CDK Regulation of Meiosis: Lessons from S. cerevisiae and S. pombe


Department of Biology, Indiana University, 1001 E. Third Street, Bloomington, IN 47405, USA
*
Author to whom correspondence should be addressed.
Genes 2020, 11(7), 723; https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070723
Submission received: 16 May 2020
/
Revised: 26 June 2020
/
Accepted: 26 June 2020
/
Published: 29 June 2020
(This article belongs to the Special Issue Chromosome Segregation Defects in the Origin of Genomic Instability)
Abstract
:Meiotic progression requires precise orchestration, such that one round of DNA replication is followed by two meiotic divisions. The order and timing of meiotic events is controlled through the modulation of the phosphorylation state of proteins. Key components of this phospho-regulatory system include cyclin-dependent kinase (CDK) and its cyclin regulatory subunits. Over the past two decades, studies in budding and fission yeast have greatly informed our understanding of the role of CDK in meiotic regulation. In this review, we provide an overview of how CDK controls meiotic events in both budding and fission yeast. We discuss mechanisms of CDK regulation through post-translational modifications and changes in the levels of cyclins. Finally, we highlight the similarities and differences in CDK regulation between the two yeast species. Since CDK and many meiotic regulators are highly conserved, the findings in budding and fission yeasts have revealed conserved mechanisms of meiotic regulation among eukaryotes.
Keywords:
meiosis; Cyclin-Dependent Kinase; CDK; cyclin; APC/C; budding yeast; fission yeast; chromosome segregation1. Introduction
Control of the eukaryotic cell cycle occurs through the modulation of phosphorylation states of proteins that trigger specific events. At the forefront of this phospho-regulation are the cyclin-dependent kinases (CDKs), whose oscillatory activity results in a large number of phosphorylations that change the activation state of their substrates [1,2]. Ultimately, CDK controls the cell cycle by regulating many processes including DNA replication in S phase (synthesis) and chromosome segregation in M phase (mitosis). Similarly, CDK is also essential for meiotic regulation, with additional roles in ensuring meiosis-specific events.
The genes encoding CDKs were originally discovered in landmark genetic screens performed by Lee Hartwell and Paul Nurse in budding and fission yeast, respectively [3,4,5]. In addition, further analysis of CDK regulation in these model organisms proves to be foundational for our understanding of the eukaryotic cell cycle. Both yeasts encode one CDK, Cdk1, that governs both the mitotic and meiotic cell cycles. Cdk1 is present throughout the cell cycle, but its oscillatory activity is dependent on the regulatory subunits known as cyclins [1,2]. Cyclins stimulate the activity of CDK and specify substrates for phosphorylation. In addition, Cdk1 and cyclins can be regulated by post-translational mechanisms and through the binding of inhibitors.
There are many similarities to the regulation of CDK activity in the budding and fission yeast mitotic cell cycle. In G1 phase (Gap 1), CDK activity is low [6,7,8,9]. With signals such as cell growth, the G1 cyclin levels rise and bind CDK (G1-CDK). G1-CDK phosphorylates and inhibits the CDK inhibitors and the Anaphase Promoting Complex/Cyclosome (APC/C), a ubiquitin ligase that targets the S and M phase cyclins for proteasomal degradation [10,11,12,13,14,15]. With the inhibition of the APC/C, S phase cyclins increase and bind CDK (S-CDK). Phosphorylation of S-CDK substrates leads to the initiation of DNA replication and S phase progression [16,17]. M phase cyclins are expressed as cells exit S phase [18,19]. In fission yeast, CDK bound to M phase cyclins (M-CDK) is inhibited by the Wee1 and Mik1 kinases, which place an inhibitory phosphorylation on CDK [20,21,22]. The inhibition of CDK allows further growth in G2 (Gap 2) before transitioning into M phase. Once cells are ready to exit G2, Cdc25 phosphatase removes the inhibitory phosphorylation, unleashing active M-CDK [22,23,24,25]. Budding yeast has a Wee1 homolog called Swe1. However, Swe1 is not thought to have as important of a role in normal cell cycle progression [26,27]. Instead, M-CDK is activated with the production of the M phase cyclins. M-CDK activity controls many events in M phase such as chromosome condensation, spindle assembly, and chromosome attachment on the bipolar spindle [28,29,30]. M-CDK also activates its inhibitor, the APC/C, allowing the cells to transition into anaphase [31,32]. CDK activity declines in anaphase because the cyclins are degraded, CDK inhibitors are activated, and the phosphatase Cdc14 in budding yeast, or Clp1 in fission yeast, removes the phosphorylations from CDK substrates [28,33,34,35,36,37]. Cells will exit M phase and enter into G1 with low CDK activity.
Budding and fission yeast have also been instrumental in revealing basic principles of meiotic regulation. Starvation of key nutrients induces meiosis in both organisms [38]. Cells undergo one round of DNA replication followed by a prolonged prophase I, in which homologous chromosomes pair, synapse (in budding yeast) and recombine (Figure 1) [39,40]. In meiosis I, the paired homologous chromosomes attach to spindle microtubules and segregate. In meiosis II, sister chromatid kinetochores attach to spindle microtubules and separate. The four haploid meiotic products are packaged into spores and germinate when nutrients return.
CDK regulates many meiotic events, with important roles in S and M phases, similar to the functions of CDK in the mitotic cell cycle. In addition, CDK regulates meiosis-specific events to ensure that homologs make interactions in prophase I, that DNA is not re-replicated in between the two meiotic divisions, and that cells undergo two and only two meiotic divisions. In this review, we highlight the complex regulation of Cdk1 and cyclins in meiosis in budding and fission yeast. We discuss how the expression of cyclin genes is regulated, how post-translational mechanisms of cyclins and Cdk1 affect activity, and how degradation of the cyclins and other cell cycle proteins is regulated. Finally, we compare the regulation between the two model organisms to highlight the differences in regulation that accomplish a similar outcome.
2. Overview of S. cerevisiae CDK Activity
In budding yeast, progression through meiosis is governed by the oscillatory activity of CDK and a CDK-related kinase Ime2. Budding yeast have six conserved CDKs, but only one, Cdk1 (or Cdc28), is required for mitotic and meiotic cell cycle control [28]. One other CDK, Pho85, has minor roles in G1 and S phase and may function to fine-tune cell cycle regulation, or may have more important functions under specific environmental conditions [41,42,43]. Whether Pho85 has a similar role in meiosis has not been investigated. There are nine cyclins that bind and activate Cdk1 in the mitotic cell cycle, the G1 cyclins Cln1, Cln2, and Cln3, and six B-type cyclins Clb1-6. In meiosis, the G1 cyclins are not expressed. Instead, Ime2 is required for the meiotic G1/S transition [44,45]. Cdk1 bound to Clb5 and Clb6 are important for DNA replication in premeiotic S phase [44,46] (Figure 1). Ime2 and CDK bound to B-type cyclins regulate the meiotic divisions [47,48]. The major mitotic cyclin, Clb2, is not expressed in meiosis [49,50]. Instead, Clb1 and Clb3 are considered the major meiosis I and meiosis II cyclins, respectively [51].
Full Cdk1 activity requires the binding of cyclins as well as the phosphorylation of the T-loop by CDK activating kinase Cak1 [28]. The non-phosphorylated T-loop blocks the active site of Cdk1, and its phosphorylation by Cak1 is thought to reposition the T-loop to allow access to the substrate-binding domain and to increase the affinity for cyclins [52,53,54,55,56,57,58]. Cak1 also phosphorylates the Ime2 activation loop, which results in Ime2 autophosphorylation and activation [59]. Although Ime2 is related to CDKs, it does not interact with cyclins to function [60].
CDK activity is downregulated at meiotic entry and during anaphase I and anaphase II. Different mechanisms are used to attenuate CDK activity. The CDK inhibitor Sic1 keeps CDK activity low as cells enter meiosis [44,47]. Post-translational modifications of the cyclins, such as phosphorylation, can decrease CDK activity at anaphase I [51,61,62,63]. Also, some cyclins are degraded in prophase I, anaphase I, and anaphase II [64,65,66]. The APC/C binds to either the co-activator Cdc20, or the meiosis-specific co-activator Ama1 to ubiquitinate and target the cyclins for proteasomal degradation (Table 1) [64,65,66,67,68].
2.1. Entry into Meiosis and Premeiotic DNA Replication
In budding yeast, meiosis is orchestrated through a series of transcriptional cascades. A transcriptional activator, Ime1, is induced in diploid cells through the integration of a number of external signals including nitrogen levels, carbon source, external pH, and presence of both mating type loci [38,69,70,71,72,73]. Interestingly, G1-CDK activity prevents the expression of Ime1, as does the presence of glucose and nitrogen [73,74]. In the lab, the induction of Ime1 typically occurs through nitrogen depletion and the presence of a nonfermentable carbon source such as acetate [71]. The activity of Ime1 initiates the first transcriptional wave of early meiotic genes needed for entry into meiosis, premeiotic S phase, prophase I, and prophase I exit [45,75,76,77,78,79]. One of the genes targeted by Ime1 is IME2, whose production is important for many events throughout meiosis [70,75,76,80].
In meiosis, G1-CDK is not active [74]. Instead, Ime2 performs many functions for the initiation of meiosis. For example, Ime2 is needed for the rapid upregulation of many early meiotic genes and for the timely entry into premeiotic S phase [80,81,82,83,84]. A comparison of RNA expression between wild type and ime2Δ cells shows that the expression of many cell cycle genes is decreased or delayed in ime2Δ cells, especially those with consensus SCB (Swi4 cell cycle box) or MCB (MluI cell cycle box) elements in their promoters [82]. In the mitotic cell cycle, the expression of genes with these promoter elements is normally regulated by G1-CDK. In addition, Ime2 is needed for the timely activation of S-CDK (Cdk1 bound to Clb5, or Clb6) [44,46,47]. In contrast to the mitotic cell cycle in which the CDK inhibitor Sic1 is phosphorylated and targeted for proteasomal degradation by G1-CDK, in meiosis, Ime2 activity results in Sic1 phosphorylation, targeting it for proteasomal degradation [7,44,46,47,74,82,85,86,87,88,89,90]. However, Sic1 is eventually degraded after a long delay in ime2Δ cells, suggesting redundant mechanisms that target Sic1 for degradation in meiosis [82].
Once Sic1 is degraded, S-CDK activity promotes premeiotic DNA replication [44,46,47]. Drug inhibition of CDK or deletion of both S phase cyclins Clb5 and Clb6 resulted in cells that were unable to replicate their DNA, demonstrating that S-CDK is essential for DNA replication. Clb5 is thought to be the more dominant cyclin for S-CDK activity. Cells lacking Clb5 are delayed in DNA replication and arrest in either prophase I or in meiosis I, while cells lacking Clb6 undergo a normal meiosis [44]. How S-CDK regulates premeiotic DNA replication has not been fully characterized, but we assume that it phosphorylates DNA replication proteins, as it does in the mitotic cell cycle. In addition, another important kinase DDK (Dbf4-dependent kinase), which consists of the Cdc7 catalytic subunit and the Dbf4 regulatory subunit is also important for DNA replication in both the mitotic cell cycle and meiosis [91,92,93,94].
2.2. Prophase I
After cells replicate their DNA, they enter an extended prophase I in which chromosomes pair, undergo programmed DNA double strand breaks (DSBs), assemble synaptonemal complex, and repair the DSBs off the homolog for crossover recombination [40]. During this process, chromosomes are undergoing rapid telomere-led movements, in which telomeres first cluster together at the nuclear periphery in a “bouquet” configuration and attach to the cytoplasmic cytoskeleton through a protein complex embedded in the nuclear membrane. In budding yeast, the movements are dependent on the actin cytoskeleton and are thought to resolve heterologous interactions and chromosome entanglements [40,95,96,97,98,99,100,101].
S-CDK helps orchestrate many events throughout prophase I with roles in DSB formation and processing and synaptonemal complex assembly [102,103,104,105]. S-CDK and DDK phosphorylate Mer2, an accessory factor for the Spo11 enzyme that makes the DSBs, and this phosphorylation mediates an interaction between Mer2 and other proteins involved in DSB formation [103,105,106,107,108,109]. The timing of Mer2 phosphorylation is coordinated with replication by the recruitment of DDK to the replisome [108,109]. S-CDK also phosphorylates Sae2, a DSB repair protein needed for the removal of the covalent attachment between Spo11 and the DNA ends to allow further processing of DSBs [110]. In addition, CDK is necessary for normal synaptonemal complex formation and may phosphorylate several synaptonemal complex components [104].
During prophase I, it is imperative that M-CDK is not active. Over-expression of the genes encoding the CLB1 and CLB3 cyclins during prophase I causes premature spindle assembly and premature interactions between kinetochores and microtubules [111]. When these cells undergo anaphase I, sister chromatids separate instead of homologous chromosomes. Interestingly, premature expression of CLB4 results in premature spindle formation, but kinetochores do not engage with microtubules. When cells that prematurely expressed CLB4 are released from the prophase I arrest, they segregate homologous chromosomes. These results suggest that premature kinetochore-microtubule interactions can lead to a disruption in the meiosis I chromosome segregation pattern and that the activity of CDK, especially when bound to Clb1 or Clb3, needs to be tightly regulated.
Multiple mechanisms restrict M-CDK activity in prophase I. First, the APC/C, bound to the meiosis-specific co-activator Ama1 targets M phase regulators and M phase cyclins for proteasomal degradation during prophase I (Table 1) [66]. One of the M phase regulators that is targeted by APC/C-Ama1 is Ndd1, a transcriptional activator for the M phase cyclins in mitosis. By targeting both Ndd1 and the M phase cyclins for degradation, Ama1 prevents premature activity of M-CDK. Second, transcription of the M phase cyclins depends on Ndt80, which induces gene expression at the end of the pachytene stage of prophase I [76,112]. Third, the meiotic recombination checkpoint, which delays cells in prophase I for additional time to repair DSBs, either directly inhibits M-CDK activity or inhibits the transcription of the M phase cyclins [113,114,115,116]. The recombination checkpoint is normally always active in meiosis as cells are repairing DSBs, and the delay can be extended if DSBs are not repaired. DSBs mediate checkpoint signaling by activating either Tel1 (ATM homolog in mammals) or Mec1 (ATR homolog in mammals), depending on the processing status of the lesion [117]. Mec1 and Tel1 activate a meiosis-specific effector kinase Mek1 that helps to inhibit meiotic progression to promote repair [118,119,120,121,122,123]. Mek1 phosphorylates and inhibits the middle meiosis transcription factor Ndt80, which prevents transcription of the M phase cyclins to block entry into the meiotic divisions [114]. The recombination checkpoint is also signaled through the activation of the Swe1 kinase (Wee1 in fission yeast and mammals), which puts an inhibitory phosphorylation on CDK [113].
Transcription of NDT80 is tightly regulated to prevent premature expression of middle meiosis genes. Although Ime1 induces the transcription of NDT80, expression is delayed with respect to other Ime1-induced genes because the Sum1 repressor binds to the NDT80 promoter [112,124,125]. Sum1 is removed from the promoter once phosphorylated by Ime2, CDK, and DDK [126,127,128]. Therefore, Ndt80 is only expressed once Ime2, also induced by Ime1, is active. Ime2 also phosphorylates and activates Ndt80 [47]. The small amount of Ndt80 from Ime1 induction is held inactive until the recombination checkpoint is satisfied [129]. Once DSBs are repaired and the checkpoint is no longer active, Ndt80 can then bind its own promoter to give a high induction of NDT80 transcription [130]. Ndt80 induces the expression of greater than 300 target genes, many of which are involved in the completion of meiotic recombination, the disassembly of the synaptonemal complex, the meiotic divisions, and spore formation, ultimately resulting in prophase I exit [112,129,131].
2.3. The Meiotic Divisions
CDK activity in conjunction with the B-type cyclins Clb1, Clb3, and Clb4 promote progression through the meiotic divisions [47,49,50] (Figure 1). CLB2, which encodes the major mitotic cyclin, is not expressed in meiosis [49,50]. Cells with single deletions of CLB1, CLB3, or CLB4 undergo meiosis somewhat normally, except that loss of CLB1 causes a reduction in four-spored asci and an increase in two-spored asci [50]. Clb1 seems to be the most important cyclin for meiosis, as combined loss clb1Δ clb3Δ and clb1Δ clb4Δ mutants have a much higher frequency of dyads than clb3Δ clb4Δ mutants. The dyads produced from clb1Δ clb3Δ and clb1Δ clb4Δ cells are viable and diploid. Examination of marker segregation suggests that the cells undergo meiosis I but not meiosis II. Overall, the results demonstrate that the M phase cyclins can compensate for one another, but that Clb1 may have a more important role than other M phase cyclins.
Each B-type cyclin has a unique regulation [47,49,50,51,61]. A previous study characterized the regulation of B-type cyclin mRNA, protein, and CDK activity [51]. They find that Clb5 protein and CDK-Clb5 activity both increase at metaphase I and metaphase II, and decline in anaphase I and anaphase II. Clb4 protein accumulates in meiosis I and stays present throughout both meiotic divisions. Although CDK-Clb4 is active in meiosis I, its activity declines in meiosis II, suggesting that post-translational mechanisms regulate CDK-Clb4 activity. Clb1 protein levels increase in meiosis I and remain high until meiosis II exit. However, a different study finds that the levels of Clb1 in the nucleus decrease in anaphase I and then return in metaphase II [62]. In addition, CDK-Clb1 is only active in meiosis I, again suggesting regulation through post-translational mechanisms [51]. Although CLB3 is transcribed by Ndt80 and the RNA is present in meiosis I, CLB3 mRNA is not translated until meiosis II. CLB3 mRNA is not translated because it binds a translational repressor until meiosis II [132]. Forced production of Clb3 in meiosis I results in the loss of the meiosis I chromosome segregation pattern: sister chromatids separated instead of homologous chromosomes [51,111]. These results suggest that Clb1 serves as the major meiosis I cyclin and Clb3 serves as the major meiosis II cyclin. Further work is needed to understand the roles of Clb4 and Clb5 in the meiotic divisions. The finding that meiosis can occur mostly normally without Clb1 or Clb3 suggests that the other cyclins likely have overlapping roles in meiotic progression [50].
The transition between meiosis I and meiosis II requires unique regulation (Figure 2). As cells undergo anaphase I, the APC/C bound to the co-activator Cdc20 becomes active and ubiquitinates securin, targeting it for proteasomal degradation [67]. Securin degradation releases separase, which cleaves cohesin along the chromosome arms for chiasma release, allowing homologous chromosome segregation. The pericentromeric cohesin is protected from cleavage by shugoshin (Sgo1) so that sister chromatids stay together until meiosis II [133,134]. The levels of Clb5 also decline in anaphase I, likely due to targeting by APC/C-Cdc20 [51,135] (Table 1). With the post-translational downregulation of Cdk-Clb1 activity, APC/C-Ama1 becomes active and also targets Clb5 and securin for degradation [65]. Thus, there are two pathways of APC/C-mediated degradation to promote timely degradation of securin and cyclins during anaphase I.
With the degradation or other post-translational regulation of the cyclins, CDK activity decreases and the Cdc14 phosphatase is released from the nucleolus to reverse CDK phosphorylations. Both the low level of CDK activity and the dephosphorylation of substrates is required for meiosis I spindle disassembly [62,63]. However, origin licensing, which normally occurs with low CDK activity, is suppressed after meiosis I to prevent DNA re-replication. Budding yeast likely undertake two mechanisms to prevent DNA re-replication. First, CDK activity is not fully downregulated and Cdc14 nucleolar release is not as extensive in meiosis as in mitosis [62,63,136]. In mitosis, there are two pathways that lead to full Cdc14 release, FEAR (Cdc14 early anaphase release) and MEN (mitotic exit network) [33,137,138,139]. In meiosis, Cdc14 release primarily occurs through FEAR and not MEN, suggesting that the release may not be as prolonged [63,136]. Second, CDK and Ime2 phosphorylate many of the same substrates but at different consensus sequences, including the CDK inhibitor Sic1, and pre-Replication Complex (pre-RC) components Cdc6, Mcm6, and Orc6 [140,141]. The Ime2 phosphorylations are thought to have the same functional consequence as the CDK phosphorylations, except that they are resistant to the Cdc14 phosphatase. The pre-RC substrates do not assemble due to the activity of both CDK and Ime2 [141]. Furthermore, Ime2 was shown to be sufficient to block DNA re-replication [140]. Also, some ime2Δ cells undergo a second round of DNA replication in late meiosis, after a prolonged delay [81,142]. Therefore, by having two kinases that phosphorylate many of the same substrates and some non-overlapping substrates, cells can transition from meiosis I to meiosis II without DNA re-replication.
In meiosis II, a number of mRNAs, including CLB3 mRNA, are translated for normal meiosis II regulation, meiotic exit, and spore formation. The Rim4 translational repressor forms amyloid-like aggregates and binds to the 5′UTR of CLB3 and other mRNAs until meiosis II [132,143]. In meiosis II, Ime2 heavily phosphorylates the intrinsically disordered region of Rim4, leading to some reversal of the amyloid-like properties of Rim4 [144]. The phosphorylation of Rim4 likely leads to its autophagic and/or proteasomal degradation [144,145].
Rim4 is a substrate of autophagy, and with inhibition of autophagy, Rim4 persists and Clb3 protein is not present [145,146]. Interestingly, inhibition of autophagy blocks meiotic exit and the cells continue to undergo rounds of spindle formation, spindle elongation, and chromosome segregation. Rim4 also binds mRNA of the meiosis-specific co-factor of the APC/C, Ama1 [143]. Ama1 substrates are not degraded with inhibition of autophagy, suggesting that Ama1 is not present in high enough levels for APC/C-Ama1 activity [145]. Forced expression of CLB3 or AMA1 in meiosis II without their normal 5′UTR prevents the additional rounds of spindle formation and chromosome segregation. Therefore, autophagy has an important role in the regulation of meiotic exit and in preventing additional rounds of chromosome segregation by degrading Rim4 and allowing the translation of several mRNAs in meiosis II.
One of the key steps for meiotic exit is the inactivation of M-CDK activity [64]. The M phase cyclins are targeted for proteasomal degradation, first by APC/C-Cdc20, whose activation in meiosis II requires the activity of the casein kinase Hrr25. Proteasomal degradation of the cyclins results in decreased CDK activity, which allows APC/C-Ama1 to become active. APC/C-Ama1 targets additional substrates for degradation, such as Ndt80, which prevents further transcription of the genes encoding the B-type cyclins and other M phase regulators. With the downregulation of M-CDK activity and the activation of APC/C-Ama1, the spindle disassembles and Cdc14 is released from the nucleolus for reversal of CDK substrate phosphorylations. Thus, cells exit meiosis and initiate the sporulation program.
3. Overview of S. pombe CDK Activity
Meiosis in fission yeast is also governed by the oscillatory activity of CDK. Fission yeast encode six CDKs and two CDK activating kinases (CAKs) [147]. Only one CDK, Cdc2 (or Cdk1), is essential for mitotic and meiotic cell cycle regulation [148,149]. Although, Pef1 (similar to Pho85 in budding yeast), when bound to cyclin Pas1, promotes cell cycle start [150]. It is not known if Pef1 also has a role in meiosis. There are four cyclins that bind and activate Cdc2 in both the mitotic cell cycle and meiosis [151,152,153,154,155,156,157,158]. In addition, there are two meiosis-specific cyclins, Rem1 and Crs1 [159,160,161] (Figure 1). The cyclins Cig1, Cig2, and Puc1 are considered the G1/S cyclins with roles in entrance into S phase and DNA replication during vegetative growth [151,152,156,157,162]. In meiosis, among the G1/S cyclins, Cig2 has the most important role in DNA replication [163,164]. During prophase I, Cig1, Cig2, Crs1, and Rem1 contribute to programmed DSB formation, assembly of chromosomal architecture elements, or meiotic recombination [159,164,165,166]. The cyclin Cdc13 regulates chromosome segregation during mitosis and meiosis [148,163,167,168].
Full CDK activity requires both cyclin binding and the phosphorylation of the activation domain by CAK. Fission yeast have two CAKs, the Mcs6/Mcs2/Pmh1 complex and Csk1, an ortholog of budding yeast Cak1 [147,169,170,171,172,173,174,175,176]. Although mcs6 is an essential gene and csk1 is not essential, the Mcs6 complex and Csk1 are thought to have redundant roles in phosphorylating and activating Cdc2 [174,175]. In addition, Csk1 also phosphorylates and activates Mcs6, acting as a CAK activating kinase [177]. Although it is assumed that CAK activation of Cdc2 is also required for meiosis, roles of the Mcs6 complex and Csk1 have not been analyzed in meiosis. However, a recent study identified csk1 in a genome-scale screen for genes important for meiosis [178]. Further work is needed to determine the role of Csk1 in meiotic regulation and spore formation, especially given the similarity of Csk1 to Cak1, which has additional roles in meiosis in budding yeast [59,176].
Oscillations in CDK activity occur through several regulatory mechanisms in meiosis. First, the cyclin-dependent kinase inhibitor Rum1 is needed for cells to arrest in G1 for mating and meiotic initiation, but Rum1 must be degraded prior to premeiotic S phase [179]. Second, M-CDK activity is regulated by the Wee1 kinase and the Cdc25 phosphatase during S phase and prophase I [163,168,180]. Wee1 phosphorylates tyrosine-15 on Cdc2 to decrease Cdc2-Cdc13 activity prior to entrance into meiosis I [180]. Cdc25 removes the inhibitory phosphorylation for a burst of CDK activity as cells enter meiosis I [148,163,168]. Third, many of the cyclins are regulated by meiosis-specific transcriptional and post-transcriptional mechanisms to allow an increase in cyclin levels at specific stages of meiosis [148,160,163,166,181,182,183]. Fourth, the cyclins are ubiquitinated and targeted for proteasomal degradation in anaphase I and anaphase II by the APC/C ubiquitin ligase. In meiosis, the APC/C mainly uses two co-activators, Slp1 (Cdc20 in budding yeast) and a meiosis-specific co-activator Mfr1/Fzr1 [184,185,186,187] (Table 1).
3.1. Entry into Meiosis and Premeiotic DNA Replication
Fission yeast typically live as haploids, but cells of opposite mating type conjugate when starved of nitrogen [188,189]. The diploid zygote then undergoes meiosis and spore formation. In the lab, this method of sporulation is not very synchronous due to the conjugation step [39,190]. Often, a method is used, in which a diploid is induced to initiate meiosis through the downregulation of Pat1 activity, an inhibitor of meiosis (see below); however, some differences from zygotic meiosis can be observed [39].
Meiosis is initiated through the induction of successive waves of transcription. The expression of the transcripts at the mating loci induce the expression of mei3, which encodes an inhibitor of Pat1 protein kinase [191,192]. In vegetative cells, Pat1 blocks meiosis by phosphorylating the substrates Ste11, a transcriptional activator, and Mei2, an RRM-type RNA-binding protein under Ste11-transcriptional control [193,194]. Because Mei3 inhibits Pat1 in meiosis, Pat1 cannot promote Ste11 nuclear exclusion or target Mei2 for degradation. Thus, Ste11 induces the transcription of early meiotic genes, and levels of Mei2 accumulate [194,195]. Mei2 binds Mmi1 to inhibit DSR-Mmi1 (Determinant of Selective Removal-Mmi1), an RNA degradation system that selectively removes sporulation-specific transcripts [196,197]. Mei2 prevents Mmi1 from binding elements within the 3’UTR of meiotic transcripts and recruits the exosome to degrade them [196,198,199]. Therefore, the activation of both Ste11 and Mei2 allows hundreds of genes to be transcribed and meiotic mRNAs to be translated.
Two mRNAs regulated by Mei2 include crs1, which encodes a meiosis-specific cyclin and mei4, which encodes a transcription factor important for the expression of a large number of middle meiosis genes [160,163,181,200,201,202]. The transcription of both crs1 and mei4 occurs in vegetative and meiotic cells, but the RNA accumulates only upon Mmi1 inhibition in meiosis [181,196,202]. In addition, csr1, like many other meiotic transcripts in fission yeast, undergoes meiosis-induced splicing to restrict expression to meiosis [160,203]. Mei4 regulates expression of the genes that encode the cyclins rem1, and cig2, and the CDK regulators wee1 and cdc25 [159,163,166,204]. Interestingly, Mei4 also regulates the splicing of rem1, likely by recruiting components of the spliceosome [159,166]. Therefore, Mei2′s role in blocking RNA degradation of crs1 and mei4, along with other transcripts, promotes many events throughout early meiosis and the meiotic divisions.
CDK activity is required for DNA replication in premeiotic S phase, likely due to its role in phosphorylating key proteins that regulate replication, as it does in the mitotic cell cycle [148,205]. cig2 is transcribed just before premeiotic DNA replication, and the protein is present during S phase and then declines shortly afterwards [163]. In contrast, cig1 is transcribed, and the protein present, just after premeiotic S phase and is not thought to have an important role in DNA replication [163,184]. The Puc1 cyclin also does not have a predominant role in premeiotic DNA replication [164]. Therefore, Cig2 bound to Cdc2 has a central role in DNA replication (Figure 1). However, cells with a deletion of cig2 or deletions of both cig1 and cig2 only have a 30-minute delay in DNA replication. A triple deletion strain, cig1Δ cig2Δ puc1Δ, displays only minor additive roles in the timing of onset of premeiotic DNA replication. Therefore, the G1/S cyclins are dispensable for premeiotic DNA replication but make the process more efficient.
The meiosis-specific cyclin Rem1 compensates for the loss of Cig2 to allow premeiotic DNA replication after a short delay [159]. Loss of both cig2 and rem1 results in cells that are blocked before premeiotic DNA replication. Normally, Rem1 is not present during premeiotic S phase; rem1 is expressed prior to premeiotic DNA replication but the transcript is not spliced until just before the onset of meiosis I. However, rem1 mRNA is spliced several hours earlier in cig2Δ cells, suggesting that CDK-Cig2 activity blocks rem1 splicing.
The M-phase cyclin Cdc13, and its associated kinase activity, is also present during premeiotic S phase and increases to high levels through meiosis II [163,168]. However, Cdc13 is not required for premeiotic S phase under normal circumstances and cannot compensate for the loss of all other cyclins in premeiotic S phase. Surprisingly, expression of a Cdc13-Cdc2 fusion protein is sufficient for DNA replication in the absence of all other cyclins [164]. These results suggest that Cdc2-Cdc13 can indeed support premeiotic DNA replication but is prevented through an unknown mechanism from activating CDK during normal premeiotic S phase.
After cells complete S phase, the activity of CDK is reduced [163]. Cdc2 is phosphorylated at tyrosine-15, an inhibitory phosphorylation put on by Wee1 kinase [20,21,22,163,180,206,207]. Due to the inhibitory phosphorylation, the activity of Cdc2 bound to M phase cyclin Cdc13 is low, but not absent, until entry into meiosis I when the inhibitory phosphorylation is removed [163].
3.2. Prophase I
During prophase I, fission yeast undergo characteristic nuclear movements to help facilitate homologous chromosome pairing and recombination. Starting in premeiotic S phase, telomeres cluster together at the nuclear membrane, just underneath the SPB, and form a structure known as the telomere bouquet [208,209,210,211,212,213]. The bouquet is connected to the SPB. Although the telomere bouquet is a conserved structure found in many organisms, fission yeast have an especially remarkable bouquet stage [40,214]. Following bouquet formation, the nucleus oscillates due to microtubule and dynein-dependent drastic movements, led by the SPB inserted in the nuclear membrane [190]. This period is defined as the ‘horsetail stage’. The telomere bouquet and horsetail movements are important for a number of events including promotion of homolog interactions, disruption of ectopic interactions, meiotic recombination, spindle formation, and centromere assembly [215,216,217,218,219,220,221].
CDK activity regulates multiple events in prophase I, including DSB formation, recombination, and maturation of linear elements, which are protein complexes structurally related to elements within the synaptonemal complex in other eukaryotic organisms [165,222,223,224,225]. Along with Cig1 and Cig2, the meiosis-specific Crs1 cyclin is present in prophase I. crs1 has four introns that are spliced starting in premeiotic S phase, and the spliced messages increased by prophase I [160]. Rem1 also has a role in recombination, such that levels of gene conversion, but not crossovers, are reduced in its absence [159]. Interestingly, Rem1 is not spliced until several hours after crs1 [159,160,166]. In prophase I, the unspliced version of rem1 is translated into a truncated protein that lacks a cyclin box. The shorter Rem1 is needed for normal levels of gene conversion, suggesting that Rem1’s role in recombination does not depend on binding Cdc2 [166]. Therefore, Rem1 has CDK-independent functions in prophase I, and Cig1, Cig2, and Crs1 would be the cyclins that function with Cdc2 to regulate events in prophase I.
As in budding yeast, DSBs are induced by the Spo11 homolog Rec12 bound to accessory proteins [226,227,228]. Inhibition of an analog-sensitive allele of Cdc2 (Cdc2-asM17) resulted in a great decrease in DSB formation [165]. Deletions of the cyclin genes cig1, cig2, and crs1 individually cause a reduction in DSB formation at a strong natural hotspot, with the cig1Δ crs1Δ double mutant displaying the greatest reduction. Linear elements are required for normal double strand break formation at hotspots [161,222,224,229]. cig1Δ crs1Δ double mutants accumulate linear element protein Rec25 with similar timing as control cells but few cells display mature linear elements [165]. Similar results are found in cdc2-asM17 cells, even without addition of the analog, and the defects are enhanced upon analog addition. These results suggest that CDK activity is needed for linear element maturation and normal DSB formation.
Cig1, Cig2, and Crs1 also contribute to recombination. Deletion of cig1 or cig2 caused a reduction in gene conversion, but not a reduction in crossovers [165]. The cig1Δ cig2Δ double mutant did not have an additive effect. Loss of crs1 resulted in a decrease in both gene conversion and crossovers. The cig1Δ crs1Δ double mutant had a similar defect as the crs1Δ single mutant. In another study, loss of crs1 shows no defect in recombination [164]. It is currently unclear why the two studies gave conflicting results. However, the summary of results suggests that CDK activity, with Cig1, Cig2, and possibly Crs1, regulates DSB formation, linear element maturation, and recombination, but the substrates are currently not known. Since these events are linked, with lower levels of mature linear elements affecting DSB formation and recombination, a small number of substrates may regulate all three events. [165,222,223,224,230,231,232,233,234]. Future studies are needed to identify the substrates that regulate these events.
The completion of recombination is monitored by the recombination checkpoint, which delays prophase I progression by downregulating CDK activity in the presence of unrepaired DNA breaks. DSBs are sensed by Rad3 (ATR) and Tel1 (ATM), which phosphorylate and activate the downstream meiosis-specific effector kinase Mek1 [235,236,237]. Mek1 excludes Cdc25 from the nucleus to prevent the removal of the inhibitory phosphorylation of Cdc2, resulting in an inhibition of CDK activity [238]. Chk1 acts downstream of Rad3 and Tel1 in the mitotic DNA damage checkpoint to phosphorylate both Cdc25 and Wee1 [239,240]. However, although Chk1 is predicted to play a role in the recombination checkpoint, Chk1 is not active until meiosis II [236]. Activation of the recombination checkpoint, by mutating proteins involved in DNA repair, results in an extended meiotic prophase beyond the completion of horsetail movements [241].
3.3. The Meiotic Divisions
The normal transition from prophase I to the entrance into the first meiotic division is also controlled by Wee1 and Cdc25 regulation of CDK. Cells enter prophase I with low CDK activity due to Wee1 inhibitory phosphorylation on Cdc2 [163,168,180]. As cells progress through prophase I, and DNA breaks are repaired, the levels of phosphorylated Cdc2 decreases due to the activity of the Cdc25 phosphatase, with a corresponding increase in CDK activity [163,168]. The Mei4 middle meiosis transcription factor induces cdc25 and inhibits wee1 by binding a region upstream of the wee1 open reading frame [148,204,242]. This transcriptional regulation results in an increase in Cdc25 and a decrease in Wee1 as cells progress through prophase I, allowing a rapid induction of CDK activity and the entrance into nuclear divisions.
The M phase cyclin Cdc13 is essential for the meiotic divisions. In the absence of Cdc13, cells undergo premeiotic DNA replication, but fail to undergo the meiotic divisions [148,164,205]. Some of the cells even re-replicate their DNA [164]. Cdc13 protein is present in S phase and then increases in the meiotic divisions [163,168]. The increase in protein levels is due to meiosis-specific transcriptional and post-transcriptional regulation of cdc13. Instead of producing a single 2.5kb transcript, as found in mitosis, both a 2.5kb and a 2.2kb cdc13 transcript is produced in meiosis, with a different length in the 3’UTR [148,163]. cdc13 transcription initiates in premeiotic S phase and rises to a higher level in meiosis I and meiosis II [163]. In the absence of the transcriptional activator Mei4, the smaller transcript did not rise to high levels in meiosis I and meiosis II, as it does in wildtype cells [148,163]. The transcript is also regulated by the Spo5 RNA-binding protein [182]. Spo5 protein is present in prophase I and then disappears as cells enter meiosis II, similar to the timings when Cdc13 levels increase [182,183]. In the absence of spo5, cells had lower levels of Cdc13 in meiosis II and did not complete meiosis II, suggesting that Spo5 has an important role in allowing the proper timing of the production of Cdc13. The meiosis II defect is partially suppressed by overexpression of Cdc13, suggesting that lower levels of Cdc2-Cdc13 activity prevents spo5Δ cells from finishing meiosis II [182,183]. The transcriptional and post-transcriptional regulation of cdc13 is thought to allow an increase in Cdc13 protein in meiosis II, after degradation of Cdc13 in anaphase I, such that adequate Cdc13 protein is present for meiosis II.
With endogenous expression, Cdc13 is not sufficient for the production of viable gametes. Cells with deletions of all other cyclins besides Cdc13 (cig1Δ cig2Δ puc1Δ rem1Δ crs1Δ) form mostly empty or one-spored asci [164]. However, if Cdc13 is fused to Cdc2, cells lacking the other cyclins undergo meiosis I, but arrest before meiosis II. A higher level of expression of Cdc2 fused to Cdc13 allows cells to go through both meiotic divisions, however, most of the spores are inviable. These results suggest that Cdc13 is sufficient for progression through both divisions when tethered to Cdc2 and expressed at a high level, but other cyclins are needed for spore viability, likely due to their important roles in recombination and other meiotic events.
Cig2, Rem1, and Crs1 have more minor roles in the meiotic divisions. Although the main role of Cig2 is in premeiotic DNA replication, Cdc2-Cig2 has another peak of expression (Mei4-dependent) and activity at the end of meiosis I [163] (Figure 1). Most cig2Δ cells undergo meiosis normally, but some have a delay in meiosis II and form dyads, with two diploid spores [163,164]. The percentage of dyads increases in cig1Δcig2Δ double mutants and cig1Δ cig2Δ puc1Δ triple mutants [160,164]. Rem1 and Crs1 are only important for zygotic meiosis, in which haploid cells mate before undergoing meiosis. Some rem1Δ crs1Δ cells underwent abnormal meiotic nuclear divisions due to a multipolar spindle. In conclusion, Cig1, Cig2, and Puc1 have some additional roles in progression through the meiotic divisions.
As cells exit metaphase I, the APC/C becomes active to ubiquitinate securin and B-type cyclins, targeting them for proteasomal degradation. The degradation of securin releases separase, which cleaves cohesin and allows the separation of homologous chromosomes [243]. Cdc13 is also degraded, but a pool of Cdc13 remains for low CDK activity during the meiosis I-to-meiosis II transition (Figure 2) [244]. The decrease in some but not all CDK activity is important; CDK activity must be low enough to allow spindle disassembly and meiosis I exit, but high enough to prevent origin licensing and the re-replication of DNA. In fission yeast, an inhibitor of the APC/C, Mes1, prevents full Cdc13 degradation [244,245,246]. Mes1 is a pseudo-substrate of the APC/C co-activator Fzr1/Mfr1 and a competitive substrate of co-activator Slp1/Cdc20. As a pseudo-substrate, Mes1 binds and inactivates Fzr1 to prevent the premature activity of APC/C-Fzr1 in meiosis I. As a competitive substrate, Mes1 competes with Cdc13 for binding of APC/C-Slp1. Once bound, Mes1 is eventually ubiquitinated and released. By having Mes1 as a substrate, there is sufficient degradation of securin to facilitate chromosome segregation, but only partial degradation of Cdc13, thus reducing but not eliminating Cdc2-Cdc13 activity.
There is a precise balance in maintaining the correct levels of Cdc13 between the meiotic divisions (Figure 2). In mes1Δ mutants, Cdc13 is completely degraded and cells cannot progress into meiosis II [187,244,245,246]. A loss of fzr1 allows mes1Δ cells to undergo meiosis II, demonstrating that reducing APC/C activity is important for the transition to meiosis II [186,187,246]. However, too much of a reduction of APC/C activity can also be detrimental. A mutant of slp1 (slp1-B05) that results in reduced APC/C-Slp1 activity, does not affect vegetative growth; but, in meiosis, metaphase I is substantially delayed and slp1-B05 cells ultimately undergo only one meiotic division [186]. Deletion of both mes1 and fzr1 can rescue the second meiotic division. These results demonstrate the importance of Mes1 in allowing a decrease but not an elimination of Cdc2-Cdc13 activity.
As cells enter meiosis II, APC/C-Slp1 and APC/C-Fzr1 activity declines and Mes1 is targeted for proteasomal degradation, possibly through other APC/C co-activators Fzr2 and Ste9, promoting full CDK activity and meiosis II spindle assembly [246]. In addition, the Cuf2 transcription factor induces expression of fzr1 to increase Fzr1 levels in meiosis II [186]. The boost of APC/C-Fzr1 activity is required for normal meiotic exit and spore formation [184,185,246].
In anaphase II, both APC/C-Slp1 and APC/C-Fzr1 target securin and B-type cyclins for degradation [186]. Although fzr1Δ cells undergo meiosis I and meiosis II normally, they do not exit meiosis. Instead, they assemble abnormal spindles and undergo an additional round of chromosome segregation [186]. They also maintain levels of Cdc13 after meiosis II, unlike wildtype cells that degrade Cdc13 after meiosis II [184,186]. In mutants of cuf2, the transcription of fzr1 is decreased by approximately 50% during the meiosis I-to-meiosis II transition [186]. Approximately 20% of cuf2Δ asci display additional spores, also due to an additional round of chromosome segregation after meiosis II. Cdc13 degradation is also aberrant in cuf2Δ cells. These results suggest that degradation of Cdc13 by APC/C-Fzr1 activity is important for the termination of meiosis after meiosis II.
4. Comparisons of CDK Activity in Budding and Fission Yeast Meiosis
CDK has many important roles in regulating both budding and fission yeast meiosis. Although many of the essential activities of CDK are similar between the two organisms, there are also interesting examples where regulation is quite different. These differences demonstrate how regulatory mechanisms have evolved between the two species but accomplish the same outcome. We highlight some of the similarities and differences in this section.
In both budding and fission yeast, meiosis initiates in diploid cells heterozygous for the mating loci as a response to starvation and is coupled to spore formation [38,69,70,71,72,73,188,190]. The starvation response signals the induction of transcriptional cascades that result in meiotic initiation [45,112,194,195]. As cells enter meiosis, CDK activity is low and inhibited by CDK inhibitors Sic1 and Rum1 in budding and fission yeast, respectively [44,179]. Budding yeast has a meiosis-specific CDK-like kinase called Ime2 that functions at multiple stages in meiosis, including entrance into premeiotic S phase and in meiosis II [44,45,47,48,144]. Ime2 targets Sic1, the CDK inhibitor, for degradation to allow activation of S-CDK [47]. In fission yeast, the CDK inhibitor Rum1 must also be degraded prior to premeiotic S phase, but it is currently unclear how Rum1 is targeted for degradation in meiosis [179]. Once active, S-CDK has an essential role in DNA replication in both yeasts, likely by phosphorylating the same substrates involved in mitotic DNA replication.
In an extended prophase I, many meiosis-specific events occur, including DSB formation, homologous chromosome pairing, and recombination [40]. During this time, telomeres cluster into a bouquet structure and then make connections with proteins that pass through the nuclear envelope, linking telomeres to the cytoskeleton and facilitating rapid chromosome movements. Such telomere-led movements are thought to help facilitate homologous chromosome interactions, possibly by disrupting ectopic interactions and removing interlocks. Currently, it is unknown whether CDK regulates chromosome movements, but it is intriguing to speculate a role for CDK, given that other events in prophase I are known to require CDK. In mammals, CDK2 is involved in telomere attachment to the nuclear envelope during meiosis [247,248,249,250]. In budding yeast, CDK phosphorylates the Spo11 accessory protein Mer2 for formation of programmed DSBs, and CDK regulates Sae2 for break repair [103,105,110]. In fission yeast, CDK is also involved in DSB formation, but the substrates are unknown [165]. Cdc2 inhibition results in defective linear elements, possibly suggesting that CDK regulates DSB formation by regulating linear element assembly, which is required for normal levels of DSBs [222]. Future work is needed to determine whether there are multiple CDK substrates for DSB formation, linear element assembly, and recombination in fission yeast.
In both budding and fission yeast, a checkpoint in prophase I delays the cell cycle in the presence of unrepaired DSBs. One strategy for this checkpoint is to downregulate CDK activity by activating the Swe1/Wee1 kinase which places an inhibitory phosphorylation on CDK to block its activity. In budding yeast, Swe1 activation can result in checkpoint signaling, but is not thought to be a main mechanism of checkpoint regulation [113,117]. Instead, the checkpoint mainly functions to inhibit the Ndt80 transcription factor to prevent transcription of the M phase cyclins, thus preventing M-CDK activity [114,115,116]. In fission yeast, CDK inhibition is the main strategy for engaging the checkpoint and delaying cells in prophase I. Activation of the checkpoint prevents nuclear accumulation of the Cdc25 phosphatase, preventing CDK activation [237,238]. The difference in regulation between the two yeasts could be due to the timing of cyclin expression. In budding yeast, the M phase cyclins Clb1, Clb3, and Clb4 are not transcribed until cells are exiting prophase I, so inhibition of Ndt80, which is required for transcription of the cyclins and other meiotic regulators, keeps cells in prophase I [112]. Ndt80 also induces transcription of Polo kinase (Cdc5), which leads to disassembly of the synaptonemal complex [112,131]. Since the synaptonemal complex is needed for normal levels of homologous recombination, a checkpoint arrest through Ndt80 inhibition prevents premature synaptonemal complex disassembly, thus ensuring cells complete DSB repair through homologous recombination [118,131,251]. In fission yeast, the M phase cyclin Cdc13 is present in prophase I, and Wee1 inhibits Cdc2-Cdc13 activity throughout a normal prophase I [163,168]. Checkpoint activation prolongs CDK inhibition and extends the time in prophase I to allow DSB repair [168].
For budding and fission yeast cells to enter meiosis I, M-CDK activity increases to promote spindle assembly and chromosome attachment to spindle microtubules. In budding yeast, the M phase cyclins Clb1 and Clb3 are the main cyclins needed for meiosis I and meiosis II, respectively [51]. In fission yeast, Cdc13 regulates both meiotic divisions, but the levels increase in meiosis II [148,164,182,205,252]. In both yeasts, the cyclins that promote premeiotic DNA replication, Clb5 and Cig2 are also activated in the meiotic divisions [51,163]. However, the function of Clb5 in the meiotic divisions has not been delineated. Loss of cig2 does affect meiosis II timing and spore formation [163,164]. Further experiments are needed to identify the specific roles for these cyclins in the meiotic divisions.
In anaphase I, CDK activity must be downregulated to allow spindle disassembly and meiosis I exit. In both budding and fission yeast, APC/C-Cdc20/Slp1 becomes active to target both securin and the cyclins for degradation at the metaphase I-to-anaphase I transition [67,187]. In budding yeast, some of the cyclins are post-translationally regulated for inactivation [51]. After inactivation of Cdk1-Clb1, which inhibits Ama1, APC/C-Ama1 becomes active and targets securin and cyclins for degradation [65]. The phosphatase Cdc14 is released from the nucleolus to dephosphorylate CDK substrates. Both the decline in CDK activity and the release of the Cdc14 phosphatase is needed for spindle disassembly [62,63]. In fission yeast, APC/C-Slp1 targets securin and Cdc13 for degradation at the metaphase I-to-anaphase I transition [187,246]. The Cdc14-like phosphatase Clp1 is not needed for normal meiotic timings or spindle disassembly, but may help with the fidelity of the process [35].
During the meiosis I-to-meiosis II transition, CDK activity must decrease for spindle disassembly, but some remaining CDK activity prevents re-licensing of origins. Budding and fission yeast have two different strategies to prevent complete loss of CDK activity, and consequentially, prevent full dephosphorylation of CDK substrates (Figure 2). In budding yeast, the Ime2 kinase remains active and phosphorylates some of the same substrates as CDK, including a component of the MCM helicase [140,141]. The Ime2 phosphorylations are not removed by the Cdc14 phosphatase, rendering Ime2 substrates resistant to Cdc14 dephosphorylation between meiosis I and meiosis II [140]. In addition, Cdc14 is only quickly released in anaphase I, unlike in mitosis, during which it is first quickly and then fully released [33,62,63,136]. Therefore, some substrates will remain phosphorylated between the meiotic divisions. Fission yeast retain M-CDK activity through the partial inhibition of the APC/C. The APC/C inhibitor Mes1 blocks full APC/C activity [244,245,246]. Mes1 binds and inactivates Fzr1 to prevent APC/C-Fzr1 activity by serving as a pseudo-substrate [246]. Mes1 competes as a substrate of APC/C-Slp1 to decrease the ubiquitination of Cdc13. Therefore, the levels of Cdc13 will decrease but will not be fully eliminated to allow some Cdc2-Cdc13 activity during the meiosis I-to-meiosis II transition.
In both budding and fission yeast, the increase of cyclins in meiosis II is regulated by RNA binding proteins [51,132,143,182]. In budding yeast, the Rim4 amyloid-like translational repressor binds CLB3 mRNA and is targeted for autophagic degradation in meiosis II, releasing CLB3 mRNA for translation, ultimately resulting in an increase in Clb3 protein and Cdk1-Clb3 activity [145]. In fission yeast, the Spo5 RNA binding protein protects and maintains cdc13 mRNA until meiosis II [182]. Cdc13 levels rise through de novo synthesis at the entrance into meiosis II. The current model is that Spo5 protects cdc13 mRNA and promotes its translation in meiosis II. The increase in CDK activity allows meiosis II spindle formation and chromosome attachment.
As cells undergo anaphase II the cyclins are targeted for degradation. In budding yeast, APC/C-Cdc20 and APC/C-Ama1 target the cyclins and other meiotic regulators for degradation, allowing meiotic exit [64]. The Cdc14 phosphatase is released from the nucleolus and removes phosphorylation from CDK substrates [62,63,253]. In fission yeast, APC/C-Slp1 and APC/C-Fzr1 target cyclins for degradation [184,185,186,187,246]. Clp1 likely reverses the phosphorylation of CDK substrates.
Interestingly, both the meiosis-specific APC/C-Ama1 and APC/C-Fzr1 have key roles in ensuring two and only two meiotic divisions. In budding yeast, if autophagy is inhibited and Rim4 is not targeted for degradation, cells fail to exit meiosis and instead undergo additional rounds of spindle assembly, spindle elongation, and chromosome segregation [145]. AMA1 mRNA is also a target of Rim4, and inhibition of degradation of Rim4 likely prevents an increase in Ama1 levels and APC/C-Ama1 activity [132,143]. Induced expression of AMA1 in meiosis II suppresses the extra divisions and allows cells to exit meiosis after meiosis II [145]. Thus, APC/C-Ama1-mediated degradation of M phase proteins plays a critical role in meiotic exit. In fission yeast, deletion of fzr1 prevents cells from exiting meiosis II and results in an additional round of chromosome segregation [186]. The results suggest that a failure to degrade key substrates at the end of meiosis II prevents meiotic exit. Additionally, the transcription factor Cuf2 increases expression of fzr1 during meiosis II. Loss of Cuf2 results in an additional round of chromosome segregation, although the defect is not as severe as loss of Fzr1. Therefore, mechanisms in both budding and fission yeast ensure the production of specific co-activators of the APC/C to limit the number of rounds of chromosome segregation by promoting meiotic exit after meiosis II.
5. Future Directions
The investigation of CDK regulation in meiosis in budding and fission yeast has greatly improved our understanding of how key meiotic events are orchestrated. Furthermore, research on CDK activity in meiosis has uncovered additional layers of regulation not found in the mitotic cell cycle. Nevertheless, further work is needed to identify a comprehensive list of meiotic Cdk1 substrates and to determine how phosphorylation regulates protein function. While we assume that for some events in meiosis, Cdk1 phosphorylates similar substrates as in the mitotic cell cycle, further investigation of meiosis-specific events is warranted. An exciting future direction would be to determine how CDK coordinates multiple events in prophase I, such as rapid chromosome movement, DSB formation, synaptonemal complex assembly, and recombination. Although some substrates have been identified, a more extensive analysis of substrates should uncover additional meiotic regulators. Similarly, further study of how CDK regulates meiosis II and meiotic exit could uncover novel regulatory mechanisms that limit meiosis to two and only two meiotic divisions. Because cell cycle regulators are highly conserved, work from budding and fission yeast has proved to be foundational for our understanding of meiotic regulation in all eukaryotes.
Author Contributions
A.M.M. and S.L. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the NIH to S.L. (GM105755).
Acknowledgments
We thank the members of the Lacefield lab for their critical reading of the manuscript.
Conflicts of Interest
The authors have no conflicts to report.
References
- Murray, A.W. Recycling the cell cycle: Cyclins revisited. Cell 2004, 116, 221–234. [Google Scholar] [CrossRef] [Green Version]
- Morgan, D.O. Cyclin-dependent kinases: Engines, clocks, and microprocessors. Annu. Rev. Cell Dev. Biol. 1997, 13, 261–291. [Google Scholar] [CrossRef]
- Hartwell, L.H.; Culotti, J.; Pringle, J.R.; Reid, B.J. Genetic control of the cell division cycle in yeast. Science 1974, 183, 46–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartwell, L.H.; Mortimer, R.K.; Culotti, J.; Culotti, M. Genetic Control of the Cell Division Cycle in Yeast: V. Genetic Analysis of cdc Mutants. Genetics 1973, 74, 267–286. [Google Scholar] [PubMed]
- Nurse, P.; Thuriaux, P.; Nasmyth, K. Genetic control of the cell division cycle in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 1976, 146, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Moreno, S.; Hayles, J.; Nurse, P. Regulation of the cell cycle timing of mitosis. J. Cell Sci. 1989, 12, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Schwob, E.; Bohm, T.; Mendenhall, M.D.; Nasmyth, K. The B-type cyclin kinase inhibitor p40SIC1 controls the G1 to S transition in S. cerevisiae. Cell 1994, 79, 233–244. [Google Scholar] [CrossRef]
- Alberghina, L.; Rossi, R.L.; Querin, L.; Wanke, V.; Vanoni, M. A cell sizer network involving Cln3 and Far1 controls entrance into S phase in the mitotic cycle of budding yeast. J. Cell Biol. 2004, 167, 433–443. [Google Scholar] [CrossRef]
- Coudreuse, D.; Nurse, P. Driving the cell cycle with a minimal CDK control network. Nature 2010, 468, 1074–1079. [Google Scholar] [CrossRef]
- Nash, P.; Tang, X.; Orlicky, S.; Chen, Q.; Gertler, F.B.; Mendenhall, M.D.; Sicheri, F.; Pawson, T.; Tyers, M. Multisite phosphorylation of a CDK inhibitor sets a threshold for the onset of DNA replication. Nature 2001, 414, 514–521. [Google Scholar] [CrossRef]
- Verma, R.; Annan, R.S.; Huddleston, M.J.; Carr, S.A.; Reynard, G.; Deshaies, R.J. Phosphorylation of Sic1p by G1 Cdk required for its degradation and entry into S phase. Science 1997, 278, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Zachariae, W.; Shevchenko, A.; Andrews, P.D.; Ciosk, R.; Galova, M.; Stark, M.J.; Mann, M.; Nasmyth, K. Mass spectrometric analysis of the anaphase-promoting complex from yeast: Identification of a subunit related to cullins. Science 1998, 279, 1216–1219. [Google Scholar] [CrossRef] [PubMed]
- Jaspersen, S.L.; Charles, J.F.; Morgan, D.O. Inhibitory phosphorylation of the APC regulator Hct1 is controlled by the kinase Cdc28 and the phosphatase Cdc14. Curr. Biol. 1999, 9, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Okayama, H.; Nurse, P. Fission yeast Fizzy-related protein srw1p is a G(1)-specific promoter of mitotic cyclin B degradation. EMBO J. 2000, 19, 3968–3977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.A.; Sanchez-Diaz, A.; de Prada, J.M.; Moreno, S. APC(ste9/srw1) promotes degradation of mitotic cyclins in G(1) and is inhibited by cdc2 phosphorylation. EMBO J. 2000, 19, 3945–3955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, S.P.; Labib, K. Chromosome Duplication in Saccharomyces cerevisiae. Genetics 2016, 203, 1027–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labib, K. How do Cdc7 and cyclin-dependent kinases trigger the initiation of chromosome replication in eukaryotic cells? Genes Dev. 2010, 24, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Spellman, P.T.; Sherlock, G.; Zhang, M.Q.; Iyer, V.R.; Anders, K.; Eisen, M.B.; Brown, P.O.; Botstein, D.; Futcher, B. Comprehensive identification of cell cycle-regulated genes of the yeast Saccharomyces cerevisiae by microarray hybridization. Mol. Biol. Cell 1998, 9, 3273–3297. [Google Scholar] [CrossRef]
- Peng, X.; Karuturi, R.K.; Miller, L.D.; Lin, K.; Jia, Y.; Kondu, P.; Wang, L.; Wong, L.S.; Liu, E.T.; Balasubramanian, M.K.; et al. Identification of cell cycle-regulated genes in fission yeast. Mol. Biol. Cell 2005, 16, 1026–1042. [Google Scholar] [CrossRef] [Green Version]
- Lundgren, K.; Walworth, N.; Booher, R.; Dembski, M.; Kirschner, M.; Beach, D. mik1 and wee1 cooperate in the inhibitory tyrosine phosphorylation of cdc2. Cell 1991, 64, 1111–1122. [Google Scholar] [CrossRef]
- Russell, P.; Nurse, P. Negative regulation of mitosis by wee1+, a gene encoding a protein kinase homolog. Cell 1987, 49, 559–567. [Google Scholar] [CrossRef]
- Gould, K.L.; Nurse, P. Tyrosine phosphorylation of the fission yeast cdc2+ protein kinase regulates entry into mitosis. Nature 1989, 342, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Russell, P.; Nurse, P. cdc25+ functions as an inducer in the mitotic control of fission yeast. Cell 1986, 45, 145–153. [Google Scholar] [CrossRef]
- Moreno, S.; Nurse, P.; Russell, P. Regulation of mitosis by cyclic accumulation of p80cdc25 mitotic inducer in fission yeast. Nature 1990, 344, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Gould, K.L.; Moreno, S.; Tonks, N.K.; Nurse, P. Complementation of the mitotic activator, p80cdc25, by a human protein-tyrosine phosphatase. Science 1990, 250, 1573–1576. [Google Scholar] [CrossRef]
- Booher, R.N.; Deshaies, R.J.; Kirschner, M.W. Properties of Saccharomyces cerevisiae wee1 and its differential regulation of p34CDC28 in response to G1 and G2 cyclins. EMBO J. 1993, 12, 3417–3426. [Google Scholar] [CrossRef]
- Russell, P.; Moreno, S.; Reed, S.I. Conservation of mitotic controls in fission and budding yeasts. Cell 1989, 57, 295–303. [Google Scholar] [CrossRef]
- Enserink, J.M.; Kolodner, R.D. An overview of Cdk1-controlled targets and processes. Cell Div. 2010, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Sherwin, D.; Wang, Y. The Opposing Functions of Protein Kinases and Phosphatases in Chromosome Bipolar Attachment. Int. J. Mol. Sci. 2019, 20, 6182. [Google Scholar] [CrossRef] [Green Version]
- Nigg, E.A. Mitotic kinases as regulators of cell division and its checkpoints. Nat. Rev. Mol. Cell Biol. 2001, 2, 21–32. [Google Scholar] [CrossRef]
- Rudner, A.D.; Murray, A.W. Phosphorylation by Cdc28 activates the Cdc20-dependent activity of the anaphase-promoting complex. J. Cell Biol. 2000, 149, 1377–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rudner, A.D.; Hardwick, K.G.; Murray, A.W. Cdc28 activates exit from mitosis in budding yeast. J. Cell Biol. 2000, 149, 1361–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stegmeier, F.; Amon, A. Closing mitosis: The functions of the Cdc14 phosphatase and its regulation. Annu. Rev. Genet. 2004, 38, 203–232. [Google Scholar] [CrossRef] [PubMed]
- Trautmann, S.; Wolfe, B.A.; Jorgensen, P.; Tyers, M.; Gould, K.L.; McCollum, D. Fission yeast Clp1p phosphatase regulates G2/M transition and coordination of cytokinesis with cell cycle progression. Curr. Biol. 2001, 11, 931–940. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.S.; Broadus, M.R.; McLean, J.R.; Feoktistova, A.; Ren, L.; Gould, K.L. Comprehensive proteomics analysis reveals new substrates and regulators of the fission yeast clp1/cdc14 phosphatase. Mol. Cell. Proteom. 2013, 12, 1074–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Queralt, E.; Uhlmann, F. Cdk-counteracting phosphatases unlock mitotic exit. Curr. Opin. Cell Biol. 2008, 20, 661–668. [Google Scholar] [CrossRef] [Green Version]
- Amon, A.; Irniger, S.; Nasmyth, K. Closing the cell cycle circle in yeast: G2 cyclin proteolysis initiated at mitosis persists until the activation of G1 cyclins in the next cycle. Cell 1994, 77, 1037–1050. [Google Scholar] [CrossRef]
- van Werven, F.J.; Amon, A. Regulation of entry into gametogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 3521–3531. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, A.; Sakuno, T.; Watanabe, Y.; Yamamoto, M. Analysis of Schizosaccharomyces pombe Meiosis. Cold Spring Harb. Protoc. 2017, 2017, pdb top079855. [Google Scholar] [CrossRef]
- Zickler, D.; Kleckner, N. Recombination, Pairing, and Synapsis of Homologs during Meiosis. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, J.; Ricco, N.; Grijota-Martinez, C.; Fado, R.; Clotet, J. Redundancy or specificity? The role of the CDK Pho85 in cell cycle control. Int. J. Biochem. Mol. Biol. 2013, 4, 140–149. [Google Scholar] [PubMed]
- Huang, D.; Friesen, H.; Andrews, B. Pho85, a multifunctional cyclin-dependent protein kinase in budding yeast. Mol. Microbiol. 2007, 66, 303–314. [Google Scholar] [CrossRef]
- Mirallas, O.; Ballega, E.; Samper-Martin, B.; Garcia-Marquez, S.; Carballar, R.; Ricco, N.; Jimenez, J.; Clotet, J. Intertwined control of the cell cycle and nucleocytoplasmic transport by the cyclin-dependent kinase Pho85 and RanGTPase Gsp1 in Saccharomyces cerevisiae. Microbiol. Res. 2018, 206, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Dirick, L.; Goetsch, L.; Ammerer, G.; Byers, B. Regulation of meiotic S phase by Ime2 and a Clb5,6-associated kinase in Saccharomyces cerevisiae. Science 1998, 281, 1854–1857. [Google Scholar] [CrossRef] [PubMed]
- Smith, H.E.; Mitchell, A.P. A transcriptional cascade governs entry into meiosis in Saccharomyces cerevisiae. Mol. Cell. Biol. 1989, 9, 2142–2152. [Google Scholar] [CrossRef] [Green Version]
- Stuart, D.; Wittenberg, C. CLB5 and CLB6 are required for premeiotic DNA replication and activation of the meiotic S/M checkpoint. Genes Dev. 1998, 12, 2698–2710. [Google Scholar] [CrossRef] [Green Version]
- Benjamin, K.R.; Zhang, C.; Shokat, K.M.; Herskowitz, I. Control of landmark events in meiosis by the CDK Cdc28 and the meiosis-specific kinase Ime2. Genes Dev. 2003, 17, 1524–1539. [Google Scholar] [CrossRef] [Green Version]
- Schindler, K.; Winter, E. Phosphorylation of Ime2 regulates meiotic progression in Saccharomyces cerevisiae. J. Biol. Chem. 2006, 281, 18307–18316. [Google Scholar] [CrossRef] [Green Version]
- Grandin, N.; Reed, S.I. Differential function and expression of Saccharomyces cerevisiae B-type cyclins in mitosis and meiosis. Mol. Cell. Biol. 1993, 13, 2113–2125. [Google Scholar] [CrossRef] [Green Version]
- Dahmann, C.; Futcher, B. Specialization of B-type cyclins for mitosis or meiosis in S. cerevisiae. Genetics 1995, 140, 957–963. [Google Scholar]
- Carlile, T.M.; Amon, A. Meiosis I is established through division-specific translational control of a cyclin. Cell 2008, 133, 280–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enke, D.A.; Kaldis, P.; Holmes, J.K.; Solomon, M.J. The CDK-activating kinase (Cak1p) from budding yeast has an unusual ATP-binding pocket. J. Biol. Chem. 1999, 274, 1949–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaldis, P.; Sutton, A.; Solomon, M.J. The Cdk-activating kinase (CAK) from budding yeast. Cell 1996, 86, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Thuret, J.Y.; Valay, J.G.; Faye, G.; Mann, C. Civ1 (CAK in vivo), a novel Cdk-activating kinase. Cell 1996, 86, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Jeffrey, P.D.; Russo, A.A.; Polyak, K.; Gibbs, E.; Hurwitz, J.; Massague, J.; Pavletich, N.P. Mechanism of CDK activation revealed by the structure of a cyclinA-CDK2 complex. Nature 1995, 376, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, F.H.; Farrell, A.; Erdjument-Bromage, H.; Tempst, P.; Morgan, D.O. A cyclin-dependent kinase-activating kinase (CAK) in budding yeast unrelated to vertebrate CAK. Science 1996, 273, 1714–1717. [Google Scholar] [CrossRef]
- Enke, D.A.; Kaldis, P.; Solomon, M.J. Kinetic analysis of the cyclin-dependent kinase-activating kinase (Cak1p) from budding yeast. J. Biol. Chem. 2000, 275, 33267–33271. [Google Scholar] [CrossRef] [Green Version]
- Ross, K.E.; Kaldis, P.; Solomon, M.J. Activating phosphorylation of the Saccharomyces cerevisiae cyclin-dependent kinase, cdc28p, precedes cyclin binding. Mol. Biol. Cell 2000, 11, 1597–1609. [Google Scholar] [CrossRef] [Green Version]
- Schindler, K.; Benjamin, K.R.; Martin, A.; Boglioli, A.; Herskowitz, I.; Winter, E. The Cdk-activating kinase Cak1p promotes meiotic S phase through Ime2p. Mol. Cell. Biol. 2003, 23, 8718–8728. [Google Scholar] [CrossRef] [Green Version]
- Honigberg, S.M. Ime2p and Cdc28p: Co-pilots driving meiotic development. J. Cell. Biochem. 2004, 92, 1025–1033. [Google Scholar] [CrossRef]
- Tibbles, K.L.; Sarkar, S.; Novak, B.; Arumugam, P. CDK-dependent nuclear localization of B-cyclin Clb1 promotes FEAR activation during meiosis I in budding yeast. PLoS ONE 2013, 8, e79001. [Google Scholar] [CrossRef] [Green Version]
- Buonomo, S.B.; Rabitsch, K.P.; Fuchs, J.; Gruber, S.; Sullivan, M.; Uhlmann, F.; Petronczki, M.; Toth, A.; Nasmyth, K. Division of the nucleolus and its release of CDC14 during anaphase of meiosis I depends on separase, SPO12, and SLK19. Dev. Cell 2003, 4, 727–739. [Google Scholar] [CrossRef] [Green Version]
- Marston, A.L.; Lee, B.H.; Amon, A. The Cdc14 phosphatase and the FEAR network control meiotic spindle disassembly and chromosome segregation. Dev. Cell 2003, 4, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Arguello-Miranda, O.; Zagoriy, I.; Mengoli, V.; Rojas, J.; Jonak, K.; Oz, T.; Graf, P.; Zachariae, W. Casein Kinase 1 Coordinates Cohesin Cleavage, Gametogenesis, and Exit from M Phase in Meiosis II. Dev. Cell 2017, 40, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Oelschlaegel, T.; Schwickart, M.; Matos, J.; Bogdanova, A.; Camasses, A.; Havlis, J.; Shevchenko, A.; Zachariae, W. The yeast APC/C subunit Mnd2 prevents premature sister chromatid separation triggered by the meiosis-specific APC/C-Ama1. Cell 2005, 120, 773–788. [Google Scholar] [CrossRef] [Green Version]
- Okaz, E.; Arguello-Miranda, O.; Bogdanova, A.; Vinod, P.K.; Lipp, J.J.; Markova, Z.; Zagoriy, I.; Novak, B.; Zachariae, W. Meiotic prophase requires proteolysis of M phase regulators mediated by the meiosis-specific APC/CAma1. Cell 2012, 151, 603–618. [Google Scholar] [CrossRef] [Green Version]
- Salah, S.M.; Nasmyth, K. Destruction of the securin Pds1p occurs at the onset of anaphase during both meiotic divisions in yeast. Chromosoma 2000, 109, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Cooper, K.F.; Mallory, M.J.; Egeland, D.B.; Jarnik, M.; Strich, R. Ama1p is a meiosis-specific regulator of the anaphase promoting complex/cyclosome in yeast. Proc. Natl. Acad. Sci. USA 2000, 97, 14548–14553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, S.S.; Mitchell, A.P. Identification of functionally related genes that stimulate early meiotic gene expression in yeast. Genetics 1993, 133, 67–77. [Google Scholar]
- Smith, H.E.; Su, S.S.; Neigeborn, L.; Driscoll, S.E.; Mitchell, A.P. Role of IME1 expression in regulation of meiosis in Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 6103–6113. [Google Scholar] [CrossRef] [Green Version]
- Kassir, Y.; Granot, D.; Simchen, G. IME1, a positive regulator gene of meiosis in S. cerevisiae. Cell 1988, 52, 853–862. [Google Scholar] [CrossRef]
- De Silva-Udawatta, M.N.; Cannon, J.F. Roles of trehalose phosphate synthase in yeast glycogen metabolism and sporulation. Mol. Microbiol. 2001, 40, 1345–1356. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, A.P.; Herskowitz, I. Activation of meiosis and sporulation by repression of the RME1 product in yeast. Nature 1986, 319, 738–742. [Google Scholar] [CrossRef]
- Colomina, N.; Gari, E.; Gallego, C.; Herrero, E.; Aldea, M. G1 cyclins block the Ime1 pathway to make mitosis and meiosis incompatible in budding yeast. EMBO J. 1999, 18, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primig, M.; Williams, R.M.; Winzeler, E.A.; Tevzadze, G.G.; Conway, A.R.; Hwang, S.Y.; Davis, R.W.; Esposito, R.E. The core meiotic transcriptome in budding yeasts. Nat. Genet. 2000, 26, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; DeRisi, J.; Eisen, M.; Mulholland, J.; Botstein, D.; Brown, P.O.; Herskowitz, I. The transcriptional program of sporulation in budding yeast. Science 1998, 282, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Smith, H.E.; Driscoll, S.E.; Sia, R.A.; Yuan, H.E.; Mitchell, A.P. Genetic evidence for transcriptional activation by the yeast IME1 gene product. Genetics 1993, 133, 775–784. [Google Scholar] [PubMed]
- Vershon, A.K.; Pierce, M. Transcriptional regulation of meiosis in yeast. Curr. Opin. Cell Biol. 2000, 12, 334–339. [Google Scholar] [CrossRef]
- Mandel, S.; Robzyk, K.; Kassir, Y. IME1 gene encodes a transcription factor which is required to induce meiosis in Saccharomyces cerevisiae. Dev. Genet. 1994, 15, 139–147. [Google Scholar] [CrossRef]
- Mitchell, A.P.; Driscoll, S.E.; Smith, H.E. Positive control of sporulation-specific genes by the IME1 and IME2 products in Saccharomyces cerevisiae. Mol. Cell. Biol. 1990, 10, 2104–2110. [Google Scholar] [CrossRef] [Green Version]
- Foiani, M.; Nadjar-Boger, E.; Capone, R.; Sagee, S.; Hashimshoni, T.; Kassir, Y. A meiosis-specific protein kinase, Ime2, is required for the correct timing of DNA replication and for spore formation in yeast meiosis. Mol. Gen. Genet. 1996, 253, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Brush, G.S.; Najor, N.A.; Dombkowski, A.A.; Cukovic, D.; Sawarynski, K.E. Yeast IME2 functions early in meiosis upstream of cell cycle-regulated SBF and MBF targets. PLoS ONE 2012, 7, e31575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, M.; Kawaguchi, H.; Sakata, Y.; Kominami, K.; Hirano, M.; Shima, H.; Akada, R.; Yamashita, I. Initiation of meiosis and sporulation in Saccharomyces cerevisiae requires a novel protein kinase homologue. Mol. Gen. Genet. 1990, 221, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Kominami, K.; Sakata, Y.; Sakai, M.; Yamashita, I. Protein kinase activity associated with the IME2 gene product, a meiotic inducer in the yeast Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 1993, 57, 1731–1735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyers, M. The cyclin-dependent kinase inhibitor p40SIC1 imposes the requirement for Cln G1 cyclin function at Start. Proc. Natl. Acad. Sci. USA 1996, 93, 7772–7776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendenhall, M.D. An inhibitor of p34CDC28 protein kinase activity from Saccharomyces cerevisiae. Science 1993, 259, 216–219. [Google Scholar] [CrossRef] [PubMed]
- Schneider, B.L.; Patton, E.E.; Lanker, S.; Mendenhall, M.D.; Wittenberg, C.; Futcher, B.; Tyers, M. Yeast G1 cyclins are unstable in G1 phase. Nature 1998, 395, 86–89. [Google Scholar] [CrossRef]
- Feldman, R.M.; Correll, C.C.; Kaplan, K.B.; Deshaies, R.J. A complex of Cdc4p, Skp1p, and Cdc53p/cullin catalyzes ubiquitination of the phosphorylated CDK inhibitor Sic1p. Cell 1997, 91, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Verma, R.; Feldman, R.M.; Deshaies, R.J. SIC1 is ubiquitinated in vitro by a pathway that requires CDC4, CDC34, and cyclin/CDK activities. Mol. Biol. Cell 1997, 8, 1427–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowyra, D.; Craig, K.L.; Tyers, M.; Elledge, S.J.; Harper, J.W. F-box proteins are receptors that recruit phosphorylated substrates to the SCF ubiquitin-ligase complex. Cell 1997, 91, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Sclafani, R.A. Cdc7p-Dbf4p becomes famous in the cell cycle. J. Cell Sci. 2000, 113, 2111–2117. [Google Scholar]
- Masai, H.; Arai, K. Cdc7 kinase complex: A key regulator in the initiation of DNA replication. J. Cell. Physiol. 2002, 190, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Valentin, G.; Schwob, E.; Della Seta, F. Dual role of the Cdc7-regulatory protein Dbf4 during yeast meiosis. J. Biol. Chem. 2006, 281, 2828–2834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, L.; Zhang, C.; Shokat, K.M.; Hollingsworth, N.M. Chemical inactivation of cdc7 kinase in budding yeast results in a reversible arrest that allows efficient cell synchronization prior to meiotic recombination. Genetics 2006, 174, 1767–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.Y.; Bisig, C.G.; Conrad, M.M.; Ditamo, Y.; Previato de Almeida, L.; Dresser, M.E.; Pezza, R.J. Extranuclear Structural Components that Mediate Dynamic Chromosome Movements in Yeast Meiosis. Curr. Biol. 2020, 30, 1207–1216.e1204. [Google Scholar] [CrossRef]
- Wanat, J.J.; Kim, K.P.; Koszul, R.; Zanders, S.; Weiner, B.; Kleckner, N.; Alani, E. Csm4, in collaboration with Ndj1, mediates telomere-led chromosome dynamics and recombination during yeast meiosis. PLoS Genet. 2008, 4, e1000188. [Google Scholar] [CrossRef] [PubMed]
- Koszul, R.; Kim, K.P.; Prentiss, M.; Kleckner, N.; Kameoka, S. Meiotic chromosomes move by linkage to dynamic actin cables with transduction of force through the nuclear envelope. Cell 2008, 133, 1188–1201. [Google Scholar] [CrossRef] [Green Version]
- Martinez-Garcia, M.; Schubert, V.; Osman, K.; Darbyshire, A.; Sanchez-Moran, E.; Franklin, F.C.H. TOPII and chromosome movement help remove interlocks between entangled chromosomes during meiosis. J. Cell Biol. 2018, 217, 4070–4079. [Google Scholar] [CrossRef]
- Lee, C.Y.; Conrad, M.N.; Dresser, M.E. Meiotic chromosome pairing is promoted by telomere-led chromosome movements independent of bouquet formation. PLoS Genet. 2012, 8, e1002730. [Google Scholar] [CrossRef] [Green Version]
- Conrad, M.N.; Lee, C.Y.; Chao, G.; Shinohara, M.; Kosaka, H.; Shinohara, A.; Conchello, J.A.; Dresser, M.E. Rapid telomere movement in meiotic prophase is promoted by NDJ1, MPS3, and CSM4 and is modulated by recombination. Cell 2008, 133, 1175–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trelles-Sticken, E.; Adelfalk, C.; Loidl, J.; Scherthan, H. Meiotic telomere clustering requires actin for its formation and cohesin for its resolution. J. Cell Biol. 2005, 170, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.N.; Penkner, A.; Ohta, K.; Klein, F.; Nicolas, A. B-type cyclins CLB5 and CLB6 control the initiation of recombination and synaptonemal complex formation in yeast meiosis. Curr. Biol. 2001, 11, 88–97. [Google Scholar] [CrossRef] [Green Version]
- Henderson, K.A.; Kee, K.; Maleki, S.; Santini, P.A.; Keeney, S. Cyclin-dependent kinase directly regulates initiation of meiotic recombination. Cell 2006, 125, 1321–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, Z.; Mori, S.; Oshiumi, H.; Matsuzaki, K.; Shinohara, M.; Shinohara, A. Cyclin-dependent kinase promotes formation of the synaptonemal complex in yeast meiosis. Genes Cells 2010, 15, 1036–1050. [Google Scholar] [CrossRef] [PubMed]
- Panizza, S.; Mendoza, M.A.; Berlinger, M.; Huang, L.; Nicolas, A.; Shirahige, K.; Klein, F. Spo11-accessory proteins link double-strand break sites to the chromosome axis in early meiotic recombination. Cell 2011, 146, 372–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasanuma, H.; Hirota, K.; Fukuda, T.; Kakusho, N.; Kugou, K.; Kawasaki, Y.; Shibata, T.; Masai, H.; Ohta, K. Cdc7-dependent phosphorylation of Mer2 facilitates initiation of yeast meiotic recombination. Genes Dev. 2008, 22, 398–410. [Google Scholar] [CrossRef] [Green Version]
- Wan, L.; Niu, H.; Futcher, B.; Zhang, C.; Shokat, K.M.; Boulton, S.J.; Hollingsworth, N.M. Cdc28-Clb5 (CDK-S) and Cdc7-Dbf4 (DDK) collaborate to initiate meiotic recombination in yeast. Genes Dev. 2008, 22, 386–397. [Google Scholar] [CrossRef] [Green Version]
- Murakami, H.; Keeney, S. Temporospatial coordination of meiotic DNA replication and recombination via DDK recruitment to replisomes. Cell 2014, 158, 861–873. [Google Scholar] [CrossRef] [Green Version]
- Murakami, H.; Keeney, S. DDK links replication and recombination in meiosis. Cell Cycle 2014, 13, 3621–3622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manfrini, N.; Guerini, I.; Citterio, A.; Lucchini, G.; Longhese, M.P. Processing of meiotic DNA double strand breaks requires cyclin-dependent kinase and multiple nucleases. J. Biol. Chem. 2010, 285, 11628–11637. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.P.; Unal, E.; Brar, G.A.; Amon, A. Meiosis I chromosome segregation is established through regulation of microtubule-kinetochore interactions. Elife 2012, 1, e00117. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.; Herskowitz, I. Gametogenesis in yeast is regulated by a transcriptional cascade dependent on Ndt80. Mol. Cell 1998, 1, 685–696. [Google Scholar] [CrossRef]
- Leu, J.Y.; Roeder, G.S. The pachytene checkpoint in S. cerevisiae depends on Swe1-mediated phosphorylation of the cyclin-dependent kinase Cdc28. Mol. Cell 1999, 4, 805–814. [Google Scholar] [CrossRef]
- Chen, X.; Gaglione, R.; Leong, T.; Bednor, L.; de Los Santos, T.; Luk, E.; Airola, M.; Hollingsworth, N.M. Mek1 coordinates meiotic progression with DNA break repair by directly phosphorylating and inhibiting the yeast pachytene exit regulator Ndt80. PLoS Genet. 2018, 14, e1007832. [Google Scholar] [CrossRef] [PubMed]
- Tung, K.S.; Hong, E.J.; Roeder, G.S. The pachytene checkpoint prevents accumulation and phosphorylation of the meiosis-specific transcription factor Ndt80. Proc. Natl. Acad. Sci. USA 2000, 97, 12187–12192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, S.R.; Friesen, H.; Segall, J. NDT80 and the meiotic recombination checkpoint regulate expression of middle sporulation-specific genes in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 5750–5761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, V.V.; Hochwagen, A. The meiotic checkpoint network: Step-by-step through meiotic prophase. Cold Spring Harb. Perspect. Biol. 2014, 6, a016675. [Google Scholar] [CrossRef] [PubMed]
- Prugar, E.; Burnett, C.; Chen, X.; Hollingsworth, N.M. Coordination of Double Strand Break Repair and Meiotic Progression in Yeast by a Mek1-Ndt80 Negative Feedback Loop. Genetics 2017, 206, 497–512. [Google Scholar] [CrossRef] [Green Version]
- Lydall, D.; Nikolsky, Y.; Bishop, D.K.; Weinert, T. A meiotic recombination checkpoint controlled by mitotic checkpoint genes. Nature 1996, 383, 840–843. [Google Scholar] [CrossRef]
- Wu, H.Y.; Ho, H.C.; Burgess, S.M. Mek1 kinase governs outcomes of meiotic recombination and the checkpoint response. Curr. Biol. 2010, 20, 1707–1716. [Google Scholar] [CrossRef] [Green Version]
- Ho, H.C.; Burgess, S.M. Pch2 acts through Xrs2 and Tel1/ATM to modulate interhomolog bias and checkpoint function during meiosis. PLoS Genet. 2011, 7, e1002351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ontoso, D.; Acosta, I.; van Leeuwen, F.; Freire, R.; San-Segundo, P.A. Dot1-dependent histone H3K79 methylation promotes activation of the Mek1 meiotic checkpoint effector kinase by regulating the Hop1 adaptor. PLoS Genet. 2013, 9, e1003262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penedos, A.; Johnson, A.L.; Strong, E.; Goldman, A.S.; Carballo, J.A.; Cha, R.S. Essential and Checkpoint Functions of Budding Yeast ATM and ATR during Meiotic Prophase Are Facilitated by Differential Phosphorylation of a Meiotic Adaptor Protein, Hop1. PLoS ONE 2015, 10, e0134297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, M.; Wagner, M.; Xie, J.; Gailus-Durner, V.; Six, J.; Vershon, A.K.; Winter, E. Transcriptional regulation of the SMK1 mitogen-activated protein kinase gene during meiotic development in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 5970–5980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierce, M.; Benjamin, K.R.; Montano, S.P.; Georgiadis, M.M.; Winter, E.; Vershon, A.K. Sum1 and Ndt80 proteins compete for binding to middle sporulation element sequences that control meiotic gene expression. Mol. Cell. Biol. 2003, 23, 4814–4825. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.T.; Bungard, D.; Shin, M.E.; Moore, M.; Winter, E. The Ime2 protein kinase enhances the disassociation of the Sum1 repressor from middle meiotic promoters. Mol. Cell. Biol. 2009, 29, 4352–4362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, M.E.; Skokotas, A.; Winter, E. The Cdk1 and Ime2 protein kinases trigger exit from meiotic prophase in Saccharomyces cerevisiae by inhibiting the Sum1 transcriptional repressor. Mol. Cell. Biol. 2010, 30, 2996–3003. [Google Scholar] [CrossRef] [Green Version]
- Lo, H.C.; Kunz, R.C.; Chen, X.; Marullo, A.; Gygi, S.P.; Hollingsworth, N.M. Cdc7-Dbf4 is a gene-specific regulator of meiotic transcription in yeast. Mol. Cell. Biol. 2012, 32, 541–557. [Google Scholar] [CrossRef] [Green Version]
- Winter, E. The Sum1/Ndt80 transcriptional switch and commitment to meiosis in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2012, 76, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Pak, J.; Segall, J. Role of Ndt80, Sum1, and Swe1 as targets of the meiotic recombination checkpoint that control exit from pachytene and spore formation in Saccharomyces cerevisiae. Mol. Cell. Biol. 2002, 22, 6430–6440. [Google Scholar] [CrossRef] [Green Version]
- Sourirajan, A.; Lichten, M. Polo-like kinase Cdc5 drives exit from pachytene during budding yeast meiosis. Genes Dev. 2008, 22, 2627–2632. [Google Scholar] [CrossRef] [Green Version]
- Berchowitz, L.E.; Gajadhar, A.S.; van Werven, F.J.; De Rosa, A.A.; Samoylova, M.L.; Brar, G.A.; Xu, Y.; Xiao, C.; Futcher, B.; Weissman, J.S.; et al. A developmentally regulated translational control pathway establishes the meiotic chromosome segregation pattern. Genes Dev. 2013, 27, 2147–2163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marston, A.L. Shugoshins: Tension-sensitive pericentromeric adaptors safeguarding chromosome segregation. Mol. Cell. Biol. 2015, 35, 634–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitajima, T.S.; Kawashima, S.A.; Watanabe, Y. The conserved kinetochore protein shugoshin protects centromeric cohesion during meiosis. Nature 2004, 427, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Shirayama, M.; Toth, A.; Galova, M.; Nasmyth, K. APC(Cdc20) promotes exit from mitosis by destroying the anaphase inhibitor Pds1 and cyclin Clb5. Nature 1999, 402, 203–207. [Google Scholar] [CrossRef]
- Kamieniecki, R.J.; Liu, L.; Dawson, D.S. FEAR but not MEN genes are required for exit from meiosis I. Cell Cycle 2005, 4, 1093–1098. [Google Scholar] [CrossRef] [Green Version]
- Stegmeier, F.; Visintin, R.; Amon, A. Separase, polo kinase, the kinetochore protein Slk19, and Spo12 function in a network that controls Cdc14 localization during early anaphase. Cell 2002, 108, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Visintin, R.; Hwang, E.S.; Amon, A. Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus. Nature 1999, 398, 818–823. [Google Scholar] [CrossRef]
- Yoshida, S.; Toh-e, A. Budding yeast Cdc5 phosphorylates Net1 and assists Cdc14 release from the nucleolus. Biochem. Biophys. Res. Commun. 2002, 294, 687–691. [Google Scholar] [CrossRef]
- Holt, L.J.; Hutti, J.E.; Cantley, L.C.; Morgan, D.O. Evolution of Ime2 phosphorylation sites on Cdk1 substrates provides a mechanism to limit the effects of the phosphatase Cdc14 in meiosis. Mol. Cell 2007, 25, 689–702. [Google Scholar] [CrossRef] [Green Version]
- Phizicky, D.V.; Berchowitz, L.E.; Bell, S.P. Multiple kinases inhibit origin licensing and helicase activation to ensure reductive cell division during meiosis. Elife 2018, 7. [Google Scholar] [CrossRef]
- Guttmann-Raviv, N.; Boger-Nadjar, E.; Edri, I.; Kassir, Y. Cdc28 and Ime2 possess redundant functions in promoting entry into premeiotic DNA replication in Saccharomyces cerevisiae. Genetics 2001, 159, 1547–1558. [Google Scholar] [PubMed]
- Berchowitz, L.E.; Kabachinski, G.; Walker, M.R.; Carlile, T.M.; Gilbert, W.V.; Schwartz, T.U.; Amon, A. Regulated Formation of an Amyloid-like Translational Repressor Governs Gametogenesis. Cell 2015, 163, 406–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, K.; Bell, R.B.; Yunus, J.; Amon, A.; Berchowitz, L.E. Phosphorylation-Mediated Clearance of Amyloid-like Assemblies in Meiosis. Dev. Cell 2018, 45, 392–405.e396. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zhang, R.; Feng, W.; Tsuchiya, D.; Ballew, O.; Li, J.; Denic, V.; Lacefield, S. Autophagy of an Amyloid-like Translational Repressor Regulates Meiotic Exit. Dev. Cell 2020, 52, 141–151.e5. [Google Scholar] [CrossRef]
- Wang, F.; Denic, V.; Lacefield, S. Autophagy prevents runaway meiotic divisions. Autophagy 2020, 16, 969–970. [Google Scholar] [CrossRef]
- Liu, J.; Kipreos, E.T. Evolution of cyclin-dependent kinases (CDKs) and CDK-activating kinases (CAKs): Differential conservation of CAKs in yeast and metazoa. Mol. Biol. Evol. 2000, 17, 1061–1074. [Google Scholar] [CrossRef] [Green Version]
- Iino, Y.; Hiramine, Y.; Yamamoto, M. The role of cdc2 and other genes in meiosis in Schizosaccharomyces pombe. Genetics 1995, 140, 1235–1245. [Google Scholar] [PubMed]
- Nurse, P.; Bissett, Y. Gene required in G1 for commitment to cell cycle and in G2 for control of mitosis in fission yeast. Nature 1981, 292, 558–560. [Google Scholar] [CrossRef]
- Tanaka, K.; Okayama, H. A pcl-like cyclin activates the Res2p-Cdc10p cell cycle "start" transcriptional factor complex in fission yeast. Mol. Biol. Cell 2000, 11, 2845–2862. [Google Scholar] [CrossRef] [Green Version]
- Mondesert, O.; McGowan, C.H.; Russell, P. Cig2, a B-type cyclin, promotes the onset of S in Schizosaccharomyces pombe. Mol. Cell. Biol. 1996, 16, 1527–1533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Castellanos, C.; Labib, K.; Moreno, S. B-type cyclins regulate G1 progression in fission yeast in opposition to the p25rum1 cdk inhibitor. EMBO J. 1996, 15, 839–849. [Google Scholar] [CrossRef]
- Booher, R.; Beach, D. Involvement of cdc13+ in mitotic control in Schizosaccharomyces pombe: Possible interaction of the gene product with microtubules. EMBO J. 1988, 7, 2321–2327. [Google Scholar] [CrossRef] [PubMed]
- Booher, R.N.; Alfa, C.E.; Hyams, J.S.; Beach, D.H. The fission yeast cdc2/cdc13/suc1 protein kinase: Regulation of catalytic activity and nuclear localization. Cell 1989, 58, 485–497. [Google Scholar] [CrossRef]
- Moreno, S.; Hayles, J.; Nurse, P. Regulation of p34cdc2 protein kinase during mitosis. Cell 1989, 58, 361–372. [Google Scholar] [CrossRef]
- Bueno, A.; Richardson, H.; Reed, S.I.; Russell, P. A fission yeast B-type cyclin functioning early in the cell cycle. Cell 1991, 66, 149–159. [Google Scholar] [CrossRef]
- Forsburg, S.L.; Nurse, P. Identification of a G1-type cyclin puc1+ in the fission yeast Schizosaccharomyces pombe. Nature 1991, 351, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Obara-Ishihara, T.; Okayama, H. A B-type cyclin negatively regulates conjugation via interacting with cell cycle ’start’ genes in fission yeast. EMBO J. 1994, 13, 1863–1872. [Google Scholar] [CrossRef]
- Malapeira, J.; Moldon, A.; Hidalgo, E.; Smith, G.R.; Nurse, P.; Ayte, J. A meiosis-specific cyclin regulated by splicing is required for proper progression through meiosis. Mol. Cell. Biol. 2005, 25, 6330–6337. [Google Scholar] [CrossRef] [Green Version]
- Averbeck, N.; Sunder, S.; Sample, N.; Wise, J.A.; Leatherwood, J. Negative control contributes to an extensive program of meiotic splicing in fission yeast. Mol. Cell 2005, 18, 491–498. [Google Scholar] [CrossRef]
- Martin-Castellanos, C.; Blanco, M.; Rozalen, A.E.; Perez-Hidalgo, L.; Garcia, A.I.; Conde, F.; Mata, J.; Ellermeier, C.; Davis, L.; San-Segundo, P.; et al. A large-scale screen in S. pombe identifies seven novel genes required for critical meiotic events. Curr. Biol. 2005, 15, 2056–2062. [Google Scholar] [CrossRef] [Green Version]
- Martin-Castellanos, C.; Blanco, M.A.; de Prada, J.M.; Moreno, S. The puc1 cyclin regulates the G1 phase of the fission yeast cell cycle in response to cell size. Mol. Biol Cell 2000, 11, 543–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borgne, A.; Murakami, H.; Ayte, J.; Nurse, P. The G1/S cyclin Cig2p during meiosis in fission yeast. Mol. Biol. Cell 2002, 13, 2080–2090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez-Escribano, P.; Nurse, P. A single cyclin-CDK complex is sufficient for both mitotic and meiotic progression in fission yeast. Nat. Commun. 2015, 6, 6871. [Google Scholar] [CrossRef] [Green Version]
- Bustamante-Jaramillo, L.F.; Ramos, C.; Alonso, L.; Sesmero, A.; Segurado, M.; Martin-Castellanos, C. CDK contribution to DSB formation and recombination in fission yeast meiosis. PLoS Genet. 2019, 15, e1007876. [Google Scholar] [CrossRef] [PubMed]
- Moldon, A.; Malapeira, J.; Gabrielli, N.; Gogol, M.; Gomez-Escoda, B.; Ivanova, T.; Seidel, C.; Ayte, J. Promoter-driven splicing regulation in fission yeast. Nature 2008, 455, 997–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decottignies, A.; Zarzov, P.; Nurse, P. In vivo localisation of fission yeast cyclin-dependent kinase cdc2p and cyclin B cdc13p during mitosis and meiosis. J. Cell Sci. 2001, 114, 2627–2640. [Google Scholar] [PubMed]
- Murakami, H.; Nurse, P. Meiotic DNA replication checkpoint control in fission yeast. Genes Dev. 1999, 13, 2581–2593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molz, L.; Beach, D. Characterization of the fission yeast mcs2 cyclin and its associated protein kinase activity. EMBO J. 1993, 12, 1723–1732. [Google Scholar] [CrossRef]
- Bamps, S.; Westerling, T.; Pihlak, A.; Tafforeau, L.; Vandenhaute, J.; Makela, T.P.; Hermand, D. Mcs2 and a novel CAK subunit Pmh1 associate with Skp1 in fission yeast. Biochem. Biophys. Res. Commun. 2004, 325, 1424–1432. [Google Scholar] [CrossRef]
- Buck, V.; Russell, P.; Millar, J.B. Identification of a cdk-activating kinase in fission yeast. EMBO J. 1995, 14, 6173–6183. [Google Scholar] [CrossRef] [PubMed]
- Damagnez, V.; Makela, T.P.; Cottarel, G. Schizosaccharomyces pombe Mop1-Mcs2 is related to mammalian CAK. EMBO J. 1995, 14, 6164–6172. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Miklos, I.; Du, H.; Watt, S.; Szilagyi, Z.; Saiz, J.E.; Madabhushi, R.; Penkett, C.J.; Sipiczki, M.; Bahler, J.; et al. Impairment of the TFIIH-associated CDK-activating kinase selectively affects cell cycle-regulated gene expression in fission yeast. Mol. Biol. Cell 2005, 16, 2734–2745. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Saiz, J.E.; Barton, W.A.; Fisher, R.P. Cdc2 activation in fission yeast depends on Mcs6 and Csk1, two partially redundant Cdk-activating kinases (CAKs). Curr. Biol. 1999, 9, 441–444. [Google Scholar] [CrossRef] [Green Version]
- Saiz, J.E.; Fisher, R.P. A CDK-activating kinase network is required in cell cycle control and transcription in fission yeast. Curr. Biol. 2002, 12, 1100–1105. [Google Scholar] [CrossRef] [Green Version]
- Tsakraklides, V.; Solomon, M.J. Comparison of Cak1p-like cyclin-dependent kinase-activating kinases. J. Biol. Chem. 2002, 277, 33482–33489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermand, D.; Pihlak, A.; Westerling, T.; Damagnez, V.; Vandenhaute, J.; Cottarel, G.; Makela, T.P. Fission yeast Csk1 is a CAK-activating kinase (CAKAK). EMBO J. 1998, 17, 7230–7238. [Google Scholar] [CrossRef] [Green Version]
- Blyth, J.; Makrantoni, V.; Barton, R.E.; Spanos, C.; Rappsilber, J.; Marston, A.L. Genes Important for Schizosaccharomyces pombe Meiosis Identified Through a Functional Genomics Screen. Genetics 2018, 208, 589–603. [Google Scholar] [CrossRef] [Green Version]
- Daga, R.R.; Bolanos, P.; Moreno, S. Regulated mRNA stability of the Cdk inhibitor Rum1 links nutrient status to cell cycle progression. Curr. Biol. 2003, 13, 2015–2024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daya-Makin, M.; Szankasi, P.; Tang, L.; MacRae, D.; Pelech, S.L. Regulation of p105wee1 and p34cdc2 during meiosis in Schizosaccharomyces pombe. Biochem. Cell Biol. 1992, 70, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- McPheeters, D.S.; Cremona, N.; Sunder, S.; Chen, H.M.; Averbeck, N.; Leatherwood, J.; Wise, J.A. A complex gene regulatory mechanism that operates at the nexus of multiple RNA processing decisions. Nat. Struct. Mol. Biol. 2009, 16, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arata, M.; Sato, M.; Yamashita, A.; Yamamoto, M. The RNA-binding protein Spo5 promotes meiosis II by regulating cyclin Cdc13 in fission yeast. Genes Cells 2014, 19, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Kasama, T.; Shigehisa, A.; Hirata, A.; Saito, T.T.; Tougan, T.; Okuzaki, D.; Nojima, H. Spo5/Mug12, a putative meiosis-specific RNA-binding protein, is essential for meiotic progression and forms Mei2 dot-like nuclear foci. Eukaryot. Cell 2006, 5, 1301–1313. [Google Scholar] [CrossRef] [Green Version]
- Blanco, M.A.; Pelloquin, L.; Moreno, S. Fission yeast mfr1 activates APC and coordinates meiotic nuclear division with sporulation. J. Cell Sci. 2001, 114, 2135–2143. [Google Scholar] [PubMed]
- Asakawa, H.; Kitamura, K.; Shimoda, C. A novel Cdc20-related WD-repeat protein, Fzr1, is required for spore formation in Schizosaccharomyces pombe. Mol. Genet. Genom. 2001, 265, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Aoi, Y.; Arai, K.; Miyamoto, M.; Katsuta, Y.; Yamashita, A.; Sato, M.; Yamamoto, M. Cuf2 boosts the transcription of APC/C activator Fzr1 to terminate the meiotic division cycle. EMBO Rep. 2013, 14, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Chikashige, Y.; Yamane, M.; Okamasa, K.; Osakada, H.; Tsutsumi, C.; Nagahama, Y.; Fukuta, N.; Haraguchi, T.; Hiraoka, Y. Fission yeast APC/C activators Slp1 and Fzr1 sequentially trigger two consecutive nuclear divisions during meiosis. FEBS Lett. 2017, 591, 1029–1040. [Google Scholar] [CrossRef] [Green Version]
- Egel, R. Physiological aspects of conjugation in fission yeast. Planta 1971, 98, 89–96. [Google Scholar] [CrossRef]
- Yamamoto, M. The selective elimination of messenger RNA underlies the mitosis-meiosis switch in fission yeast. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2010, 86, 788–797. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, A.; Hiraoka, Y. How do meiotic chromosomes meet their homologous partners?: Lessons from fission yeast. Bioessays 2001, 23, 526–533. [Google Scholar] [CrossRef]
- McLeod, M.; Stein, M.; Beach, D. The product of the mei3+ gene, expressed under control of the mating-type locus, induces meiosis and sporulation in fission yeast. EMBO J. 1987, 6, 729–736. [Google Scholar] [CrossRef] [PubMed]
- McLeod, M.; Beach, D. A specific inhibitor of the ran1+ protein kinase regulates entry into meiosis in Schizosaccharomyces pombe. Nature 1988, 332, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; McLeod, M. Molecular mimicry in development: Identification of ste11+ as a substrate and mei3+ as a pseudosubstrate inhibitor of ran1+ kinase. Cell 1996, 87, 869–880. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, K.; Katayama, S.; Dhut, S.; Sato, M.; Watanabe, Y.; Yamamoto, M.; Toda, T. Phosphorylation of Mei2 and Ste11 by Pat1 kinase inhibits sexual differentiation via ubiquitin proteolysis and 14-3-3 protein in fission yeast. Dev. Cell 2001, 1, 389–399. [Google Scholar] [CrossRef] [Green Version]
- Sugimoto, A.; Iino, Y.; Maeda, T.; Watanabe, Y.; Yamamoto, M. Schizosaccharomyces pombe ste11+ encodes a transcription factor with an HMG motif that is a critical regulator of sexual development. Genes Dev. 1991, 5, 1990–1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harigaya, Y.; Tanaka, H.; Yamanaka, S.; Tanaka, K.; Watanabe, Y.; Tsutsumi, C.; Chikashige, Y.; Hiraoka, Y.; Yamashita, A.; Yamamoto, M. Selective elimination of messenger RNA prevents an incidence of untimely meiosis. Nature 2006, 442, 45–50. [Google Scholar] [CrossRef]
- Shichino, Y.; Yamashita, A.; Yamamoto, M. Meiotic long non-coding meiRNA accumulates as a dot at its genetic locus facilitated by Mmi1 and plays as a decoy to lure Mmi1. Open Biol. 2014, 4, 140022. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, S.; Yamashita, A.; Harigaya, Y.; Iwata, R.; Yamamoto, M. Importance of polyadenylation in the selective elimination of meiotic mRNAs in growing S. pombe cells. EMBO J. 2010, 29, 2173–2181. [Google Scholar] [CrossRef]
- St-Andre, O.; Lemieux, C.; Perreault, A.; Lackner, D.H.; Bahler, J.; Bachand, F. Negative regulation of meiotic gene expression by the nuclear poly(a)-binding protein in fission yeast. J. Biol. Chem. 2010, 285, 27859–27868. [Google Scholar] [CrossRef] [Green Version]
- Cremona, N.; Potter, K.; Wise, J.A. A meiotic gene regulatory cascade driven by alternative fates for newly synthesized transcripts. Mol. Biol. Cell 2011, 22, 66–77. [Google Scholar] [CrossRef]
- Horie, S.; Watanabe, Y.; Tanaka, K.; Nishiwaki, S.; Fujioka, H.; Abe, H.; Yamamoto, M.; Shimoda, C. The Schizosaccharomyces pombe mei4+ gene encodes a meiosis-specific transcription factor containing a forkhead DNA-binding domain. Mol. Cell. Biol. 1998, 18, 2118–2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.M.; Futcher, B.; Leatherwood, J. The fission yeast RNA binding protein Mmi1 regulates meiotic genes by controlling intron specific splicing and polyadenylation coupled RNA turnover. PLoS ONE 2011, 6, e26804. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.M.; Rosebrock, A.P.; Khan, S.R.; Futcher, B.; Leatherwood, J.K. Repression of meiotic genes by antisense transcription and by Fkh2 transcription factor in Schizosaccharomyces pombe. PLoS ONE 2012, 7, e29917. [Google Scholar] [CrossRef] [Green Version]
- Murakami-Tonami, Y.; Yamada-Namikawa, C.; Tochigi, A.; Hasegawa, N.; Kojima, H.; Kunimatsu, M.; Nakanishi, M.; Murakami, H. Mei4p coordinates the onset of meiosis I by regulating cdc25+ in fission yeast. Proc. Natl. Acad. Sci. USA 2007, 104, 14688–14693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grallert, B.; Sipiczki, M. Common genes and pathways in the regulation of the mitotic and meiotic cell cycles of Schizosaccharomyces pombe. Curr. Genet. 1991, 20, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Featherstone, C.; Russell, P. Fission yeast p107wee1 mitotic inhibitor is a tyrosine/serine kinase. Nature 1991, 349, 808–811. [Google Scholar] [CrossRef]
- Parker, L.L.; Atherton-Fessler, S.; Piwnica-Worms, H. p107wee1 is a dual-specificity kinase that phosphorylates p34cdc2 on tyrosine 15. Proc. Natl. Acad. Sci. USA 1992, 89, 2917–2921. [Google Scholar] [CrossRef] [Green Version]
- Beach, D. Sexual differentiation is controlled by a protein kinase encoded by the ran1+ gene in fission yeast. Cold Spring Harb. Symp. Quant. Biol. 1985, 50, 635–641. [Google Scholar] [CrossRef]
- Bahler, J.; Wyler, T.; Loidl, J.; Kohli, J. Unusual nuclear structures in meiotic prophase of fission yeast: A cytological analysis. J. Cell Biol. 1993, 121, 241–256. [Google Scholar] [CrossRef] [Green Version]
- Chikashige, Y.; Ding, D.Q.; Funabiki, H.; Haraguchi, T.; Mashiko, S.; Yanagida, M.; Hiraoka, Y. Telomere-led premeiotic chromosome movement in fission yeast. Science 1994, 264, 270–273. [Google Scholar] [CrossRef]
- Chikashige, Y.; Ding, D.Q.; Imai, Y.; Yamamoto, M.; Haraguchi, T.; Hiraoka, Y. Meiotic nuclear reorganization: Switching the position of centromeres and telomeres in the fission yeast Schizosaccharomyces pombe. EMBO J. 1997, 16, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikashige, Y.; Tsutsumi, C.; Yamane, M.; Okamasa, K.; Haraguchi, T.; Hiraoka, Y. Meiotic proteins bqt1 and bqt2 tether telomeres to form the bouquet arrangement of chromosomes. Cell 2006, 125, 59–69. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Jin, Y.; Cande, W.Z. Bqt2p is essential for initiating telomere clustering upon pheromone sensing in fission yeast. J. Cell Biol. 2006, 173, 845–851. [Google Scholar] [CrossRef]
- Klutstein, M.; Cooper, J.P. The Chromosomal Courtship Dance-homolog pairing in early meiosis. Curr. Opin. Cell Biol. 2014, 26, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Klutstein, M.; Fennell, A.; Fernandez-Alvarez, A.; Cooper, J.P. The telomere bouquet regulates meiotic centromere assembly. Nat. Cell Biol. 2015, 17, 458–469. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.P.; Watanabe, Y.; Nurse, P. Fission yeast Taz1 protein is required for meiotic telomere clustering and recombination. Nature 1998, 392, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.; Smith, G.R. The meiotic bouquet promotes homolog interactions and restricts ectopic recombination in Schizosaccharomyces pombe. Genetics 2006, 174, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, K.; Bez, C.; Fennell, A.; Cooper, J.P. A single internal telomere tract ensures meiotic spindle formation. EMBO Rep. 2013, 14, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Tomita, K.; Cooper, J.P. The telomere bouquet controls the meiotic spindle. Cell 2007, 130, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Niwa, O.; Shimanuki, M.; Miki, F. Telomere-led bouquet formation facilitates homologous chromosome pairing and restricts ectopic interaction in fission yeast meiosis. EMBO J. 2000, 19, 3831–3840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, D.Q.; Yamamoto, A.; Haraguchi, T.; Hiraoka, Y. Dynamics of homologous chromosome pairing during meiotic prophase in fission yeast. Dev. Cell 2004, 6, 329–341. [Google Scholar] [CrossRef] [Green Version]
- Fowler, K.R.; Gutierrez-Velasco, S.; Martin-Castellanos, C.; Smith, G.R. Protein determinants of meiotic DNA break hot spots. Mol. Cell 2013, 49, 983–996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, L.; Rozalen, A.E.; Moreno, S.; Smith, G.R.; Martin-Castellanos, C. Rec25 and Rec27, novel linear-element components, link cohesin to meiotic DNA breakage and recombination. Curr. Biol. 2008, 18, 849–854. [Google Scholar] [CrossRef] [PubMed]
- Ellermeier, C.; Smith, G.R. Cohesins are required for meiotic DNA breakage and recombination in Schizosaccharomyces pombe. Proc. Natl. Acad. Sci. USA 2005, 102, 10952–10957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenz, A.; Wells, J.L.; Pryce, D.W.; Novatchkova, M.; Eisenhaber, F.; McFarlane, R.J.; Loidl, J.S. pombe meiotic linear elements contain proteins related to synaptonemal complex components. J. Cell Sci. 2004, 117, 3343–3351. [Google Scholar] [CrossRef] [Green Version]
- Keeney, S.; Giroux, C.N.; Kleckner, N. Meiosis-specific DNA double-strand breaks are catalyzed by Spo11, a member of a widely conserved protein family. Cell 1997, 88, 375–384. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, M.D.; Farah, J.A.; Smith, G.R. Meiotic DNA breaks associated with recombination in S. pombe. Mol. Cell 2000, 5, 883–888. [Google Scholar] [CrossRef]
- Lam, I.; Keeney, S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb. Perspect. Biol. 2014, 7, a016634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Castellanos, C.; Fowler, K.R.; Smith, G.R. Making chromosomes hot for breakage. Cell Cycle 2013, 12, 1327–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phadnis, N.; Cipak, L.; Polakova, S.; Hyppa, R.W.; Cipakova, I.; Anrather, D.; Karvaiova, L.; Mechtler, K.; Smith, G.R.; Gregan, J. Casein Kinase 1 and Phosphorylation of Cohesin Subunit Rec11 (SA3) Promote Meiotic Recombination through Linear Element Formation. PLoS Genet. 2015, 11, e1005225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnar, M.; Doll, E.; Yamamoto, A.; Hiraoka, Y.; Kohli, J. Linear element formation and their role in meiotic sister chromatid cohesion and chromosome pairing. J. Cell Sci. 2003, 116, 1719–1731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molnar, M.; Bahler, J.; Sipiczki, M.; Kohli, J. The rec8 gene of Schizosaccharomyces pombe is involved in linear element formation, chromosome pairing and sister-chromatid cohesion during meiosis. Genetics 1995, 141, 61–73. [Google Scholar] [PubMed]
- Sakuno, T.; Watanabe, Y. Phosphorylation of cohesin Rec11/SA3 by casein kinase 1 promotes homologous recombination by assembling the meiotic chromosome axis. Dev. Cell 2015, 32, 220–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyoshi, T.; Ito, M.; Kugou, K.; Yamada, S.; Furuichi, M.; Oda, A.; Yamada, T.; Hirota, K.; Masai, H.; Ohta, K. A central coupler for recombination initiation linking chromosome architecture to S phase checkpoint. Mol. Cell 2012, 47, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Tougan, T.; Kasama, T.; Ohtaka, A.; Okuzaki, D.; Saito, T.T.; Russell, P.; Nojima, H. The Mek1 phosphorylation cascade plays a role in meiotic recombination of Schizosaccharomyces pombe. Cell Cycle 2010, 9, 4688–4702. [Google Scholar] [CrossRef] [Green Version]
- Shimada, M.; Nabeshima, K.; Tougan, T.; Nojima, H. The meiotic recombination checkpoint is regulated by checkpoint rad+ genes in fission yeast. EMBO J. 2002, 21, 2807–2818. [Google Scholar] [CrossRef] [Green Version]
- Perez-Hidalgo, L.; Moreno, S.; San-Segundo, P.A. The fission yeast meiotic checkpoint kinase Mek1 regulates nuclear localization of Cdc25 by phosphorylation. Cell Cycle 2008, 7, 3720–3730. [Google Scholar] [CrossRef]
- Perez-Hidalgo, L.; Moreno, S.; San-Segundo, P.A. Regulation of meiotic progression by the meiosis-specific checkpoint kinase Mek1 in fission yeast. J. Cell Sci. 2003, 116, 259–271. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, M.J.; Raleigh, J.M.; Verkade, H.M.; Nurse, P. Chk1 is a wee1 kinase in the G2 DNA damage checkpoint inhibiting cdc2 by Y15 phosphorylation. EMBO J. 1997, 16, 545–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furnari, B.; Rhind, N.; Russell, P. Cdc25 mitotic inducer targeted by chk1 DNA damage checkpoint kinase. Science 1997, 277, 1495–1497. [Google Scholar] [CrossRef]
- Moiseeva, V.; Amelina, H.; Collopy, L.C.; Armstrong, C.A.; Pearson, S.R.; Tomita, K. The telomere bouquet facilitates meiotic prophase progression and exit in fission yeast. Cell Discov. 2017, 3, 17041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami-Tonami, Y.; Ohtsuka, H.; Aiba, H.; Murakami, H. Regulation of wee1(+) expression during meiosis in fission yeast. Cell Cycle 2014, 13, 2853–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, J.M. Cyclin degradation: Don’t mes(s) with meiosis. Curr. Biol. 2005, 15, R461–R463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izawa, D.; Goto, M.; Yamashita, A.; Yamano, H.; Yamamoto, M. Fission yeast Mes1p ensures the onset of meiosis II by blocking degradation of cyclin Cdc13p. Nature 2005, 434, 529–533. [Google Scholar] [CrossRef]
- Kimata, Y.; Trickey, M.; Izawa, D.; Gannon, J.; Yamamoto, M.; Yamano, H. A mutual inhibition between APC/C and its substrate Mes1 required for meiotic progression in fission yeast. Dev. Cell 2008, 14, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Kitamura, K.; Fenner, N.; Yamano, H. Mes1 controls the meiosis I to meiosis II transition by distinctly regulating the anaphase-promoting complex/cyclosome coactivators Fzr1/Mfr1 and Slp1 in fission yeast. Mol. Biol. Cell 2011, 22, 1486–1494. [Google Scholar] [CrossRef]
- Viera, A.; Alsheimer, M.; Gomez, R.; Berenguer, I.; Ortega, S.; Symonds, C.E.; Santamaria, D.; Benavente, R.; Suja, J.A. CDK2 regulates nuclear envelope protein dynamics and telomere attachment in mouse meiotic prophase. J. Cell Sci. 2015, 128, 88–99. [Google Scholar] [CrossRef] [Green Version]
- Mikolcevic, P.; Isoda, M.; Shibuya, H.; del Barco Barrantes, I.; Igea, A.; Suja, J.A.; Shackleton, S.; Watanabe, Y.; Nebreda, A.R. Essential role of the Cdk2 activator RingoA in meiotic telomere tethering to the nuclear envelope. Nat. Commun. 2016, 7, 11084. [Google Scholar] [CrossRef] [Green Version]
- Tu, Z.; Bayazit, M.B.; Liu, H.; Zhang, J.; Busayavalasa, K.; Risal, S.; Shao, J.; Satyanarayana, A.; Coppola, V.; Tessarollo, L.; et al. Speedy A-Cdk2 binding mediates initial telomere-nuclear envelope attachment during meiotic prophase I independent of Cdk2 activation. Proc. Natl. Acad. Sci. USA 2017, 114, 592–597. [Google Scholar] [CrossRef] [Green Version]
- Palmer, N.; Talib, S.Z.A.; Kaldis, P. Diverse roles for CDK-associated activity during spermatogenesis. FEBS Lett. 2019, 593, 2925–2949. [Google Scholar] [CrossRef]
- Acosta, I.; Ontoso, D.; San-Segundo, P.A. The budding yeast polo-like kinase Cdc5 regulates the Ndt80 branch of the meiotic recombination checkpoint pathway. Mol. Biol. Cell 2011, 22, 3478–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mata, J.; Lyne, R.; Burns, G.; Bahler, J. The transcriptional program of meiosis and sporulation in fission yeast. Nat. Genet. 2002, 32, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Attner, M.A.; Amon, A. Control of the mitotic exit network during meiosis. Mol. Biol. Cell 2012, 23, 3122–3132. [Google Scholar] [CrossRef]
Figure 1.
Oscillation of cyclin-dependent kinase (CDK) activity and cyclins present in each stage of meiosis. In budding yeast, S phase CDK (S-CDK) activity peaks during premeiotic S phase and remains high until anaphase I. Following degradation of S phase cyclin, S-CDK activity increases again during metaphase II, followed by a decrease in anaphase II. In fission yeast, S-CDK activity peaks during premeiotic S phase, then declines following a decrease in Cig2 protein. In both budding yeast and fission yeast, M-CDK activity is low until entry into the nuclear divisions, during which M phase CDK (M-CDK) activity peaks at metaphase I and metaphase II. There is a highly regulated partial drop in M-CDK activity between meiosis I and meiosis II. For both yeast species, cyclins are listed under the stage in which they act. Cyclins in parenthesis have more minor or undetermined roles during these stages. Question mark after a cyclin indicates conflicting data about that cyclin’s role in the corresponding stage.
Figure 1.
Oscillation of cyclin-dependent kinase (CDK) activity and cyclins present in each stage of meiosis. In budding yeast, S phase CDK (S-CDK) activity peaks during premeiotic S phase and remains high until anaphase I. Following degradation of S phase cyclin, S-CDK activity increases again during metaphase II, followed by a decrease in anaphase II. In fission yeast, S-CDK activity peaks during premeiotic S phase, then declines following a decrease in Cig2 protein. In both budding yeast and fission yeast, M-CDK activity is low until entry into the nuclear divisions, during which M phase CDK (M-CDK) activity peaks at metaphase I and metaphase II. There is a highly regulated partial drop in M-CDK activity between meiosis I and meiosis II. For both yeast species, cyclins are listed under the stage in which they act. Cyclins in parenthesis have more minor or undetermined roles during these stages. Question mark after a cyclin indicates conflicting data about that cyclin’s role in the corresponding stage.

Figure 2.
Regulation of CDK activity in meiosis I. (A) At the metaphase I-to-anaphase. I transition in budding yeast meiosis Cdc14 is held inactive by Net1. APC/C-Cdc20 targets Clb5 for degradation while the APC/C co-activator Ama1 is inhibited by CDK-Clb1. Ime2 phosphorylates a subset of Cdk1 substrates. At anaphase I, Ama1 becomes active and APC/C-Ama1 targets Clb5 for further degradation. Clb1 and Clb4 are likely post-translationally modified (denoted with star) to downregulate CDK activity. Cdc14 activation is promoted by only the FEAR pathway, causing a partial but not full dephosphorylation of CDK substrates. Cdc14 reverses CDK, but not Ime2, phosphorylation on substrates. Combined, this regulation leads to a decline in CDK activity. (B) In fission yeast meiosis, Mes1 inhibits the APC/C. Mes1 is a competitive substrate of the APC/C co-activator Slp1 (Cdc20) and a pseudo-substrate of the co-activator Fzr1/Mfr1. Mes1 prevents full degradation of Cdc13 during meiosis I, thus allowing for retention of some CDK activity. Progression into anaphase I requires APC/C ubiquitination and proteasomal degradation of Mes1. Mes1 inhibits Fzr1/Mfr1 until meiosis II. The role of Clp1 (Cdc14) in meiosis is unclear, as deletion of clp1 has no effect on the timing of meiosis, but has a minor effect on the formation of four-spore asci.
Figure 2.
Regulation of CDK activity in meiosis I. (A) At the metaphase I-to-anaphase. I transition in budding yeast meiosis Cdc14 is held inactive by Net1. APC/C-Cdc20 targets Clb5 for degradation while the APC/C co-activator Ama1 is inhibited by CDK-Clb1. Ime2 phosphorylates a subset of Cdk1 substrates. At anaphase I, Ama1 becomes active and APC/C-Ama1 targets Clb5 for further degradation. Clb1 and Clb4 are likely post-translationally modified (denoted with star) to downregulate CDK activity. Cdc14 activation is promoted by only the FEAR pathway, causing a partial but not full dephosphorylation of CDK substrates. Cdc14 reverses CDK, but not Ime2, phosphorylation on substrates. Combined, this regulation leads to a decline in CDK activity. (B) In fission yeast meiosis, Mes1 inhibits the APC/C. Mes1 is a competitive substrate of the APC/C co-activator Slp1 (Cdc20) and a pseudo-substrate of the co-activator Fzr1/Mfr1. Mes1 prevents full degradation of Cdc13 during meiosis I, thus allowing for retention of some CDK activity. Progression into anaphase I requires APC/C ubiquitination and proteasomal degradation of Mes1. Mes1 inhibits Fzr1/Mfr1 until meiosis II. The role of Clp1 (Cdc14) in meiosis is unclear, as deletion of clp1 has no effect on the timing of meiosis, but has a minor effect on the formation of four-spore asci.


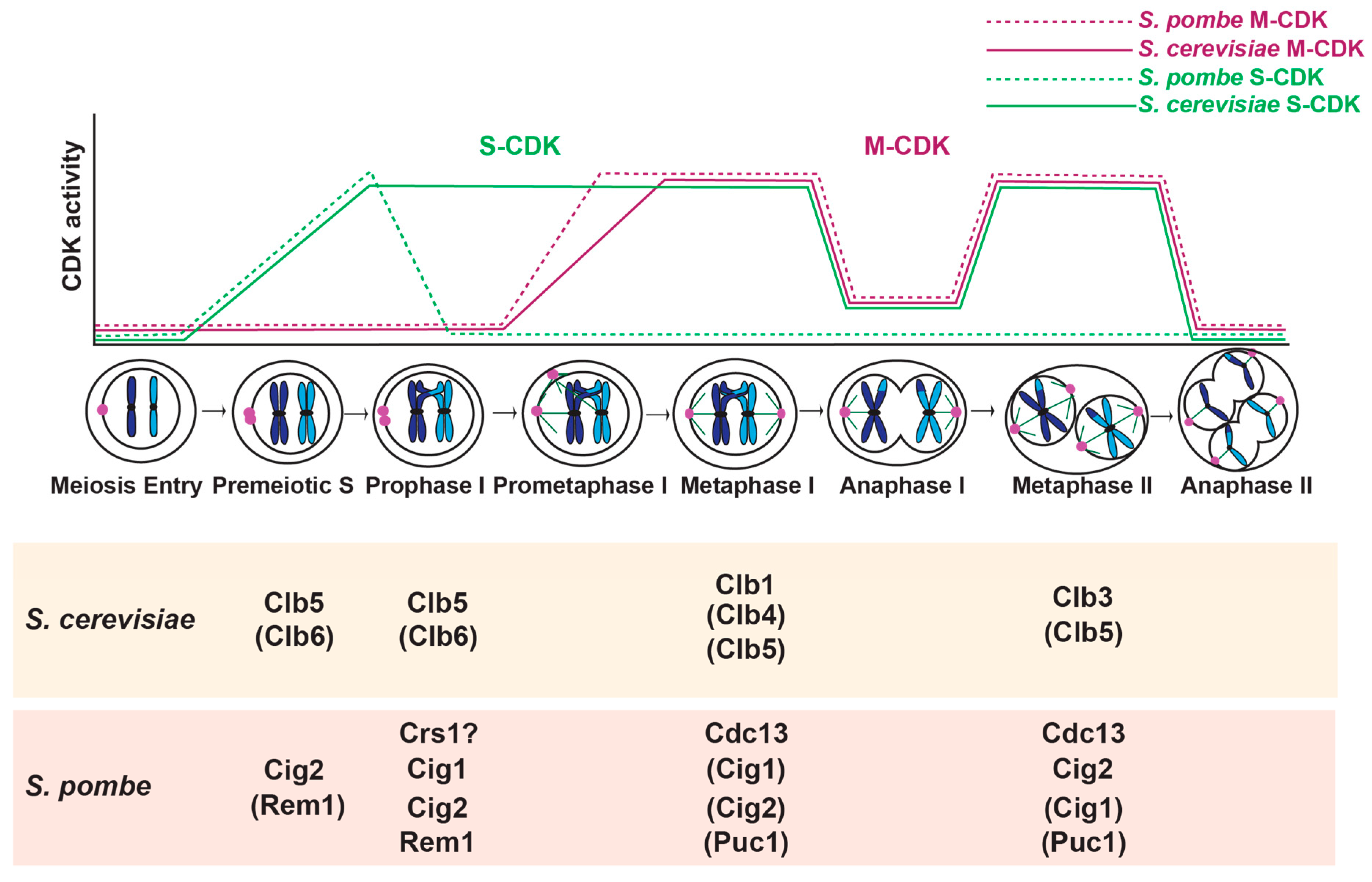{kind=link}
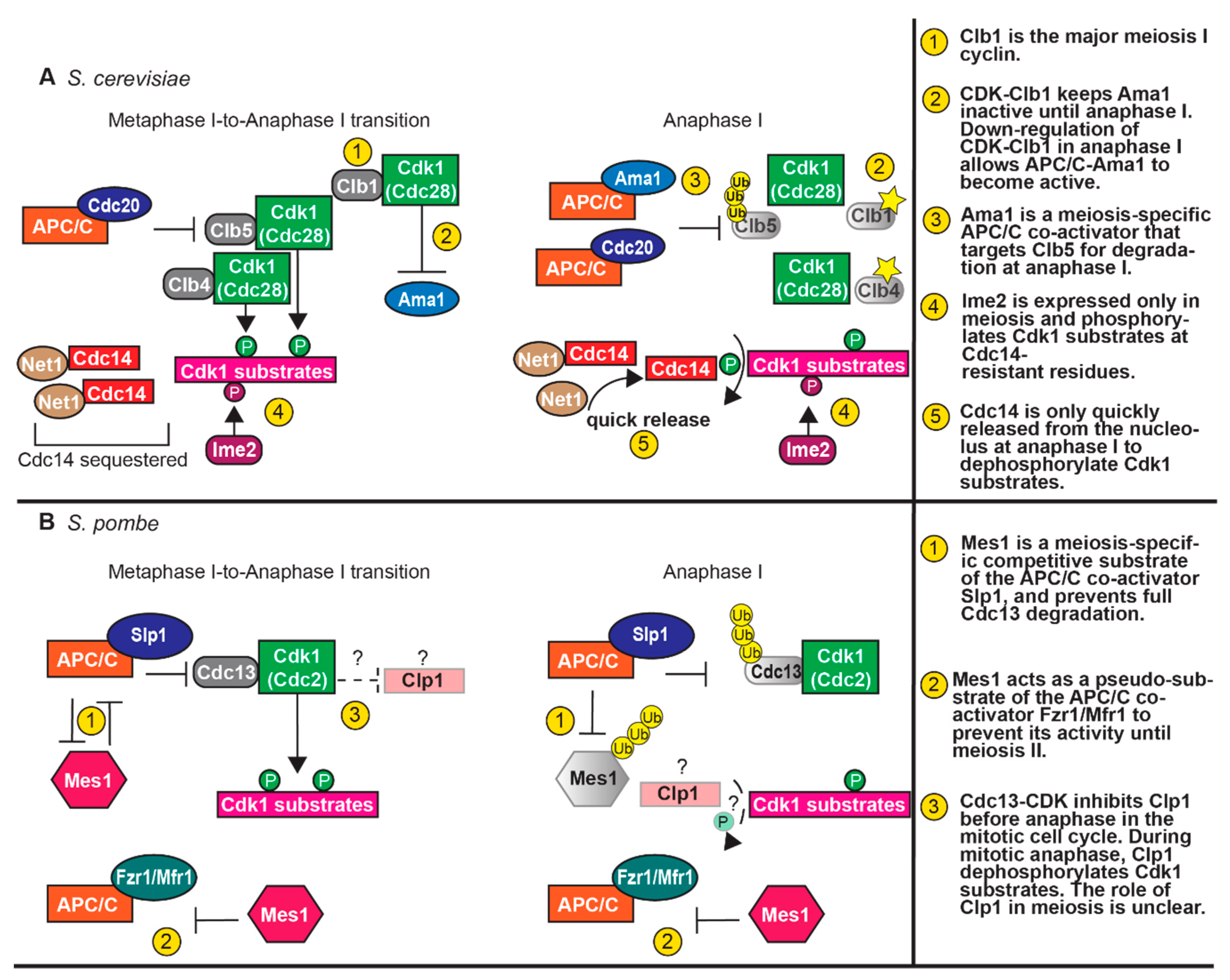{kind=link}
Table 1.
Anaphase Promoting Complex/Cyclosome (APC/C) Co-activators and their Cell Cycle Targets in Meiosis.
Table 1.
Anaphase Promoting Complex/Cyclosome (APC/C) Co-activators and their Cell Cycle Targets in Meiosis.
| APC/C Co-activator | Target(s) | Active Stage(s) of Meiosis | |
|---|---|---|---|
| S. cerevisiae | Cdc20 | Clb5 Pds1 | Meta I- to Ana I transition |
| Clb5 Pds1 Clb1 | Meta II- to Ana II transition | ||
| Ama1 | Ndd1 Clb1 Clb4 Cdc5 | Prophase I | |
| Clb5 Pds1 | Anaphase I | ||
| Ndt80 Cdc5 Clb3 | Anaphase II/Meiotic Exit | ||
| S. pombe | Slp1 | Cdc13 Cut2 Mes1 | Meta I- to Ana I transition |
| Cdc13 Cut2 | Meta II- to Ana II transition | ||
| Fzr1/Mfr1 | Cdc13 Cut2 | Meta II- to Ana II transition/ Anaphase II/Meiotic Exit |
Clb, Cyclin B; Pds1, Securin; Cdc5, Polo-like kinase; Cut2, Securin.
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
MacKenzie, A.M.; Lacefield, S. CDK Regulation of Meiosis: Lessons from S. cerevisiae and S. pombe. Genes 2020, 11, 723. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070723
AMA Style
MacKenzie AM, Lacefield S. CDK Regulation of Meiosis: Lessons from S. cerevisiae and S. pombe. Genes. 2020; 11(7):723. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070723
Chicago/Turabian StyleMacKenzie, Anne M., and Soni Lacefield. 2020. "CDK Regulation of Meiosis: Lessons from S. cerevisiae and S. pombe" Genes 11, no. 7: 723. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070723
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.