Ectopic Expressions of the GhLETM1 Gene Reveal Sensitive Dose Effects on Precise Stamen Development and Male Fertility in Cotton


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Cloning RT-PCR
2.3. Vector Construction and Plant Transformation
2.4. Molecular Analysis of Transformants
2.5. Statistical Analysis
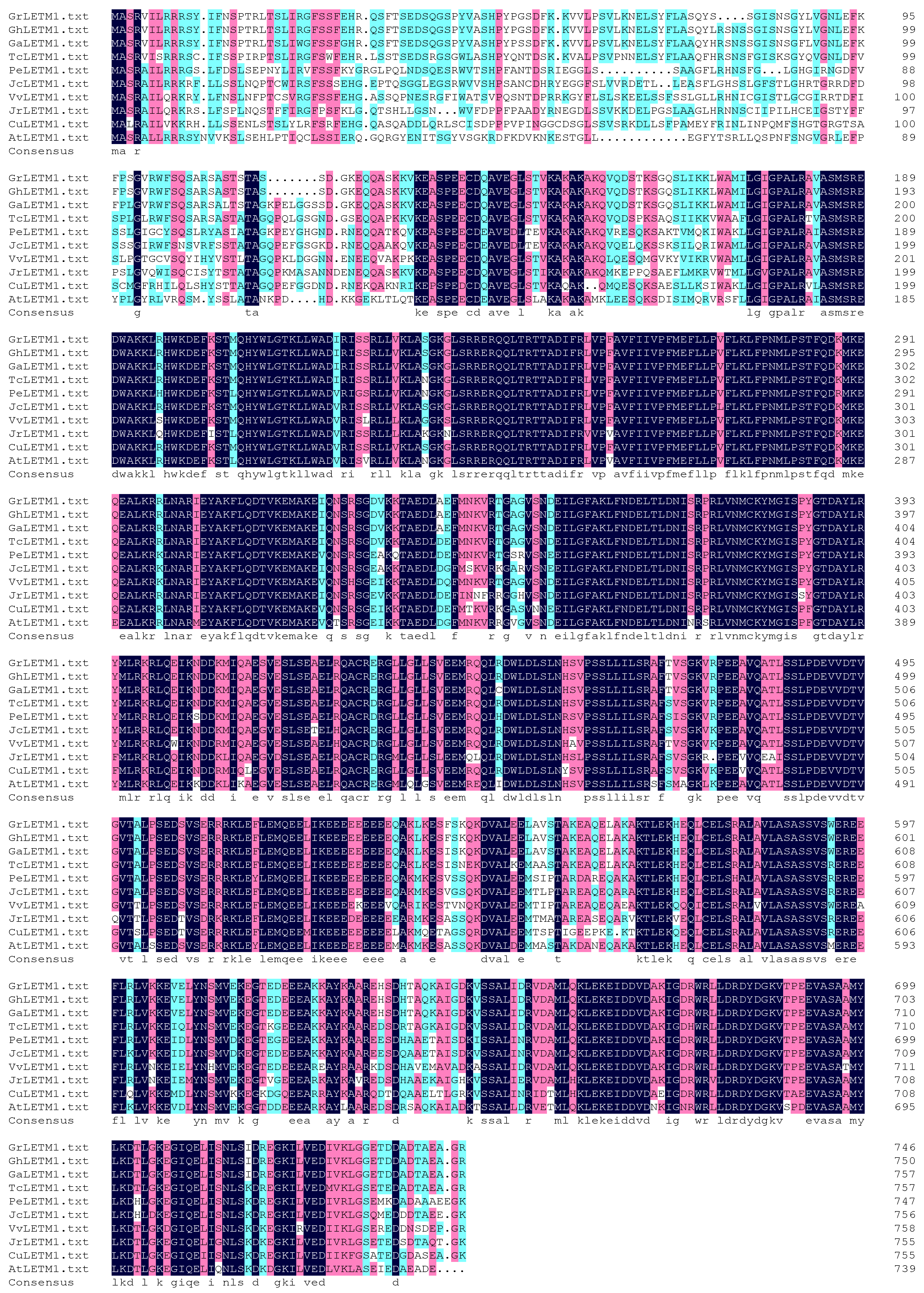
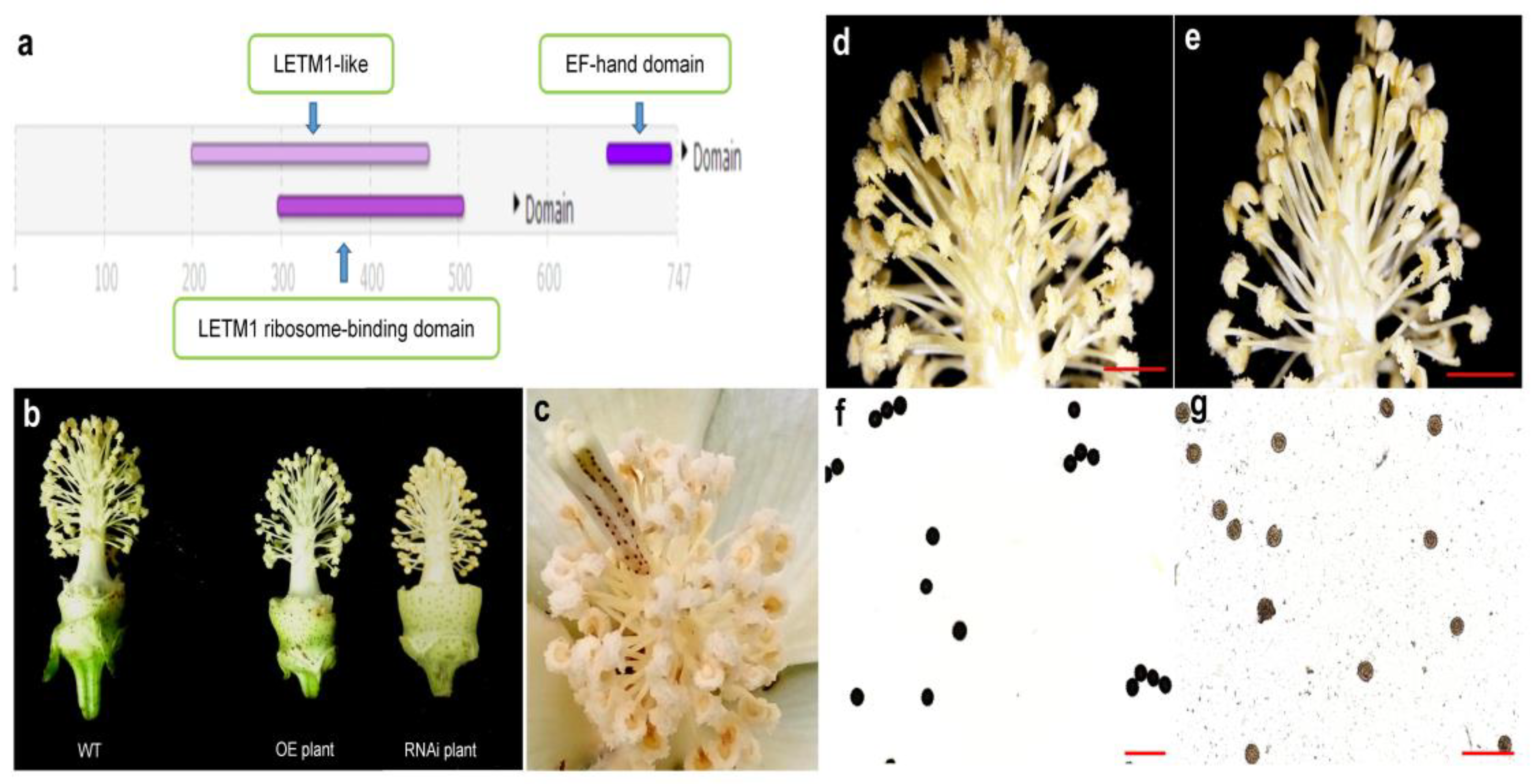
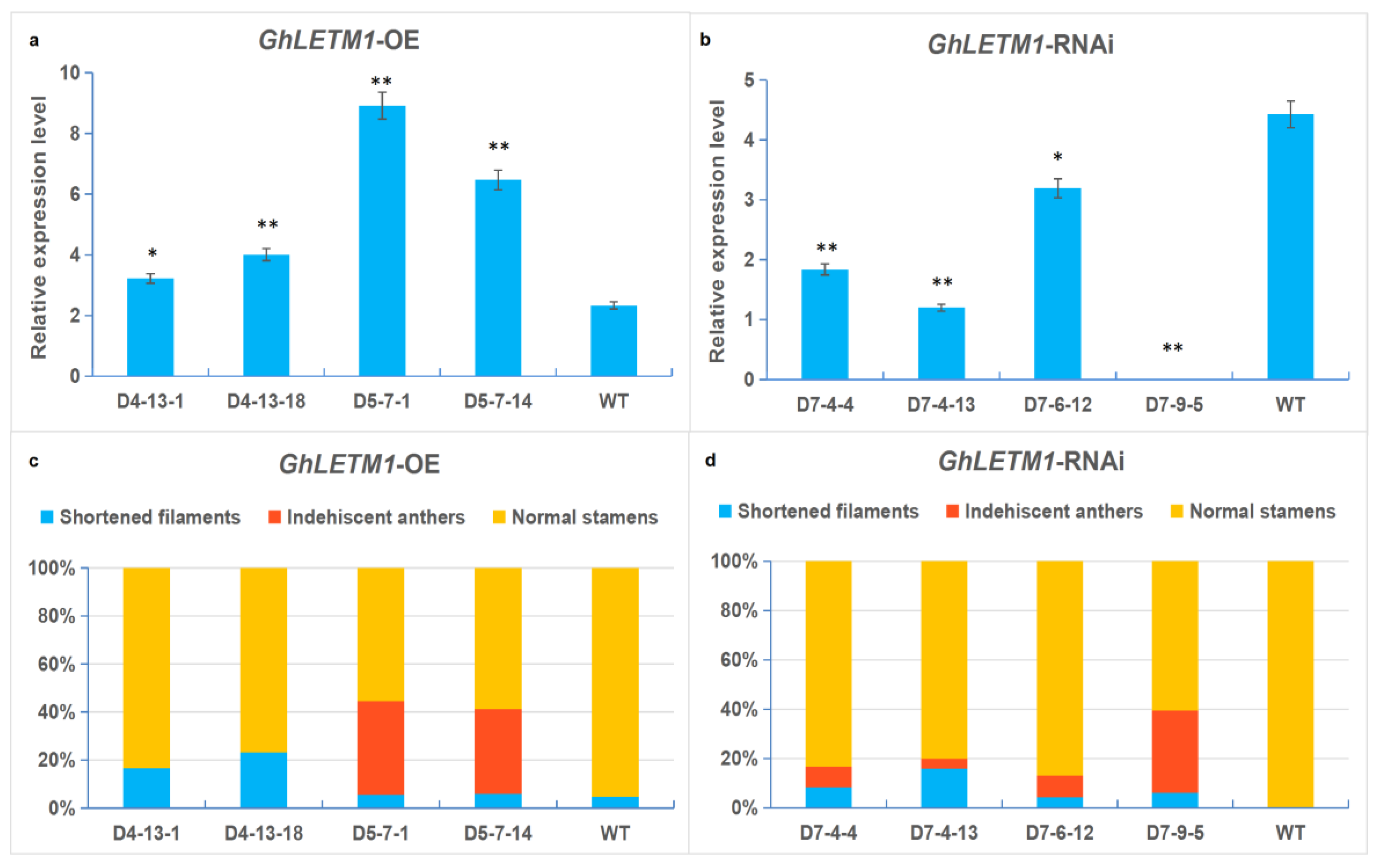
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Gh | Gossypium hirsutum L. |
| LETM | Leucine zipper/EF-hand-containing transmembrane protein |
| OE | Overexpression |
| RNAi | RNA interference |
| CMS | Cytoplasmic Male Sterility |
| qPCR | Quantitative Polymerase Chain Reaction |
References
- Schlickum, S.; Moghekar, A.; Simpson, J.C.; Steglich, C.; O’Brien, R.J.; Winterpacht, A.; Endele, S.U. LETM1, a gene deleted in Wolf-Hirschhorn syndrome, encodes an evolutionarily conserved mitochondrial protein. Genomics 2004, 83, 254–261. [Google Scholar] [CrossRef]
- Piao, L.; Li, Y.; Kim, S.J.; Byun, H.S.; Huang, S.M.; Hwang, S.K.; Yang, K.J.; Park, K.A.; Won, M.; Hong, J.; et al. Association of LETM1 and MRPL36 contributes to the regulation of mitochondrial ATP production and necrotic cell death. Cancer Res. 2009, 69, 3397–3404. [Google Scholar] [CrossRef] [Green Version]
- Jiang, D.; Zhao, L.; Clapham, D.E. Genome-wide RNAi screen identifies Letm1 as a mitochondrial Ca2+/H+ antiporter. Science 2009, 326, 144–147. [Google Scholar] [CrossRef] [Green Version]
- Levings, C.S.; Pring, D.R. Restriction endonuclease analysis of mitochondrial DNA from normal and Texas cytoplasmic male-sterile maize. Science 1976, 193, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Ducos, E.; Touzet, P.; Boutry, M. The male sterile G cytoplasm of wild beet displays modified mitochondrial respiratory complexes. Plant J. 2001, 26, 171–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Xu, H.; Liu, Z.; Guo, J.; Li, H.; Chen, L.; Fang, C.; Zhang, Q.; Bai, M.; Yao, N.; et al. A detrimental mitochondrial-nuclear interaction causes cytoplasmic male sterility in rice. Nat. Genet. 2013, 45, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Touzet, P.; Meyer, E.H. Cytoplasmic male sterility and mitochondrial metabolism in plants. Mitochondrion 2014, 16, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.J.; Guan, Y.C.; Ren, H.B.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Helliwell, C.A.; Wesley, S.V.; Wielopolska, A.J.; Waterhouse, P.M. High-throughput vectors for efficient gene silencing in plants. Funct. Plant Biol. 2002, 29, 1217–1225. [Google Scholar] [CrossRef]
- Liu, Z.; Zhu, Z.; Zhang, T. Development of Transgenic CryIA(c) + GNA Cotton Plants via Pollen Tube Pathway Method Confers Resistance to Helicoverpa Armigera and Aphis Gossypii Glover. Methods Mol. Biol. 2013, 958, 199–210. [Google Scholar] [PubMed]
- Bibi, N.; Fan, K.; Yuan, S.; Ni, M.; Ahmed, I.M.; Malik, W.; Wang, X. An efficient and highly reproducible approach for the selection of upland transgenic cotton produced by pollen tube pathway method. Aust. J. Crop Sci. 2013, 7, 1714–1722. [Google Scholar]
- Wang, M.; Zhang, B.; Wang, Q. Cotton Transformation via Pollen Tube Pathway. Methods Mol. Biol. 2013, 958, 71–77. [Google Scholar] [PubMed]
- Guo, K.; Du, X.; Tu, L.; Tang, W.; Wang, P.; Wang, M.; Liu, Z.; Zhang, X.J. Fibre elongation requires normal redox homeostasis modulated by cytosolic ascorbate peroxidase in cotton (Gossypium hirsutum). Exp. Bot. 2016, 67, 3289–3301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birchler, J.A.; Veitia, R.A. Gene Balance Hypothesis: Connecting Issues of Dosage Sensitivity across Biological Disciplines. Proc. Natl. Acad. Sci. USA 2012, 109, 14746–14753. [Google Scholar] [CrossRef] [PubMed] [Green Version]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Zhang, Y.; Fan, Y.; Guo, H.; Guo, H.; Wu, J.; Wang, H.; Zhao, Y.; Lian, X.; Gou, Z.; et al. Ectopic Expressions of the GhLETM1 Gene Reveal Sensitive Dose Effects on Precise Stamen Development and Male Fertility in Cotton. Genes 2020, 11, 772. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070772
Zhang L, Zhang Y, Fan Y, Guo H, Guo H, Wu J, Wang H, Zhao Y, Lian X, Gou Z, et al. Ectopic Expressions of the GhLETM1 Gene Reveal Sensitive Dose Effects on Precise Stamen Development and Male Fertility in Cotton. Genes. 2020; 11(7):772. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070772
Chicago/Turabian StyleZhang, Li, Yao Zhang, Yijie Fan, Haixia Guo, Huihui Guo, Jianfei Wu, Hongmei Wang, Yunlei Zhao, Xin Lian, Zhongyuan Gou, and et al. 2020. "Ectopic Expressions of the GhLETM1 Gene Reveal Sensitive Dose Effects on Precise Stamen Development and Male Fertility in Cotton" Genes 11, no. 7: 772. https://0-doi-org.brum.beds.ac.uk/10.3390/genes11070772