGYHRGGGFHRGGGFYGGGFRGGYNRGYLG | 46 | 4829.2 | 45.7 | 10.6 |
| P. pruinosus | SYGRGSYGGGSIGRGSFGHGGGSFGRGGGRFGHGGGRFGGIGGGGRYGGGHIGGYRG | 57 | 5307.6 | 54.4 | 11.6 |
| P. scaber | GYIRRPVGYYGGGGGRYGGGRFGGGGGGIGGGRYGGGGRYGGGSYGGYHG | 50 | 4709.0 | 58.0 | 10.2 |
| O. asellus | TYRPSYGGGGGFNRGGGRGGGIHRGGGIGGGIYRGGGIGGGHRGGGGGRFNRGYGYR | 57 | 5485.9 | 52.6 | 11.7 |
| MIC of CE (µM) | MIC of PP (µM) | MIC of Armadillidin H (µM) (Verdon et al., 2016) | ||
|---|---|---|---|---|
| Gram-positive bacteria | Bacillus megaterium F04 | >9.5 | >19 | 2.37 |
| Bacillus pumilus NG1 | 9.5 | 19 | 4.75 | |
| Micrococcus lysodeikticus ATCC 4698 | 4.75 | 4.75 | 2.37 | |
| Staphylococcus aureus ATCC 29213 | >9.5 | >19 | >19 | |
| Staphylococcus warneri RK | >9.5 | >19 | >19 | |
| Gram-negative bacteria | Escherichia coli LMG 2092 | >9.5 | 19 | 9.5 |
| Klebsiella pneumoniae 0502083 | >9.5 | 19 | 19 | |
| Pseudomonas aeruginosa PA14 | >9.5 | >19 | 19 | |
| Pseudomonas fluorescens MFE01 | >9.5 | >19 | 19 | |
| Pseudomonas syringae pv tomato DC3000 | >9.5 | >19 | 4.75 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Becking, T.; Delaunay, C.; Cordaux, R.; Berjeaud, J.-M.; Braquart-Varnier, C.; Verdon, J. Shedding Light on the Antimicrobial Peptide Arsenal of Terrestrial Isopods: Focus on Armadillidins, a New Crustacean AMP Family. Genes 2020, 11, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010093
Becking T, Delaunay C, Cordaux R, Berjeaud J-M, Braquart-Varnier C, Verdon J. Shedding Light on the Antimicrobial Peptide Arsenal of Terrestrial Isopods: Focus on Armadillidins, a New Crustacean AMP Family. Genes. 2020; 11(1):93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010093
Chicago/Turabian StyleBecking, Thomas, Carine Delaunay, Richard Cordaux, Jean-Marc Berjeaud, Christine Braquart-Varnier, and Julien Verdon. 2020. "Shedding Light on the Antimicrobial Peptide Arsenal of Terrestrial Isopods: Focus on Armadillidins, a New Crustacean AMP Family" Genes 11, no. 1: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11010093