A New Intra-Specific and High-Resolution Genetic Map of Eggplant Based on a RIL Population, and Location of QTLs Related to Plant Anthocyanin Pigmentation and Seed Vigour

 ,
,  , ,
, ,  , , ,
, , ,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Library Construction and Sequencing
2.3. Sequence Analysis and Map Construction
2.4. Phenotypic Traits Evaluation
2.5. Statistical Analyses and QTL Detection
3. Results
3.1. Sequencing and Linkage Map Construction
3.2. Phenotypic Variation and Inter-Trait Correlations
3.3. QTL Analysis
3.3.1. QTL Affecting Plant Pigmentation in Eggplant
Adaxial Leaf Lamina Anthocyanin (adlan)
Stem Anthocyanins (stean)
Leaf Venation Anthocyanins (lvean)
Flower Anthocyanin Intensity (flian)
Anthocyanin Tonality (toan)
Hypocotyl Anthocyanins (hyan)
Corolla Colour (corcol)
3.3.2. QTL Affecting Speed of Plant Emergence Index
3.4. Candidate Genes Identification
sei2.1
sei2.2
sei4.1
stean10.1, lvean10.1, flian10.1, hyan10.1
corcol10.1-toan10.1
adlan10.1
stean5.1, toan5.2
toan5.1-corcol5.1-hyan5.1
hyan7.1
stean2.1
4. Discussion
4.1. Genetic Map Construction and Phenotyping
4.2. QTLs and Underlying Candidate Genes
4.2.1. Seed Emergency Index
4.2.2. Anthocyanins
4.2.3. Anthocyanin Related Candidate Genes Identifications
Cluster on Chromosome E10
QTL cluster on Chromosome E05
QTLs for hyan7.1 and stean2.1
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Fukuoka, H.; Yamaguchi, H.; Nunome, T.; Negoro, S.; Miyatake, K.; Ohyama, A. Accumulation, functional annotation, and comparative analysis of expressed sequence tags in eggplant (Solanum melongena L.), the third pole of the genus Solanum species after tomato and potato. Gene 2010, 450, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Hirakawa, H.; Shirasawa, K.; Miyatake, K.; Nunome, T.; Negoro, S.; Ohyama, A.; Yamaguchi, H.; Sato, S.; Isobe, S.; Tabata, S.; et al. Draft genome sequence of eggplant (Solanum melongena L.): The representative solanum species indigenous to the old world. DNA Res. Int. J. Rapid Publ. Rep. Genes Genomes 2014, 21, 649–660. [Google Scholar] [CrossRef] [Green Version]
- Albert, V.A.; Chang, T.H. Evolution of a hot genome. Proc. Natl. Acad. Sci. USA 2014, 111, 5069–5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lester, R.N.; Hasan, S.M.Z. Origin and Domestication of the Brinjal Eggplant, Solanum Melongena, from S. incanum, in Africa and Asia. In Solanaceae III: Taxonomy, Chemistry, and Evolution; Hawkes, J.G., Lester, R.N., Nee, M., Estrada, N., Eds.; Royal Botanic Gardens: Kew, UK, 1991; pp. 369–387. [Google Scholar]
- Meyer, R.S.; Karol, K.G.; Little, D.P.; Nee, M.H.; Litt, A. Phylogeographic relationships among Asian eggplants and new perspectives on eggplant domestication. Mol. Phylogenetics Evol. 2012, 63, 685–701. [Google Scholar] [CrossRef] [PubMed]
- Cericola, F.; Portis, E.; Toppino, L.; Barchi, L.; Acciarri, N.; Ciriaci, T.; Sala, T.; Rotino, G.L.G.L.; Lanteri, S. The Population Structure and diversity of eggplant from Asia and the Mediterranean Basin. PLoS ONE 2013, 8, e73702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.; Gibson, J.; Meyer, R.S.; Chapman, M.A. Eggplant domestication: Pervasive gene flow, feralization, and transcriptomic divergence. Mol. Biol. Evol. 2019, 36, 1359–1372. [Google Scholar] [CrossRef]
- FAO. Available online: http://faostat3.fao.org/home/E.org/ (accessed on 1 June 2020).
- Zhang, Y.; Liu, H.; Shen, S.; Zhang, X. Improvement of eggplant seed germination and seedling emergence at low temperature by seed priming with incorporation SA into KNO 3 solution. Front. Agric. China 2011, 5, 534–537. [Google Scholar] [CrossRef]
- Gisbert, C.; Prohens, J.; Nuez, F. Treatments for improving seed germination in eggplant and related species. Acta Hortic. 2009, 898, 45–51. [Google Scholar] [CrossRef] [Green Version]
- Forti, C.; Ottobrino, V.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Pagano, A.; Macovei, A.; Balestrazzi, A. Molecular dynamics of pre-germinative metabolism in primed eggplant (Solanum melongena L.) seeds. Hortic. Res. 2020, 7, 87. [Google Scholar] [CrossRef]
- Doganlar, S.; Frary, A.; Daunay, M.C.; Lester, R.N.; Tanksley, S.D. A comparative genetic linkage map of eggplant (Solanum melongena) and its implications for genome evolution in the Solanaceae. Genetics 2002, 161, 1697–1711. [Google Scholar]
- Doğanlar, S.; Frary, A.; Daunay, M.-C.; Huvenaars, K.; Mank, R.; Frary, A. High resolution map of eggplant (Solanum melongena) reveals extensive chromosome rearrangement in domesticated members of the Solanaceae. Euphytica 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.N.; Eannetta, N.T.; Xu, Y.M.; Tanksley, S.D. A detailed synteny map of the eggplant genome based on conserved ortholog set II (COSII) markers. Theor. Appl. Genet. 2009, 118, 927–935. [Google Scholar] [CrossRef]
- Frary, A.; Doganlar, S.; Daunay, M.C.; Tanksley, S.D. QTL analysis of morphological traits in eggplant and implications for conservation of gene function during evolution of solanaceous species. Theor. Appl. Genet. 2003, 107, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Frary, A.; Frary, A.; Daunay, M.-C.; Huvenaars, K.; Mank, R.; Doğanlar, S. QTL hotspots in eggplant (Solanum melongena) detected with a high resolution map and CIM analysis. Euphytica 2014, 197, 211–228. [Google Scholar] [CrossRef] [Green Version]
- Gramazio, P.; Prohens, J.; Plazas, M.; Andújar, I.; Herraiz, F.J.; Castillo, E.; Knapp, S.; Meyer, R.S.; Vilanova, S. Location of chlorogenic acid biosynthesis pathway and polyphenol oxidase genes in a new interspecific anchored linkage map of eggplant. BMC Plant Biol. 2014, 14, 350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunseri, F.; Sciancalepore, A.; Martelli, G.; Acciarri, N.; Rotino, G.L.; Valentino, D.; Tamietti, G. Development of RAPD-AFLP Map of Eggplant and Improvement of Tolerance to Verticillium Wilt. In Proceedings of the Acta Horticulturae, Leuven, Belgium, 30 September 2003; pp. 107–115. [Google Scholar]
- Nunome, T.; Ishiguro, K.; Yoshida, T.; Hirai, M. Mapping of fruit shape and color development traits in eggplant (Solanum melongena L.) based on RAPD and AFLP markers. Breed. Sci. 2001, 51, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Nunome, T.; Suwabe, K.; Iketani, H.; Hirai, M. Identification and characterization of microsatellites in eggplant. Plant Breed. 2003, 122, 256–262. [Google Scholar] [CrossRef]
- Nunome, T.; Negoro, S.; Kono, I.; Kanamori, H.; Miyatake, K.; Yamaguchi, H.; Ohyama, A.; Fukuoka, H. Development of SSR markers derived from SSR-enriched genomic library of eggplant (Solanum melongena L.). Theor. Appl. Genet. 2009, 119, 1143–1153. [Google Scholar] [CrossRef]
- Barchi, L.; Lanteri, S.; Portis, E.; Stàgel, A.; Valè, G.; Toppino, L.; Leonardo Rotino, G. Segregation distortion and linkage analysis in eggplant (Solanum melongena L.). Genome 2010, 53. [Google Scholar] [CrossRef]
- Fukuoka, H.; Miyatake, K.; Nunome, T.; Negoro, S.; Shirasawa, K.; Isobe, S.; Asamizu, E.; Yamaguchi, H.; Ohyama, A. Development of gene-based markers and construction of an integrated linkage map in eggplant by using Solanum orthologous (SOL) gene sets. Theor. Appl. Genet. 2012, 125, 47–56. [Google Scholar] [CrossRef]
- Miyatake, K.; Saito, T.; Negoro, S.; Yamaguchi, H.; Nunome, T.; Ohyama, A.; Fukuoka, H. Development of selective markers linked to a major QTL for parthenocarpy in eggplant (Solanum melongena L.). Theor. Appl. Genet. 2012, 124, 1–11. [Google Scholar] [CrossRef]
- Miyatake, K.; Saito, T.; Negoro, S.; Yamaguchi, H.; Nunome, T.; Ohyama, A.; Fukuoka, H. Detailed mapping of a resistance locus against Fusarium wilt in cultivated eggplant (Solanum melongena). Theor. Appl. 2016, 129, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Lanteri, S.; Portis, E.; Acquadro, A.; Valè, G.; Toppino, L.; Rotino, G.L.G.L.L.G.L.; Vale, G.; Toppino, L.; Rotino, G.L.G.L.L.G.L.; et al. Identification of SNP and SSR markers in eggplant using RAD tag sequencing. BMC Genom. 2011, 12. [Google Scholar] [CrossRef] [Green Version]
- Baird, N.A.; Etter, P.D.; Atwood, T.S.; Currey, M.C.; Shiver, A.L.; Lewis, Z.A.; Selker, E.U.; Cresko, W.A.; Johnson, E.A. Rapid SNP Discovery and Genetic Mapping Using Sequenced RAD Markers. PLoS ONE 2008, 3, e3376. [Google Scholar] [CrossRef] [PubMed]
- Barchi, L.; Lanteri, S.; Portis, E.; Valè, G.; Volante, A.; Pulcini, L.; Ciriaci, T.; Acciarri, N.; Barbierato, V.; Toppino, L.; et al. A RAD tag derived marker based eggplant linkage map and the location of QTLs determining anthocyanin pigmentation. PLoS ONE 2012, 7, e43740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portis, E.; Barchi, L.; Toppino, L.; Lanteri, S.; Acciarri, N.; Felicioni, N.; Fusari, F.; Barbierato, V.; Cericola, F.; Valè, G.; et al. QTL mapping in eggplant reveals clusters of yield-related loci and orthology with the tomato genome. PLoS ONE 2014, 9, e89499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toppino, L.; Barchi, L.; Lo Scalzo, R.; Palazzolo, E.; Francese, G.; Fibiani, M.; D’ Alessandro, A.; Papa, V.; Laudicina, V.A.; Sabatino, L.; et al. Mapping Quantitative Trait Loci Affecting Biochemical and Morphological Fruit Properties in Eggplant (Solanum melongena L.). Front. Plant Sci. 2016. [Google Scholar] [CrossRef] [Green Version]
- Barchi, L.; Toppino, L.; Valentino, D.; Bassolino, L.; Portis, E.; Lanteri, S.; Rotino, G.L. QTL analysis reveals new eggplant loci involved in resistance to fungal wilts. Euphytica 2018, 214, 20. [Google Scholar] [CrossRef]
- Cericola, F.; Portis, E.; Lanteri, S.; Toppino, L.; Barchi, L.; Acciarri, N.; Pulcini, L.; Sala, T.; Rotino, G.L. Linkage disequilibrium and genome-wide association analysis for anthocyanin pigmentation and fruit color in eggplant. BMC Genom. 2014, 15, 896. [Google Scholar] [CrossRef] [Green Version]
- Portis, E.; Cericola, F.; Barchi, L.; Toppino, L.; Acciarri, N.; Pulcini, L.; Sala, T.; Lanteri, S.; Rotino, G.L. Association mapping for fruit, plant and leaf morphology traits in eggplant. PLoS ONE 2015, 10, e0135200. [Google Scholar] [CrossRef] [Green Version]
- Salgon, S.; Raynal, M.; Lebon, S.; Baptiste, J.M.; Daunay, M.C.; Dintinger, J.; Jourda, C. Genotyping by sequencing highlights a polygenic resistance to ralstonia pseudosolanacearum in eggplant (Solanum melongena L.). Int. J. Mol. Sci. 2018, 19, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, Q.; Wang, W.; Hu, T.; Hu, H.; Wang, J.; Bao, C. Construction of a SNP-based genetic map using SLAF-Seq and QTL analysis of morphological traits in eggplant. Front. Genet. 2020, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Barchi, L.; Pietrella, M.; Venturini, L.; Minio, A.; Toppino, L.; Acquadro, A.; Andolfo, G.; Aprea, G.; Avanzato, C.; Bassolino, L.; et al. A chromosome-anchored eggplant genome sequence reveals key events in Solanaceae evolution. Sci. Rep. 2019, 9, 11769. [Google Scholar] [CrossRef] [PubMed]
- Sulli, M.; Barchi, L.; Francese, G.; Toppino, L.; Diretto, G.; Mennella, G.; Lanteri, S.; Rotino, G.; Giuliano, G. Metabolic Profiling of a Recombinant Inbred Eggplant Population Reveals Key Metabolic QTLs controlling Fruit Nutritional Quality. In Proceedings of the Plant & Animal Genome XXVII Conference (pe0948), San Diego, CA, USA, 12–16 January 2019. [Google Scholar]
- Van Eck, H.J.; Jacobs, J.M.E.; Dijk, J.; Stiekema, W.J.; Jacobsen, E. Identification and mapping of three flower colour loci of potato (S. tuberosum L.) by RFLP analysis. Theor. Appl. Genet. 1993, 86, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Chaim, A.B.C.; Borovsky, Y.B.; De Jong, W.D.J.; Paran, I.P. Linkage of the A locus for the presence of anthocyanin and fs10.1, a major fruit-shape QTL in pepper. Theor. Appl. Genet. 2003, 106, 889–894. [Google Scholar] [CrossRef]
- Van Eck, H.J.; Jacobs, J.M.E.; van den Berg, P.M.M.M.; Stiekema, W.J.; Jacobsen, E. The inheritance of anthocyanin pigmentation in potato (Solanum tuberosum L.) and mapping of tuber skin colour loci using RFLPs. Heredity 1994, 73, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Borovsky, Y.; Oren-Shamir, M.; Ovadia, R.; De Jong, W.; Paran, I. The A locus that controls anthocyanin accumulation in pepper encodes a MYB transcription factor homologous to Anthocyanin2 of Petunia. Theor. Appl. Genet. 2004, 109, 23–29. [Google Scholar] [CrossRef]
- Bovy, A.; Schijlen, E.; Hall, R. Metabolic engineering of flavonoids in tomato (Solanum lycopersicum): The potential for metabolomics. Metabolomics 2007, 3, 399–412. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Mazzucato, A.; Perata, P. Purple as a tomato: Towards high anthocyanin tomatoes. Trends Plant Sci. 2009, 14, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Ge, H.; Liu, Y.; Jiang, M.; Zhang, J.; Han, H.; Chen, H. Analysis of genetic diversity and structure of eggplant populations (Solanum melongena L.) in China using simple sequence repeat markers. Sci. Hortic. 2013, 162, 71–75. [Google Scholar] [CrossRef]
- Toppino, L.; Vale, G.; Rotino, G.L. Inheritance of Fusarium wilt resistance introgressed from Solanum aethiopicum Gilo and Aculeatum groups into cultivated eggplant (S.melongena) and development of associated PCR-based markers. Mol. Breed. 2008, 22, 237–250. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–14. [Google Scholar]
- Acquadro, A.; Lanteri, S.; Scaglione, D.; Arens, P.; Vosman, B.; Portis, E. Genetic mapping and annotation of genomic microsatellites isolated from globe artichoke. Theor. Appl. Genet. 2009, 118, 1573–1587. [Google Scholar] [CrossRef]
- Acquadro, A.; Barchi, L.; Gramazio, P.; Portis, E.; Vilanova, S.; Comino, C.; Plazas, M.; Prohens, J.; Lanteri, S. Coding SNPs analysis highlights genetic relationships and evolution pattern in eggplant complexes. PLoS ONE 2017, 12, e0180774. [Google Scholar] [CrossRef] [PubMed]
- Li, H. Aligning Sequence Reads, Clone Sequences and Assembly Contigs with BWA-MEM. arXiv 2013, arXiv:1303.3997. [Google Scholar]
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinform. 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [Green Version]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A.; Sen, Ś.; Churchill, G.A.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]
- Van Ooijen, J.W. JoinMap ® 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2006. [Google Scholar]
- Van Os, H.; Stam, P.; Visser, R.G.F.; van Eck, H.J. SMOOTH: A statistical method for successful removal of genotyping errors from high-density genetic linkage data. Theor. Appl. Genet. 2005, 112, 187–194. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing. 2020. Available online: https://www.r-project.org/ (accessed on 15 May 2020).
- Jansen, R.C. Interval mapping of multiple quantitative trait loci. Genetics 1993, 135, 205–211. [Google Scholar]
- Jansen, R.C.; Stam, P. High-resolution of quantitative traits into multiple loci via interval mapping. Genetics 1994, 136, 1447–1455. [Google Scholar] [PubMed]
- Van Ooijen, J.W. MapQTL 5, Software for the Mapping of Quantitative Trait Loci in Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2004; Volume 63. [Google Scholar]
- Churchill, G.A.; Do erge, R.W. Empirical Threshold Values for Quantitative Trait Mapping. Genetics 1994, 138, 963–971. [Google Scholar] [PubMed]
- CMplot. Available online: https://github.com/YinLiLin/R-CMplot (accessed on 15 June 2020).
- Stàgel, A.; Portis, E.; Toppino, L.; Rotino, G.L.; Lanteri, S. Gene-based microsatellite development for mapping and phylogeny studies in eggplant. BMC Genom. 2008, 9, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchi, L.; Acquadro, A.; Alonso, D.; Aprea, G.; Bassolino, L.; Demurtas, O.; Ferrante, P.; Gramazio, P.; Mini, P.; Portis, E.; et al. Single Primer Enrichment Technology (SPET) for high-throughput genotyping in tomato and eggplant germplasm. Front. Plant Sci. 2019, 10, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barchi, L.; Portis, E.; Toppino, L.; Rotino, G.L. Molecular Mapping, QTL Identification, and GWA Analysis. In The Eggplant Genome; Springer International Publishing: Cham, Switzerland, 2019; pp. 41–54. [Google Scholar]
- Acquadro, A.; Barchi, L.; Portis, E.; Mangino, G.; Valentino, D.; Mauromicale, G.; Lanteri, S. Genome reconstruction in Cynara cardunculus taxa gains access to chromosome-scale DNA variation. Sci. Rep. 2017, 7, 5617. [Google Scholar] [CrossRef]
- Zhang, X.F.; Wang, G.Y.; Dong, T.T.; Chen, B.; Du, H.S.; Li, C.B.; Zhang, F.L.; Zhang, H.Y.; Xu, Y.; Wang, Q.; et al. High-density genetic map construction and QTL mapping of first flower node in pepper (Capsicum annuum L.). BMC Plant Biol. 2019, 19, 167. [Google Scholar] [CrossRef]
- Hu, X.H.; Zhang, S.Z.; Miao, H.R.; Cui, F.G.; Shen, Y.; Yang, W.Q.; Xu, T.T.; Chen, N.; Chi, X.Y.; Zhang, Z.M.; et al. High-density genetic map construction and identification of QTLs controlling oleic and linoleic acid in peanut using SLAF-seq and SSRs. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef]
- Collard, B.; Jahufer, M.; Brouwer, J.; Pang, E. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Li, Z.; Jakkula, L.; Hussey, R.S.; Tamulonis, J.P.; Boerma, H.R. SSR mapping and confirmation of the QTL from PI96354 conditioning soybean resistance to southern root-knot nematode. Theor. Appl. Genet. 2001, 103, 1167–1173. [Google Scholar] [CrossRef]
- Lindhout, P. The perspectives of polygenic resistance in breeding for durable disease resistance. Euphytica 2002, 124, 217–226. [Google Scholar] [CrossRef]
- Pilet-Nayel, M.P.-N.; Muehlbauer, F.M.; McGee, R.M.; Kraft, J.K.; Baranger, A.B.; Coyne, C.C. Quantitative trait loci for partial resistance to Aphanomyces root rot in pea. Theor. Appl. Genet. 2002, 106, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Botstein, D. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics 1989, 121, 185–199. [Google Scholar] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Müller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [Green Version]
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed Germination and Vigor. Annu. Rev. Plant Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [Green Version]
- Adebola, P.O.; Afolayan, A.J. Germination responses of Solanum aculeastrum, a medicinal species of the Eastern Cape, South Africa. Seed Sci. Technol. 2006, 34, 735–740. [Google Scholar] [CrossRef]
- Demir, I.; Ermis, S.; Okçu, G.; Matthews, S. Vigour tests for predicting seedling emergence of aubergine (Solanum melongena L.) seed lots. Seed Sci. Technol. 2005, 33, 481–484. [Google Scholar] [CrossRef]
- Joshua, A. Seed germination of Solanum incanum: An example of germination programs of tropical vegetable crops. Acta Hortic. 1977, 83, 155–162. [Google Scholar] [CrossRef]
- Ibrahim, M.; Munira, M.K.; Kabir, M.S.; Islam, A.K.M.S.; Miah, M.M.U. Seed germination and graft compatibility of wild solanum as rootstock of tomato. J. Biol. Sci. 2001, 1, 701–703. [Google Scholar] [CrossRef] [Green Version]
- Taab, A.; Andersson, L. Seed dormancy dynamics and germination characteristics of Solanum nigrum. Weed Res. 2009, 49, 490–498. [Google Scholar] [CrossRef]
- Wang, Z.; Cao, H.; Sun, Y.; Li, X.; Chen, F.; Carles, A.; Li, Y.; Ding, M.; Zhang, C.; Deng, X.; et al. Arabidopsis paired amphipathic helix proteins SNL1 and SNL2 redundantly regulate primary seed dormancy via abscisic acid-ethylene antagonism mediated by histone deacetylation. Plant Cell 2013, 25, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Song, W.M.; Pan, J.; Jiang, C.M.; Srivastava, R.; Li, B.; Zhu, L.Y.; Su, H.Y.; Gao, X.S.; Liu, H.; et al. Overexpression of the NDR1/HIN1-Like gene NHL6 modifies seed germination in response to abscisic acid and abiotic stresses in Arabidopsis. PLoS ONE 2016, 11. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, J. Membrane bound: C2-domain abscisic acid-related proteins help abscisic acid receptors get where they need to go. Plant Cell 2014, 26, 4566. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Hernández, A.A.; Ortega-Amaro, M.A.; Delgado-Sánchez, P.; Salinas, J.; Jiménez-Bremont, J.F. AtGRDP1 Gene Encoding a Glycine-Rich domain protein is involved in germination and responds to ABA Signalling. Plant Mol. Biol. Report. 2014, 32, 1187–1202. [Google Scholar] [CrossRef]
- Arc, E.; Chibani, K.; Grappin, P.; Jullien, M.; Godin, B.; Cueff, G.; Valot, B.; Balliau, T.; Job, D.; Rajjou, L. Cold stratification and exogenous nitrates entail similar functional proteome adjustments during Arabidopsis seed dormancy release. J. Proteome Res. 2012, 11, 5418–5432. [Google Scholar] [CrossRef]
- Chatelain, E.; Hundertmark, M.; Leprince, O.; Gall, S.L.; Satour, P.; Deligny-Penninck, S.; Rogniaux, H.; Buitink, J. Temporal profiling of the heat-stable proteome during late maturation of Medicago truncatula seeds identifies a restricted subset of late embryogenesis abundant proteins associated with longevity. Plant Cell Environ. 2012, 35, 1440–1455. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Davydov, O.; Fluhr, R. Plant serpin protease inhibitors: Specificity and duality of function. J. Exp. Bot. 2019, 70, 2077–2085. [Google Scholar] [CrossRef]
- Tsay, Y.F.; Chiu, C.C.; Tsai, C.B.; Ho, C.H.; Hsu, P.K. Nitrate transporters and peptide transporters. FEBS Lett. 2007, 581, 2290–2300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, C.H.; Lin, S.H.; Hu, H.C.; Tsay, Y.F. CHL1 functions as a nitrate sensor in plants. Cell 2009, 138, 1184–1194. [Google Scholar] [CrossRef] [Green Version]
- Miranda, M.; Borisjuk, L.; Tewes, A.; Dietrich, D.; Rentsch, D.; Weber, H.; Wobus, U. Peptide and amino acid transporters are differentially regulated during seed development and germination in faba bean. Plant Physiol. 2003, 132, 1950–1960. [Google Scholar] [CrossRef] [Green Version]
- Footitt, S.; Huang, Z.; Clay, H.A.; Mead, A.; Finch-Savage, W.E. Temperature, light and nitrate sensing coordinate Arabidopsis seed dormancy cycling, resulting in winter and summer annual phenotypes. Plant J. 2013, 74, 1003–1015. [Google Scholar] [CrossRef] [PubMed]
- Osuna, D.; Prieto, P.; Aguilar, M. Control of seed germination and plant development by carbon and nitrogen availability. Front. Plant Sci. 2015, 6, 1023. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [Green Version]
- Alboresi, A.; Gestin, C.; Leydecker, M.T.; Bedu, M.; Meyer, C.; Truong, H.N. Nitrate, a signal relieving seed dormancy in Arabidopsis. Plant Cell Environ. 2005, 28, 500–512. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Hou, X.; Fang, J.; Wei, P.; Xu, B.; Chen, M.; Feng, Y.; Chu, C. The rice GERMINATION DEFECTIVE 1, encoding a B3 domain transcriptional repressor, regulates seed germination and seedling development by integrating GA and carbohydrate metabolism. Plant J. 2013, 75, 403–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Li, M.; He, D.; Wang, K.; Yang, P. Mutations on ent-kaurene oxidase 1 encoding gene attenuate its enzyme activity of catalyzing the reaction from ent-kaurene to ent-kaurenoic acid and lead to delayed germination in rice. PLoS Genet. 2020, 16, e1008562. [Google Scholar] [CrossRef]
- Harborne, J.B.; Williams, C.A. Advances in flavonoid research since 1992. Phytochemistry 2000, 55, 481–504. [Google Scholar] [CrossRef]
- Schemske, D.W.; Bradshaw, H.D. Pollinator preference and the evolution of floral traits in monkeyflowers (Mimulus). Proc. Natl. Acad. Sci. USA 1999, 96, 11910–11915. [Google Scholar] [CrossRef] [Green Version]
- Close, D.C.; Beadle, C.L. The Ecophysiology of foliar anthocyanin. Bot. Rev. 2003, 69, 149–161. [Google Scholar] [CrossRef]
- Gross, J. Pigments in Vegetables: Chlorophylls and Carotenoids; Springer Science & Business Media: New York, NY, USA, 2012; ISBN 1461520339. [Google Scholar]
- De Jong, W.S.; Eannetta, N.T.; Jong, D.M.; Bodis, M. Candidate gene analysis of anthocyanin pigmentation loci in the Solanaceae. Theor. Appl. Genet. 2004, 108, 423–432. [Google Scholar] [CrossRef]
- Stommel, J.R.; Dumm, J.M. Coordinated regulation of biosynthetic and regulatory genes coincides with anthocyanin accumulation in developing eggplant fruit. J. Amer. Soc. Hort. Sci. 2015, 140, 129–135. [Google Scholar] [CrossRef]
- Tatebe, T. On inheritance of color in Solanum melongena L. Jpn. J. Genet. 1939, 15, 261–271. [Google Scholar] [CrossRef] [Green Version]
- Tigchelaar, E.C.; Janick, J.; Erickson, H.T. The genetics of anthocyanin coloration in eggplant (Solanum melongena L.). Genetics 1968, 60, 475–491. [Google Scholar] [PubMed]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables: A review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef]
- Docimo, T.; Francese, G.; Ruggiero, A.; Batelli, G.; de Palma, M.; Bassolino, L.; Toppino, L.; Rotino, G.L.; Mennella, G.; Tucci, M. phenylpropanoids accumulation in eggplant fruit: Characterization of biosynthetic genes and regulation by a MYB transcription factor. Front. Plant Sci. 2015, 6, 1233. [Google Scholar] [CrossRef] [Green Version]
- Moglia, A.; Florio, F.E.; Iacopino, S.; Guerrieri, A.; Milani, A.M.; Comino, C.; Barchi, L.; Marengo, A.; Cagliero, C.; Rubiolo, P.; et al. Identification of a new R3 MYB type repressor and functional characterization of the members of the MBW transcriptional complex involved in anthocyanin biosynthesis in eggplant (S. melongena L.). PLoS ONE 2020, 15, e0232986. [Google Scholar] [CrossRef]
- Zhang, Y.; Hu, Z.; Chu, G.; Huang, C.; Tian, S.; Zhao, Z.; Chen, G. Anthocyanin accumulation and molecular analysis of anthocyanin biosynthesis-associated genes in eggplant (Solanum melongena L.). J. Agric. Food Chem. 2014, 62, 2906–2912. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.O.; Lin, W.Q.; Li, K.; Feng, X.F.; Jin, H.; Zou, H. Transcriptome analyses reveal anthocyanin biosynthesis in eggplants. PeerJ Prepr. 2018, 6, e27289v1. [Google Scholar]
- Saito, K.; Kobayashi, M.; Gong, Z.; Tanaka, Y.; Yamazaki, M. Direct evidence for anthocyanidin synthase as a 2-oxoglutarate-dependent oxygenase: Molecular cloning and functional expression of cDNA from a red forma of Perilla frutescens. Plant J. 1999, 17, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Hirsch, C.D.; Springer, N.M. Transposable element influences on gene expression in plants. Biochim. Et Biophys. Acta—Gene Regul. Mech. 2017, 1860, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.E.; Torabinejad, J.; Cohick, E.; Parker, K.; Drake, E.J.; Thompson, J.E.; Hortter, M.; DeWald, D.B. Mutations in the Arabidopsis phosphoinositide phosphatase gene SAC9 lead to overaccumulation of PtdIns(4,5)P2 and constitutive expression of the stress-response pathway. Plant Physiol. 2005, 138, 686–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.; Ho Shin, D.; Cho, M.-H.; Kim, T.-L.; Hee Bhoo, S.; Hahn, T.-R. An ankyrin repeat protein is involved in anthocyanin biosynthesis in Arabidopsis. Physiol. Plant. 2011, 142, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.F.; Zhao, Y.; Jiang, J.P.; Guan, W.X.; Du, J.F. Phosphatase Wip1 in immunity: An overview and update. Front. Immunol. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, H.; Li, W.; Zhang, X.; Deng, S.; Xu, Q.; Hou, T.; Pang, X.; Zhang, Z.; Zhang, X. In planta high levels of hydrolysable tannins inhibit peroxidase mediated anthocyanin degradation and maintain abaxially red leaves of Excoecaria Cochinchinensis. BMC Plant Biol. 2019, 19, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Fang, F.; Zhang, X.L.; Luo, H.H.; Zhou, J.J.; Gong, Y.H.; Li, W.J.; Shi, Z.W.; He, Q.; Wu, Q.; Li, L.; et al. An intracellular Laccase is responsible for epicatechin-mediated Anthocyanin degradation in litchi fruit Pericarp. Plant Physiol. 2015, 169, 2391–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zipor, G.; Duarte, P.; Carqueijeiro, I.; Shahar, L.; Ovadia, R.; Teper-Bamnolker, P.; Eshel, D.; Levin, Y.; Doron-Faigenboim, A.; Sottomayor, M.; et al. In planta anthocyanin degradation by a vacuolar class III peroxidase in Brunfelsia calycina flowers. New Phytol. 2015, 205, 653–665. [Google Scholar] [CrossRef]
- Sytar, O.; Zivcak, M.; Bruckova, K.; Brestic, M.; Hemmerich, I.; Rauh, C.; Simko, I. Shift in accumulation of flavonoids and phenolic acids in lettuce attributable to changes in ultraviolet radiation and temperature. Sci. Hortic. 2018, 239, 193–204. [Google Scholar] [CrossRef]
- Ino, I.; Yamaguchi, M.A. Acetyl-coenzyme A: Anthocyanidin 3-glucoside acetyltransferase from flowers of Zinnia elegans. Phytochemistry 1993. [Google Scholar] [CrossRef]
- Chong, J.; Baltz, R.; Schmitt, C.; Beffa, R.; Fritig, B.; Saindrenan, P. Downregulation of a pathogen-responsive tobacco UDP-Glc:phenylpropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virus resistance. Plant Cell 2002. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Brugliera, F. Flower colour and cytochromes P450. Philos. Trans. R. Soc. B Biol. Sci. 2013, 368, 20120432. [Google Scholar] [CrossRef] [Green Version]
- Su, V.; Hsu, B.D. Transient expression of the cytochrome p450 CYP78A2 enhances anthocyanin production in flowers. Plant Mol. Biol. Report. 2010. [Google Scholar] [CrossRef]
- Höll, J.; Vannozzi, A.; Czemmel, S.; Donofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogsa, J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis in Vitis vinifera. Plant Cell 2013. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.B.; McNear, D.H. Induced transcriptional profiling of phenylpropanoid pathway genes increased flavonoid and lignin content in Arabidopsis leaves in response to microbial products. BMC Plant Biol. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albert, N.W. Subspecialization of R2R3-MYB repressors for anthocyanin and proanthocyanidin regulation in forage legumes. Front. Plant Sci. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Gou, M.; Liu, C.J. Arabidopsis kelch repeat F-Box proteins regulate phenylpropanoid biosynthesis via controlling the turnover of phenylalanine ammonia-lyase. Plant Cell 2013. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
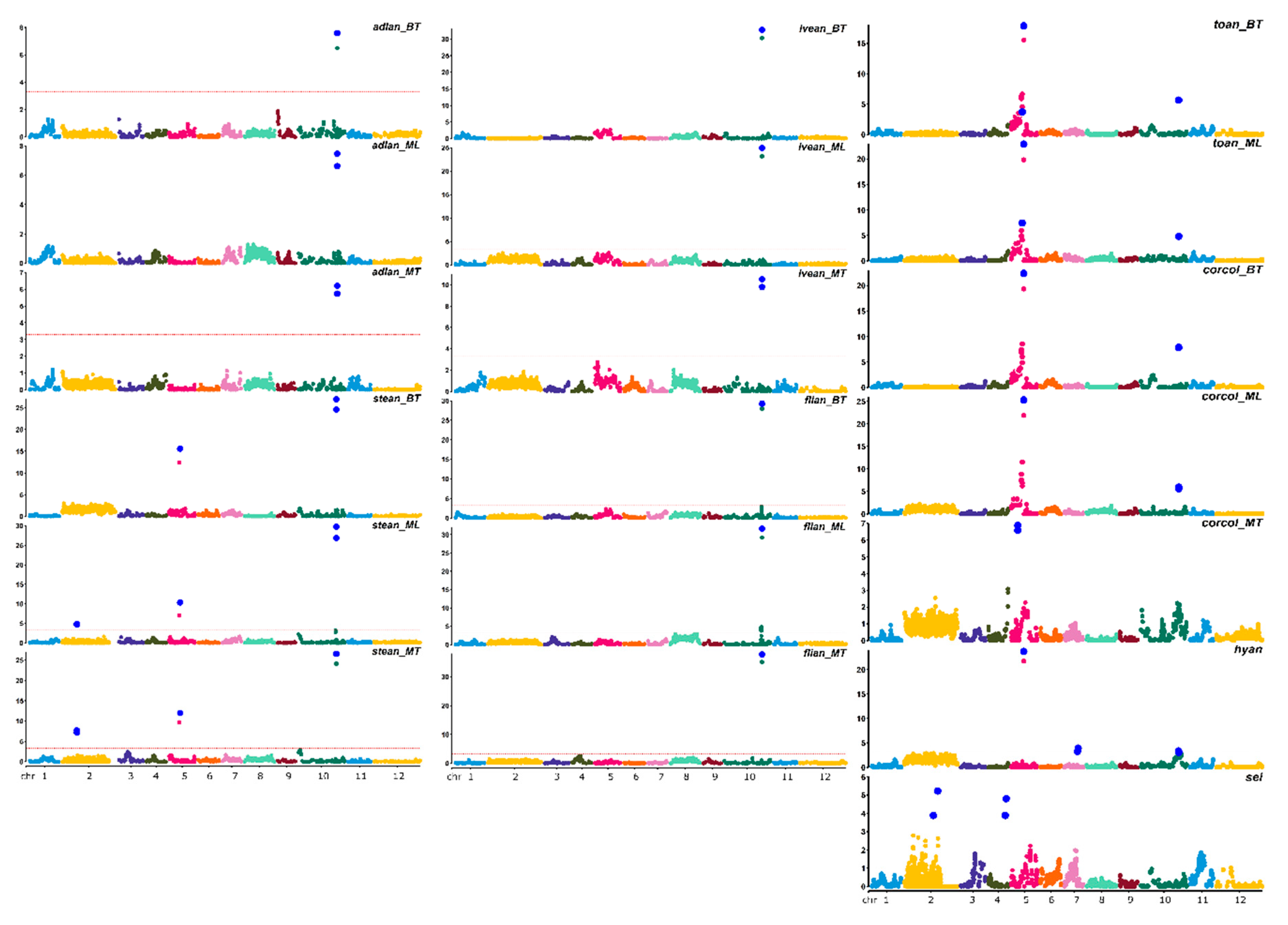{kind=link}
| Trait | Trait Code | 305E40 Mean | ±SD | 67/3 Mean | ±SD | F1 Mean | ±SD | cv | RIL pop Means | ±SD | cv | Shapiro-Wilks | Skewness | SE | Kurtosis | SE | Heritability | Transgressive Respect 305E40 | Transgressive Respect 67/3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Adaxial Leaf Lamina Anthocyanin | adlan_BT | 0.42 | 0.20 | 5.00 | 0.00 | 1.67 | 1.17 | 0.70 | 1.76 | 1.58 | 0.89 | 0.87 | 0.72 | 0.08 | −0.75 | 0.16 | 0.97 | 2 | 0 |
| adlan_ML | 0.42 | 0.20 | 4.67 | 0.41 | 2.25 | 0.42 | 0.19 | 2.04 | 1.58 | 0.77 | 0.90 | 0.43 | 0.08 | −1.15 | 0.16 | 0.94 | 0 | 0 | |
| adlan_MT | 0.00 | 0.00 | 2.67 | 0.87 | 1.11 | 0.60 | 0.54 | 0.69 | 1.03 | 1.49 | 0.71 | 1.63 | 0.07 | 2.14 | 0.13 | 0.92 | 0 | 0 | |
| Stem Anthocyanin | stean_BT | 2.17 | 0.25 | 5.00 | 0.00 | 4.83 | 0.25 | 0.05 | 3.80 | 1.02 | 0.27 | 0.92 | −0.62 | 0.07 | −0.14 | 0.13 | 0.96 | 3 | 0 |
| stean_ML | 2.33 | 0.43 | 5.00 | 0.00 | 4.78 | 0.36 | 0.08 | 3.80 | 0.98 | 0.26 | 0.92 | −0.35 | 0.07 | −0.80 | 0.13 | 0.96 | 0 | 0 | |
| stean_MT | 1.11 | 0.33 | 4.89 | 0.33 | 4.44 | 0.53 | 0.12 | 3.39 | 1.08 | 0.32 | 0.91 | −0.22 | 0.07 | −0.53 | 0.13 | 0.95 | 0 | 0 | |
| Leaf Venation Anthocyanin | lvean_BT | 1.67 | 0.26 | 5.00 | 0.00 | 4.33 | 0.26 | 0.06 | 3.61 | 1.33 | 0.37 | 0.76 | 4.26 | 0.08 | 71.86 | 0.16 | 0.86 | 1 | 0 |
| lvean_ML | 2.75 | 0.42 | 4.92 | 0.20 | 4.75 | 0.27 | 0.06 | 3.74 | 0.99 | 0.27 | 0.92 | −0.54 | 0.08 | −0.24 | 0.16 | 0.87 | 2 | 0 | |
| lvean_MT | 1.00 | 0.71 | 4.22 | 0.97 | 4.44 | 1.01 | 0.23 | 3.06 | 1.53 | 0.50 | 0.91 | −0.38 | 0.07 | −0.95 | 0.13 | 0.87 | 0 | 0 | |
| Flower Anthocyanin Intensity | flian_BT | 2.28 | 1.12 | 5.00 | 0.00 | 4.89 | 0.22 | 0.05 | 4.27 | 0.89 | 0.21 | 0.79 | −1.05 | 0.07 | 0.40 | 0.13 | 0.94 | 0 | 0 |
| flian_ML | 2.89 | 0.89 | 4.89 | 0.33 | 4.67 | 0.43 | 0.09 | 3.95 | 1.06 | 0.27 | 0.83 | −0.75 | 0.07 | −0.06 | 0.13 | 0.95 | 0 | 0 | |
| flian_MT | 3.22 | 1.48 | 4.89 | 0.33 | 5.00 | 0.00 | 0.00 | 4.12 | 0.98 | 0.24 | 0.84 | −0.79 | 0.07 | −0.43 | 0.13 | 0.95 | 0 | 0 | |
| Anthocianin Tonality | toan_BT | 1.00 | 0.00 | 5.00 | 0.00 | 3.00 | 0.00 | 0.00 | 3.02 | 1.86 | 0.62 | 0.72 | −0.02 | 0.11 | −1.85 | 0.23 | 0.94 | 0 | 0 |
| toan_ML | 1.00 | 0.00 | 5.00 | 0.00 | 3.00 | 0.00 | 0.00 | 3.21 | 1.86 | 0.58 | 0.71 | −0.21 | 0.11 | −1.80 | 0.22 | 0.89 | 0 | 0 | |
| Corolla Colour | corcol_BT | 1.00 | 0.00 | 5.00 | 0.00 | 4.11 | 0.33 | 0.08 | 3.00 | 1.78 | 0.59 | 0.76 | −0.01 | 0.07 | −1.78 | 0.13 | 0.97 | 0 | 0 |
| corcol_ML | 1.00 | 0.00 | 5.00 | 0.00 | 3.33 | 1.32 | 0.40 | 3.23 | 1.78 | 0.55 | 0.76 | −0.20 | 0.07 | −1.75 | 0.13 | 0.97 | 0 | 0 | |
| corcol_MT | 4.33 | 1.00 | 5.00 | 0.00 | 4.67 | 0.50 | 0.11 | 4.49 | 1.10 | 0.24 | 0.51 | −2.08 | 0.07 | 3.26 | 0.13 | 0.87 | 0 | 0 | |
| Hypocotyl Anthocyanin | hyan | 0.67 | 0.29 | 4.83 | 1.26 | 3.00 | 0.50 | 0.17 | 2.88 | 1.34 | 0.47 | 0.94 | −0.18 | 0.12 | −0.98 | 0.23 | 0.98 | 1 | 0 |
| Speed of emergence index | sei | 11.67 | 1.53 | 5.67 | 1.53 | 5.67 | 1.15 | 0.20 | 13.65 | 3.00 | 0.22 | 0.90 | 1.27 | 0.12 | 4.80 | 0.23 | 0.98 | 44 | 0 |
| Marker Density | Ratio cM/Mb (Estimated Recombination Rate) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LG/Chromosome | Size (cM) | Size (Mp) | Markers | LG Physical Start (bp)a | LG Physical End (bp)b | Physical Span (Mb)c | Max Gap (cM) | Gap <5 cM | Markers from Different CH | Markers from CH0 | Distorted Markers | cM | kb | |
| E01 | 180.09 | 136.53 | 426 | 1,761,388 | 136,521,595 | 134.76 | 14.89 | 0.984 | 0 | 0 | 54 | 0.4 | 321.3 | 1.32 |
| E02 | 326.00 | 83.34 | 1454 | 5796 | 77,157,371 | 77.15 | 6.71 | 0.999 | 9 | 3 | 1425 | 0.2 | 57.4 | 3.91 |
| E03 | 148.51 | 97.01 | 408 | 36,214 | 82,766,928 | 82.73 | 18.00 | 0.985 | 9 | 19 | 7 | 0.4 | 238.4 | 1.53 |
| E04 | 119.40 | 105.67 | 258 | 1,557,104 | 105,339,521 | 103.78 | 14.46 | 0.973 | 0 | 0 | 0 | 0.5 | 411.2 | 1.13 |
| E05 | 157.15 | 43.85 | 211 | 2,017,627 | 43,756,820 | 41.74 | 13.75 | 0.957 | 0 | 13 | 72 | 0.7 | 208.8 | 3.58 |
| E06 | 126.94 | 108.97 | 838 | 2187 | 99,842,165 | 99.84 | 12.19 | 0.995 | 24 | 104 | 8 | 0.2 | 130.2 | 1.16 |
| E07 | 115.69 | 142.38 | 535 | 16,820 | 140,599,277 | 140.58 | 14.79 | 0.994 | 1 | 16 | 36 | 0.2 | 266.6 | 0.81 |
| E08 | 183.94 | 109.58 | 730 | 2123 | 10,680,425 | 106.80 | 8.89 | 0.999 | 0 | 2 | 1 | 0.3 | 150.3 | 1.68 |
| E09 | 107.06 | 36.10 | 230 | 11,386 | 34,244,824 | 34.23 | 18.18 | 0.978 | 1 | 15 | 1 | 0.5 | 157.6 | 2.97 |
| E10 | 281.23 | 106.64 | 386 | 701,005 | 106,482,885 | 105.78 | 22.29 | 0.956 | 0 | 0 | 68 | 0.7 | 277.0 | 2.64 |
| E11 | 141.34 | 72.29 | 231 | 213,621 | 72,208,518 | 71.99 | 12.24 | 0.974 | 3 | 59 | 39 | 0.6 | 314.3 | 1.96 |
| E12 | 281.90 | 100.42 | 1542 | 10,639 | 96,335,130 | 96.32 | 22.45 | 0.995 | 0 | 1 | 33 | 0.2 | 65.2 | 2.81 |
| Total | 2169.23 | 1142.80 | 7249.00 | 1095.72 | 47 | 232 | 1744 | 0.4 | 216.5 | 2.12 | ||||
| adlanML | adlanMT | corcolBT | corcolML | corcolMT | flianBT | flianML | flianMT | sei | hyan | lveanBT | lveanML | lveanMT | steanBT | steanML | steanMT | toanBT | toanML | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| adlanBT | 0.90 | 0.81 | 0.21 | 0.21 | 0.21 | 0.43 | 0.45 | 0.50 | 0.02 | 0.19 | 0.49 | 0.42 | 0.66 | 0.38 | 0.38 | 0.39 | 0.20 | 0.16 |
| adlanML | 0.83 | 0.20 | 0.19 | 0.18 | 0.37 | 0.43 | 0.45 | 0.00 | 0.17 | 0.40 | 0.41 | 0.66 | 0.32 | 0.37 | 0.38 | 0.19 | 0.18 | |
| adlanMT | 0.15 | 0.15 | 0.18 | 0.39 | 0.42 | 0.45 | 0.00 | 0.11 | 0.42 | 0.39 | 0.65 | 0.30 | 0.37 | 0.36 | 0.12 | 0.15 | ||
| corcolBT | 0.85 | 0.60 | 0.57 | 0.53 | 0.53 | 0.03 | 0.69 | 0.57 | 0.61 | 0.42 | 0.74 | 0.65 | 0.69 | 0.92 | 0.83 | |||
| corcolML | 0.68 | 0.51 | 0.54 | 0.54 | 0.05 | 0.68 | 0.58 | 0.58 | 0.44 | 0.75 | 0.68 | 0.71 | 0.81 | 0.91 | ||||
| corcolMT | 0.42 | 0.37 | 0.41 | −0.12 | 0.48 | 0.45 | 0.44 | 0.34 | 0.60 | 0.56 | 0.55 | 0.58 | 0.63 | |||||
| flianBT | 0.85 | 0.81 | 0.04 | 0.42 | 0.80 | 0.79 | 0.55 | 0.79 | 0.77 | 0.73 | 0.50 | 0.45 | ||||||
| flianML | 0.85 | 0.02 | 0.37 | 0.83 | 0.81 | 0.60 | 0.75 | 0.83 | 0.78 | 0.42 | 0.47 | |||||||
| flianMT | 0.02 | 0.44 | 0.82 | 0.78 | 0.61 | 0.77 | 0.80 | 0.80 | 0.45 | 0.46 | ||||||||
| sei | 0.09 | 0.07 | −0.01 | −0.02 | 0.05 | −0.01 | 0.00 | 0.07 | −0.01 | |||||||||
| hyan | 0.52 | 0.43 | 0.27 | 0.65 | 0.53 | 0.54 | 0.66 | 0.65 | ||||||||||
| lveanBT | 0.83 | 0.59 | 0.83 | 0.82 | 0.80 | 0.52 | 0.50 | |||||||||||
| lveanML | 0.52 | 0.78 | 0.83 | 0.83 | 0.55 | 0.57 | ||||||||||||
| lveanMT | 0.54 | 0.59 | 0.62 | 0.35 | 0.36 | |||||||||||||
| steanBT | 0.88 | 0.88 | 0.70 | 0.68 | ||||||||||||||
| steanML | 0.91 | 0.59 | 0.62 | |||||||||||||||
| steanMT | 0.62 | 0.65 | ||||||||||||||||
| toanBT | 0.84 |
| Battipaglia | Montanaso Lombardo | Monsampolo del Tronto | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Trait | GW | QTL | Position | LOD | CI | PVE | A | GW | QTL | Position | LOD | CI | PVE | A | GW | QTL | Position | LOD | CI | PVE | A | ||||||
| Chr. | cM | Locus | Chr. | cM | Locus | Chr. | cM | Locus | |||||||||||||||||||
| adlan | 3.3 | adlan10.1_BT | E10 | 236.98 | CH10_95003635 | 7.59 | 236.98 | 21.0 | −0.6002 | 3.3 | adlan10.1_ML | E10 | 236.98 | CH10_95003635 | 7.47 | 236.98 | 20.4 | −0.5872 | 3.3 | adlan10.1_MT | E10 | 236.98 | CH10_95003635 | 6.21 | 236.987 | 17.7 | −0.3465 |
| stean | 3.2 | 3.2 | stean2.1_ML | E02 | 88.73 | CH02_30555633 | 4.90 | 88.73 | 5.6 | −0.2294 | 3.2 | stean2.1_ML | E02 | 88.73 | CH02_30555713 | 7.71 | 88.733 | 9.5 | −0.3255 | ||||||||
| stean5.1_BT | E05 | 66.39 | CH05_36124744 | 15.58 | 62.45–66.51 | 21.5 | −0.4263 | stean5.1_ML | E05 | 66.39 | CH05_36124744 | 9.22 | 66.39 | 13.0 | −0.3156 | stean5.1_ML | E05 | 64.51 | CH05_36124744 | 9.70 | 62.45–64.516 | 16.8 | −0.3882 | ||||
| stean10.1_BT | E10 | 231.46 | CH10_94779014 | 26.93 | 231.46 | 45.1 | −0.6000 | stean10.1_ML | E10 | 231.46 | CH10_94779014 | 25.92 | 231.46 | 48.1 | −0.5894 | stean10.1_MT | E10 | 231.46 | CH10_94779014 | 20.76 | 231.465 | 40.3 | −0.5851 | ||||
| lvean | 3.2 | lvean10.1_BT | E10 | 231.46 | CH10_94779014 | 32.78 | 231.46 | 64.2 | −0.7883 | 3.2 | lvean10.1_ML | E10 | 231.46 | CH10_94779014 | 24.92 | 231.46 | 53.2 | −0.6202 | 3.2 | lvean10.1_MT | E10 | 231.46 | CH10_94779014 | 10.52 | 231.465 | 28.2 | −0.5629 |
| flian | 3.1 | flian10.1_BT | E10 | 231.46 | CH10_94779014 | 29.26 | 231.46 | 59.5 | −0.5680 | 3.1 | flian10.1_ML | E10 | 231.46 | CH10_94779014 | 31.64 | 231.46 | 61.9 | −0.6544 | 3.1 | flian10.1_MT | E10 | 231.46 | CH10_94779014 | 37.70 | 231.465 | 69.3 | −0.7393 |
| toan | 3.4 | toan5.1_BT | E05 | 75.48 | CH05_37533757 | 17.89 | 75.48 | 35.7 | −110.9 | 3.4 | toan5.1_ML | E05 | 75.48 | CH05_37533757 | 22.92 | 75.48 | 43.9 | −116208 | |||||||||
| toan5.2_BT | E05 | 66.39 | CH05_36076199 | 3.72 | 61.55–66.39 | 5.2 | −0.6245 | toan5.2_ML | E05 | 66.39 | CH05_36124744 | 7.44 | 61.55–66.39 | 8.7 | −0.7422 | ||||||||||||
| toan10.1_BT | E10 | 232.77 | CH10_94275882 | 5.67 | 232.77 | 9.3 | −0.5498 | toan10.1_ML | E10 | 232.77 | CH10_94275882 | 4.82 | 232.77 | 6.8 | −0.4464 | ||||||||||||
| corcol | 3.3 | corcol5.1_BT | E05 | 75.48 | CH05_37533757 | 22.39 | 75.484 | 40.2 | −107.477 | 3.3 | corcol5.1_ML | E05 | 75.48 | CH05_37533757 | 25.31 | 75.48 | 45.2 | −112175 | 3.3 | corcol5.1_MT | E05 | 39.39 | CH05_17086140 | 6.88 | 39.397 | 19.3 | −0.3440 |
| corcol10.1_BT | E10 | 232.77 | CH10_94275882 | 7.88 | 232.77 | 11.1 | −0.5493 | corcol10.1_ML | E10 | 232.77 | CH10_94281914 | 5.95 | 232.77 | 7.7 | −0.4507 | ||||||||||||
| hyan | 3.3 | hyan5.1 | E05 | 75.48 | CH05_37533757 | 23.72 | 75.48 | 48.1 | −0.9234 | ||||||||||||||||||
| hyan7.1 | E07 | 83.94 | CH07_132761839 | 3.93 | 79.32–83.94 | 5.8 | −0.3223 | ||||||||||||||||||||
| hyan10.1 | E10 | 231.46 | CH10_94684020 | 3.37 | 230.12–233.45 | 4.5 | −0.2807 | ||||||||||||||||||||
| sei | 3.0 | sei2.1 | E02 | 204.18 | CH02_54633733 | 5.23 | 204.18 | 10.4 | −5.689 | ||||||||||||||||||
| sei2.2 | E02 | 176.24 | CH02_63996392 | 3.89 | 176.24 | 7.6 | 5.109 | ||||||||||||||||||||
| sei4.1 | E04 | 108.42 | CH04_102121728 | 4.81 | 102.22–114.85 | 9.7 | −0.8733 | ||||||||||||||||||||
| QTL | Approximative Position | Gene | Predicted Function | |
|---|---|---|---|---|
| IDs SMEL_ | Abbreviation | |||
| sei 2.1 | 9.2 Mb | 002g153950.960 | NHL6 | 2x NDR1/HIN1-like protein 6 |
| 54 Mb | 002g158620 | PECS−2.1 | Pectinesterases 2 | |
| 002g158940 | SNL2 | Paired amphipathic helix protein Sin3-like 2 | ||
| between sei 2.1 and sei 2.2 | ~54–60 Mb | 002g159100 | LAC11 | Laccase |
| 002g159470 | ENY | Zinc finger ENHYDROUS | ||
| 002g159370.380 | PLT6 | 2x polyol transporter | ||
| 002g159480 | GBF1 | G-box-binding factor 1 | ||
| sei 2.2 | 60–63 Mb | 002g160070.080 | CAR2 | 2x C2-Domain Abscisic Acid-Related Proteins |
| 002g160170 | GRDP1 | Glycine-rich domain-containing protein 1 | ||
| 002g159720 | TCP1 | T-complex protein 1 subunit zeta 1 | ||
| 002g159870 | MSR4 | Peptide methionine sulfoxide reductase | ||
| sei 4.1 | 102 Mb | 004g219910.920 | 2x Serpins-ZX | |
| 004g220200–220 | NPF4.5/NPF4.3 | 3x NRT1/ PTR protein fam 4.5/4.3 | ||
| 004g220270 | KO | Ent-kaurene oxidase | ||
| 004g220280 | REM16 | B3 domain transcription factor | ||
| 004g220780 | GAF1 | Zinc finger GAI-ASSOCIATED FACTOR 1 | ||
| ~102 Mb | 004g221390 | abscisic acid 8’ hydrolase 4 | ||
| 004g221470 | BZIP44 | bZIP transcription factor 44 | ||
| toan10.1 corcol 10.1 | ~94 Mb | 010g352310–490 | ANS | 4x 2-oxoglutarate/Fe(II)-dependent dioxygenase |
| 010g352500 | JRG21 | 2-oxoglutarate/Fe(II)-dependent dioxygenase | ||
| 94 Mb | 010g352790 | Myb family transcription factor | ||
| 010g352650 | SAC8 | Phosphoinositide phosphatase | ||
| lvean 10.1 stean 10.1 flian10.1 hyan 10.1 | 94.7 Mb | 010g352910 | BES1/BZR1 | BES1/BZR1 transcription factor |
| 010g352980 | Ankyrin repeat-containing protein | |||
| 010g352930 | DREB2C | Dehydration-Responsive Element-Binding Protein 2C | ||
| 010g353040 | PPC6–1 | protein phosphatase 2C | ||
| ~94.7 Mb | 010g353090–110 | RAPTOR | 3x RAPTOR - Regulatory-associated protein of TOR | |
| adlan 10.1 | 95 Mb | 010g353170–200 | 5x peroxidase | |
| 010g353200 | Protein disulfide-isomerase | |||
| hyan 7.1 | 132.7 Mb | 007g289310–410 | MYBs | 6x similar to MYB15/14/58/102 |
| 007g289700.710 | NDB | 2x NAD(P)H dehydrogenase | ||
| 007g289780 | F-box/kelch-repeat protein | |||
| 007g289410 | PPC6–6 | protein phosphatase 2C | ||
| stean 2.1 | 30.5 Mb | 002g155860 | Ankyrin repeat-containing protein | |
| 002g155880 | LIMYB | L10-interacting MYB domain-containing protein | ||
| 002g155890 | HSP70 | Similar to Heat shock 70 kDa protein | ||
| 002g155950.970 | LARP1C | 2x La-related protein 1C-like | ||
| stean 5.1 toan 5.2 | 36.2 Mb | 005g235930 | HSP | 17.3 kDa class II heat shock protein |
| 005g236210 | BHLH93 | similar to Transcription factor bHLH93 | ||
| ~36.2 Mb | 005g236240 | AAT | Acetyl-CoA-benzylalcohol acetyltransferase | |
| hyan 5.1 toan 5.1 corcol 5.1 | ~37.5 Mb | 005g236840–90 | 7x Calmodulin-like genes | |
| 37.5 Mb | 005g236910.20 | BKI1 | 2x BR1 kinase inhibitors | |
| ~37.5 Mb | 005g236720.30 | TOGT1 | 2x Scopoletin glucosyltransferase | |
| 005g236480 | BHLH84 | Transcription factor bHLH84 | ||
| 005g236490.00 | AZF3,ZAT10 | 2x Zinc finger protein | ||
| 005g236570–620 | CYP81Q32,VQ31 | cytochrome p450 | ||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toppino, L.; Barchi, L.; Mercati, F.; Acciarri, N.; Perrone, D.; Martina, M.; Gattolin, S.; Sala, T.; Fadda, S.; Mauceri, A.; et al. A New Intra-Specific and High-Resolution Genetic Map of Eggplant Based on a RIL Population, and Location of QTLs Related to Plant Anthocyanin Pigmentation and Seed Vigour. Genes 2020, 11, 745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070745
Toppino L, Barchi L, Mercati F, Acciarri N, Perrone D, Martina M, Gattolin S, Sala T, Fadda S, Mauceri A, et al. A New Intra-Specific and High-Resolution Genetic Map of Eggplant Based on a RIL Population, and Location of QTLs Related to Plant Anthocyanin Pigmentation and Seed Vigour. Genes. 2020; 11(7):745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070745
Chicago/Turabian StyleToppino, Laura, Lorenzo Barchi, Francesco Mercati, Nazzareno Acciarri, Domenico Perrone, Matteo Martina, Stefano Gattolin, Tea Sala, Stefano Fadda, Antonio Mauceri, and et al. 2020. "A New Intra-Specific and High-Resolution Genetic Map of Eggplant Based on a RIL Population, and Location of QTLs Related to Plant Anthocyanin Pigmentation and Seed Vigour" Genes 11, no. 7: 745. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070745