
Suitability of Pedigree Information and Genomic Methods for Analyzing Inbreeding of Polish Cold-Blooded Horses Covered by Conservation Programs

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
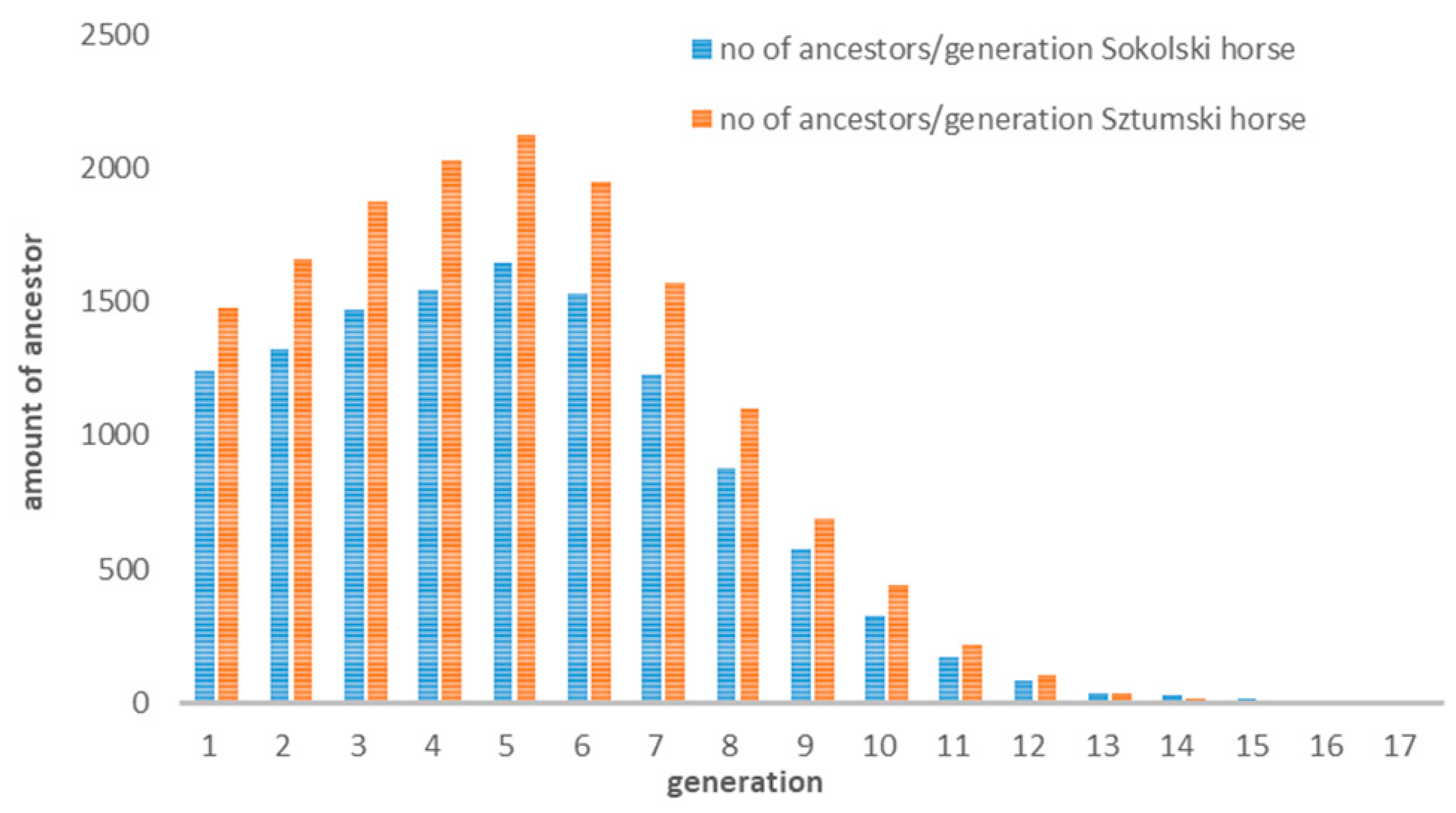
3.1. Pedigree Data Analysis
3.2. Genomic Analysis and Its Comparison within Pedigree Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- VanRaden, P.M. Accounting for Inbreeding and Crossbreeding in Genetic Evaluation of Large Populations. J. Dairy Sci. 1992, 75, 3136–3144. [Google Scholar] [CrossRef]
- Kardos, M.; Luikart, G.; Allendorf, F.W. Measuring individual inbreeding in the age of genomics: Marker-based measures are better than pedigrees. Heredity 2015, 115, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ablondi, M.; Dadousis, C.; Vasini, M.; Eriksson, S.; Mikko, S.; Sabbioni, A. Genetic Diversity and Signatures of Selection in a Native Italian Horse Breed Based on SNP Data. Animals 2020, 10, 1005. [Google Scholar] [CrossRef] [PubMed]
- Mancin, E.; Ablondi, M.; Mantovani, R.; Pigozzi, G.; Sabbioni, A.; Sartori, C. Genetic Variability in the Italian Heavy Draught Horse from Pedigree Data and Genomic Information. Animals 2020, 10, 1310. [Google Scholar] [CrossRef] [PubMed]
- Schiavo, G.; Bovo, S.; Bertolini, F.; Tinarelli, S.; Dall’Olio, S.; Nanni Costa, L.; Fontanesi, L. Comparative evaluation of genomic inbreeding parameters in seven commercial and autochthonous pig breeds. Animal 2020, 14, 910–920. [Google Scholar] [CrossRef]
- Chachuła, J.; Rudowski, M. Studia nad hodowlą koni zimnokrwistych i pogrubionych w Polsce. In Roczniki Nauk Rolniczych; PWRiL: Warszawa, Poland, 1967; Volume 123. (In Polish) [Google Scholar]
- Pruski, W. Hodowla Koni; Tom I; PWRiL: Warszawa, Poland, 1960. (In Polish) [Google Scholar]
- Nozdrzyn-Płotnickim, J. Koń Sokólski; PWRiL: Warszawa, Poland, 1966. (In Polish) [Google Scholar]
- PZHK. Księga Stadna koni Zimnokrwistych i Pogrubionych (Kzp); Tom I; PWRiL: Warszawa, Poland, 1964. (In Polish) [Google Scholar]
- Chrzanowski, S.; Chachuła, J.; Szelągowska-Wąsik, U.; Oleksiak, S.; Wilczak, J. Konie Zimnokrwiste w Polsce Środkowej, Środkowowschodniej i Południowej; PWN: Warszawa, Poland, 1989. (In Polish) [Google Scholar]
- PZHK. Księga Stadna koni Zimnokrwistych i Pogrubionych (Kzp); Tom II; PWRiL: Warszawa, Poland, 1978. (In Polish) [Google Scholar]
- IZ PIB. Program Ochrony Zasobów Genetycznych koni Sokólskich. 2021. Available online: http://www.bioroznorodnosc.izoo.krakow.pl/konie/programy-ochrony (accessed on 21 January 2021).
- IZ PIB. Program Ochrony Zasobów Genetycznych koni Sztumskich. 2021. Available online: http://www.bioroznorodnosc.izoo.krakow.pl/konie/programy-ochrony (accessed on 21 January 2021).
- Polak, G. Genetic Variability of Cold-Blooded Horses Participating in Genetic Resources Conservation Programs, Using Pedigree Analysis. Ann. Anim. Sci. 2019, 19, 49–60. [Google Scholar] [CrossRef]
- Meuwissen, T.; Lou, Z. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 1992, 24, 305–313. [Google Scholar] [CrossRef]
- Gurgul, A.; Jasielczuk, I.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Szmatała, T.; Polak, G.; Monika Bugno-Poniewierska, M. Genetic Differentiation of the Two Types of Polish Cold-Blooded Horses Included in the National Conservation Program. Animals 2020, 10, 542. [Google Scholar] [CrossRef] [Green Version]
- Gurgul, A.; Jasielczuk, I.; Semik-Gurgul, E.; Pawlina-Tyszko, K.; Stefaniuk-Szmukier, M.; Szmatola, T.; Polak, G.; Tomczyk-Wrona, I.; Bugno-Poniewierska, M. A Genome-Wide Scan for Diversifying Selection Signatures in Selected Horse Breeds. PLoS ONE 2019, 14, e0210751. [Google Scholar] [CrossRef] [Green Version]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- McQuillan, R.; Leutenegger, A.L.; Abdel-Rahman, R.; Franklin, C.S.; Pericic, M.; Barac-Lauc, L.; Smolej-Narancic, N.; Janicijevic, B.; Polasek, O.; Tenesa, A.; et al. Runs of Homozygosity in European Populations. Am. J. Hum. Genet. 2008, 83, 359–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Sham, P.C. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szmatoła, T.; Gurgul, A.; Jasielczuk, I.; Ząbek, T.; Ropka-Molik, K.; Litwińczuk, Z.; Bugno-Poniewierska, M. A Comprehensive Analysis of Runs of Homozygosity of Eleven Cattle Breeds Representing Different Production Types. Animals 2019, 9, 1024. [Google Scholar] [CrossRef] [Green Version]
- JASP. Version 0.14, Computer Software; JASP Team: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Alexander, D.H.; Novembre, J.; Lange, K. Fast Model-Based Estimation of Ancestry in Unrelated Individuals. Genome Res. 2009, 19, 1655–1664. [Google Scholar] [CrossRef] [Green Version]
- Wright, S. Evolution in Mendelian Populations. Genetics 1931, 16, 97–159. [Google Scholar] [CrossRef]
- Dolgin, E.S.; Charlesworth, B.; Baird, S.E.; Cutter, A.D. Inbreeding and outbreeding depression in Caenorhabditis nematodes. Evolution 2007, 61, 1339–1352. [Google Scholar] [CrossRef]
- Wright, S. Coefficients of inbreeding and relationship. Am. Nat. 1922, 56, 330–339. [Google Scholar] [CrossRef] [Green Version]
- VanRaden, P.M. Efficient Methods to Compute Genomic Predictions. J. Dairy Sci. 2008, 91, 4414–4423. [Google Scholar] [CrossRef] [Green Version]
- Alemu, S.W.; Kadri, N.K.; Harland, C.; Faux, P.; Charlier, C.; Caballero, A.; Druet, T. An Evaluation of Inbreeding Measures Using a Whole-Genome Sequenced Cattle Pedigree. Heredity 2021, 126, 410–423. [Google Scholar] [CrossRef]
- Milligan, B. Maximum-Likelihood Estimation of Relatedness. Genetics 2003, 163, 1153–1167. [Google Scholar]
- Ritland, K. Estimators for Pairwise Relatedness and Individual Inbreeding Coefficients. Genet. Res. 1966, 67, 175–185. [Google Scholar] [CrossRef]
- Bjelland, D.W.; Weigel, K.A.; Vukasinovic, N.; Nkrumah, J.D. Evaluation of Inbreeding Depression in Holstein Cattle Using Whole-Genome SNP Markers and Alternative Measures of Genomic Inbreeding. J. Dairy Sci. 2013, 96, 4697–4706. [Google Scholar] [CrossRef]
- Ackerman, M.S.; Johri, P.; Spitze, K.; Xu, S.; Doak, T.G.; Young, K.; Lynch, M. Estimating Seven Coefficients of Pairwise Relatedness Using Population-Genomic Data. Genetics 2017, 206, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Ferenčaković, M.; Hamzić, E.; Gredler, B.; Solberg, T.T.; Klemetsdal, G.; Curik, I.; Sölkner, J. Estimates of Autozygosity Derived from Runs of Homozygosity: Empirical Evidence from Selected Cattle Populations. J. Anim. Breed. Genet. 2013, 130, 286–293. [Google Scholar] [CrossRef]
- Kasarda, R.; Moravčíková, N.; Kadlečík, O.; Trakovická, A.; Halo, M.; Candrák, J. Level of inbreeding in Norik of muran horse: Pedigree vs. Genomic data. Acta Univ. Agric. Silvic. Mendel. Brun. 2019, 67, 1457–1463. [Google Scholar] [CrossRef] [Green Version]
- Gurgul, A.; Szmatoła, T.; Topolski, P.; Jasielczuk, I.; Żukowski, K.; Bugno-Poniewierska, M. The Use of Runs of Homozygosity for Estimation of Recent Inbreeding in Holstein Cattle. J. Appl. Genet. 2016, 57, 527–530. [Google Scholar] [CrossRef] [PubMed]
- Purfield, D.C.; Berry, D.P.; McParland, S.; Bradley, D.G. Runs of homozygosity and population history in cattle. BMC Genet. 2012, 13, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Keller, M.C.; Visscher, P.M.; Goddard, M.E. Quantification of Inbreeding Due to Distant Ancestors and Its Detection Using Dense Single Nucleotide Polymorphism Data. Genetics 2011, 189, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, A.; Villanueva, B.; Druet, T. On the Estimation of Inbreeding Depression Using Different Measures of Inbreeding from Molecular Markers. Evol. Appl. 2021, 14, 416–428. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Sztumski | Sokólski | ||
|---|---|---|---|---|
| Sire | Dam | Sire | Dam | |
| Total horses in database | 30,331 | |||
| Year of birth of oldest ancestors | 1932 | 1938 | 1935 | 1940 |
| Total horses under conservation (2008–2020) | 6531 | |||
| 784 | 2594 | 555 | 2598 | |
| Reference population (under conservation in 2020) | 3736 | |||
| 533 | 1509 | 335 | 1359 | |
| Statistics | FPED | FGRM | FIS | FUNI | FROH |
|---|---|---|---|---|---|
| Mean | 0.018 | −0.002 | 0.135 | 0.067 | 0.061 |
| Median | 0.012 | −0.003 | 0.134 | 0.064 | 0.057 |
| Std. Deviation | 0.016 | 0.018 | 0.022 | 0.018 | 0.018 |
| Shapiro–Wilk test | 0.690 | 0.976 | 0.986 | 0.965 | 0.931 |
| p-value of Shapiro–Wilk | <0.001 | 0.004 | 0.072 | <0.001 | <0.001 |
| Minimum | 0.002 | −0.044 | 0.069 | 0.023 | 0.026 |
| Maximum | 0.135 | 0.081 | 0.221 | 0.151 | 0.149 |
| Statistic | FPED | FGRM | FIS | FUNI | FROH | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| SOK | SZTUM | SOK | SZTUM | SOK | SZTUM | SOK | SZTUM | SOK | SZTUM | |
| Mean | 0.018 | 0.017 | −9.6 × 10−4 | −0.004 | 0.137 | 0.133 | 0.068 | 0.064 | 0.063 | 0.058 |
| Std. Error of Mean | 0.001 | 0.003 | 0.002 | 0.002 | 0.002 | 0.003 | 0.002 | 0.002 | 0.002 | 0.002 |
| Median | 0.014 | 0.011 | −0.003 | −0.005 | 0.137 | 0.132 | 0.069 | 0.062 | 0.061 | 0.054 |
| Std. Deviation | 0.013 | 0.021 | 0.017 | 0.019 | 0.020 | 0.024 | 0.017 | 0.019 | 0.017 | 0.019 |
| Minimum | 0.002 | 0.002 | −0.038 | −0.044 | 0.084 | 0.069 | 0.023 | 0.034 | 0.026 | 0.028 |
| Maximum | 0.070 | 0.135 | 0.046 | 0.081 | 0.199 | 0.221 | 0.123 | 0.151 | 0.123 | 0.149 |
| F Coefficient | Spearman | |||||
|---|---|---|---|---|---|---|
| rho | p | Lower 95% CI | Upper 95% CI | |||
| FPED | - | FGRM | 0.351 | <0.001 | 0.213 | 0.474 |
| FPED | - | FIS | 0.381 | <0.001 | 0.247 | 0.501 |
| FPED | - | FUNI | 0.430 | <0.001 | 0.301 | 0.544 |
| FPED | - | FROH | 0.443 | <0.001 | 0.316 | 0.555 |
| FGRM | - | FIS | 0.522 | <0.001 | 0.405 | 0.622 |
| FGRM | - | FUNI | 0.837 | <0.001 | 0.787 | 0.877 |
| FGRM | - | FROH | 0.733 | <0.001 | 0.655 | 0.795 |
| FIS | - | FUNI | 0.888 | <0.001 | 0.852 | 0.916 |
| FIS | - | FROH | 0.875 | <0.001 | 0.835 | 0.906 |
| FUNI | - | FROH | 0.919 | <0.001 | 0.892 | 0.939 |
| Statistic | Q-Value to SOK Population | Q-Value to SZTUM Population | Lineage (Pedigree) | |||
|---|---|---|---|---|---|---|
| SOK | SZTUM | SOK | SZTUM | SOK | SZTUM | |
| Mean | 0.752 | 0.166 | 0.248 | 0.834 | 0.856 | 0.865 |
| Median | 0.759 | 0.167 | 0.241 | 0.833 | 0.844 | 0.875 |
| Std. Deviation | 0.144 | 0.124 | 0.144 | 0.124 | 0.064 | 0.089 |
| Variance | 0.021 | 0.015 | 0.021 | 0.015 | 0.004 | 0.008 |
| Minimum | 0.267 | 1.000 × 10−5 | 1.000 × 10−5 | 0.54 | 0.719 | 0.563 |
| Maximum | 1 | 0.46 | 0.733 | 1 | 0.969 | 1 |
| Sum | 79.747 | 11.435 | 26.253 | 57.565 | 90.752 | 59.656 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polak, G.; Gurgul, A.; Jasielczuk, I.; Szmatoła, T.; Krupiński, J.; Bugno-Poniewierska, M. Suitability of Pedigree Information and Genomic Methods for Analyzing Inbreeding of Polish Cold-Blooded Horses Covered by Conservation Programs. Genes 2021, 12, 429. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030429
Polak G, Gurgul A, Jasielczuk I, Szmatoła T, Krupiński J, Bugno-Poniewierska M. Suitability of Pedigree Information and Genomic Methods for Analyzing Inbreeding of Polish Cold-Blooded Horses Covered by Conservation Programs. Genes. 2021; 12(3):429. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030429
Chicago/Turabian StylePolak, Grażyna, Artur Gurgul, Igor Jasielczuk, Tomasz Szmatoła, Jędrzej Krupiński, and Monika Bugno-Poniewierska. 2021. "Suitability of Pedigree Information and Genomic Methods for Analyzing Inbreeding of Polish Cold-Blooded Horses Covered by Conservation Programs" Genes 12, no. 3: 429. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12030429