
Nanopore Sequencing Unveils Diverse Transcript Variants of the Epithelial Cell-Specific Transcription Factor Elf-3 in Human Malignancies


Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Total RNA Extraction and First-Strand cDNA Synthesis
2.3. PCR Amplification of ELF3 mRNA Transcripts
2.4. Library Preparation and Nanopore Sequencing
2.5. Post Processing and Bioinformatics Analysis
2.6. Validation of the Novel Splice Junctions
3. Results
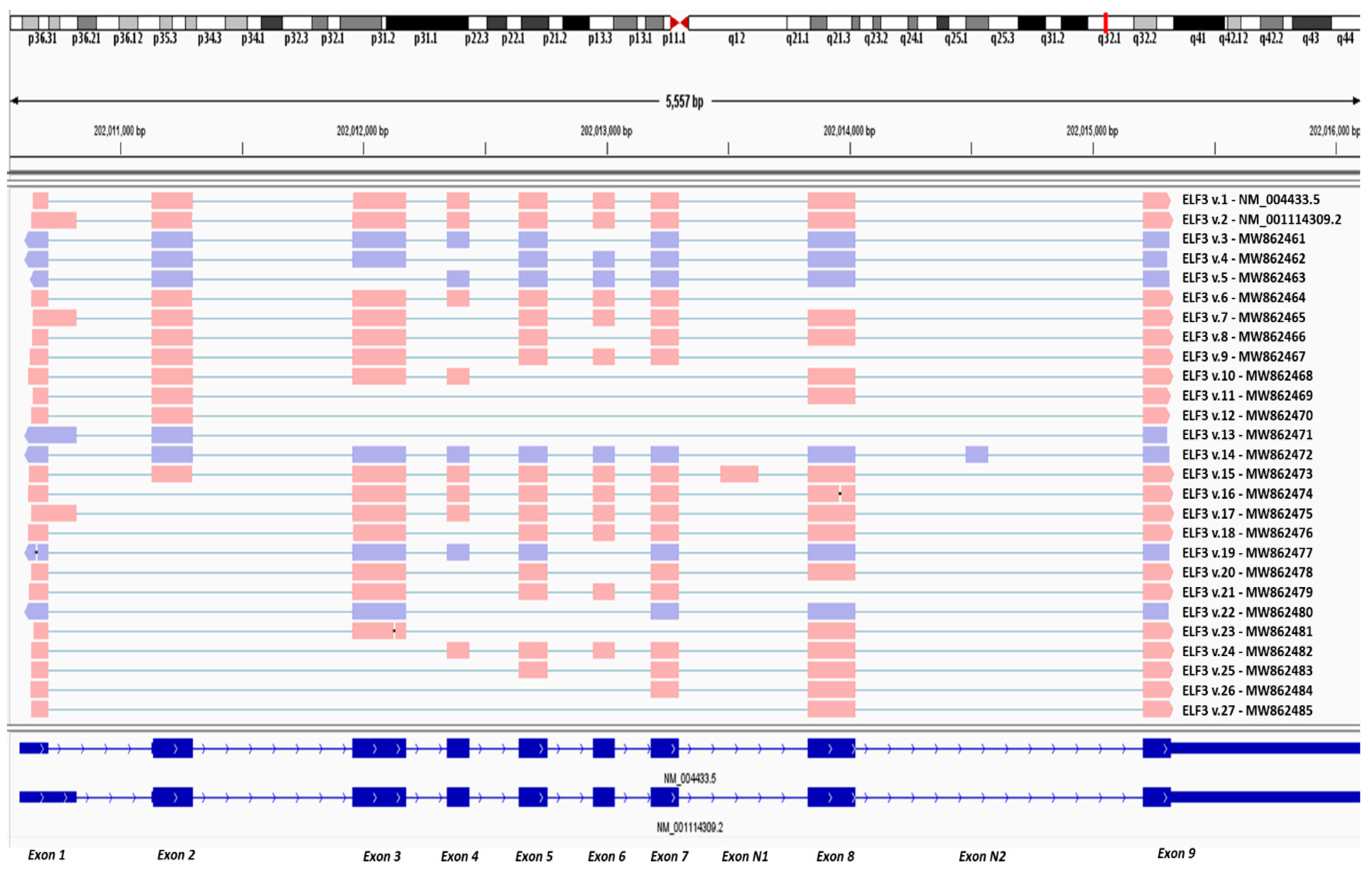
3.1. Nanopore Sequencing Reveals New Alternative Splicing Events of ELF3
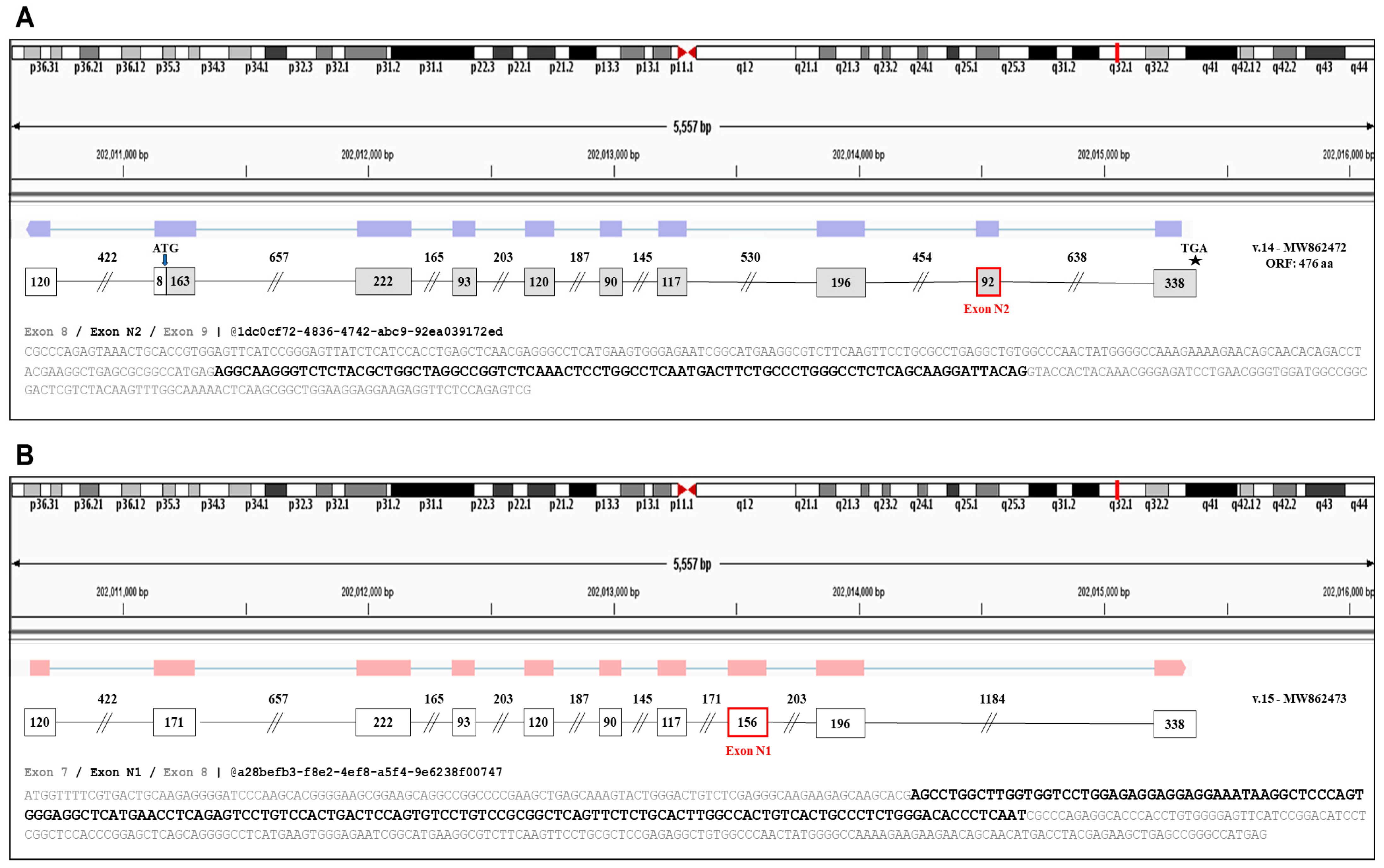
3.2. Detection of Two Novel Cryptic Exons of ELF3
3.3. Identification of Novel ELF3 Transcripts Sharing the Annotated Initiation Codon
3.4. Identification of Novel ELF3 Transcripts Bearing Novel Translation Initiation Codons
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef] [PubMed]
- Baralle, F.E.; Giudice, J. Alternative splicing as a regulator of development and tissue identity. Nat. Rev. Mol. Cell Biol. 2017, 18, 437–451. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.T.; Sandberg, R.; Luo, S.; Khrebtukova, I.; Zhang, L.; Mayr, C.; Kingsmore, S.F.; Schroth, G.P.; Burge, C.B. Alternative isoform regulation in human tissue transcriptomes. Nature 2008, 456, 470–476. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Weiss, W.A. Alternative splicing in cancer: Implications for biology and therapy. Oncogene 2015, 34, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Pinto, S.M.; Getnet, D.; Nirujogi, R.S.; Manda, S.S.; Chaerkady, R.; Madugundu, A.K.; Kelkar, D.S.; Isserlin, R.; Jain, S.; et al. A draft map of the human proteome. Nature 2014, 509, 575–581. [Google Scholar] [CrossRef] [Green Version]
- Karapetyan, A.R.; Buiting, C.; Kuiper, R.A.; Coolen, M.W. Regulatory roles for long ncrna and mrna. Cancers 2013, 5, 462–490. [Google Scholar] [CrossRef]
- Kalsotra, A.; Cooper, T.A. Functional consequences of developmentally regulated alternative splicing. Nat. Rev. Genet. 2011, 12, 715–729. [Google Scholar] [CrossRef]
- Paronetto, M.P.; Passacantilli, I.; Sette, C. Alternative splicing and cell survival: From tissue homeostasis to disease. Cell Death Differ. 2016, 23, 1919–1929. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, D.G.; Woolard, J.; Amin, E.M.; Konopatskaya, O.; Saleem, M.A.; Churchill, A.J.; Ladomery, M.R.; Harper, S.J.; Bates, D.O. Expression of pro- and anti-angiogenic isoforms of vegf is differentially regulated by splicing and growth factors. J. Cell Sci. 2008, 121, 3487–3495. [Google Scholar] [CrossRef] [Green Version]
- Cloutier, P.; Toutant, J.; Shkreta, L.; Goekjian, S.; Revil, T.; Chabot, B. Antagonistic effects of the srp30c protein and cryptic 5 ′ splice sites on the alternative splicing of the apoptotic regulator bcl-x. J. Biol. Chem. 2008, 283, 21315–21324. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhou, M.; Shi, B.; Zhang, Q.; Jiang, H.; Sun, Y.; Liu, J.; Zhou, K.; Yao, M.; Gu, J.; et al. Identification of an exon 4-deletion variant of epidermal growth factor receptor with increased metastasis-promoting capacity. Neoplasia 2011, 13, 461–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yae, T.; Tsuchihashi, K.; Ishimoto, T.; Motohara, T.; Yoshikawa, M.; Yoshida, G.J.; Wada, T.; Masuko, T.; Mogushi, K.; Tanaka, H.; et al. Alternative splicing of cd44 mrna by esrp1 enhances lung colonization of metastatic cancer cell. Nat. Commun. 2012, 3, 883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warzecha, C.C.; Jiang, P.; Amirikian, K.; Dittmar, K.A.; Lu, H.; Shen, S.; Guo, W.; Xing, Y.; Carstens, R.P. An esrp-regulated splicing programme is abrogated during the epithelial–mesenchymal transition. EMBO J. 2010, 29, 3286–3300. [Google Scholar] [CrossRef] [PubMed]
- Adamopoulos, P.G.; Kontos, C.K.; Tsiakanikas, P.; Scorilas, A. Identification of novel alternative splice variants of the bcl2l12 gene in human cancer cells using next-generation sequencing methodology. Cancer Lett 2016, 373, 119–129. [Google Scholar] [CrossRef]
- Adamopoulos, P.G.; Kontos, C.K.; Scorilas, A. Molecular cloning of novel transcripts of human kallikrein-related peptidases 5, 6, 7, 8 and 9 (klk5–klk9), using next-generation sequencing. Sci. Rep. 2017, 7, 17299. [Google Scholar] [CrossRef] [Green Version]
- Adamopoulos, P.G.; Tsiakanikas, P.; Adam, E.E.; Scorilas, A. Unraveling novel survivin mrna transcripts in cancer cells using an in-house developed targeted high-throughput sequencing approach. Genomics 2021, 113, 573–581. [Google Scholar] [CrossRef]
- Mak, A.C.; Lai, Y.Y.; Lam, E.T.; Kwok, T.P.; Leung, A.K.; Poon, A.; Mostovoy, Y.; Hastie, A.R.; Stedman, W.; Anantharaman, T.; et al. ; et al. Genome-wide structural variation detection by genome mapping on nanochannel arrays. Genetics 2016, 202, 351–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.; Popic, V.; Batzoglou, S. Short read alignment with populations of genomes. Bioinformatics 2013, 29, i361–i370. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Gurtowski, J.; Yoo, S.; Nattestad, M.; Marcus, S.; Goodwin, S.; Richard McCombie, W.; Schatz, M.C. Third-generation sequencing and the future of genomics. bioRxiv 2016. bioRxiv:048603. [Google Scholar]
- Van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The third revolution in sequencing technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, S.; McPherson, J.D.; McCombie, W.R. Coming of age: Ten years of next-generation sequencing technologies. Nat. Rev. Genet. 2016, 17, 333–351. [Google Scholar] [CrossRef] [PubMed]
- Michael, T.P.; Jupe, F.; Bemm, F.; Motley, S.T.; Sandoval, J.P.; Lanz, C.; Loudet, O.; Weigel, D.; Ecker, J.R. High contiguity arabidopsis thaliana genome assembly with a single nanopore flow cell. Nat. Commun. 2018, 9, 541. [Google Scholar] [CrossRef] [Green Version]
- Roberts, R.J.; Carneiro, M.O.; Schatz, M.C. The advantages of smrt sequencing. Genome Biol. 2013, 14, 405. [Google Scholar] [CrossRef]
- Chaisson, M.J.; Huddleston, J.; Dennis, M.Y.; Sudmant, P.H.; Malig, M.; Hormozdiari, F.; Antonacci, F.; Surti, U.; Sandstrom, R.; Boitano, M.; et al. ; et al. Resolving the complexity of the human genome using single-molecule sequencing. Nature 2015, 517, 608–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharon, D.; Tilgner, H.; Grubert, F.; Snyder, M. A single-molecule long-read survey of the human transcriptome. Nat. Biotechnol. 2013, 31, 1009–1014. [Google Scholar] [CrossRef]
- Oettgen, P.; Alani, R.M.; Barcinski, M.A.; Brown, L.; Akbarali, Y.; Boltax, J.; Kunsch, C.; Munger, K.; Libermann, T.A. Isolation and characterization of a novel epithelium-specific transcription factor, ese-1, a member of the ets family. Mol. Cell. Biol. 1997, 17, 4419–4433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kas, K.; Finger, E.; Grall, F.; Gu, X.; Akbarali, Y.; Boltax, J.; Weiss, A.; Oettgen, P.; Kapeller, R.; Libermann, T.A. Ese-3, a novel member of an epithelium-specific ets transcription factor subfamily, demonstrates different target gene specificity from ese-1. J. Biol. Chem. 2000, 275, 2986–2998. [Google Scholar] [CrossRef] [Green Version]
- Tymms, M.J.; Ng, A.Y.; Thomas, R.S.; Schutte, B.C.; Zhou, J.; Eyre, H.J.; Sutherland, G.R.; Seth, A.; Rosenberg, M.; Papas, T.; et al. A novel epithelial-expressed ets gene, elf3: Human and murine cdna sequences, murine genomic organization, human mapping to 1q32.2 and expression in tissues and cancer. Oncogene 1997, 15, 2449–2462. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.H.; Scott, G.K.; Kuo, W.L.; Xiong, X.; Suzdaltseva, Y.; Park, J.W.; Sayre, P.; Erny, K.; Collins, C.; Gray, J.W.; et al. Esx: A structurally unique ets overexpressed early during human breast tumorigenesis. Oncogene 1997, 14, 1617–1622. [Google Scholar] [CrossRef] [Green Version]
- Asada, S.; Choi, Y.; Uesugi, M. A gene-expression inhibitor that targets an α-helix-mediated protein interaction. J. Am. Chem. Soc. 2003, 125, 4992–4993. [Google Scholar] [CrossRef]
- Asada, S.; Choi, Y.; Yamada, M.; Wang, S.C.; Hung, M.C.; Qin, J.; Uesugi, M. External control of her2 expression and cancer cell growth by targeting a ras-linked coactivator. Proc. Natl. Acad. Sci. USA 2002, 99, 12747–12752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.G.; Yi, Y.; Kim, Y.S.; Kato, M.; Chang, J.; Chung, H.W.; Hahm, K.B.; Yang, H.K.; Rhee, H.H.; Bang, Y.J.; et al. A novel ets-related transcription factor, ert/esx/ese-1, regulates expression of the transforming growth factor-β type ii receptor. J. Biol. Chem. 1998, 273, 110–117. [Google Scholar] [CrossRef] [Green Version]
- Eckel, K.L.; Tentler, J.J.; Cappetta, G.J.; Diamond, S.E.; Gutierrez-Hartmann, A. The epithelial-specific ets transcription factor esx/ese-1/elf-3 modulates breast cancer-associated gene expression. DNA Cell Biol. 2003, 22, 79–94. [Google Scholar] [CrossRef]
- Feldman, R.J.; Sementchenko, V.I.; Watson, D.K. The epithelial-specific ets factors occupy a unique position in defining epithelial proliferation, differentiation and carcinogenesis. Anticancer Res. 2003, 23, 2125–2131. [Google Scholar] [PubMed]
- Neve, R.; Chang, C.H.; Scott, G.K.; Wong, A.; Friis, R.R.; Hynes, N.E.; Benz, C.C. The epithelium-specific ets transcription factor esx is associated with mammary gland development and involution. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1998, 12, 1541–1550. [Google Scholar] [CrossRef]
- Ng, A.Y.; Waring, P.; Ristevski, S.; Wang, C.; Wilson, T.; Pritchard, M.; Hertzog, P.; Kola, I. Inactivation of the transcription factor elf3 in mice results in dysmorphogenesis and altered differentiation of intestinal epithelium. Gastroenterology 2002, 122, 1455–1466. [Google Scholar] [CrossRef] [PubMed]
- Cabral, A.; Fischer, D.F.; Vermeij, W.P.; Backendorf, C. Distinct functional interactions of human skn-1 isoforms with ese-1 during keratinocyte terminal differentiation. J. Biol. Chem. 2003, 278, 17792–17799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oettgen, P.; Kas, K.; Dube, A.; Gu, X.; Grall, F.; Thamrongsak, U.; Akbarali, Y.; Finger, E.; Boltax, J.; Endress, G.; et al. Characterization of ese-2, a novel ese-1-related ets transcription factor that is restricted to glandular epithelium and differentiated keratinocytes. J. Biol. Chem. 1999, 274, 29439–29452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, S.P.; Vuong, H.; Adiseshaiah, P. Interplay between proximal and distal promoter elements is required for squamous differentiation marker induction in the bronchial epithelium: Role for ese-1, sp1, and ap-1 proteins. J. Biol. Chem. 2003, 278, 21378–21387. [Google Scholar] [CrossRef] [Green Version]
- Jobling, A.I.; Fang, Z.; Koleski, D.; Tymms, M.J. Expression of the ets transcription factor elf3 in the retinal pigment epithelium. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3530–3537. [Google Scholar]
- Sengez, B.; Aygün, I.; Shehwana, H.; Toyran, N.; Tercan Avci, S.; Konu, O.; Stemmler, M.P.; Alotaibi, H. The transcription factor elf3 is essential for a successful mesenchymal to epithelial transition. Cells 2019, 8, 858. [Google Scholar] [CrossRef] [Green Version]
- Neve, R.M.; Ylstra, B.; Chang, C.H.; Albertson, D.G.; Benz, C.C. Erbb2 activation of esx gene expression. Oncogene 2002, 21, 3934–3938. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, N.; Yoshida, S.; Araie, M.; Handa, H.; Nabeshima, Y. Ets family transcription factor ese-1 is expressed in corneal epithelial cells and is involved in their differentiation. Mech. Dev. 2000, 97, 27–34. [Google Scholar] [CrossRef]
- Schedin, P.J.; Eckel-Mahan, K.L.; McDaniel, S.M.; Prescott, J.D.; Brodsky, K.S.; Tentler, J.J.; Gutierrez-Hartmann, A. Esx induces transformation and functional epithelial to mesenchymal transition in mcf-12a mammary epithelial cells. Oncogene 2004, 23, 1766–1779. [Google Scholar] [CrossRef] [Green Version]
- Prescott, J.D.; Koto, K.S.; Singh, M.; Gutierrez-Hartmann, A. The ets transcription factor ese-1 transforms mcf-12a human mammary epithelial cells via a novel cytoplasmic mechanism. Mol. Cell. Biol. 2004, 24, 5548–5564. [Google Scholar] [CrossRef] [Green Version]
- Kopp, J.L.; Wilder, P.J.; Desler, M.; Kinarsky, L.; Rizzino, A. Different domains of the transcription factor elf3 are required in a promoter-specific manner and multiple domains control its binding to DNA. J. Biol. Chem. 2007, 282, 3027–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidel, J.J.; Graves, B.J. An erk2 docking site in the pointed domain distinguishes a subset of ets transcription factors. Genes Dev. 2002, 16, 127–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.H.; Scott, G.K.; Baldwin, M.A.; Benz, C.C. Exon 4-encoded acidic domain in the epithelium-restricted ets factor, esx, confers potent transactivating capacity and binds to tata-binding protein (tbp). Oncogene 1999, 18, 3682–3695. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Holt, K.E. Performance of neural network basecalling tools for oxford nanopore sequencing. Genome Biol. 2019, 20, 129. [Google Scholar] [CrossRef] [Green Version]
- Li, H. Minimap2: Pairwise alignment for nucleotide sequences. Bioinformatics 2018, 34, 3094–3100. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The sequence alignment/map format and samtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorvaldsdottir, H.; Robinson, J.T.; Mesirov, J.P. Integrative genomics viewer (igv): High-performance genomics data visualization and exploration. Brief. Bioinform 2013, 14, 178–192. [Google Scholar] [CrossRef] [Green Version]
- Depledge, D.P.; Srinivas, K.P.; Sadaoka, T.; Bready, D.; Mori, Y.; Placantonakis, D.G.; Mohr, I.; Wilson, A.C. Direct RNA sequencing on nanopore arrays redefines the transcriptional complexity of a viral pathogen. Nat. Commun. 2019, 10, 754. [Google Scholar] [CrossRef] [Green Version]
- Brembeck, F.H.; Opitz, O.G.; Libermann, T.A.; Rustgi, A.K. Dual function of the epithelial specific ets transcription factor, elf3, in modulating differentiation. Oncogene 2000, 19, 1941–1949. [Google Scholar] [CrossRef] [Green Version]
- Kar, A.; Gutierrez-Hartmann, A. Ese-1/elf3 mrna expression associates with poor survival outcomes in her2(+) breast cancer patients and is critical for tumorigenesis in her2(+) breast cancer cells. Oncotarget 2017, 8, 69622–69640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Chen, W.; Zhang, X.; Hu, L.; Tang, G.; Kong, J.; Wang, Z. E26 transformation (ets)-specific related transcription factor-3 (elf3) orchestrates a positive feedback loop that constitutively activates the mapk/erk pathway to drive thyroid cancer. Oncol. Rep. 2019, 41, 570–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Splice Junction between Known Exons | Nanopore Sequencing Reads Confirming Each Splice Site | |
|---|---|---|
| Annotated | Exon 1 (120 bp)–Exon 2 | 205.399 |
| Exon 1 (234 bp)–Exon 2 | 3.328 | |
| Exon 2–Exon 3 | 206.117 | |
| Exon 3–Exon 4 | 210.848 | |
| Exon 4–Exon 5 | 188.589 | |
| Exon 5–Exon 6 | 221.945 | |
| Exon 6–Exon 7 | 218.907 | |
| Exon 7–Exon 8 | 213.796 | |
| Exon 8–Exon 9 | 207.222 | |
| Novel | Exon 1 (120 bp)–Exon 3 | 11.469 |
| Exon 1 (120 bp)–Exon 4 | 2.138 | |
| Exon 1 (120 bp)–Exon 5 | 5.321 | |
| Exon 1 (120 bp)–Exon 7 | 1.114 | |
| Exon 1 (120 bp)–Exon 8 | 1.627 | |
| Exon 1 (234 bp)–Exon 3 | 22.197 | |
| Exon 2–Exon 4 | 1.274 | |
| Exon 2–Exon 8 | 6.339 | |
| Exon 2–Exon 9 | 1.828 | |
| Exon 3–Exon 5 | 4.211 | |
| Exon 3–Exon 7 | 828 | |
| Exon 3–Exon 8 | 8.047 | |
| Exon 4–Exon 8 | 4.896 | |
| Exon 5–Exon 7 | 140.394 | |
| Exon 7–Exon 9 | 1.495 |
| Novel Transcripts | Elf-3 Domains | ||||
|---|---|---|---|---|---|
| PNT | TAD | SAR | A/T | ETS | |
| v.3 | ✓ | ✓ | - | ✓ | ✓ |
| v.4 | ✓ | - | ✓ | ✓ | ✓ |
| v.5 | - | ✓ | ✓ | ✓ | ✓ |
| v.6 | ✓ | ✓ | ✓ | ✓ | - |
| v.7 | ✓ | - | ✓ | ✓ | ✓ |
| v.8 | ✓ | - | - | ✓ | ✓ |
| v.9 | ✓ | - | ✓ | ✓ | - |
| v.10 | ✓ | ✓ | - | - | ✓ |
| v.11 | - | - | - | - | ✓ |
| v.12 | - | - | - | - | - |
| v.13 | - | - | - | - | - |
| v.14 | ✓ | ✓ | ✓ | ✓ | - |
| v.16 | - | ✓ | ✓ | ✓ | ✓ |
| v.17 | - | ✓ | ✓ | ✓ | ✓ |
| v.18 | - | - | ✓ | ✓ | ✓ |
| v.19 | - | ✓ | - | ✓ | ✓ |
| v.20 | - | - | - | ✓ | ✓ |
| v.21 | - | - | ✓ | ✓ | - |
| v.22 | - | - | - | ✓ | ✓ |
| v.23 | - | - | - | - | ✓ |
| v.24 | - | - | ✓ | ✓ | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boti, M.A.; Adamopoulos, P.G.; Tsiakanikas, P.; Scorilas, A. Nanopore Sequencing Unveils Diverse Transcript Variants of the Epithelial Cell-Specific Transcription Factor Elf-3 in Human Malignancies. Genes 2021, 12, 839. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060839
Boti MA, Adamopoulos PG, Tsiakanikas P, Scorilas A. Nanopore Sequencing Unveils Diverse Transcript Variants of the Epithelial Cell-Specific Transcription Factor Elf-3 in Human Malignancies. Genes. 2021; 12(6):839. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060839
Chicago/Turabian StyleBoti, Michaela A., Panagiotis G. Adamopoulos, Panagiotis Tsiakanikas, and Andreas Scorilas. 2021. "Nanopore Sequencing Unveils Diverse Transcript Variants of the Epithelial Cell-Specific Transcription Factor Elf-3 in Human Malignancies" Genes 12, no. 6: 839. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060839