Comparative Transcriptomic Analysis of Subcutaneous Adipose Tissue from Local Pig Breeds

 ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Experimental Design and Sampling
2.2. RNA Extraction and Sequencing
2.3. Quality Control, Mapping and Assembly
2.4. Differential Expression Analysis
2.5. Functional Enrichment Analysis
2.6. Real Time Quantitative PCR and Statistical Analysis
3. Results and Discussion
3.1. Alentejano and Bísaro: Local Pig Breeds with Recognizable Meat Quality Traits
3.2. Mapping and Annotation
3.3. Gene Expression Analysis: DESeq2
3.4. Validation by Real Time qPCR
3.5. Functional Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Chen, K.; Baxter, T.; Muir, W.M.; Groenen, M.A.; Schook, L.B. Genetic resources, genome mapping and evolutionary genomics of the pig (Sus scrofa). Int. J. Biol. Sci. 2007, 3, 153–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat deposition, fatty acid composition and meat quality: A review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, C.; Sirtori, F. Quality of meat and meat products produced from southern European pig breeds. Meat Sci. 2012, 90, 511–518. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F. Ham: Dry-cured Ham. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Oxford, UK, 2016; pp. 307–310. [Google Scholar] [CrossRef]
- Gama, L.T.; Martínez, A.M.; Carolino, I.; Landi, V.; Delgado, J.V.; Vicente, A.A.; Vega-Pla, J.L.; Cortés, O.; Sousa, C.O. Genetic structure, relationships and admixture with wild relatives in native pig breeds from Iberia and its islands. Genet. Sel. Evol. GSE 2013, 45, 18. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, M.; Bozzi, R.; García, F.; Núñez, Y.; Geraci, C.; Crovetti, A.; García-Casco, J.; Alves, E.; Škrlep, M.; Charneca, R.; et al. Diversity across major and candidate genes in European local pig breeds. PLoS ONE 2018, 13, e0207475. [Google Scholar] [CrossRef] [Green Version]
- Charneca, R.; Martins, J.; Freitas, A.; Neves, J.; Nunes, J.; Paixim, H.; Bento, P.; Batorek-Lukač, N. Alentejano pig. In European Local Pig Breeds—Diversity and Performance; Candek-Potokar, M., Linan, R.M.N., Eds.; IntechOpen: London, UK, 2019; p. 24. [Google Scholar] [CrossRef] [Green Version]
- Charneca, R.; Nunes, J.; Dividich, J.L. Reproductive and productive traits of sows from Alentejano compared to sows Large-White x Landrace genotype. Rev. Port. Zootec. Edição Electrón. 2012, 1, 17. [Google Scholar]
- Martins, J.M.; Albuquerque, A.; Neves, J.A.; Freitas, A.B.; Charneca, R.; Tirapicos, J.L. Influence of outdoor rearing and oleic acid supplementation on lipid characteristics of muscle and adipose tissues from obese Alentejano pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e578–e590. [Google Scholar] [CrossRef]
- López-Bote, C.J. Sustained utilization of the Iberian pig breed. Meat Sci. 1998, 49, S17–S27. [Google Scholar] [CrossRef]
- Charneca, R.; Martins, J.; Freitas, A.; Neves, J.; Nunes, J.; Elias, M.; Laranjo, M.; Bento, P.; Silva, D.; Paixim, H.; et al. El cruce Alentejano x Bísaro: Un estudio en TREASURE. Sólo Cerdo Ibérico 2018, 39, 26–33. [Google Scholar] [CrossRef]
- Santos e Silva, J.; Ferreira-Cardoso, J.; Bernardo, A.; Costa, J.S.P.d. Conservation and development of the Bísaro pig. Characterization and zootechnical evaluation of the breed for production and genetic management. In Proceedings of the Quality of Meat and Fat in Pigs as Affected by Genetics and Nutrition, Zurich, Switzerland, 25 August 1999; pp. 85–92. [Google Scholar]
- Araújo, J.P.; Cerqueira, J.; Pires, P.; Amorim, I.; Carneiro, M.; Silva, J.S.; Dominguez, R.; Bermúdez, R.; Lorenzo, J.M. Diversidade das raças locais de suínos e produtos tradicionais de qualidade. In Proceedings of the Forum Politécnico #1: Bio-Regiões, Valorização Agro-Industrial e Produção Animal, Beja, Portugal, 21 April 2016; p. 72. [Google Scholar]
- Araújo, J.P.; Cerqueira, J.L.; Pires, P.; Silva, J.S.; Candek-Potokar, M. Influence of traditional and innovative feed on finishing Bísaro pigs in a hoop barn with outdoor access. Rev. Port. Zootec. 2018, III, 436–442. [Google Scholar]
- Amorim, A.; Gonçalves, A.; Leite, A.; Paulos, K.; Oliveira, A.; Pereira, E.; Rodrigues, S.; Teixeira, A. Características da Carcaça. In Porco Bísaro—Qualidade da Carcaça e da Carne; Fernandes, A., Teixeira, A., Eds.; Quinta do Bísaro: Bragança, Portugal, 2015; pp. 33–79. [Google Scholar]
- Araújo, J.P.; Cerqueira, J.; Pires, P.; Amorim, I.; Carneiro, M.; Silva, J.S.; Dominguez, R.; Bermúdez, R.; Lorenzo, J.M. Influence of rearing systems on carcass quality of Bísaro pig breed. In Proceedings of the X Congresso Ibérico Sobre Recursos Genéticos Animais, Castelo Branco, Portugal, 15 September 2016; p. 72. [Google Scholar]
- Santos Silva, J.; Araújo, J.P.; Cerqueira, J.O.; Pires, P.; Alves, C.; Batorek-Lukač, N. Bísaro Pig. In European Local Pig Breeds—Diversity and Performance; Candek-Potokar, M., Linan, R.M.N., Eds.; IntechOpen: London, UK, 2019; p. 13. [Google Scholar] [CrossRef] [Green Version]
- Santos e Silva, J.; Nunes, J.L.T. Inventory and characterization of traditional mediterranean pig production systems. Advantages and constraints towards its development. Acta Agric. Slov. 2013, 4, 61–67. [Google Scholar]
- Paixão, G.; Esteves, A.; Payan-Carreira, R.; Carolino, N. Demographic structure and genetic diversity of the endangered Bísaro pig: Evolution and current status. Czech J. Anim. Sci. 2018, 63, 452–461. [Google Scholar] [CrossRef] [Green Version]
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonergan, S.M.; Topel, D.G.; Marple, D.N. Chapter 5—Fat and fat cells in domestic animals. In The Science of Animal Growth and Meat Technology, 2nd ed.; Lonergan, S.M., Topel, D.G., Marple, D.N., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 51–69. [Google Scholar] [CrossRef]
- Nieto, R.; Miranda, A.; García, M.A.; Aguilera, J.F. The effect of dietary protein content and feeding level on the rate of protein deposition and energy utilization in growing Iberian pigs from 15 to 50 kg body weight. Br. J. Nutr. 2002, 88, 39–49. [Google Scholar] [CrossRef]
- Neel, J.V. Diabetes Mellitus: A “Thrifty” Genotype Rendered Detrimental by “Progress”? Am. J. Hum. Genet. 1962, 14, 353–362. [Google Scholar]
- Zhang, Y.; Proenca, R.; Maffei, M.; Barone, M.; Leopold, L.; Friedman, J.M. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–432. [Google Scholar] [CrossRef]
- Torres-Rovira, L.; Astiz, S.; Caro, A.; Lopez-Bote, C.; Ovilo, C.; Pallares, P.; Perez-Solana, M.L.; Sanchez-Sanchez, R.; Gonzalez-Bulnes, A. Diet-induced swine model with obesity/leptin resistance for the study of metabolic syndrome and type 2 diabetes. Sci. World J. 2012, 2012, 510149. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, G.; Ovilo, C.; Silió, L.; Tomás, A.; Noguera, J.L.; Rodríguez, M.C. Single- and joint-population analyses of two experimental pig crosses to confirm quantitative trait loci on Sus scrofa chromosome 6 and leptin receptor effects on fatness and growth traits. J. Anim. Sci. 2009, 87, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Óvilo, C.; Fernández, A.; Fernández, A.I.; Folch, J.M.; Varona, L.; Benítez, R.; Nuñez, Y.; Rodríguez, C.; Silió, L. Hypothalamic expression of porcine leptin receptor (LEPR), neuropeptide Y (NPY), and cocaine- and amphetamine-regulated transcript (CART) genes is influenced by LEPR genotype. Mamm. Genome 2010, 21, 583–591. [Google Scholar] [CrossRef]
- Kersten, S. Mechanisms of nutritional and hormonal regulation of lipogenesis. EMBO Rep. 2001, 2, 282–286. [Google Scholar] [CrossRef] [Green Version]
- Arner, P. The adipocyte in insulin resistance: Key molecules and the impact of the thiazolidinediones. Trends Endocrinol. Metab. TEM 2003, 14, 137–145. [Google Scholar] [CrossRef]
- Rodríguez-López, J.M.; Lachica, M.; González-Valero, L.; Fernández-Fígares, I. Determining insulin sensitivity from glucose tolerance tests in Iberian and Landrace pigs. bioRxiv 2019. [Google Scholar] [CrossRef]
- Fernandez-Veledo, S.; Nieto-Vazquez, I.; de Castro, J.; Ramos, M.P.; Bruderlein, S.; Moller, P.; Lorenzo, M. Hyperinsulinemia induces insulin resistance on glucose and lipid metabolism in a human adipocytic cell line: Paracrine interaction with myocytes. J. Clin. Endocrinol. Metab. 2008, 93, 2866–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rask-Madsen, C.; King, G.L. Mechanisms of Disease: Endothelial dysfunction in insulin resistance and diabetes. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 46–56. [Google Scholar] [CrossRef]
- Martins, J.M.; Fialho, R.; Albuquerque, A.; Neves, J.; Freitas, A.; Nunes, J.T.; Charneca, R. Growth, blood, carcass and meat quality traits from local pig breeds and their crosses. Animal 2020, 14, 636–647. [Google Scholar] [CrossRef]
- FastQC. Available online: http://www.bioinformatics.bbsrc.ac.uk/projects/fastqc/ (accessed on 25 October 2019).
- Trim Galore. Available online: https://www.bioinformatics.babraham.ac.uk/projects/trim_galore/ (accessed on 25 October 2019).
- SAMtools. Available online: http://www.htslib.org/ (accessed on 31 October 2019).
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2014, 31, 166–169. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [Green Version]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2013, 30, 523–530. [Google Scholar] [CrossRef]
- Peffer, P.L.; Lin, X.; Odle, J. Hepatic beta-oxidation and carnitine palmitoyltransferase I in neonatal pigs after dietary treatments of clofibric acid, isoproterenol, and medium-chain triglycerides. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1518–R1524. [Google Scholar] [CrossRef] [Green Version]
- Vandesompele, J.; Preter, K.D.; Pattyn, F.; Poppe, B.; Roy, N.V.; Paepe, A.D.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, I.K.L. A Concordance Correlation Coefficient to Evaluate Reproducibility. Biometrics 1989, 45, 255–268. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, L.; Zhao, W.; Ren, S.; Tu, F.; Fu, Y.; Li, B.; Wang, X.; Fang, X. Transcriptome sequencing analysis of porcine MDM response to FSL-1 stimulation. Microb. Pathog. 2020, 138, 103830. [Google Scholar] [CrossRef]
- Wang, Y.; Hu, T.; Wu, L.; Liu, X.; Xue, S.; Lei, M. Identification of non-coding and coding RNAs in porcine endometrium. Genomics 2017, 109, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Qi, R.; Wang, J.; Huang, W.; Wu, Y.; Huang, X.; Yang, F.; Huang, J. Differential expression profile of miRNAs in porcine muscle and adipose tissue during development. Gene 2017, 618, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Corominas, J.; Ramayo-Caldas, Y.; Puig-Oliveras, A.; Estellé, J.; Castelló, A.; Alves, E.; Pena, R.N.; Ballester, M.; Folch, J.M. Analysis of porcine adipose tissue transcriptome reveals differences in de novo fatty acid synthesis in pigs with divergent muscle fatty acid composition. BMC Genom. 2013, 14, 843. [Google Scholar] [CrossRef]
- Puig-Oliveras, A.; Ramayo-Caldas, Y.; Corominas, J.; Estellé, J.; Pérez-Montarelo, D.; Hudson, N.J.; Casellas, J.; Folch, J.M.; Ballester, M. Differences in Muscle Transcriptome among Pigs Phenotypically Extreme for Fatty Acid Composition. PLoS ONE 2014, 9, e99720. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.; Fernández, A.; Núñez, Y.; Benítez, R.; Isabel, B.; Barragán, C.; Fernández, A.I.; Rey, A.I.; Medrano, J.F.; Cánovas, A.; et al. Comparative Analysis of Muscle Transcriptome between Pig Genotypes Identifies Genes and Regulatory Mechanisms Associated to Growth, Fatness and Metabolism. PLoS ONE 2015, 10, e0145162. [Google Scholar] [CrossRef] [Green Version]
- Ramayo-Caldas, Y.; Mach, N.; Esteve-Codina, A.; Corominas, J.; Castello, A.; Ballester, M.; Estelle, J.; Ibanez-Escriche, N.; Fernandez, A.I.; Perez-Enciso, M.; et al. Liver transcriptome profile in pigs with extreme phenotypes of intramuscular fatty acid composition. BMC Genom. 2012, 13, 18. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Ai, H.; Ren, J.; Li, W.; Li, P.; Qiao, R.; Ouyang, J.; Yang, M.; Ma, J.; Huang, L. A global view of porcine transcriptome in three tissues from a full-sib pair with extreme phenotypes in growth and fat deposition by paired-end RNA sequencing. BMC Genom. 2011, 12, 448. [Google Scholar] [CrossRef] [Green Version]
- Benítez, R.; Trakooljul, N.; Núñez, Y.; Isabel, B.; Murani, E.; De Mercado, E.; Gómez-Izquierdo, E.; García-Casco, J.; López-Bote, C.; Wimmers, K.; et al. Breed, Diet, and Interaction Effects on Adipose Tissue Transcriptome in Iberian and Duroc Pigs Fed Different Energy Sources. Genes (Basel) 2019, 10, 589. [Google Scholar] [CrossRef] [Green Version]
- Caicedo, A.; Pereira, E.; Margolskee, R.F.; Roper, S.D. Role of the G-protein subunit alpha-gustducin in taste cell responses to bitter stimuli. J. Neurosci. 2003, 23, 9947–9952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribani, A.; Bertolini, F.; Schiavo, G.; Scotti, E.; Utzeri, V.J.; Dall’Olio, S.; Trevisi, P.; Bosi, P.; Fontanesi, L. Next generation semiconductor based sequencing of bitter taste receptor genes in different pig populations and association analysis using a selective DNA pool-seq approach. Anim. Genet. 2017, 48, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gnoni, G.V.; Priore, P.; Geelen, M.J.; Siculella, L. The mitochondrial citrate carrier: Metabolic role and regulation of its activity and expression. IUBMB Life 2009, 61, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Hayakawa, K.; Bateman, K.S.; Fraser, M.E. Identification of the Citrate-binding Site of Human ATP-Citrate Lyase Using X-ray Crystallography. J. Biol. Chem. 2010, 285, 27418–27428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berg, J.M.; Tymoczko, J.L.; Gatto, G.J.; Stryer, L. Biochemistry, 8th ed.; W.H. Freeman: New York, NY, USA, 2015. [Google Scholar]
- Lehninger, A.L.; Nelson, D.L.; Cox, M.M. Lehninger Principles of Biochemistry; W.H. Freeman: New York, NY, USA, 2008. [Google Scholar]
- Matsuzaka, T.; Shimano, H. Elovl6: A new player in fatty acid metabolism and insulin sensitivity. J. Mol. Med. 2009, 87, 379–384. [Google Scholar] [CrossRef] [Green Version]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-CoA desaturase. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E28–E37. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, A.; Rodrigues, S. Pork Meat Quality of Preto Alentejano and Commercial Largewhite Landrace Cross. J. Integr. Agric. 2013, 12, 1961–1971. [Google Scholar] [CrossRef]
- Liang, Y.-J.; Jiang, J.-G. Characterization of malic enzyme and the regulation of its activity and metabolic engineering on lipid production. RSC Adv. 2015, 5, 45558–45570. [Google Scholar] [CrossRef]
- González-Manchón, C.; Ferrer, M.; Ayuso, M.S.; Parrilla, R. Cloning, sequencing and functional expression of a cDNA encoding a NADP-dependent malic enzyme from human liver. Gene 1995, 159, 255–260. [Google Scholar] [CrossRef]
- Millward, C.A.; Desantis, D.; Hsieh, C.-W.; Heaney, J.D.; Pisano, S.; Olswang, Y.; Reshef, L.; Beidelschies, M.; Puchowicz, M.; Croniger, C.M. Phosphoenolpyruvate carboxykinase (Pck1) helps regulate the triglyceride/fatty acid cycle and development of insulin resistance in mice. J. Lipid Res. 2010, 51, 1452–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latorre, P.; Burgos, C.; Hidalgo, J.; Varona, L.; Carrodeguas, J.A.; López-Buesa, P. c.A2456C-substitution in Pck1 changes the enzyme kinetic and functional properties modifying fat distribution in pigs. Sci. Rep. 2016, 6, 19617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montez, J.M.; Soukas, A.; Asilmaz, E.; Fayzikhodjaeva, G.; Fantuzzi, G.; Friedman, J.M. Acute leptin deficiency, leptin resistance, and the physiologic response to leptin withdrawal. Proc. Natl. Acad. Sci. USA 2005, 102, 2537–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stern, J.H.; Rutkowski, J.M.; Scherer, P.E. Adiponectin, Leptin, and Fatty Acids in the Maintenance of Metabolic Homeostasis through Adipose Tissue Crosstalk. Cell Metab. 2016, 23, 770–784. [Google Scholar] [CrossRef]
- Benítez, R.; Fernández, A.; Isabel, B.; Núñez, Y.; Mercado, E.D.; Gómez-Izquierdo, E.; García-Casco, J.; López-Bote, C.; Óvilo, C. Modulatory Effects of Breed, Feeding Status, and Diet on Adipogenic, Lipogenic, and Lipolytic Gene Expression in Growing Iberian and Duroc Pigs. Int. J. Mol. Sci. 2018, 19, 22. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Fígares, I.; Lachica, M.; Nieto, R.; Rivera-Ferre, M.G.; Aguilera, J.F. Serum profile of metabolites and hormones in obese (Iberian) and lean (Landrace) growing gilts fed balanced or lysine deficient diets. Livest. Sci. 2007, 110, 73–81. [Google Scholar] [CrossRef]
- Pedersen, T.A.; Bereshchenko, O.; Garcia-Silva, S.; Ermakova, O.; Kurz, E.; Mandrup, S.; Porse, B.T.; Nerlov, C. Distinct C/EBPalpha motifs regulate lipogenic and gluconeogenic gene expression in vivo. EMBO J. 2007, 26, 1081–1093. [Google Scholar] [CrossRef] [Green Version]
- Olofsson, L.E.; Orho-Melander, M.; William-Olsson, L.; Sjoholm, K.; Sjostrom, L.; Groop, L.; Carlsson, B.; Carlsson, L.M.; Olsson, B. CCAAT/enhancer binding protein alpha (C/EBPalpha) in adipose tissue regulates genes in lipid and glucose metabolism and a genetic variation in C/EBPalpha is associated with serum levels of triglycerides. J. Clin. Endocrinol. Metab. 2008, 93, 4880–4886. [Google Scholar] [CrossRef] [Green Version]
- Matulewicz, N.; Stefanowicz, M.; Nikołajuk, A.; Karczewska-Kupczewska, M. Markers of Adipogenesis, but Not Inflammation, in Adipose Tissue Are Independently Related to Insulin Sensitivity. J. Clin. Endocrinol. Metab. 2017, 102, 3040–3049. [Google Scholar] [CrossRef] [Green Version]
- Acosta, J.R.; Douagi, I.; Andersson, D.P.; Bäckdahl, J.; Rydén, M.; Arner, P.; Laurencikiene, J. Increased fat cell size: A major phenotype of subcutaneous white adipose tissue in non-obese individuals with type 2 diabetes. Diabetologia 2016, 59, 560–570. [Google Scholar] [CrossRef] [Green Version]
- Dixon, T.M.; Daniel, K.W.; Farmer, S.R.; Collins, S. CCAAT/enhancer-binding protein alpha is required for transcription of the beta 3-adrenergic receptor gene during adipogenesis. J. Biol. Chem. 2001, 276, 722–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michal, J.J.; Zhang, Z.W.; Gaskins, C.T.; Jiang, Z. The bovine fatty acid binding protein 4 gene is significantly associated with marbling and subcutaneous fat depth in Wagyu x Limousin F2 crosses. Anim. Genet. 2006, 37, 400–402. [Google Scholar] [CrossRef] [PubMed]
- Shi-Zheng, G.; Zhao, S.-M. Physiology, Affecting Factors and Strategies for Control of Pig Meat Intramuscular Fat. Recent Pat. Food Nutr. Agric. 2009, 1, 59–74. [Google Scholar] [CrossRef] [Green Version]
- Garin-Shkolnik, T.; Rudich, A.; Hotamisligil, G.S.; Rubinstein, M. FABP4 attenuates PPARgamma and adipogenesis and is inversely correlated with PPARgamma in adipose tissues. Diabetes 2014, 63, 900–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furuhashi, M.; Saitoh, S.; Shimamoto, K.; Miura, T. Fatty Acid-Binding Protein 4 (FABP4): Pathophysiological Insights and Potent Clinical Biomarker of Metabolic and Cardiovascular Diseases. Clin. Med. Insights Cardiol. 2015, 8, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Alfares, M.N.; Perks, C.M.; Hamilton-Shield, J.P.; Holly, J.M.P. Insulin-like growth factor-II in adipocyte regulation: Depot-specific actions suggest a potential role limiting excess visceral adiposity. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1098–E1107. [Google Scholar] [CrossRef] [PubMed]
- Van Laere, A.-S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef]
- Criado-Mesas, L.; Ballester, M.; Crespo-Piazuelo, D.; Castelló, A.; Benítez, R.; Fernández, A.I.; Folch, J.M. Analysis of porcine IGF2 gene expression in adipose tissue and its effect on fatty acid composition. PLoS ONE 2019, 14, e0220708. [Google Scholar] [CrossRef] [Green Version]
- Óvilo, C.; Benítez, R.; Fernández, A.; Núñez, Y.; Ayuso, M.; Fernández, A.I.; Rodríguez, C.; Isabel, B.; Rey, A.I.; López-Bote, C.; et al. Longissimus dorsi transcriptome analysis of purebred and crossbred Iberian pigs differing in muscle characteristics. BMC Genom. 2014, 15, 413. [Google Scholar] [CrossRef] [Green Version]
- Munroe, M.E.; Bishop, G.A. A costimulatory function for T cell CD40. J. Immunol. 2007, 178, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Yi, Z.; Bishop, G.A. Regulatory role of CD40 in obesity-induced insulin resistance. Adipocyte 2014, 4, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.-A.; Kogan, S.; Amano, S.U.; Wang, M.; Dagdeviren, S.; Friedline, R.H.; Aouadi, M.; Kim, J.K.; Czech, M.P. CD40 deficiency in mice exacerbates obesity-induced adipose tissue inflammation, hepatic steatosis, and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E951–E963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopchick, J.J.; Berryman, D.E.; Puri, V.; Lee, K.Y.; Jorgensen, J.O.L. The effects of growth hormone on adipose tissue: Old observations, new mechanisms. Nat. Rev. Endocrinol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Hajjari, M.; Salavaty, A. HOTAIR: An oncogenic long non-coding RNA in different cancers. Cancer Biol. Med. 2015, 12, 1–9. [Google Scholar] [CrossRef]
- Divoux, A.; Karastergiou, K.; Xie, H.; Guo, W.; Perera, R.J.; Fried, S.K.; Smith, S.R. Identification of a novel lncRNA in gluteal adipose tissue and evidence for its positive effect on preadipocyte differentiation. Obesity (Silver Spring) 2014, 22, 1781–1785. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.D.; Yuan, L.L.; Lin, L.Q. LncRNA HOTAIR contributes to the tumorigenesis of nasopharyngeal carcinoma via up-regulating FASN. Eur. Rev. Med Pharmacol. Sci. 2017, 21, 5143–5152. [Google Scholar] [CrossRef]
- Nilsson, L.A.; Roepstorff, C.; Kiens, B.; Billig, H.; Ling, C. Prolactin suppresses malonyl-CoA concentration in human adipose tissue. Horm. Metab. Res. 2009, 41, 747–751. [Google Scholar] [CrossRef]
- Pala, N.A.; Laway, B.A.; Misgar, R.A.; Dar, R.A. Metabolic abnormalities in patients with prolactinoma: Response to treatment with cabergoline. Diabetol. Metab. Syndr. 2015, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, L.; Binart, N.; Bohlooly, Y.M.; Bramnert, M.; Egecioglu, E.; Kindblom, J.; Kelly, P.A.; Kopchick, J.J.; Ormandy, C.J.; Ling, C.; et al. Prolactin and growth hormone regulate adiponectin secretion and receptor expression in adipose tissue. Biochem. Biophys. Res. Commun. 2005, 331, 1120–1126. [Google Scholar] [CrossRef] [Green Version]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef]
- Kim, S.J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase to Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol. Cell. Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.J.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [Green Version]
- García-Contreras, C.; Madsen, O.; Groenen, M.A.M.; López-García, A.; Vázquez-Gómez, M.; Astiz, S.; Núñez, Y.; Benítez, R.; Fernández, A.; Isabel, B.; et al. Impact of genotype, body weight and sex on the prenatal muscle transcriptome of Iberian pigs. PLoS ONE 2020, 15, e0227861. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.R.; Kim, H.J.; Xu, X.; Ferrante, A.W., Jr. Macrophage and adipocyte IGF1 maintain adipose tissue homeostasis during metabolic stresses. Obesity (Silver Spring) 2016, 24, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Musarò, A.; McCullagh, K.J.A.; Naya, F.J.; Olson, E.N.; Rosenthal, N. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature 1999, 400, 581–585. [Google Scholar] [CrossRef]
- LeRoith, D.; Yakar, S. Mechanisms of Disease: Metabolic effects of growth hormone and insulin-like growth factor 1. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 302–310. [Google Scholar] [CrossRef]
- Yen, P.M. Physiological and Molecular Basis of Thyroid Hormone Action. Physiol. Rev. 2001, 81, 1097–1142. [Google Scholar] [CrossRef] [Green Version]
- Febbraio, M.A. Role of interleukins in obesity: Implications for metabolic disease. Trends Endocrinol. Metab. 2014, 25, 312–319. [Google Scholar] [CrossRef]
- Karagiannis, F.; Wilhelm, C. More Is Less: IL-9 in the Resolution of Inflammation. Immunity 2017, 47, 403–405. [Google Scholar] [CrossRef] [Green Version]
- Matsuzawa, Y. Adiponectin: Identification, physiology and clinical relevance in metabolic and vascular disease. Atheroscler. Suppl. 2005, 6, 7–14. [Google Scholar] [CrossRef]
- Fang, H.; Judd, R.L. Adiponectin Regulation and Function. In Comprehensive Physiology; Pollock, D.M., Ed.; American Physiological Society: Rockville, MD, USA, 2018; pp. 1031–1063. [Google Scholar] [CrossRef]
- Pivovarova, O.; Gogebakan, O.; Sucher, S.; Groth, J.; Murahovschi, V.; Kessler, K.; Osterhoff, M.; Rudovich, N.; Kramer, A.; Pfeiffer, A.F. Regulation of the clock gene expression in human adipose tissue by weight loss. Int. J. Obes. 2016, 40, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.; Chatterjee, S.; Yin, H.; Liu, R.; Lee, J.; Yechoor, V.K.; Ma, K. Novel Function of Rev-erbα in Promoting Brown Adipogenesis. Sci. Rep. 2015, 5, 11239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kojetin, D.J.; Burris, T.P. A role for rev-erbalpha ligands in regulation of adipogenesis. Curr. Pharm. Des. 2011, 17, 320–324. [Google Scholar] [CrossRef]
- Madison, B.B. Srebp2: A master regulator of sterol and fatty acid synthesis. J. Lipid Res. 2016, 57, 333–335. [Google Scholar] [CrossRef] [Green Version]
- Miserez, A.R.; Muller, P.Y.; Barella, L.; Barella, S.; Staehelin, H.B.; Leitersdorf, E.; Kark, J.D.; Friedlander, Y. Sterol-regulatory element-binding protein (SREBP)-2 contributes to polygenic hypercholesterolaemia. Atherosclerosis 2002, 164, 15–26. [Google Scholar] [CrossRef]
- Crewe, C.; Zhu, Y.; Paschoal, V.A.; Joffin, N.; Ghaben, A.L.; Gordillo, R.; Oh, D.Y.; Liang, G.; Horton, J.D.; Scherer, P.E. SREBP-regulated adipocyte lipogenesis is dependent on substrate availability and redox modulation of mTORC1. JCI Insight 2019, 4. [Google Scholar] [CrossRef]
- Ge, K.; Cho, Y.-W.; Guo, H.; Hong, T.B.; Guermah, M.; Ito, M.; Yu, H.; Kalkum, M.; Roeder, R.G. Alternative Mechanisms by Which Mediator Subunit MED1/TRAP220 Regulates Peroxisome Proliferator-Activated Receptor γ-Stimulated Adipogenesis and Target Gene Expression. Mol. Cell. Biol. 2008, 28, 1081–1091. [Google Scholar] [CrossRef] [Green Version]
- Sarjeant, K.; Stephens, J.M. Adipogenesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008417. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, M.; Takahashi, H.; Saeki, Y.; Ozaki, T.; Itoh, S.; Suzuki, M.; Mizushima, W.; Tanaka, K.; Hatakeyama, S. The E3 ubiquitin ligase TRIM23 regulates adipocyte differentiation via stabilization of the adipogenic activator PPARγ. eLife 2015, 4, e05615. [Google Scholar] [CrossRef]
- Rosen, E.D.; MacDougald, O.A. Adipocyte differentiation from the inside out. Nat. Rev. Mol. Cell Biol. 2006, 7, 885–896. [Google Scholar] [CrossRef]
- Albuquerque, A.; Neves, J.A.; Redondeiro, M.; Laranjo, M.; Félix, M.R.; Freitas, A.; Tirapicos, J.L.; Martins, J.M. Long term betaine supplementation regulates genes involved in lipid and cholesterol metabolism of two muscles from an obese pig breed. Meat Sci. 2017, 124, 25–33. [Google Scholar] [CrossRef]
- Dong, X.-Y.; Tang, S.-Q.; Chen, J.-D. Dual functions of Insig proteins in cholesterol homeostasis. Lipids Health Dis. 2012, 11, 173. [Google Scholar] [CrossRef] [Green Version]
- Burg, J.S.; Espenshade, P.J. Regulation of HMG-CoA reductase in mammals and yeast. Prog. Lipid Res. 2011, 50, 403–410. [Google Scholar] [CrossRef] [Green Version]
- Junye, C.; Yi, L.; Mengyuan, T.; Qiren, H. Molecular mechanisms of FOXO1 in adipocyte differentiation. J. Mol. Endocrinol. 2019, 62, R239–R253. [Google Scholar] [CrossRef]
- Xu, M.; Chen, X.; Chen, D.; Yu, B.; Huang, Z. FoxO1: A novel insight into its molecular mechanisms in the regulation of skeletal muscle differentiation and fiber type specification. Oncotarget 2017, 8, 10662–10674. [Google Scholar] [CrossRef] [Green Version]



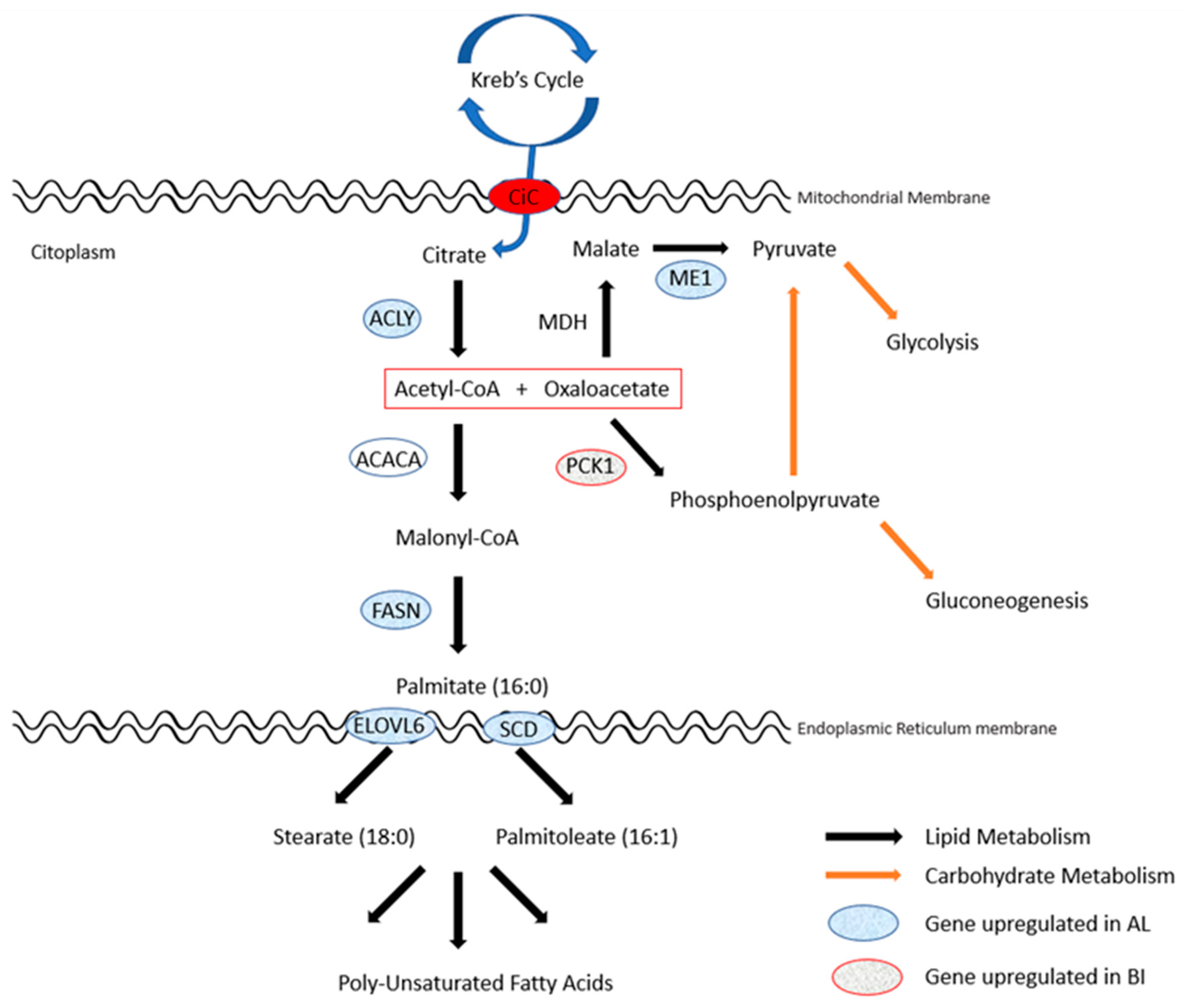{kind=link}
{kind=link}
| Genes | RNA-seq | Real Time qPCR | Correlation | |||
|---|---|---|---|---|---|---|
| Log2 FC | q-Value | Log2 FC | p-Value | r | p-Value | |
| ACACA | 0.854 | 0.116 | 1.055 | 0.077 | 0.986 | 7.07 × 10−6 |
| ACLY | 1.853 | 0.005 | 1.601 | 0.068 | 0.864 | 0.006 |
| ADIPOQ | −0.625 | 0.297 | −0.685 | 0.110 | 0.707 | 0.050 |
| ELOVL6 | 1.236 | 0.009 | 0.671 | 0.136 | 0.650 | 0.081 |
| FASN | 1.691 | 0.002 | 1.359 | 0.100 | 0.920 | 0.001 |
| LEP | 1.376 | 0.037 | 0.929 | 0.046 | 0.739 | 0.036 |
| ME1 | 2.090 | 8.42 × 10−6 | 1.008 | 0.106 | 0.757 | 0.030 |
| PCK1 | −1.959 | 0.037 | −2.660 | 0.001 | 0.820 | 0.013 |
| SCD | 2.311 | 0.026 | 1.351 | 0.087 | 0.643 | 0.086 |
| n = 4 | ||||||
| Master Regulator | Participating Regulators | Predicted Activation (AL) | Z-Score | p-Value | Target Molecules in Dataset |
|---|---|---|---|---|---|
| INSIG2 | INSIG2 | Inhibited | −2.236 | 3.74 × 10−6 | 5 |
| FLCN | Esrra, FLCN, MTORC1, PPARGC1A | Inhibited | −2.236 | 7.72 × 10−6 | 20 |
| PKIA | CREB1, Pka, PKIA | Inhibited | −2.887 | 1.26 × 10−5 | 12 |
| LRAT | Akt, INSR, JAK2, LRAT, RPE65, STAT5a/b | Inhibited | −3.300 | 3.03 × 10−5 | 18 |
| NRG4 | NRG4 | Inhibited | −2.000 | 3.76 × 10−5 | 4 |
| PDGF BB | PDGF BB | Inhibited | −2.333 | 1.54 × 10−4 | 9 |
| CREB1 | CREB1 | Inhibited | −2.828 | 1.69 × 10−4 | 8 |
| GNAS | GNAS | Inhibited | −2.236 | 1.97 × 10−4 | 5 |
| MFSD2A | MFSD2A | Inhibited | −2.000 | 4.85 × 10−4 | 4 |
| MEF2C | MEF2C | Inhibited | −2.236 | 1.47 × 10−3 | 5 |
| HSP27 | Hsp27 | Inhibited | −2.000 | 1.63 × 10−3 | 4 |
| NDFIP1 | Akt, Jnk, Map3k7, NDFIP1, PTEN, SRC | Activated | 2.449 | 2.31 × 10−6 | 24 |
| NR1D1 | NR1D1 | Activated | 2.236 | 5.36 × 10−6 | 5 |
| NSUN3 | NSUN3 | Activated | 2.000 | 5.61 × 10−6 | 4 |
| ALKBH1 | ALKBH1 | Activated | 2.000 | 5.61 × 10−6 | 4 |
| UCHL3 | Akt, AMPK, FOXO1, GSK3B, INSR, UCHL3 | Activated | 2.449 | 1.05 × 10−5 | 24 |
| MIR-29B-3P (AND OTHER MIRNAS W/SEED AGCACCA) | Akt, ERK, miR-29b-3p (and other miRNAs w/seed AGCACCA), PMP22 | Activated | 2.500 | 1.06 × 10−5 | 16 |
| ARNTL | Akt, ARNTL, CLOCK, NFE2L2 | Activated | 2.200 | 2.85 × 10−5 | 25 |
| IDH1 | IDH1 | Activated | 2.236 | 2.97 × 10−5 | 5 |
| EPCAM | EPCAM | Activated | 2.000 | 3.76 × 10−5 | 4 |
| ATP7B | Akt, ATP7B | Activated | 2.887 | 4.58 × 10−5 | 12 |
| DAP3 | DAP3 | Activated | 2.000 | 5.36 × 10−5 | 4 |
| BACE1 | BACE1, CREB1, Pka | Activated | 2.714 | 7.70 × 10−5 | 11 |
| ATP7B | ATP7B | Activated | 2.236 | 3.98 × 10−4 | 5 |
| MLXIPL | MLXIPL | Activated | 2.000 | 4.02 × 10−4 | 4 |
| SCAP | SCAP | Activated | 2.449 | 4.21 × 10−4 | 6 |
| SREBF2 | SREBF2 | Activated | 2.449 | 6.09 × 10−4 | 6 |
| GHRL | GHRL | Activated | 2.000 | 8.06 × 10−4 | 4 |
| MED1 | MED1 | Activated | 2.449 | 1.00 × 10−3 | 6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albuquerque, A.; Óvilo, C.; Núñez, Y.; Benítez, R.; López-Garcia, A.; García, F.; Félix, M.d.R.; Laranjo, M.; Charneca, R.; Martins, J.M. Comparative Transcriptomic Analysis of Subcutaneous Adipose Tissue from Local Pig Breeds. Genes 2020, 11, 422. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040422
Albuquerque A, Óvilo C, Núñez Y, Benítez R, López-Garcia A, García F, Félix MdR, Laranjo M, Charneca R, Martins JM. Comparative Transcriptomic Analysis of Subcutaneous Adipose Tissue from Local Pig Breeds. Genes. 2020; 11(4):422. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040422
Chicago/Turabian StyleAlbuquerque, André, Cristina Óvilo, Yolanda Núñez, Rita Benítez, Adrián López-Garcia, Fabián García, Maria do Rosário Félix, Marta Laranjo, Rui Charneca, and José Manuel Martins. 2020. "Comparative Transcriptomic Analysis of Subcutaneous Adipose Tissue from Local Pig Breeds" Genes 11, no. 4: 422. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11040422