
ANGPTL3 Variants Associate with Lower Levels of Irisin and C-Peptide in a Cohort of Arab Individuals

 ,
,  ,
, 
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recruitment of Participants and Study Cohort
2.2. Blood Sample Collection and Processing
2.3. Estimation of Plasma Levels of Various Biomarkers
2.4. Targeted Genotyping of the ANGPTL3 Study Variants rs1748197 and rs12130333
2.5. Quality Procedures for SNP and Trait Measurements
2.6. Allele-Based Association Tests and Thresholds for Ascertaining Statistical Significance
2.7. Assessing the Interaction of Correlations between Study Variants and Traits
3. Results
3.1. Characteristics of the Two ANGPTL3 Variants
3.2. Characteristics of the Study Cohort
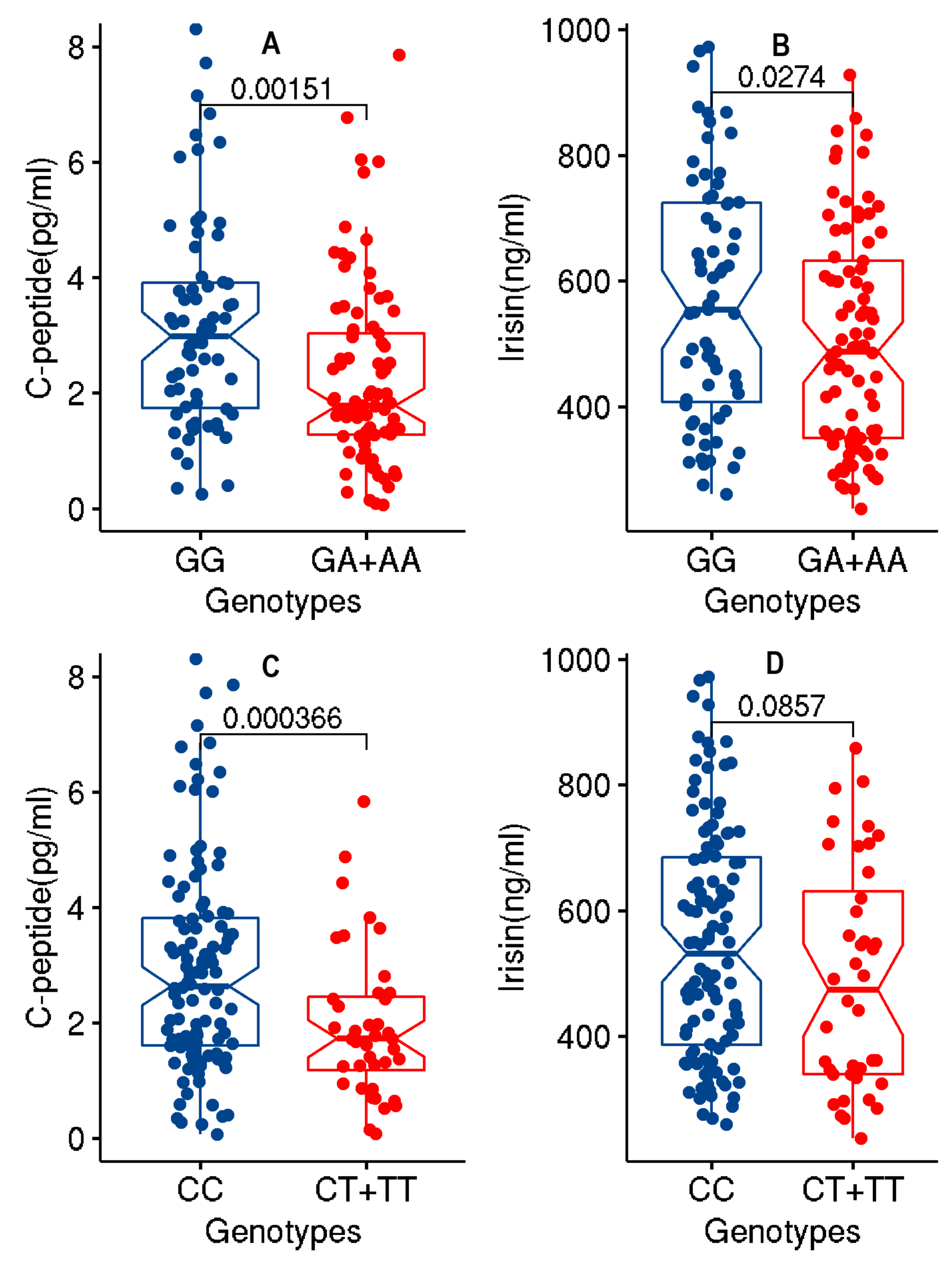
3.3. Association of the Two ANGPTL3 Study Variants with Lower Levels of c-Peptide and Irisin at Significant p-Values
3.4. Associations of the Haplotype of the ANGPTL3 Study Variants with the Levels of c-Peptide, and Irisin
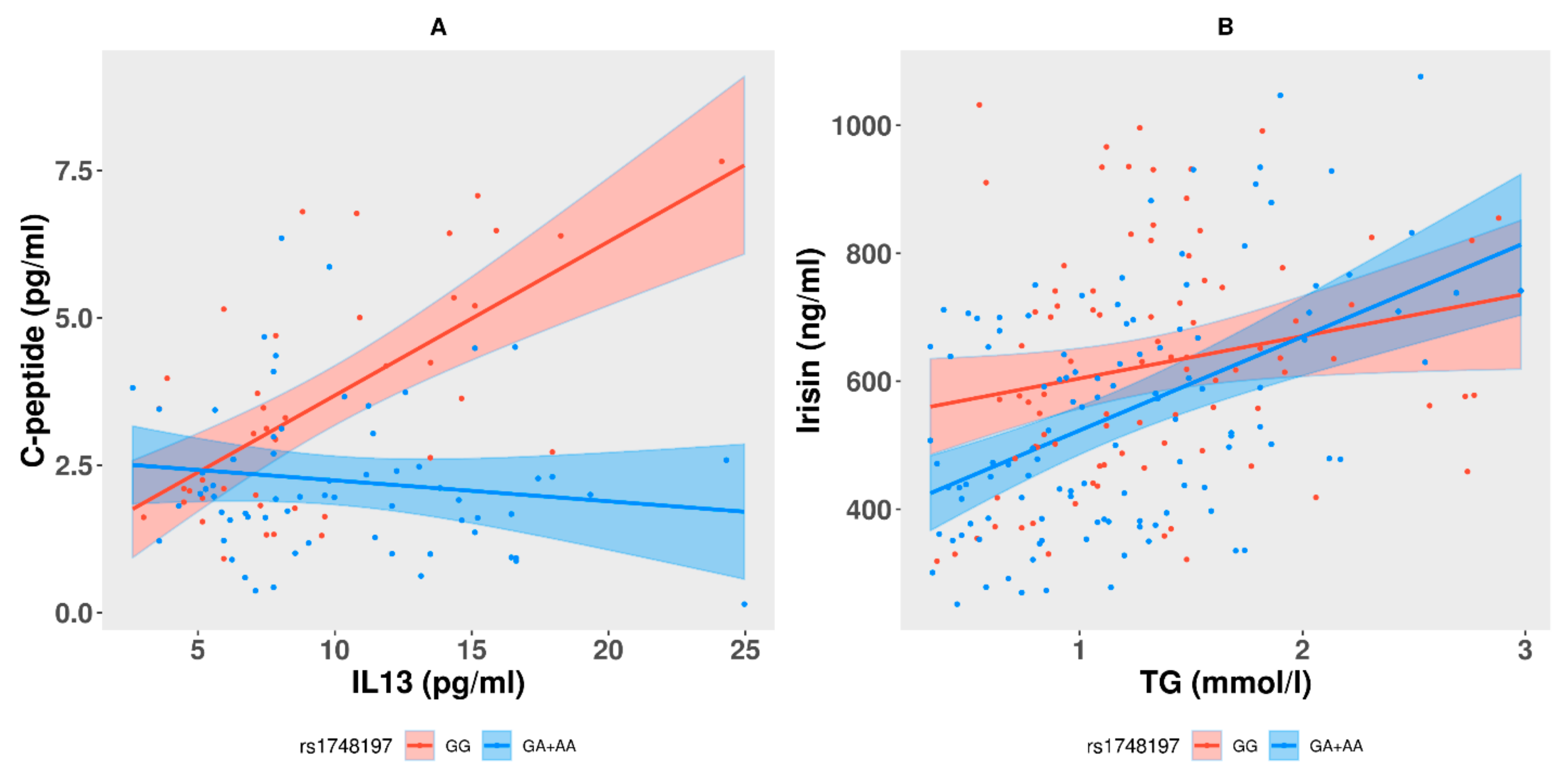
3.5. Interactions between Genotypes at the Study Variants and Correlations among the Levels of c-Peptide or Irisin and Other Traits
3.6. Disease Status of the Cohort Participants and the Impact of the Effect Alleles at the Study Variants on the Levels of c-Peptide, Irisin and TG
3.7. Power Calculation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Unger, R.H. Minireview: Weapons of lean body mass destruction: The role of ectopic lipids in the metabolic syndrome. Endocrinology 2003, 144, 5159–5165. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.B.; Petersen, K.F.; Shulman, G.I. Disordered lipid metabolism and the pathogenesis of insulin resistance. Physiol. Rev. 2007, 87, 507–520. [Google Scholar] [CrossRef] [PubMed]
- Abu-Farha, M.; Abubaker, J.; Tuomilehto, J. Angptl8 (betatrophin) role in diabetes and metabolic diseases. Diabetes Metab. Res. Rev. 2017, 33, e2919. [Google Scholar] [CrossRef]
- Su, X.; Peng, D.Q. New insights into angplt3 in controlling lipoprotein metabolism and risk of cardiovascular diseases. Lipids Health Dis. 2018, 17, 12. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.J.; Lee, R.G.; Brandt, T.A.; Tai, L.J.; Fu, W.; Peralta, R.; Yu, R.; Hurh, E.; Paz, E.; McEvoy, B.W.; et al. Cardiovascular and metabolic effects of angptl3 antisense oligonucleotides. N. Engl. J. Med. 2017, 377, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Buniello, A.; MacArthur, J.A.L.; Cerezo, M.; Harris, L.W.; Hayhurst, J.; Malangone, C.; McMahon, A.; Morales, J.; Mountjoy, E.; Sollis, E.; et al. The nhgri-ebi gwas catalog of published genome-wide association studies, targeted arrays and summary statistics 2019. Nucleic Acids Res. 2019, 47, D1005–D1012. [Google Scholar] [CrossRef]
- Ma, L.; Ballantyne, C.M.; Belmont, J.W.; Keinan, A.; Brautbar, A. Interaction between snps in the rxra and near angptl3 gene region inhibits apob reduction after statin-fenofibric acid therapy in individuals with mixed dyslipidemia. J. Lipid Res. 2012, 53, 2425–2428. [Google Scholar] [CrossRef]
- Wang, J.; Ban, M.R.; Zou, G.Y.; Cao, H.; Lin, T.; Kennedy, B.A.; Anand, S.; Yusuf, S.; Huff, M.W.; Pollex, R.L.; et al. Polygenic determinants of severe hypertriglyceridemia. Hum. Mol. Genet. 2008, 17, 2894–2899. [Google Scholar] [CrossRef]
- Jaromi, L.; Csongei, V.; Polgar, N.; Rappai, G.; Szolnoki, Z.; Maasz, A.; Horvatovich, K.; Safrany, E.; Sipeky, C.; Magyari, L.; et al. Triglyceride level-influencing functional variants of the angptl3, cilp2, and trib1 loci in ischemic stroke. Neuromol. Med. 2011, 13, 179–186. [Google Scholar] [CrossRef]
- Minicocci, I.; Montali, A.; Robciuc, M.R.; Quagliarini, F.; Censi, V.; Labbadia, G.; Gabiati, C.; Pigna, G.; Sepe, M.L.; Pannozzo, F.; et al. Mutations in the angptl3 gene and familial combined hypolipidemia: A clinical and biochemical characterization. J. Clin. Endocrinol. Metab. 2012, 97, E1266–E1275. [Google Scholar] [CrossRef]
- Gallois, A.; Mefford, J.; Ko, A.; Vaysse, A.; Julienne, H.; Ala-Korpela, M.; Laakso, M.; Zaitlen, N.; Pajukanta, P.; Aschard, H. A comprehensive study of metabolite genetics reveals strong pleiotropy and heterogeneity across time and context. Nat. Commun. 2019, 10, 4788. [Google Scholar] [CrossRef]
- Rader, D.J.; Hovingh, G.K. Hdl and cardiovascular disease. Lancet 2014, 384, 618–625. [Google Scholar] [CrossRef]
- Emerging Risk Factors, C.; Di Angelantonio, E.; Sarwar, N.; Perry, P.; Kaptoge, S.; Ray, K.K.; Thompson, A.; Wood, A.M.; Lewington, S.; Sattar, N.; et al. Major lipids, apolipoproteins, and risk of vascular disease. JAMA 2009, 302, 1993–2000. [Google Scholar] [CrossRef]
- Holmes, M.V.; Millwood, I.Y.; Kartsonaki, C.; Hill, M.R.; Bennett, D.A.; Boxall, R.; Guo, Y.; Xu, X.; Bian, Z.; Hu, R.; et al. Lipids, lipoproteins, and metabolites and risk of myocardial infarction and stroke. J. Am. Coll. Cardiol. 2018, 71, 620–632. [Google Scholar] [CrossRef]
- Girona, J.; Amigo, N.; Ibarretxe, D.; Plana, N.; Rodriguez-Borjabad, C.; Heras, M.; Ferre, R.; Gil, M.; Correig, X.; Masana, L. Hdl triglycerides: A new marker of metabolic and cardiovascular risk. Int. J. Mol. Sci. 2019, 20, 3151. [Google Scholar] [CrossRef]
- Teslovich, T.M.; Musunuru, K.; Smith, A.V.; Edmondson, A.C.; Stylianou, I.M.; Koseki, M.; Pirruccello, J.P.; Ripatti, S.; Chasman, D.I.; Willer, C.J.; et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 2010, 466, 707–713. [Google Scholar] [CrossRef]
- Oldoni, F.; Palmen, J.; Giambartolomei, C.; Howard, P.; Drenos, F.; Plagnol, V.; Humphries, S.E.; Talmud, P.J.; Smith, A.J. Post-gwas methodologies for localisation of functional non-coding variants: Angptl3. Atherosclerosis 2016, 246, 193–201. [Google Scholar] [CrossRef]
- Raal, F.J.; Rosenson, R.S.; Reeskamp, L.F.; Hovingh, G.K.; Kastelein, J.J.P.; Rubba, P.; Ali, S.; Banerjee, P.; Chan, K.C.; Gipe, D.A.; et al. Evinacumab for homozygous familial hypercholesterolemia. N. Engl. J. Med. 2020, 383, 711–720. [Google Scholar] [CrossRef]
- Grundy, S.M.; Stone, N.J.; Bailey, A.L.; Beam, C.; Birtcher, K.K.; Blumenthal, R.S.; Braun, L.T.; de Ferranti, S.; Faiella-Tommasino, J.; Forman, D.E.; et al. 2018 aha/acc/aacvpr/aapa/abc/acpm/ada/ags/apha/aspc/nla/pcna guideline on the management of blood cholesterol: A report of the american college of cardiology/american heart association task force on clinical practice guidelines. Circulation 2019, 139, e1082–e1143. [Google Scholar]
- AlKhairi, I.; Cherian, P.; Abu-Farha, M.; Madhoun, A.A.; Nizam, R.; Melhem, M.; Jamal, M.; Al-Sabah, S.; Ali, H.; Tuomilehto, J.; et al. Increased expression of meteorin-like hormone in type 2 diabetes and obesity and its association with irisin. Cells 2019, 8, 1283. [Google Scholar] [CrossRef]
- Perakakis, N.; Triantafyllou, G.A.; Fernandez-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and role of irisin in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef]
- Arhire, L.I.; Mihalache, L.; Covasa, M. Irisin: A hope in understanding and managing obesity and metabolic syndrome. Front. Endocrinol. 2019, 10, 524. [Google Scholar] [CrossRef] [PubMed]
- Mahgoub, M.O.; D’Souza, C.; Al Darmaki, R.; Baniyas, M.; Adeghate, E. An update on the role of irisin in the regulation of endocrine and metabolic functions. Peptides 2018, 104, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Tine Kartinah, N.; Rosalyn Sianipar, I.; Nafi’ah; Rabia. The effects of exercise regimens on irisin levels in obese rats model: Comparing high-intensity intermittent with continuous moderate-intensity training. BioMed Res. Int. 2018, 2018, 4708287. [Google Scholar] [CrossRef]
- Bostrom, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostrom, E.A.; Choi, J.H.; Long, J.Z.; et al. A pgc1-alpha-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Villarroya, F. Irisin, turning up the heat. Cell Metab. 2012, 15, 277–278. [Google Scholar] [CrossRef]
- Timmons, J.A.; Baar, K.; Davidsen, P.K.; Atherton, P.J. Is irisin a human exercise gene? Nature 2012, 488, E9–E10. [Google Scholar] [CrossRef]
- Maalouf, G.E.; El Khoury, D. Exercise-induced irisin, the fat browning myokine, as a potential anticancer agent. J. Obes. 2019, 2019, 6561726. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation plink: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, 7. [Google Scholar] [CrossRef]
- Li, J.; Ji, L. Adjusting multiple testing in multilocus analyses using the eigenvalues of a correlation matrix. Heredity 2005, 95, 221–227. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Liu, D.J.; Peloso, G.M.; Yu, H.; Butterworth, A.S.; Wang, X.; Mahajan, A.; Saleheen, D.; Emdin, C.; Alam, D.; Alves, A.C.; et al. Exome-wide association study of plasma lipids in >300,000 individuals. Nat. Genet. 2017, 49, 1758–1766. [Google Scholar] [CrossRef]
- Kathiresan, S.; Melander, O.; Guiducci, C.; Surti, A.; Burtt, N.P.; Rieder, M.J.; Cooper, G.M.; Roos, C.; Voight, B.F.; Havulinna, A.S.; et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat. Genet. 2008, 40, 189–197. [Google Scholar] [CrossRef]
- Kulminski, A.M.; Huang, J.; Loika, Y.; Arbeev, K.G.; Bagley, O.; Yashkin, A.; Duan, M.; Culminskaya, I. Strong impact of natural-selection-free heterogeneity in genetics of age-related phenotypes. Aging 2018, 10, 492–514. [Google Scholar] [CrossRef]
- Wojcik, G.L.; Graff, M.; Nishimura, K.K.; Tao, R.; Haessler, J.; Gignoux, C.R.; Highland, H.M.; Patel, Y.M.; Sorokin, E.P.; Avery, C.L.; et al. Genetic analyses of diverse populations improves discovery for complex traits. Nature 2019, 570, 514–518. [Google Scholar] [CrossRef]
- Willer, C.J.; Sanna, S.; Jackson, A.U.; Scuteri, A.; Bonnycastle, L.L.; Clarke, R.; Heath, S.C.; Timpson, N.J.; Najjar, S.S.; Stringham, H.M.; et al. Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat. Genet. 2008, 40, 161–169. [Google Scholar] [CrossRef]
- Hebbar, P.; Abubaker, J.A.; Abu-Farha, M.; Alsmadi, O.; Elkum, N.; Alkayal, F.; John, S.E.; Channanath, A.; Iqbal, R.; Pitkaniemi, J.; et al. Genome-wide landscape establishes novel association signals for metabolic traits in the arab population. Hum. Genet. 2020, 140, 505–528. [Google Scholar] [CrossRef]
- Katzmann, J.L.; Packard, C.J.; Chapman, M.J.; Katzmann, I.; Laufs, U. Targeting rna with antisense oligonucleotides and small interfering rna: Jacc state-of-the-art review. J. Am. Coll. Cardiol. 2020, 76, 563–579. [Google Scholar] [CrossRef]
- Musunuru, K.; Pirruccello, J.P.; Do, R.; Peloso, G.M.; Guiducci, C.; Sougnez, C.; Garimella, K.V.; Fisher, S.; Abreu, J.; Barry, A.J.; et al. Exome sequencing, angptl3 mutations, and familial combined hypolipidemia. N. Engl. J. Med. 2010, 363, 2220–2227. [Google Scholar] [CrossRef]
- Abu-Farha, M.; Ghosh, A.; Al-Khairi, I.; Madiraju, S.R.M.; Abubaker, J.; Prentki, M. The multi-faces of angptl8 in health and disease: Novel functions beyond lipoprotein lipase modulation. Prog. Lipid Res. 2020, 80, 101067. [Google Scholar] [CrossRef]
- Mo, L.; Shen, J.; Liu, Q.; Zhang, Y.; Kuang, J.; Pu, S.; Cheng, S.; Zou, M.; Jiang, W.; Jiang, C.; et al. Irisin is regulated by car in liver and is a mediator of hepatic glucose and lipid metabolism. Mol. Endocrinol. 2016, 30, 533–542. [Google Scholar] [CrossRef]
- Zhang, R. The angptl3-4-8 model, a molecular mechanism for triglyceride trafficking. Open Biol. 2016, 6, 150272. [Google Scholar] [CrossRef] [PubMed]
- Rana, K.S.; Pararasa, C.; Afzal, I.; Nagel, D.A.; Hill, E.J.; Bailey, C.J.; Griffiths, H.R.; Kyrou, I.; Randeva, H.S.; Bellary, S.; et al. Plasma irisin is elevated in type 2 diabetes and is associated with increased e-selectin levels. Cardiovasc. Diabetol. 2017, 16, 147. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Zaichenko, L.; Brinkoetter, M.; Thakkar, B.; Sahin-Efe, A.; Joung, K.E.; Tsoukas, M.A.; Geladari, E.V.; Huh, J.Y.; Dincer, F.; et al. Circulating irisin in relation to insulin resistance and the metabolic syndrome. J. Clin. Endocrinol. Metab. 2013, 98, 4899–4907. [Google Scholar] [CrossRef]
- Zhang, H.J.; Zhang, X.F.; Ma, Z.M.; Pan, L.L.; Chen, Z.; Han, H.W.; Han, C.K.; Zhuang, X.J.; Lu, Y.; Li, X.J.; et al. Irisin is inversely associated with intrahepatic triglyceride contents in obese adults. J. Hepatol. 2013, 59, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Avramoglu, R.K.; Basciano, H.; Adeli, K. Lipid and lipoprotein dysregulation in insulin resistant states. Clin. Chim. Acta 2006, 368, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Ginsberg, H.N.; Zhang, Y.L.; Hernandez-Ono, A. Regulation of plasma triglycerides in insulin resistance and diabetes. Arch. Med. Res. 2005, 36, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Forst, T.; Kunt, T.; Pfutzner, A.; Beyer, J.; Wahren, J. New aspects on biological activity of c-peptide in iddm patients. Exp. Clin. Endocrinol. Diabetes 1998, 106, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Cardellini, M.; Farcomeni, A.; Ballanti, M.; Morelli, M.; Davato, F.; Cardolini, I.; Grappasonni, G.; Rizza, S.; Guglielmi, V.; Porzio, O.; et al. C-peptide: A predictor of cardiovascular mortality in subjects with established atherosclerotic disease. Diabetes Vasc. Dis. Res. 2017, 14, 395–399. [Google Scholar] [CrossRef]
- Becht, F.S.; Walther, K.; Martin, E.; Nauck, M.A. Fasting c-peptide and related parameters characterizing insulin secretory capacity for correctly classifying diabetes type and for predicting insulin requirement in patients with type 2 diabetes. Exp. Clin. Endocrinol. Diabetes 2016, 124, 148–156. [Google Scholar] [CrossRef]
- Cabrera de Leon, A.; Oliva Garcia, J.G.; Marcelino Rodriguez, I.; Almeida Gonzalez, D.; Aleman Sanchez, J.J.; Brito Diaz, B.; Dominguez Coello, S.; Bertomeu Martinez, V.; Aguirre Jaime, A.; Rodriguez Perez Mdel, C. C-peptide as a risk factor of coronary artery disease in the general population. Diabetes Vasc. Dis. Res. 2015, 12, 199–207. [Google Scholar] [CrossRef]
- Williams, G.V.; Gambhir, K.K.; Nunlee-Bland, G.; Abrams, C.K.; Ganta, V.; Odonkor, W. Significance of plasma c-peptide in obese african american adolescents. J. Natl. Med. Assoc. 2011, 103, 907–916. [Google Scholar] [CrossRef]
- Khan, H.A.; Sobki, S.H.; Ekhzaimy, A.; Khan, I.; Almusawi, M.A. Biomarker potential of c-peptide for screening of insulin resistance in diabetic and non-diabetic individuals. Saudi J. Biol. Sci. 2018, 25, 1729–1732. [Google Scholar] [CrossRef]
- Yanai, H.; Hirowatari, Y. Fasting serum c-peptide levels (>1.6 ng/ml) can predict the presence of insulin resistance in Japanese patients with type 2 diabetes. Diabetes Metab. 2017, 43, 97–98. [Google Scholar] [CrossRef]
- Martinez-Reyes, C.P.; Gomez-Arauz, A.Y.; Torres-Castro, I.; Manjarrez-Reyna, A.N.; Palomera, L.F.; Olivos-Garcia, A.; Mendoza-Tenorio, E.; Sanchez-Medina, G.A.; Islas-Andrade, S.; Melendez-Mier, G.; et al. Serum levels of interleukin-13 increase in subjects with insulin resistance but do not correlate with markers of low-grade systemic inflammation. J. Diabetes Res. 2018, 2018, 7209872. [Google Scholar] [CrossRef]
- Stanya, K.J.; Jacobi, D.; Liu, S.; Bhargava, P.; Dai, L.; Gangl, M.R.; Inouye, K.; Barlow, J.L.; Ji, Y.; Mizgerd, J.P.; et al. Direct control of hepatic glucose production by interleukin-13 in mice. J. Clin. Investig. 2013, 123, 261–271. [Google Scholar] [CrossRef]
- López, J.C. Il-13 controls blood sugar. Nature Med. 2013, 19, 142. [Google Scholar] [CrossRef]
- Zhang, J.; Valverde, P.; Zhu, X.; Murray, D.; Wu, Y.; Yu, L.; Jiang, H.; Dard, M.M.; Huang, J.; Xu, Z.; et al. Exercise-induced irisin in bone and systemic irisin administration reveal new regulatory mechanisms of bone metabolism. Bone Res. 2017, 5, 16056. [Google Scholar] [CrossRef]
- Huh, J.Y.; Panagiotou, G.; Mougios, V.; Brinkoetter, M.; Vamvini, M.T.; Schneider, B.E.; Mantzoros, C.S. Fndc5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and ii. Mrna expression and circulating concentrations in response to weight loss and exercise. Metabolism 2012, 61, 1725–1738. [Google Scholar] [CrossRef]
- Albrecht, E.; Schering, L.; Buck, F.; Vlach, K.; Schober, H.C.; Drevon, C.A.; Maak, S. Irisin: Still chasing shadows. Mol. Metab. 2020, 34, 124–135. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Traits | Number of Participants Measured | All Participants (Mean ± SD) | Non-Diabetic Participants (Mean ± SD) | Diabetic Participants (Mean ± SD) | p-Value (a) (for Differences in Mean Values between Diabetic and Non-Diabetic Participants) |
|---|---|---|---|---|---|
| Male:Female | 278 | 125:153 | 65:93 | 60:60 | 0.1772 |
| Age (in years) | 278 | 46.25 ± 12.38 | 42.04 ± 12.57 | 51.8 ± 9.70 | 3.03 × 10−12 |
| Height (in meter) | 278 | 1.64 ± 0.09 | 1.64 ± 0.09 | 1.65 ± 0.09 | 0.516 |
| Weight (in kg) | 278 | 81.40 ± 16.23 | 77.65 ± 16.49 | 86.27 ± 14.57 | 7.07 × 10−6 |
| BMI (kg/m2) | 278 | 29.93 ± 5.17 | 28.81 ± 5.46 | 31.41 ± 4.37 | 1.36 × 10−5 |
| WC (in cm) | 177 | 99.36 ± 13.36 | 93.95 ± 13.43 | 105.22 ± 10.57 | 3.58 × 10−9 |
| HDL (in mmol/L) | 260 | 1.20 ± 0.32 | 1.25 ± 0.31 | 1.13 ± 0.31 | 0.0018 |
| TC (b) (in mmol/L) | 272 | 5.27 ± 1.04 | 5.56 ± 1.42 | 5.40 ± 1.22 | 0.068 |
| LDL (b) (in mmol/L) | 269 | 3.38 ± 0.94 | 3.68 ± 1.39 | 3.51 ± 1.16 | 0.049 |
| Non-HDL (b) (in mmol/L) | 258 | 4.018 ± 1.05 | 4.34 ± 1.31 | 4.16 ± 1.18 | 0.031 |
| TG (in mmol/L) | 260 | 1.22 ± 0.59 | 1.07 ± 0.57 | 1.43 ± 0.56 | 7.09 × 10−7 |
| FPG (in mmol/L) | 241 | 5.77 ± 1.24 | 5.21 ± 0.63 | 6.73 ± 1.42 | 5.19 × 10−16 |
| HbA1 c (%) | 254 | 6.31 ± 1.29 | 5.61 ± 0.60 | 7.30 ± 1.36 | <2.2 × 10−16 |
| Irisin (ng/mL) | 219 | 556.95 ± 192.26 | 507.96 ± 170.77 | 620.89 ± 200.67 | 1.83 × 10−5 |
| IL7 (pg/mL) (b) | 163 | 13.23 ± 5.75 | 12.09 ± 5.44 | 14.86 ± 5.82 | 0.0026 |
| IL13 (pg/mL) | 158 | 9.67 ± 4.84 | 9.21 ± 4.95 | 10.33 ± 4.63 | 0.1491 |
| Insulin (pg/mL) | 195 | 14.90 ± 11.97 | 13.41 ± 11.19 | 16.83 ± 12.71 | 0.0511 |
| c-peptide (pg/mL) | 161 | 2.66 ± 1.73 | 2.76 ± 1.71 | 2.56 ± 1.75 | 0.4636 |
| ANGPTL3 (ng/mL) | 195 | 37.42 ± 10.29 | 36.77 ± 10.33 | 38.21 ± 10.24 | 0.3337 |
| TNFa (pg/mL) | 167 | 127.67 ± 32.16 | 125.31 ± 32.97 | 131.19 ± 30.81 | 0.2417 |
| Obese status | 278 | 135:143 | 62:96 | 73:47 | 0.00056 |
| Diabetes status | 278 | 120:158 | 0:158 | 120:0 | - |
| Anti-diabetic medication | 278 | 101(med):177 (no med) | 158 (no med):0 | 19 (No med):101 (med) | 0.001 |
| Lipid-lowering medication | 278 | 88(med):190 | 21(med):137 | 67(med):53 | 1.43 × 10−13 |
| Traits | SNP with Effect Allele | Correction | Sample Size (a) | β | p-Value (b) | Empirical p-Value (Pemp-Value) (b) |
|---|---|---|---|---|---|---|
| c-peptide | rs1748197 | R | 160 | −0.6976 | 0.000127 | 0.00679 |
| DM | 160 | −0.6944 | 0.000161 | 0.00939 | ||
| LLM | 160 | −0.6964 | 0.000138 | 0.00739 | ||
| rs12130333 | R | 161 | −0.9002 | 0.00032 | 0.0154 | |
| R + DM | 161 | −0.8991 | 0.000335 | 0.0174 | ||
| R + LLM | 161 | −0.9117 | 0.000288 | 0.0167 | ||
| Irisin | rs1748197 | R | 217 | −63.1 | 0.000299 | 0.0149 |
| DM | 216 | −67.78 | 9.58 × 10−5 | 0.0047 | ||
| LLM | 216 | −63 | 0.000357 | 0.0184 | ||
| rs12130333 | R | 218 | −72.61 | 0.002135 | 0.0979 | |
| R + DM | 217 | −70.87 | 0.002436 | 0.1184 | ||
| R + LLM | 217 | −74.3 | 0.001806 | 0.0898 |
| Trait | Haplotype (GC Forms the Reference Haplotype) | Frequency | β | p-Value (a) | Empirical p-Value (Pemp-Value) (a) |
|---|---|---|---|---|---|
| Irisin | AT | 0.134 | −74.9 | 0.00313 | 0.0122 |
| GT | 0.013 | −110 | 0.262 | 0.6224 | |
| AC | 0.221 | −36.4 | 0.0951 | 0.2853 | |
| GC | 0.632 | 68.2 | 0.000162 | 0.00059 | |
| c-peptide | AT | 0.134 | −1.45 | 0.0368 | 0.1363 |
| GT | 0.013 | −2.92 | 0.304 | 0.6326 | |
| AC | 0.221 | −0.49 | 0.413 | 0.7832 | |
| GC | 0.632 | 1.17 | 0.0182 | 0.0891 |
| Trait (Response variable) | Genotype and Interacting Trait (Predict Variable) | Estimate | Std. Error | p-Value | Adj. R-Square | Model p-Value |
|---|---|---|---|---|---|---|
| Model:c-peptide~rs1748197 + age + sex + IL13 + rs1748197*IL13 | ||||||
| c-peptide | (Intercept) | −0.2927 | 0.8453 | 0.7299 | 0.3416 | 4.84 × 10−9 |
| rs1748197 (GA + AA) | 1.5210 | 0.6446 | 0.0202 | |||
| IL13 | 0.261 | 0.046 | 1.96 × 10−7 | |||
| rs1748197 (GA + AA):IL13 | −0.2961 | 0.0584 | 1.88 × 10−6 | |||
| Model:Irisin~ rs1748197 + age + sex + TG + rs1748197*TG | ||||||
| Irisin | (Intercept) | 296.583 | 74.581 | 9.78 × 10−5 | 0.2163 | 2.27 × 10−10 |
| rs1748197 (GA + AA) | −161.75 | 57.993 | 0.0058 | |||
| TG | 66.04 | 32.31 | 0.042 | |||
| rs1748197 (GA + AA):TG | 80.551 | 41.507 | 0.053 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alanbaei, M.; Abu-Farha, M.; Hebbar, P.; Melhem, M.; Chandy, B.S.; Anoop, E.; Cherian, P.; Al-Khairi, I.; Alkayal, F.; Al-Mulla, F.; et al. ANGPTL3 Variants Associate with Lower Levels of Irisin and C-Peptide in a Cohort of Arab Individuals. Genes 2021, 12, 755. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050755
Alanbaei M, Abu-Farha M, Hebbar P, Melhem M, Chandy BS, Anoop E, Cherian P, Al-Khairi I, Alkayal F, Al-Mulla F, et al. ANGPTL3 Variants Associate with Lower Levels of Irisin and C-Peptide in a Cohort of Arab Individuals. Genes. 2021; 12(5):755. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050755
Chicago/Turabian StyleAlanbaei, Muath, Mohamed Abu-Farha, Prashantha Hebbar, Motasem Melhem, Betty S. Chandy, Emil Anoop, Preethi Cherian, Irina Al-Khairi, Fadi Alkayal, Fahd Al-Mulla, and et al. 2021. "ANGPTL3 Variants Associate with Lower Levels of Irisin and C-Peptide in a Cohort of Arab Individuals" Genes 12, no. 5: 755. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12050755