
New Bird Sexing Strategy Developed in the Order Psittaciformes Involves Multiple Markers to Avoid Sex Misidentification: Debunked Myth of the Universal DNA Marker

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Extraction and Biological Samples
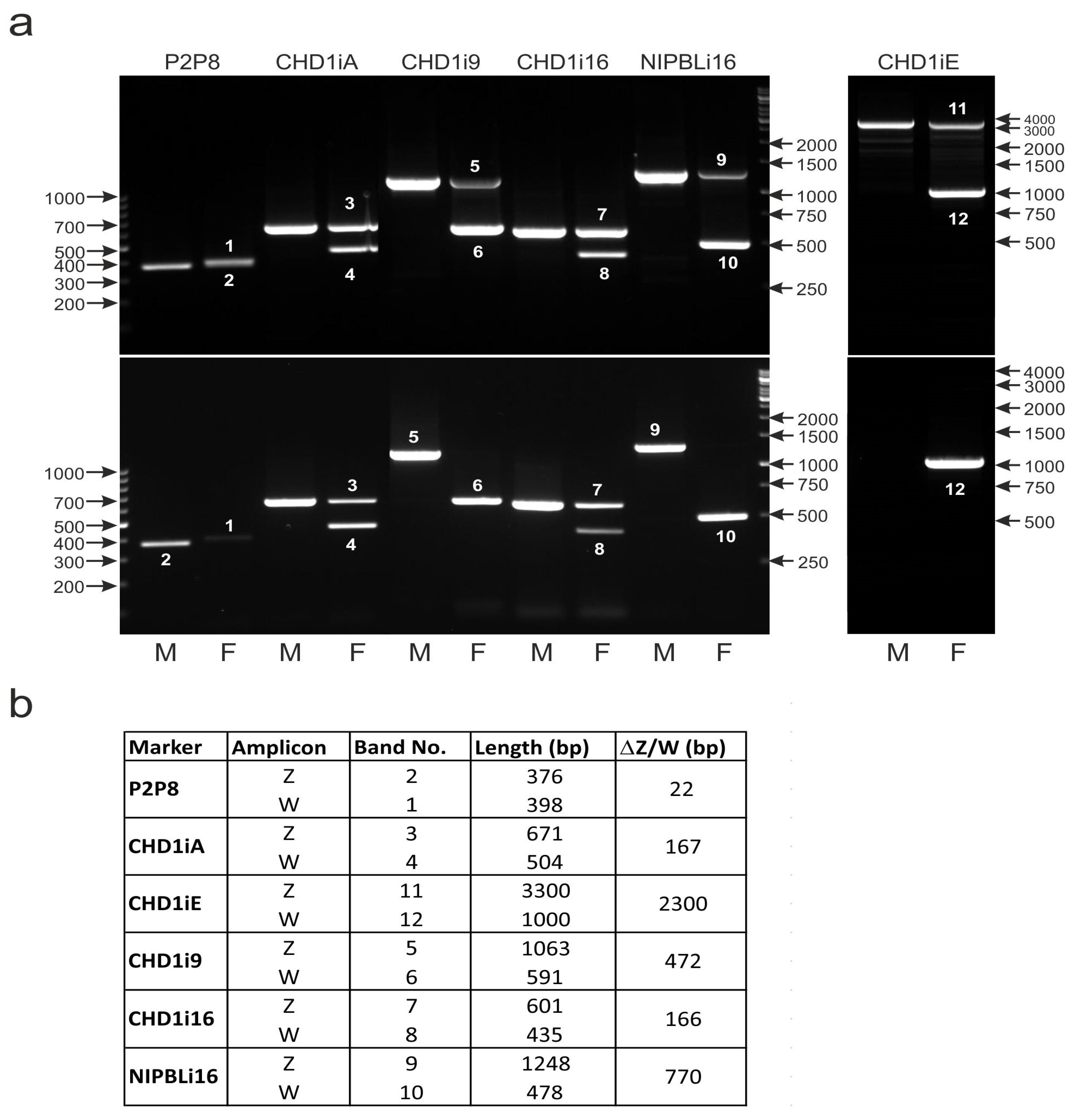
2.2. DNA Amplification
3. Results and Discussion
3.1. The Set of Parrot Species Representative for Phylogenetic Divergence of Psittaciformes Order
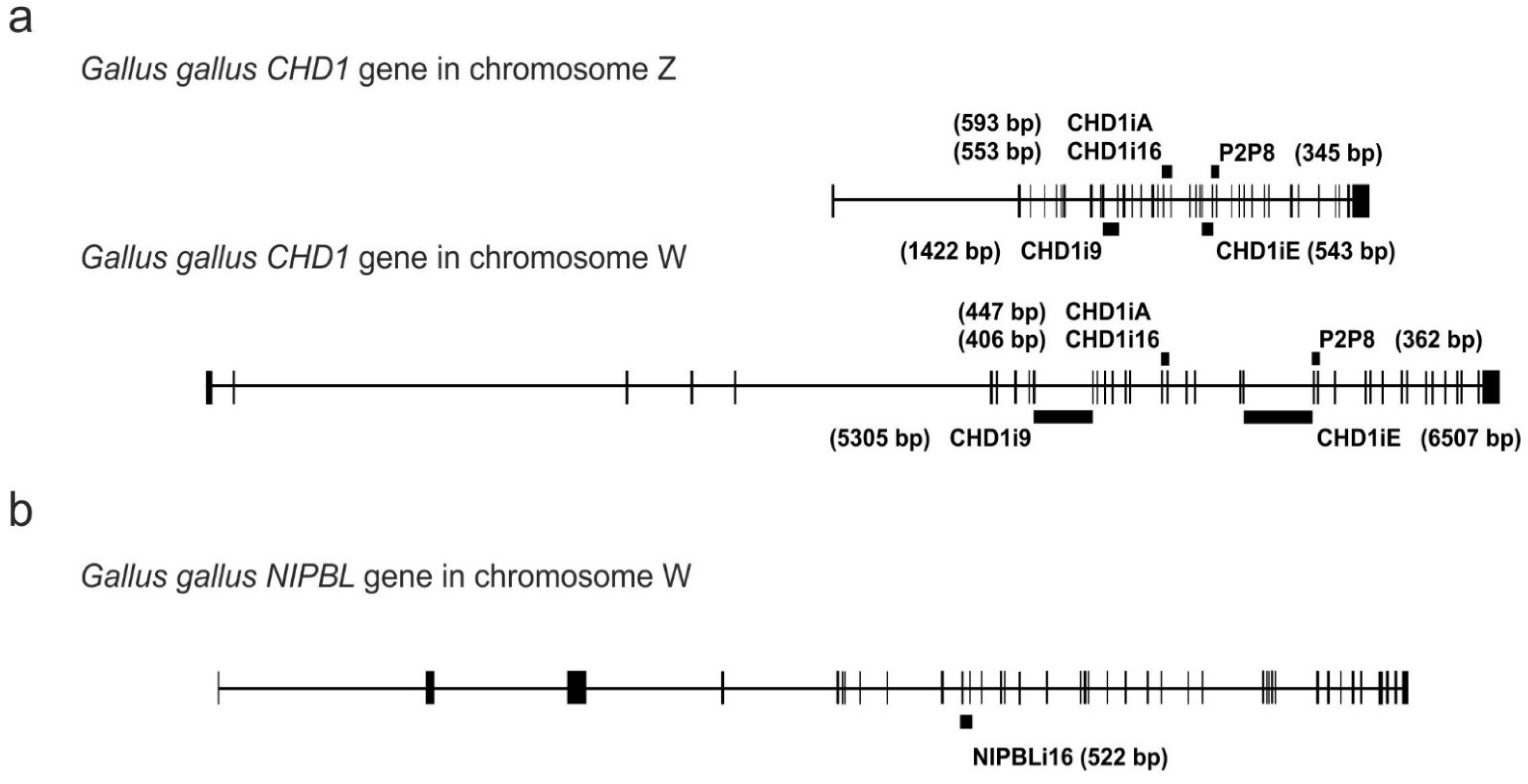
3.2. Genomic Localization of the Analyzed Markers
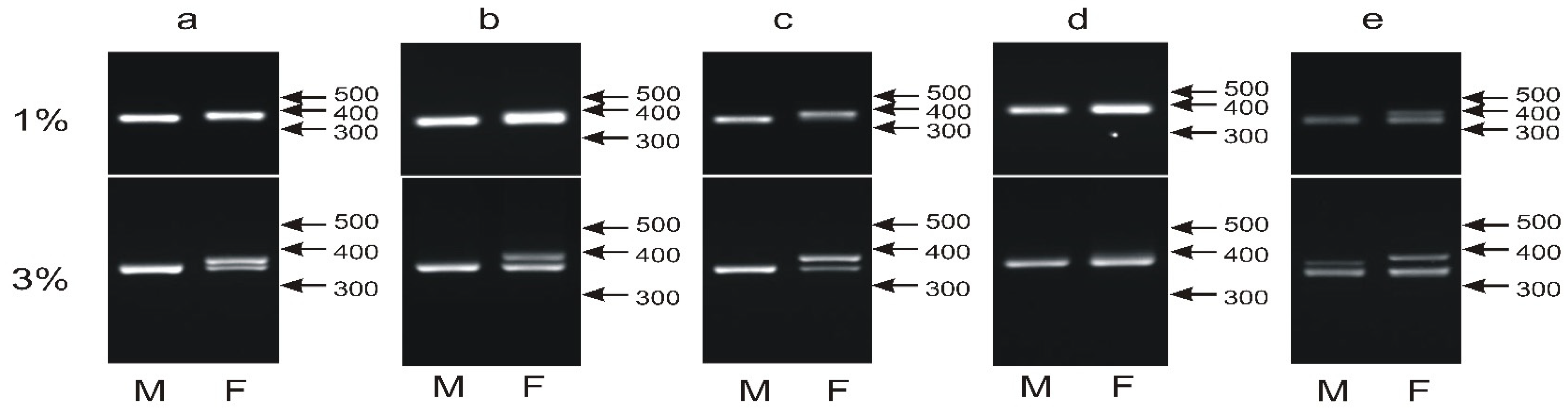
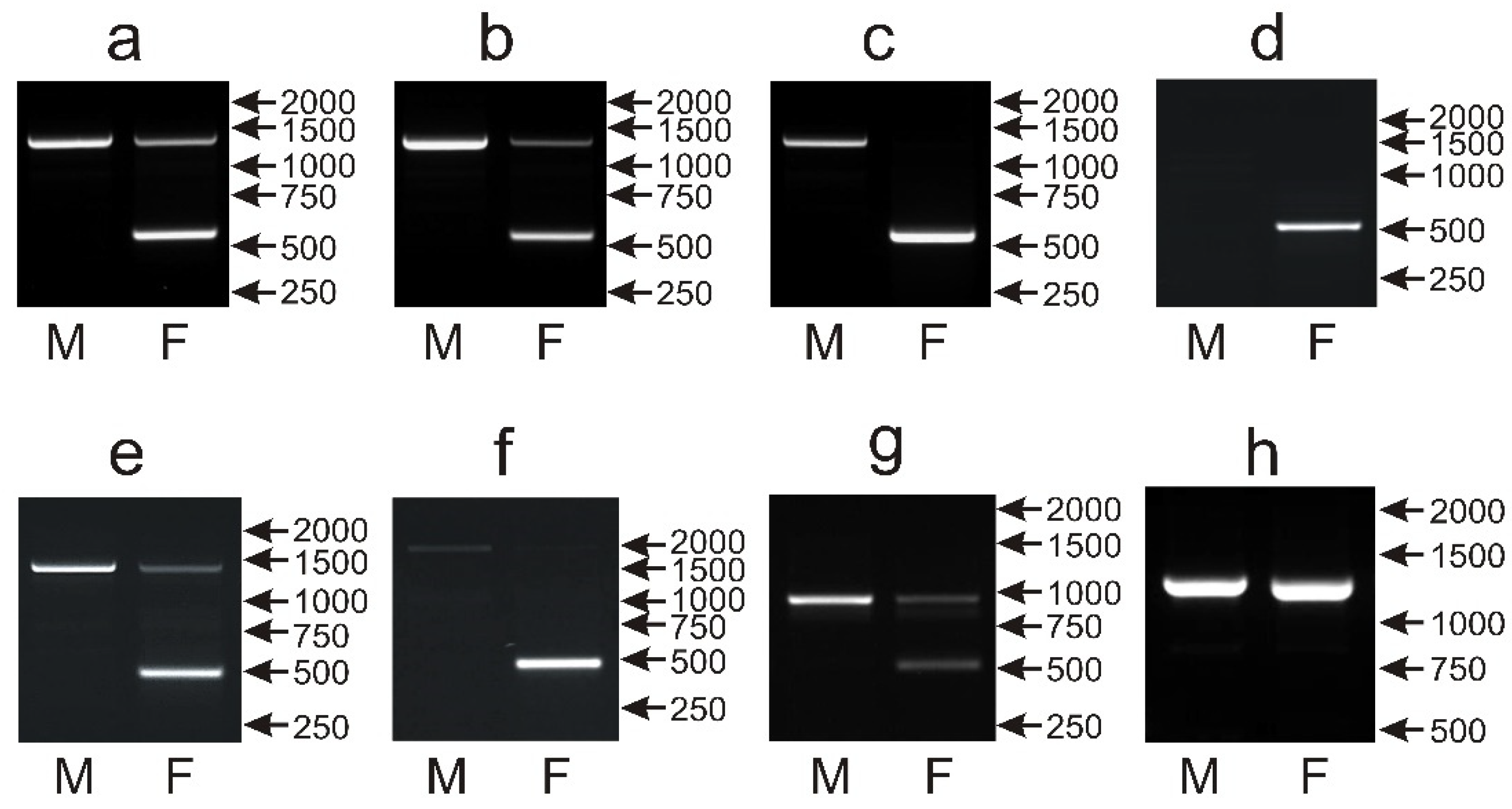
3.3. The Male- and Female-Specific Patterns of the P2P8 Marker Are Difficult to Distinguish
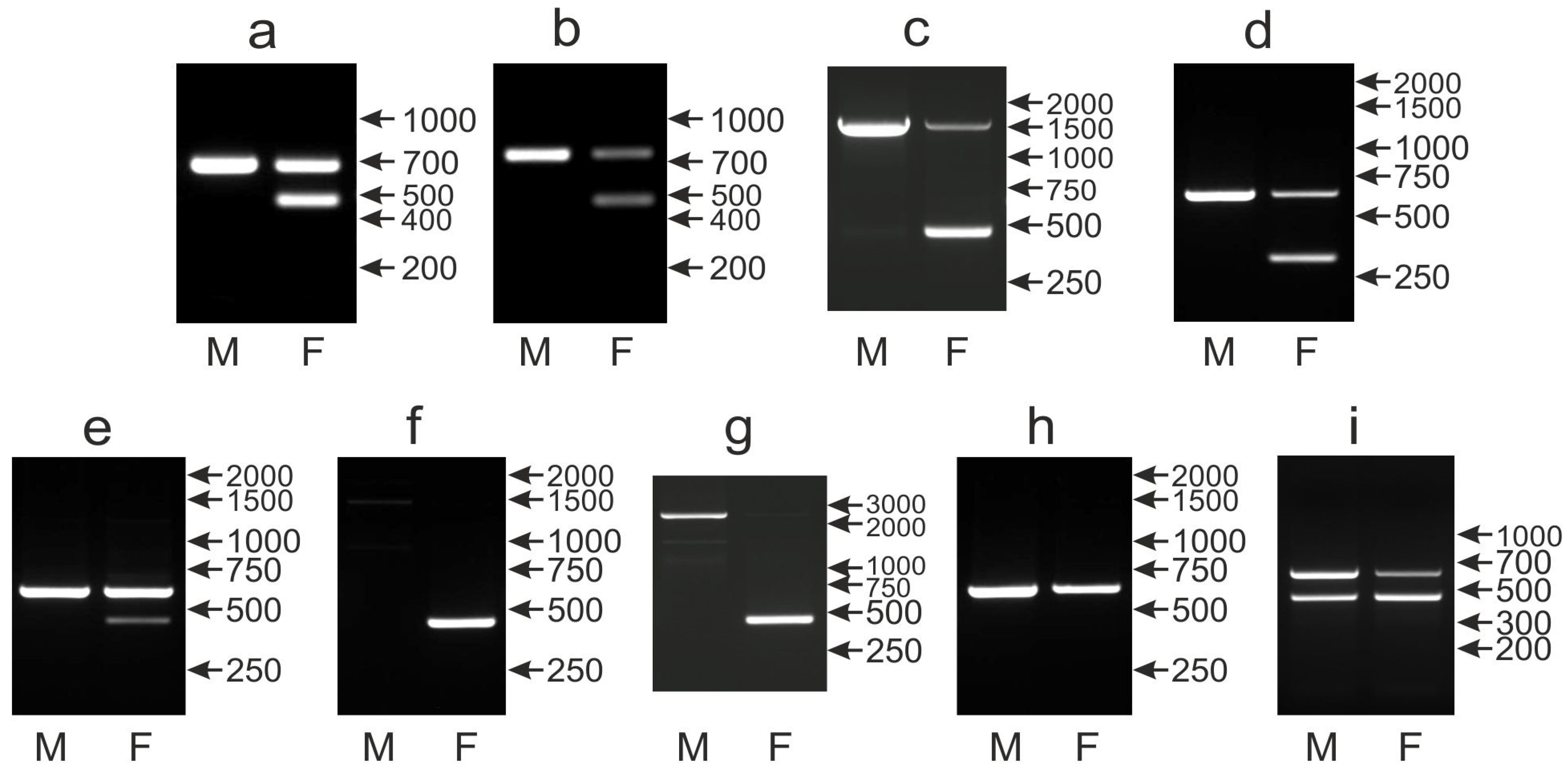
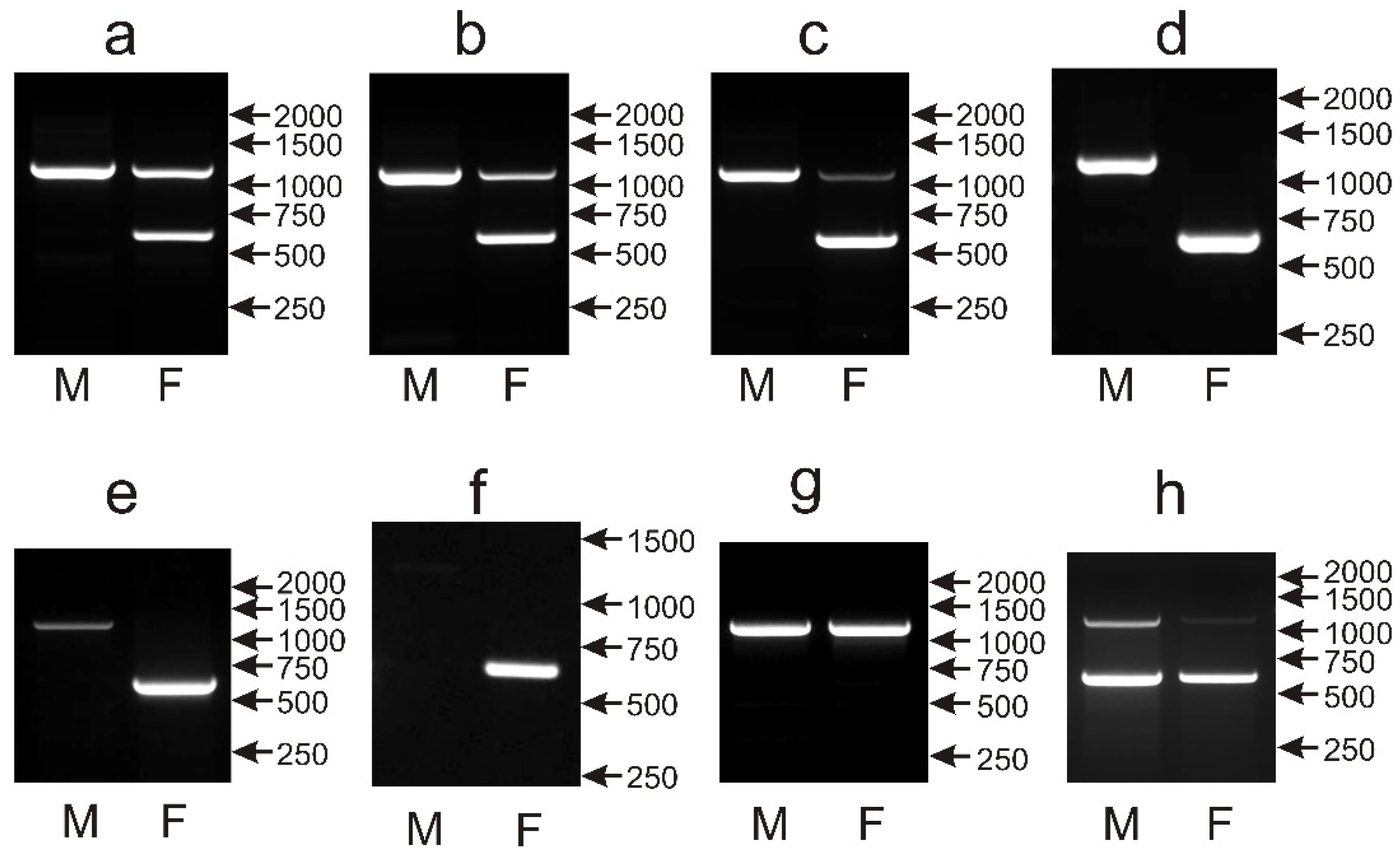
3.4. The CHD1iA Marker Shows Limitations in Sex Determination While Tested in New Species
3.5. Application of New CHD1iE Marker Is Difficult Due to Technical Drawbacks in PCR
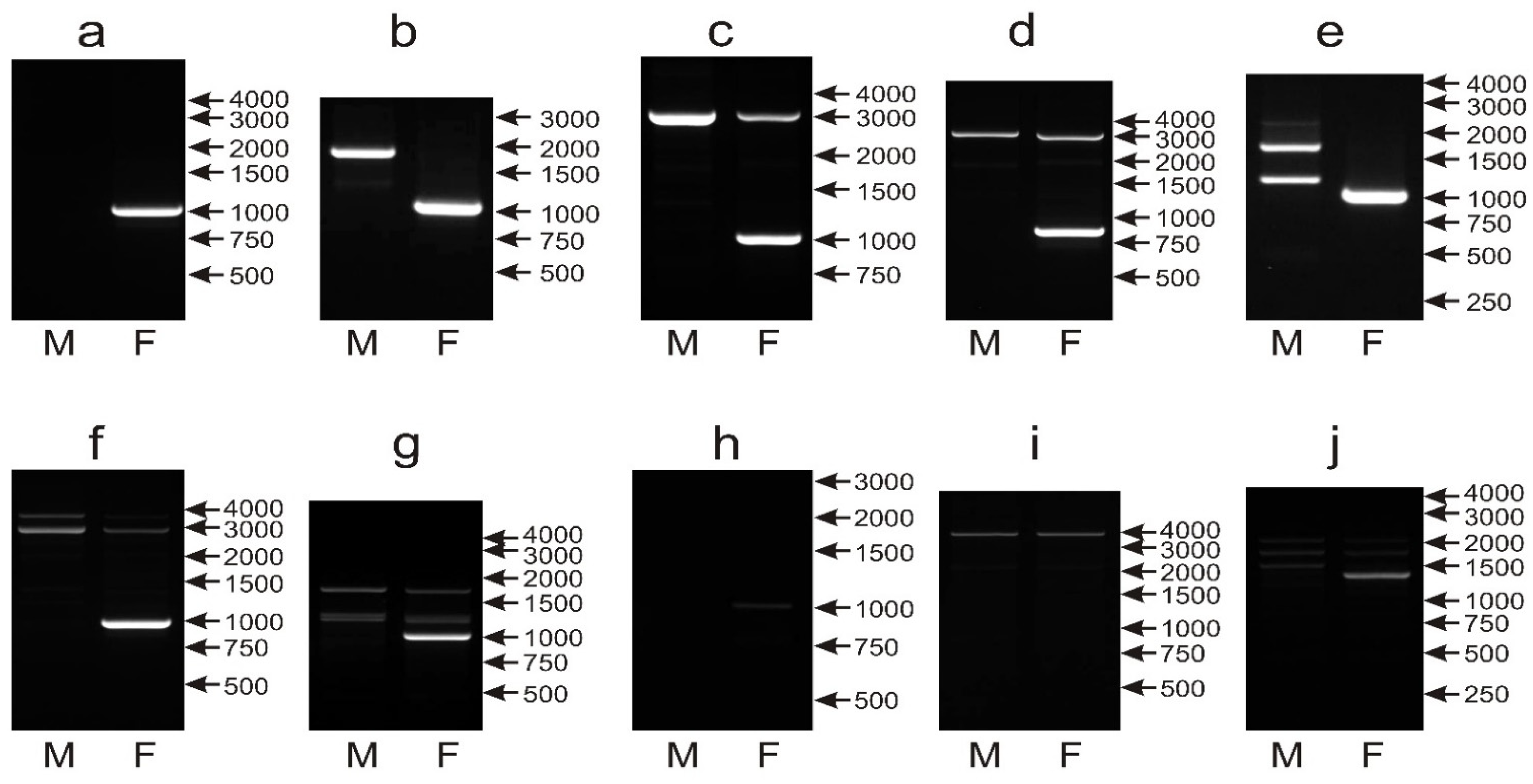
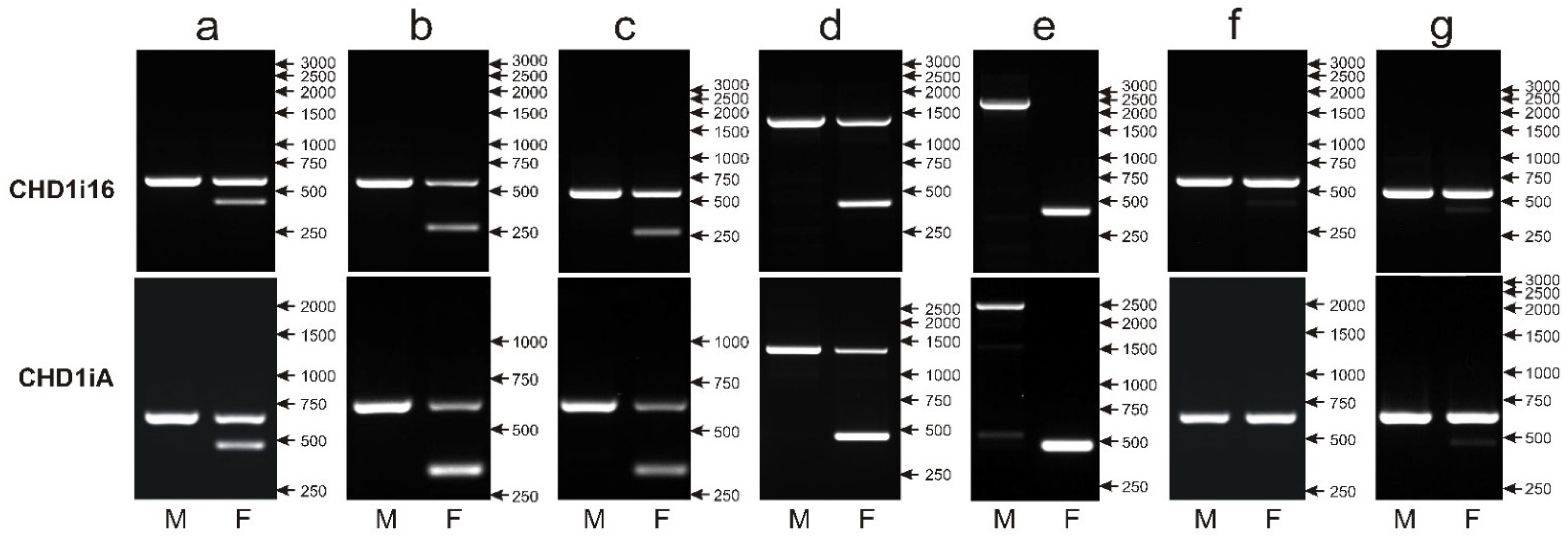
3.6. None of the CHD1i16, CHD1i9 and NIPBLi16 Markers Turn Universal When Applied to the Sexing of a Large Number of Parrot Species
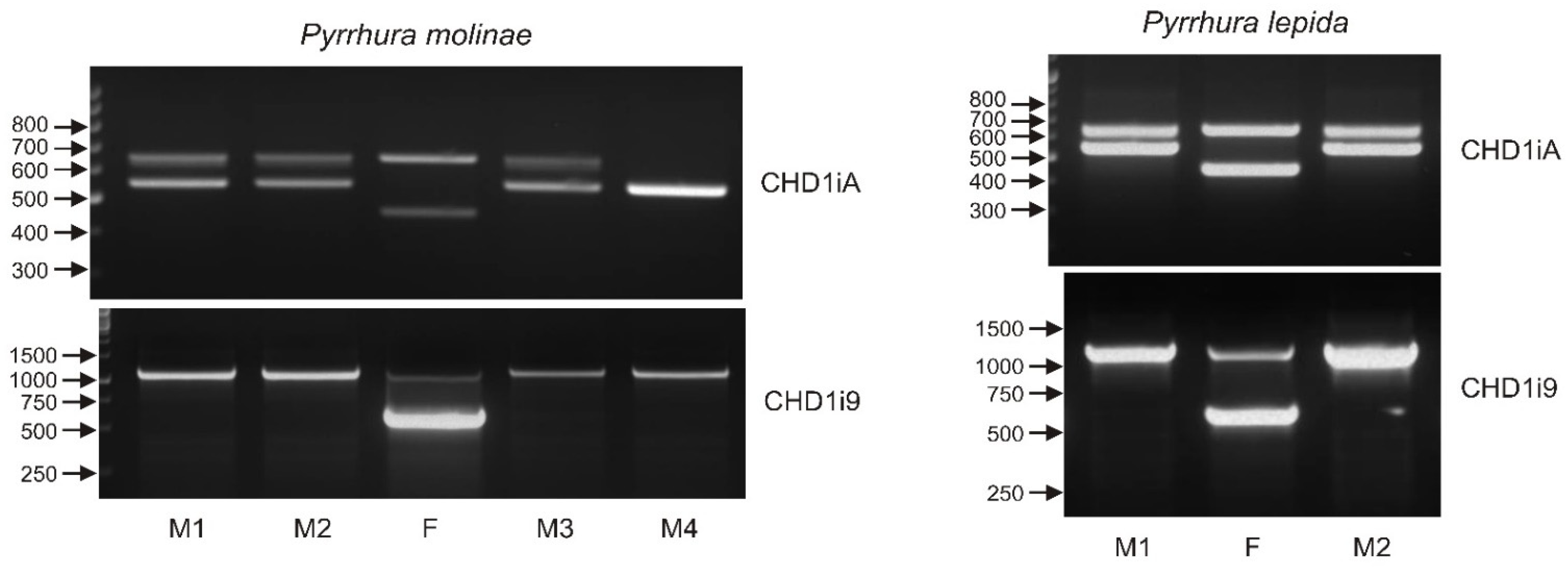
3.7. Sexing with Only One Marker May Lead to Sex Misidentification—The Case of Genus Pyrrhura
3.8. Molecular Markers Detecting Polymorphisms of CHD1 and NIPBL Genes Can Be Amplified via Direct PCR without DNA Isolation from Blood
3.9. Strategy to Sex Birds with Combination of Markers
3.10. There Is No Universal Marker
3.11. Sex Misidentification May Occur Due to Chromosomal Rearrangements
3.12. Application of Multiple Markers Ensures Effective and Reliable Sexing
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Olah, G.; Butchart, S.; Symes, A.; Medina, I.; Cunningham, R.; Brightsmith, D.; Heinsohn, R. Ecological and socio-economic factors affecting extinction risk in parrots. Biodivers. Conserv. 2016, 25, 205–223. [Google Scholar] [CrossRef]
- Toft, C.A.; Wright, T.F. Parrots of the Wild A Natural History of the World’s Most Captivating Birds; University of California Press: Berkeley, CA, USA, 2015. [Google Scholar]
- Heinsohn, R.; Buchanan, K.L.; Joseph, L. Parrots move to centre stage in conservation and evolution. Emu Austral Ornithol. 2018, 118, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bosé, M.; Le Gouar, P.; Arthur, C.; Lambourdie‘re, J.; Choisy, J.P.; Henriquet, S.; Lecuyer, P.; Richard, M.; Tessier, C.; Sarrazin, F. Does sex matter in reintroduction of griffon vultures Gyps fulvus? Oryx 2007, 41, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Bessa-Gomes, C.; Legendre, S.; Clobert, J. Allee effects, mating systems and the extinction risk in populations with two sexes. Ecol. Lett. 2004, 7, 802–812. [Google Scholar] [CrossRef]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 4326, 95–716. [Google Scholar]
- Zhu, Q.; Zhang, J.; Bachtrog, D.; An, N.; Huang, Q.; Jarvis, E.D. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 6215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solari, A.J.; Dresser, M.E. High-resolution cytological localization of the XhoI and EcoRI repeat sequences in the pachytene ZW bivalent of the chicken. Chromosom. Res. 1995, 3, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Lessells, C.; Mateman, A. Sexing birds using random amplified polymorphic DNA (RAPD) markers. Mol. Ecol. 1998, 7, 187–195. [Google Scholar] [CrossRef]
- Smeds, L.; Warmuth, V.; Bolivar, P.; Uebbing, S.; Burri, R.; Suh, A.; Nater, A.; Bureš, S.; Garamszegi, L.Z.; Hogner, S.; et al. Evolutionary analysis of the female-specific avian W chromosome. Nat. Commun. 2015, 6, 7330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, R.; Daan, S.; Dijkstra, C. Sex identification in birds using two CHD genes. Proc. R. Soc. Lond. B. 1996, 263, 1251–1256. [Google Scholar]
- Fridolfsson, A.K.; Ellegren, H. A simple and universal method for molecular sexing of non-ratite birds. J. Avian Biol. 1999, 30, 116–121. [Google Scholar] [CrossRef]
- de Kloet, R.S.; de Kloet, S.R. Evolution of the spindlin gene in birds: Independent cessation of the recombination of sex chromosomes at the spindlin locus in neognathous birds and tinamous, a palaeognathous avian family. Genetica 2003, 119, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Suh, A.; Kriegs, J.O.; Brosius, J.; Schmitz, J. Retroposon insertions and the chronology of avian sex chromosome evolution. Mol. Biol. Evol. 2011, 28, 2993–2997. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xue, F.; Li, L.; Li, X.; Yue, B.; Li, J. A triple-primer PCR approach for the sex identification of endangered Phasianidae birds. Eur. J. Wildl. Res. 2012, 58, 289–294. [Google Scholar] [CrossRef]
- Griffiths, R.; Double, M.C.; Orr, K.; Dawson, R.J.G. A DNA test to sex most birds. Mol. Ecol. 1998, 7, 1071–1075. [Google Scholar] [CrossRef]
- Miyaki, C.Y.; Griffiths, R.; Orr, K.; Nahum, L.A.; Pereira, S.; Wajntal, A. Sex identification of parrots, toucans, and Curassows by PCR: Perspectives for wild and captive population studies. Zoo. Biol. 1998, 17, 415–423. [Google Scholar] [CrossRef]
- Cortés, O.; Barroso, A.; Dunner, S. Avian Sexing: An optimized protocol using Polymerase Chain Reaction-Single-Strand Conformation Polymorphism. J. Vet. Diagn. Investig. 1999, 11, 297–299. [Google Scholar] [CrossRef] [Green Version]
- Canon, N.R.; Tell, L.A.; Needham, M.L.; Gardner, I.A. Low cytometric analysis of nuclear DNA for sex identification in three psittacine species. Am. J. Vet. Res. 2000, 61, 847–850. [Google Scholar] [CrossRef]
- Bermúdez-Humarán, L.G.; García-García, A.; Leal-Garza, C.H.; Riojas-Valdes, V.M.; Jaramillo-Range, G.; Montes-de-oca-Luna, R. Molecular sexing of monomorphic endangered Ara birds. J. Exp. Zool. 2002, 292, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Boutette, J.B.; Ramsay, E.C.; Potgieter, L.N.D.; Kania, S.A. An improved Polymerase Chain Reaction–Restriction Fragment Length Polymorphism assay for gender identification in birds. J. Avian Med. Surg. 2002, 16, 198–202. [Google Scholar] [CrossRef]
- Jensen, T.; Pernasetti, F.M.; Durrant, B. Conditions for rapid sex determination in 47 avian species by PCR of genomic DNA from blood, shell-membrane blood vessels and feathers. Zoo. Biol. 2003, 22, 561–571. [Google Scholar] [CrossRef]
- Sundstrom, H.; Webster, M.T.; Ellegren, H. Is the Rate of Insertion and Deletion Mutation Male Biased?: Molecular Evolutionary Analysis of Avian and Primate Sex Chromosome Sequences. Genetics 2003, 164, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Chen, C.T.; Lee, H.Y.; Li, S.H.; Lir, J.T.; Chin, S.C. Sexing a wider range of avian species based on two CHD1 introns with a unified reaction condition. Zoo Biol. 2007, 26, 425–431. [Google Scholar] [CrossRef]
- Ong, A.H.K.; Vellayan, S. An evaluation of CHD-specific primer sets for sex typing of birds from feathers. Zoo. Biol. 2008, 27, 62–69. [Google Scholar] [CrossRef]
- Taylor, T.D.; Parkin, D.T. Sex ratios observed in 80 species of parrots. J. Zool. 2008, 276, 89–94. [Google Scholar] [CrossRef]
- Han, J.I.; Kim, J.H.; Kim, S.; Park, S.R.; Na, K.J. A simple and improved DNA test for avian sex determination. Auk 2009, 126, 779–783. [Google Scholar] [CrossRef]
- Sulandari, S.; Zein, M.S.A. Application of two molecular sexing methods for Indonesian bird species: Implication for captive breeding programs in Indonesia. HAYATI J. Biosci. 2012, 19, 183–190. [Google Scholar] [CrossRef] [Green Version]
- Bosnjak, J.; Stevanov-Pavlovic, M.; Vucicevic, M.; Stevanovic, J.; Simeunovic, P.; Resanovic, R.; Stanimirovic, Z. Feasibility of non-invasive molecular method for sexing of parrots. Pak. J. Zool. 2013, 45, 715–720. [Google Scholar]
- Braun, M.P.; Wink, M. Nestling development of ring-necked parakeets (Psittacula krameri) in a nest box population. Open Ornithol. J. 2013, 6, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Presti, F.T.; Meyer, J.; Antas, P.T.Z.; Guedes, N.M.R.; Miyaki, C.Y. Non-invasive genetic sampling for molecular sexing and microsatellite genotyping of hyacinth macaw (Anodorhynchus hyacinthinus). Genet. Mol. Biol. 2013, 36, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Dawson, D.A.; dos Remedios, N.; Horsburgh, G.J. A new marker based on the avian spindlin gene that is able to sex most birds, including species problematic to sex with CHD markers. Zoo Biol. 2016, 35, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Cakmak, E.; Akın Peksen, C.; Bilgin, C.C. Comparison of three different primer sets for sexing birds. J. Vet. Diagn. Investig. 2017, 29, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Clements, J.F.; Schulenberg, T.S.; Iliff, M.J.; Roberson, D.; Fredericks, T.A.; Sullivan, B.L.; Wood, C.L. The eBird/Clements Checklist of Birds of the World: v2018. Available online: http://www.birds.cornell.edu/clementschecklist (accessed on 25 September 2018).
- Joseph, L.; Toon, A.; Schirtzinger, E.E.; Wright, T.F.; Schodde, R. A revised nomenclature and classification for family-group taxa of parrots (Psittaciformes). Zootaxa 2012, 3205, 26–40. [Google Scholar] [CrossRef]
- Schodde, R.; Remsen, J.V.; Schirtzinger, E.E.; Joseph, L.; Wright, T.F. Higher classification of new world parrots (Psittaciformes; Arinae), with diagnoses of tribes. Zootaxa 2013, 3691, 591–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawson, D.A. Genomic Analysis of Passerine Birds Using Conserved Microsatellite Loci. Ph.D. Thesis, University of Sheffield, Sheffield, UK, 2007. [Google Scholar]
- Cerit, H.; Avanus, K. Sex determination by CHDW and CHDZ genes of avian sex chromosomes in Nymphicus hollandicus. Turk. J. Vet. Anim. Sci. 2007, 31, 371–374. [Google Scholar]
- Russello, M.A.; Amato, G. Application of a noninvasive, PCR-based test for sex identification in an endangered parrot, Amazona guildingii. Zoo Biol. 2001, 20, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Symes, C.T.; Perrin, M.R. Breeding biology of the Greyheaded parrot (Poicephalus fuscicollis suahelicus) in the wild. Emu 2004, 104, 45–57. [Google Scholar] [CrossRef]
- Ellegren, H. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc. Biol. Sci. 1996, 263, 1635–1641. [Google Scholar] [PubMed]
- Ogawa, A.; Solovei, I.; Hutchison, N.; Saitoh, Y.; Ikeda, J.-E.; MacGregor, H.; Mizuno, S. Molecular characterization and cytological mapping of a non-repetitive DNA sequence region from the W chromosome of chicken and its use as a universal probe for sexing carinatae birds. Chromosom. Res. 1997, 5, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Kahn, N.W.; John, J.S.T.; Quinn, T.W. Chromosome-specific intron size differences in the avian CHD gene provide an efficient method for sex identification in birds. Auk 1998, 115, 1074–1078. [Google Scholar]
- Griffiths, R.; Phil, D. Sex Identification in Birds. Semin. Avian Exot. Pet Med. 2000, 9, 14–26. [Google Scholar] [CrossRef]
- Huynen, L.; Miles, J.; Lambert, D. Unusual electrophoretic mobility of a DNA fragment of the universal ‘non-ratite’ sexing marker CHD allows sexing of New Zealand’s endangered kiwi ratite Apteryx spp. IBIS 2006, 148, 167–168. [Google Scholar]
- Vucicevic, M.; Stevanov-Pavlovic, M.; Stevanovic, J.; Bosnjak, J.; Gajic, B.; Aleksic, N.; Stanimirovic, Z. Sex determination in 58 bird species and evaluation of CHD gene as a universal molecular marker in bird sexing. Zoo Biol. 2013, 32, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Furo, I.D.O.; Kretschmer, R.; Dos Santos, M.S.; Carvalho, C.A.D.L.; Gunski, R.J.; O’Brien, P.C.; Ferguson-Smith, M.A.; Cioffi, M.B.; De Oliveira, E.H. Chromosomal mapping of repetitive DNAs in Myiopsitta monachus and Amazona aestiva (Psittaciformes, Psittacidae) with emphasis on the sex chromosomes. Cytogenet. Genome Res. 2017, 151, 151–160. [Google Scholar] [CrossRef]
- Dawson, D.A.; Darby, S.; Hunter, F.M.; Krupa, A.P.; Jones, I.L.; Burke, T. A critique of avian CHD-based molecular sexing protocols illustrated by a Z-chromosome polymorphism detected in auklets. Mol. Ecol. Notes 2001, 1, 201–204. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SPECIES | Sexing Strategy b | No | CHD1iE | CHD1iA | CHD1i9 | NIPBLi16 |
|---|---|---|---|---|---|---|
| Agapornis canusa | 2 | 68 | ― | 3e | 6b | 7c |
| Agapornis fischeria | 1 | 95 | ― | 3a | 6b | 7a |
| Agapornis lilianea | 2 | 102 | ― | 3a | 6b | 7c |
| Agapornis nigrigenisa | 2 | 62 | 4d | 3e | 6b | 7a |
| Agapornis personataa | 1 | 73 | ― | 3a | 6c | 7a |
| Agapornis roseicollis | 1 | 24 | 4d | 3a | 6c | 7a |
| Agapornis tarantaa | 1 | 67 | ― | 3a | 6b | 7a |
| Alipiopsitta xanthopsa | 1 | 119 | ― | 3a | 6g | 7a |
| Alisterus scapularisa | 2 | 39 | 4i | 3d | 6a | 7c |
| Amazona aestiva | 1 | 43 | 4a | 3a | 6g | 7b |
| Amazona albifrons | 1 | 78 | ― | 3a | 6g | 7e |
| Amazona agilisa | 1 | 120 | ― | 3a | 6a | 7e |
| Amazona amazonica | 1 | 44 | 4a | 3a | 6g | 7a |
| Amazona auropalliata | 1 | 99 | ― | 3a | 6g | 7a |
| Amazona autumnalis | 1 | 79 | ― | 3a | 6g | 7a |
| Amazona barbadensis | 1 | 1 | 4a | 3a | 6g | 7a |
| Amazona brasiliensis | 1 | 121 | ― | 3a | 6g | 7b |
| Amazona collariaa | 1 | 122 | ― | 3a | 6g | 7e |
| Amazona dufresnianaa | 1 | 123 | ― | 3a | 6g | 7a |
| Amazona farinosa | 1 | 94 | ― | 3a | 6c | 7a |
| Amazona festivaa | 1 | 124 | ― | 3a | 6g | 7a |
| Amazona finschi | 1 | 59 | 4a | 3a | 6g | 7b |
| Amazona guatemalaea | 1 | 125 | ― | 3a | 6c | 7a |
| Amazona guildingiia | 1 | 132 | ― | 3a | 6g | 7a |
| Amazona leucocephala | 1 | 88 | ― | 3a | 6g | 7b |
| Amazona ochrocephala | 1 | 100 | ― | 3a | 6g | 7a |
| Amazona oratrixa | 1 | 2 | 4a | 3a | 6g | 7a |
| Amazona pretrei | 1 | 76 | ― | 3a | 6g | 7a |
| Amazona rhodocorythaa | 1 | 126 | ― | 3a | 6a | 7a |
| Amazona tucumana | 1 | 127 | ― | 3a | 6g | 7a |
| Amazona versicolora | 1 | 131 | ― | 3a | 6g | 7a |
| Amazona vinacea | 1 | 77 | ― | 3a | 6g | 7a |
| Amazona viridigenalis | 1 | 87 | ― | 3a | 6g | 7b |
| Amazona xantholoraa | 1 | 128 | ― | 3a | 6a | 7e |
| Anodorhynchus hyacinthinus | 2 | 53 | 4f | 3h | 6b | 7a |
| Aprosmictus erythropterus | 1 | 40 | 4h | 3d | 6c | 7a |
| Ara ararauna | 1 | 3 | 4a | 3a | 6b | 7a |
| Ara chloroptera | 1 | 52 | 4a | 3a | 6c | 7b |
| Ara glaucogularis | 1 | 54 | 4a | 3a | 6a | 7a |
| Ara macao | 1 | 42 | 4a | 3a | 6a | 7a |
| Ara militaris | 1 | 98 | ― | 3a | 6c | 7a |
| Ara rubrogenys | 1 | 4 | 4a | 3a | 6b | 7a |
| Ara severa | 1 | 109 | ― | 3a | 6a | 7a |
| Aratinga auricapillusa | 1 | 108 | ― | 3a | 6b | 7a |
| Aratinga jandaya | 1 | 97 | ― | 3a | 6b | 7a |
| Aratinga nenday | 1 | 49 | 4f | 3a | 6b | 7a |
| Aratinga solstitialis | 1 | 26 | 4a | 3a | 6b | 7a |
| Aratinga weddelliia | 1 | 117 | ― | 3a | 6c | 7b |
| Barnardius zonarius | 2 | 38 | 4c | 3e | 6b | 7a |
| Bolborhynchus lineola | 1 | 7 | 4h | 3a | 6e | 7a |
| Brotogeris jugularisa | 1 | 112 | ― | 3a | 6c | 7a |
| Cacatua ducorpsii | 1 | 29 | 4a | 3a | 6a | 7b |
| Cacatua goffini | 1 | 51 | 4a | 3a | 6c | 7b |
| Cacatua moluccensis | 1 | 30 | 4a | 3a | 6b | 7b |
| Cacatua sulphurea | 1 | 8 | 4b | 3a | 6b | 7a |
| Calyptorhynchus banksiia | 1 | 130 | ― | 3a | 6c | 7a |
| Chalcopsitta atra | 2 | 92 | ― | 3a | 6e | 7d |
| Chalcopsitta duivenbodei | 2 | 50 | 4h | 3a | 6e | 7c |
| Coracopsis vasa | 2 | 57 | 4a | 3i | 6b | 7g |
| Cyanoliseus patagonus | 1 | 9 | 4b | 3a | 6b | 7a |
| Cyanopsitta spixiia | 1 | 135 | ― | 3a | 6b | 7a |
| Cyanoramphus novaezelandiaea | 1 | 31 | 4a | 3a | 6a | 7a |
| Deroptyus accipitrinus | 1 | 80 | ― | 3a | 6b | 7a |
| Diopsittaca nobilisa | 1 | 28 | 4a | 3a | 6b | 7a |
| Eclectus roratus | 2 | 58 | 4a | 3f | 6b | 7b |
| Enicognathus ferrugineusa | 1 | 134 | ― | 3a | 6c | 7b |
| Enicognathus leptorhynchusa | 1 | 133 | ― | 3a | 6c | 7b |
| Eolophus roseicapillus | 1 | 32 | 4c | 3a | 6b | 7a |
| Eupsittula aurea | 1 | 6 | 4a | 3a | 6b | 7a |
| Forpus coelestisa | 1 | 106 | ― | 3c | 6a | 7a |
| Forpus crassirostrisa | 2 | 10 | 4i | 3g | 6a | 7b |
| Forpus xanthopsa | 1 | 96 | ― | 3c | 6a | 7a |
| Guaruba guarouba | 1 | 11 | 4c | 3a | 6b | 7a |
| Lathamus discolora | 1 | 104 | ― | 3a | 6b | 7a |
| Lophochroa leadbeateria | 1 | 55 | 4f | 3a | 6b | 7b |
| Loriculus galgulus | 1 | 60 | 4f | 3a | 6a | 7a |
| Lorius chlorocercusa | 2 | 89 | ― | 3a | 6e | 7c |
| Lorius garrulus | 1 | 90 | ― | 3a | 6e | 7b |
| Lorius lory | 1 | 91 | ― | 3a | 6e | 7b |
| Melopsitacus undulatus | 1 | 25 | 4a | 3a | 6a | 7b |
| Myiopsitta monachus | 2 | 47 | 4a | 3b | 6e | 7h |
| Neophema elegansa | 2 | 103 | ― | 3h | 6b | 7a |
| Neophema pulchellaa | 2 | 13 | 4a | 3h | 6b | 7b |
| Neophema splendidaa | 2 | 72 | ― | 3h | 6b | 7a |
| Neopsephotus bourkii | 1 | 12 | 4a | 3a | 6b | 7a |
| Nestor notabilis | 2 | 56 | 4j | 3i | 6b | 7a |
| Nymphicus hollandicus | 1 | 14 | 4a | 3a | 6b | 7b |
| Orthopsittaca manilataa | 3 | 113 | ― | 3i | 6h | 7b |
| Pionites melanocephalus | 1 | 15 | 4c | 3a | 6b | 7a |
| Pionopsitta pileata | 1 | 101 | ― | 3a | 6b | 7b |
| Pionus chalcopterus | 1 | 69 | ― | 3a | 6g | 7a |
| Pionus maximiliani | 1 | 86 | ― | 3a | 6g | 7b |
| Pionus menstruus | 1 | 16 | 4a | 3a | 6g | 7a |
| Pionus senilis | 1 | 46 | 4a | 3a | 6g | 7a |
| Platycercus adscitusa | 1 | 74 | ― | 3a | 6a | 7a |
| Platycercus caledoniusa | 2 | 110 | ― | 3h | 6a | 7b |
| Platycercus elegans | 2 | 17 | 4a | 3e | 6c | 7a |
| Platycercus eximiusa | 1 | 37 | 4a | 3a | 6b | 7a |
| Platycercus icterotisa | 2 | 64 | 4a | 3h | 6a | 7a |
| Poicephalus fuscicollisa | 2 | 116 | ― | 3a | 6b | 7h |
| Poicephalus gulielmi | 3 | 18 | 4a | 3a | 6g | 7h |
| Poicephalus meyeri | 2 | 118 | ― | 3a | 6e | 7h |
| Poicephalus senegalus | 2 | 33 | 4a | 3a | 6b | 7h |
| Polytelis alexandraea | 2 | 19 | 4h | 3d | 6d | 7c |
| Polytelis anthopeplusa | 1 | 48 | 4h | 3d | 6b | 7a |
| Polytelis swainsonii | 1 | 20 | 4h | 3d | 6b | 7b |
| Primolius auricollisa | 1 | 82 | ― | 3a | 6c | 7a |
| Primolius coulonia | 1 | 61 | 4c | 3a | 6b | 7a |
| Primolius maracanaa | 1 | 41 | 4a | 3a | 6b | 7a |
| Probosciger aterrimus | 1 | 34 | 4a | 3a | 6f | 7a |
| Psephotus haematonotusa | 1 | 21 | 4d | 3a | 6b | 7a |
| Psilopsiagon aymaraa | 2 | 71 | ― | 3a | 6b | 7h |
| Psittacara brevipesa | 1 | 115 | ― | 3a | 6b | 7a |
| Psittacara finschia | 1 | 111 | ― | 3a | 6b | 7b |
| Psittacara mitrataa | 1 | 85 | ― | 3a | 6b | 7b |
| Psittacara wagleria | 1 | 107 | ― | 3a | 6a | 7a |
| Psittacula alexandrii | 2 | 105 | ― | 3f | 6b | 7c |
| Psittacula cyanocephala | 2 | 70 | ― | 3g | 6c | 7f |
| Psittacula derbianaa | 2 | 35 | 4e | 3g | 6c | 7b |
| Psittacula eupatria | 2 | 63 | 4g | 3f | 6b | 7a |
| Psittacula krameri | 2 | 45 | 4a | 3f | 6b | 7a |
| Psittacus erithacus | 2 | 22 | 4a | 3a | 6b | 7h |
| Psittinus cyanurusa | 2 | 129 | ― | 3g | 6c | 7e |
| Purpureicephalus spuriusa | 1 | 81 | ― | 3a | 6b | 7a |
| Pyrhura roseifronsa | 1 | 83 | ― | 3a | 6a | 7a |
| Pyrrhura frontalis | 1 | 27 | 4a | 3a | 6b | 7a |
| Pyrrhura molinae | 1 | 23 | 4a | 3a | 6b | 7a |
| Pyrrhura perlata | 1 | 75 | ― | 3a | 6a | 7a |
| Pyrrhura picta | 1 | 65 | ― | 3a | 6b | 7a |
| Pyrrhura rhodocephala | 2 | 84 | ― | 3a | 6b | 7c |
| Pyrrhura rupicolaa | 1 | 66 | ― | 3a | 6b | 7a |
| Rhynchopsitta terrisia | 2 | 114 | ― | 3e | 6b | 7a |
| Thectocercus acuticaudatus | 1 | 5 | 4a | 3a | 6b | 7b |
| Trichoglossus euteles | 2 | 93 | ― | 3a | 6e | 7c |
| Trichoglossus haematodus | 2 | 36 | 4h | 3a | 6f | 7c |
| Superfamily | Family | Subfamily | Tribe | Genus |
|---|---|---|---|---|
| Strigopoidea | Strigopidae | Strigops | ||
| Nestoridae | Nestor | |||
| Cacatuoidea | Cacatuidae | Nymphicinae | Nymphicus | |
| Calyptorhynchinae | Calyptorhynchus | |||
| Cacatuinae | Microglossini | Probosciger | ||
| Cacatuini | Cacatua, Callocephalon, Eolophus, Lophochroa | |||
| Psittacoidea | Psittacidae | Psittacinae | Poicephalus, Psittacus | |
| Arinae | Arini | Anodorhynchus, Ara, Aratinga, Cyanoliseus, Cyanopsitta, Deroptyus, | ||
| Diopsittaca, Enicognathus, Eupsittula, Guaruba, Leptosittaca, Ognorhynchus, | ||||
| Orthopsittaca, Pionites, Primolius, Psittacara, Pyrrhura, Rhynchopsitta, Thectocercus | ||||
| Androglossini | Alipiopsitta, Amazona, Brotogeris, Graydidascalus, Hapalopsittaca, | |||
| Myiopsitta, Pionopsitta, Pionus, Pyrilia, Triclaria | ||||
| Amoropsittacini | Bolborhynchus, Nannopsittaca, Psilopsiagon, Touit | |||
| Forpini | Forpus | |||
| Psittrichasidae | Psittrichasinae | Psittrichas | ||
| Coracopseinae | Coracopsis | |||
| Psittaculidae | Platycercinae | Platycercini | Barnardius, Cyanoramphus, Eunymphicus, Lathamus, Northiella, | |
| Platycercus, Prosopeia, Psephotus, Purpureicephalus | ||||
| Pezoporini | Neophema, Neopsephotus, Pezoporus | |||
| Psittacellinae | Psittacella | |||
| Loriinae | Loriini | Chalcopsitta, Charmosyna, Eos, Glossopsitta, Lorius, Neopsittacus, | ||
| Oreopsittacus, Phigys, Pseudeos, Psitteuteles, Trichoglossus, Vini | ||||
| Melopsittacini | Melopsittacus | |||
| Cyclopsittini | Cyclopsitta, Psittaculirostris | |||
| Agapornithinae | Agapornis, Bolbopsittacus, Loriculus | |||
| Psittaculinae | Polytelini | Alisterus, Aprosmictus, Polytelis | ||
| Psittaculini | Eclectus, Geoffroyus, Prioniturus, Psittacula, Psittinus, Tanygnathus | |||
| Micropsittini | Micropsitta |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kroczak, A.; Wołoszyńska, M.; Wierzbicki, H.; Kurkowski, M.; Grabowski, K.A.; Piasecki, T.; Galosi, L.; Urantówka, A.D. New Bird Sexing Strategy Developed in the Order Psittaciformes Involves Multiple Markers to Avoid Sex Misidentification: Debunked Myth of the Universal DNA Marker. Genes 2021, 12, 878. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060878
Kroczak A, Wołoszyńska M, Wierzbicki H, Kurkowski M, Grabowski KA, Piasecki T, Galosi L, Urantówka AD. New Bird Sexing Strategy Developed in the Order Psittaciformes Involves Multiple Markers to Avoid Sex Misidentification: Debunked Myth of the Universal DNA Marker. Genes. 2021; 12(6):878. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060878
Chicago/Turabian StyleKroczak, Aleksandra, Magdalena Wołoszyńska, Heliodor Wierzbicki, Marcin Kurkowski, Krzysztof Aleksander Grabowski, Tomasz Piasecki, Livio Galosi, and Adam Dawid Urantówka. 2021. "New Bird Sexing Strategy Developed in the Order Psittaciformes Involves Multiple Markers to Avoid Sex Misidentification: Debunked Myth of the Universal DNA Marker" Genes 12, no. 6: 878. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12060878