
Embryo Screening for Polygenic Disease Risk: Recent Advances and Ethical Considerations

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
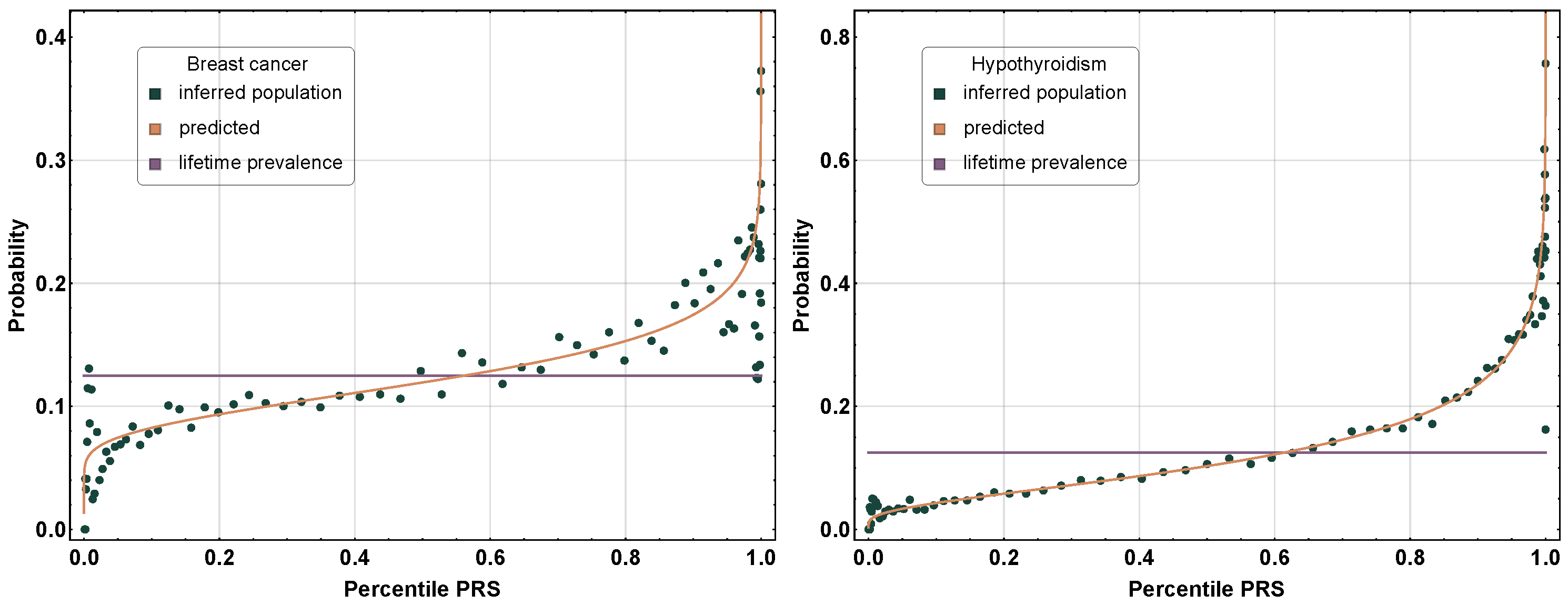
2. Polygenic Risk Scores (PRSs)
3. Precision Embryo Genotyping
4. Ethical Considerations
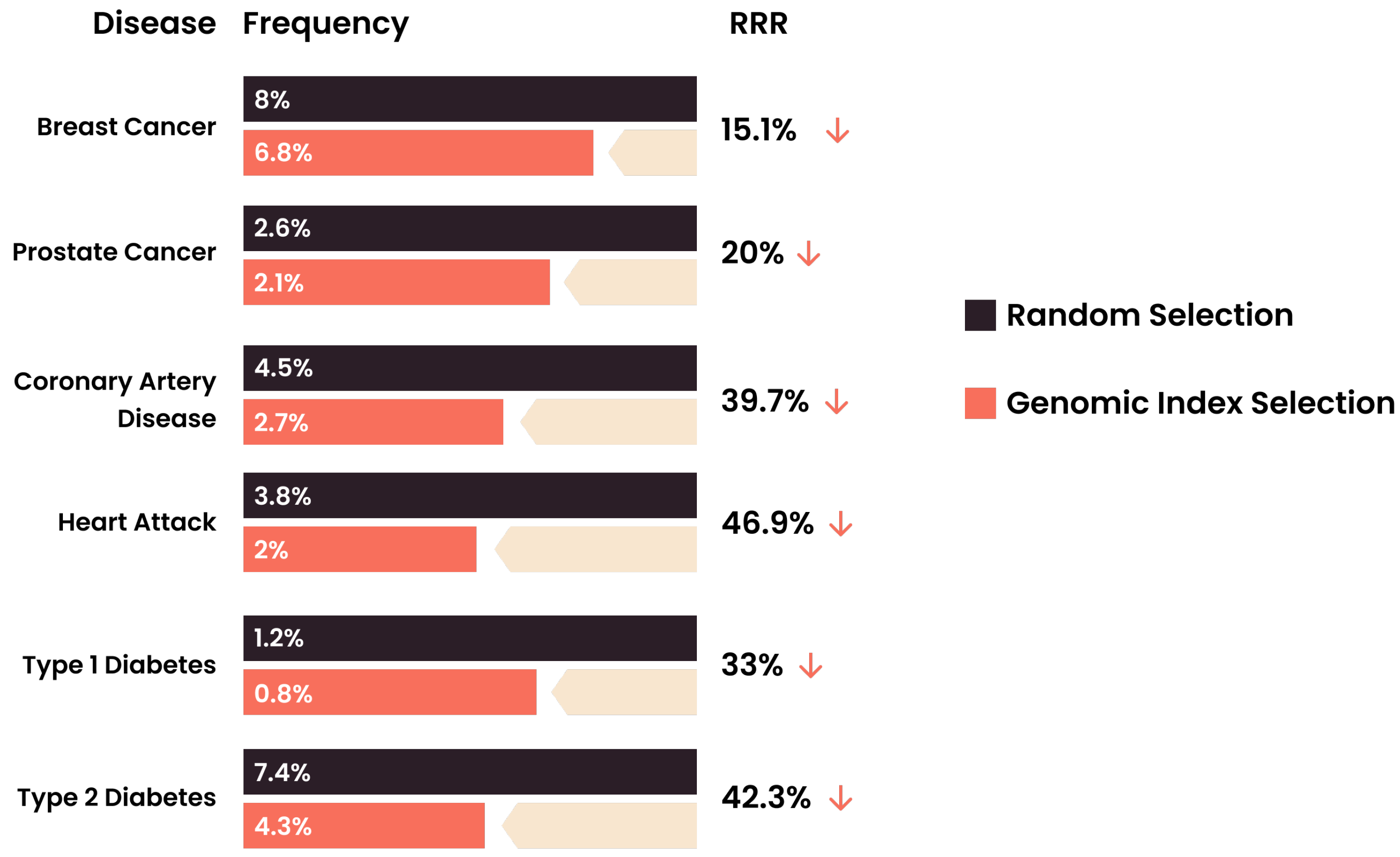
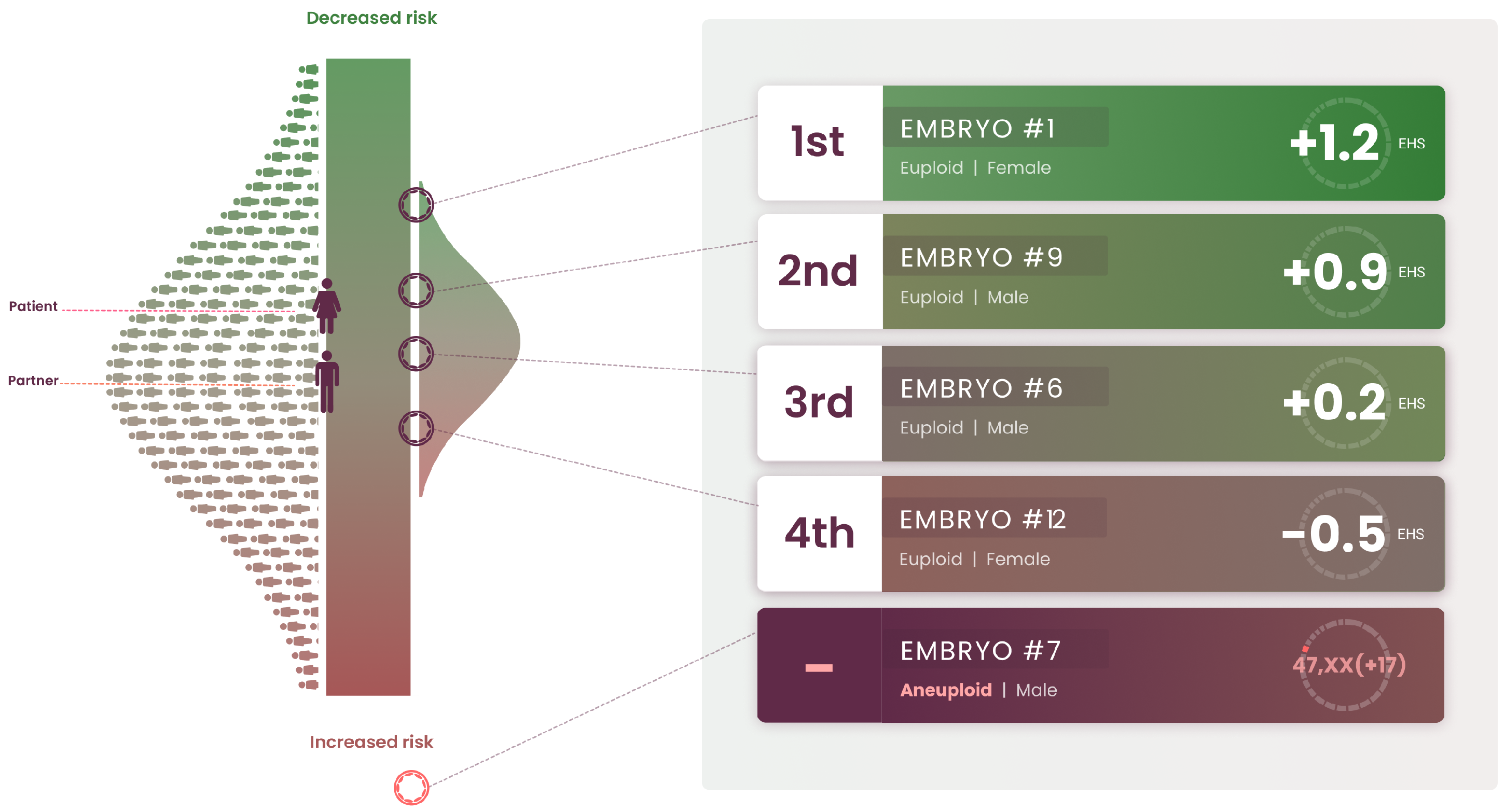
5. Clinical Application of the Embryo Health Score
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fauser, B.C. Towards the global coverage of a unified registry of IVF outcomes. Reprod. Biomed. Online 2019, 38, P133–P137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Society of Human Reproduction and Embryology. ART Fact Sheet. Available online: https://www.eshre.eu/-/media/sitecore-files/Press-room/ART-fact-sheet-2020-data-2016.pdf?la=en&hash=AB68A67B4FEA7723F2125B02BCB93FB837139CD4 (accessed on 7 July 2021).
- Raben, T.G.; Lello, L.; Widen, E.; Hsu, S.D.H. From Genotype to Phenotype: Polygenic Prediction of Complex Human Traits. 2021. Available online: http://xxx.lanl.gov/abs/2101.05870 (accessed on 7 July 2021).
- Munday, S.; Savulescu, J. Three models for the regulation of polygenic scores in reproduction. J. Med. Ethics 2021. [Google Scholar] [CrossRef]
- Lello, L.; Raben, T.G.; Yong, S.Y.; Tellier, L.C.; Hsu, S.D.H. Genomic prediction of 16 complex disease risks including heart attack, diabetes, breast and prostate cancer. Sci. Rep. 2019, 9, 15286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khera, A.V.; Chaffin, M.; Aragam, K.G.; Haas, M.E.; Roselli, C.; Choi, S.H.; Natarajan, P.; Lander, E.S.; Lubitz, S.A.; Ellinor, P.T.; et al. Genome-wide polygenic scores for common diseases identify individuals with risk equivalent to monogenic mutations. Nat. Genet. 2018, 50, 1219. [Google Scholar] [CrossRef]
- Khera, A.V.; Chaffin, M.; Wade, K.H.; Zahid, S.; Brancale, J.; Xia, R.; Distefano, M.; Senol-Cosar, O.; Haas, M.E.; Bick, A.; et al. Polygenic prediction of weight and obesity trajectories from birth to adulthood. Cell 2019, 177, 587–596. [Google Scholar] [CrossRef] [Green Version]
- Lewis, C.M.; Vassos, E. Prospects for using risk scores in polygenic medicine. Genome Med. 2017, 9, 96. [Google Scholar] [CrossRef] [Green Version]
- Lambert, S.A.; Gil, L.; Jupp, S.; Ritchie, S.C.; Xu, Y.; Buniello, A.; Abraham, G.; Chapman, M.; Parkinson, H.; Danesh, J.; et al. The Polygenic Score Catalog: An open database for reproducibility and systematic evaluation. medRxiv 2020. [Google Scholar] [CrossRef]
- Wünnemann, F.; Sin Lo, K.; Langford-Avelar, A.; Busseuil, D.; Dubé, M.P.; Tardif, J.C.; Lettre, G. Validation of Genome-Wide Polygenic Risk Scores for Coronary Artery Disease in French Canadians. Circ. Genom. Precis. Med. 2019, 12, e002481. [Google Scholar] [CrossRef] [Green Version]
- Privé, F.; Aschard, H.; Carmi, S.; Folkersen, L.; Hoggart, C.; O’Reilly, P.F.; Vilhjálmsson, B.J. High-resolution portability of 245 polygenic scores when derived and applied in the same cohort. medRxiv 2021. [Google Scholar] [CrossRef]
- Torkamani, A.; Wineinger, N.E.; Topol, E.J. The personal and clinical utility of polygenic risk scores. Nat. Rev. Genet. 2018, 19, 581. [Google Scholar] [CrossRef]
- Belsky, D.W.; Domingue, B.W.; Wedow, R.; Arseneault, L.; Boardman, J.D.; Caspi, A.; Conley, D.; Fletcher, J.M.; Freese, J.; Herd, P.; et al. Genetic analysis of social-class mobility in five longitudinal studies. Proc. Natl. Acad. Sci. USA 2018, 115, E7275–E7284. [Google Scholar] [CrossRef] [Green Version]
- Mars, N.; Koskela, J.T.; Ripatti, P.; Kiiskinen, T.T.J.; Havulinna, A.S.; Lindbohm, J.V.; Ahola-Olli, A.; Kurki, M.; Karjalainen, J.; Palta, P.; et al. Polygenic and clinical risk scores and their impact on age at onset and prediction of cardiometabolic diseases and common cancers. Nat. Med. 2020, 26, 549–557. [Google Scholar] [CrossRef]
- Durvasula, A.; Lohmueller, K.E. Negative selection on complex traits limits phenotype prediction accuracy between populations. Am. J. Hum. Genet. 2021, 108, 620–631. [Google Scholar] [CrossRef]
- Lello, L.; Raben, T.G.; Hsu, S.D.H. Sibling validation of polygenic risk scores and complex trait prediction. Sci. Rep. 2020, 10, 13190. [Google Scholar] [CrossRef]
- Lello, L.; Avery, S.G.; Tellier, L.; Vazquez, A.I.; de los Campos, G.; Hsu, S.D. Accurate genomic prediction of human height. Genetics 2018, 210, 477–497, [PMC621659]. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Kiryluk, K. Genome-wide polygenic risk predictors for kidney disease. Nat. Rev. Nephrol. 2018, 14, 723–724. [Google Scholar] [CrossRef]
- Chatterjee, N.; Shi, J.; García-Closas, M. Developing and evaluating polygenic risk prediction models for stratified disease prevention. Nat. Rev. Genet. 2016, 17, 392. [Google Scholar] [CrossRef]
- Euesden, J.; Lewis, C.M.; O’reilly, P.F. PRSice: Polygenic risk score software. Bioinformatics 2014, 31, 1466–1468. [Google Scholar] [CrossRef] [Green Version]
- Shieh, Y.; Shieh, Y.; Hu, D.; Ma, L.; Huntsman, S.; Gard, C.C.; Leung, J.W.T.; Tice, J.A.; Vachon, C.M.; Cummings, S.R.; et al. Breast cancer risk prediction using a clinical risk model and polygenic risk score. Breast Cancer Res. Treat. 2016, 159, 513–525. [Google Scholar] [CrossRef]
- Abraham, G.; Inouye, M. Genomic risk prediction of complex human disease and its clinical application. Curr. Opin. Genet. Dev. 2015, 33, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Priest, J.R.; Ashley, E.A. Genomics in clinical practice. BMJ Heart 2014, 100, 1569–1570. [Google Scholar] [CrossRef]
- Jacob, H.J.; Abrams, K.; Bick, D.P.; Brodie, K.; Dimmock, D.P.; Farrell, M.; Geurts, J.; Harris, J.; Helbling, D.; Joers, B.J.; et al. Genomics in clinical practice: Lessons from the front lines. Sci. Transl. Med. 2013, 5, 194cm5. [Google Scholar] [CrossRef] [Green Version]
- Veenstra, D.L.; Roth, J.A.; Garrison, L.P.; Ramsey, S.D.; Burke, W. A formal risk-benefit framework for genomic tests: Facilitating the appropriate translation of genomics into clinical practice. Genet. Med. 2010, 12, 686–693. [Google Scholar] [CrossRef] [Green Version]
- Bowdin, S.; Gilbert, A.; Bedoukian, E.; Carew, C.; Adam, M.P.; Belmont, J.; Bernhardt, B.; Biesecker, L.; Bjornsson, H.T.; Blitzer, M.; et al. Recommendations for the integration of genomics into clinical practice. Genet. Med. 2016, 18, 1075–1084. [Google Scholar] [CrossRef] [Green Version]
- Nelson, H.D.; Pappas, M.; Cantor, A.; Haney, E.; Holmes, R. Risk assessment, genetic counseling, and genetic testing for BRCA-related cancer in women: Updated evidence report and systematic review for the US Preventive Services Task Force. JAMA 2019, 322, 666–685. [Google Scholar] [CrossRef] [Green Version]
- Amir, E.; Freedman, O.C.; Seruga, B.; Evans, D.G. Assessing women at high risk of breast cancer: A review of risk assessment models. J. Natl. Cancer Inst. 2010, 102, 680–691. [Google Scholar] [CrossRef] [Green Version]
- Offit, K. BRCA Mutation Frequency and Penetrance: New Data, Old Debate. J. Natl. Cancer Inst. 2006, 98, 23. [Google Scholar] [CrossRef]
- Ford, D.; Easton, D.F.; Peto, J. Estimates of the gene frequency of BRCA1 and its contribution to breast and ovarian cancer incidence. Am. J. Hum. Genet. 1995, 57, 1457–1462. [Google Scholar]
- Whittemore, A.S.; Gong, G.; John, E.M.; McGuire, V.; Li, F.P.; Ostrow, K.L.; Dicioccio, R.; Felberg, A.; West, D.W. Prevalence of BRCA1 mutation carriers among U.S. non-Hispanic Whites. Cancer Epidemoiol. Biomark. Prev. 2004, 13, 2078–2083. [Google Scholar]
- Kuchenbaecker, K.; McGuffog, L.; Barrowdale, D.; Lee, A.; Soucy, P.; Dennis, J.; Domchek, S.M.; Robson, M.; Spurdle, A.B.; Ramus, S.J.; et al. Evaluation of Polygenic Risk Scores for Breast and Ovarian Cancer Risk Prediction in BRCA1 and BRCA2 Mutation Carriers. J. Natl. Cancer Inst. 2017, 109, 7. [Google Scholar] [CrossRef] [Green Version]
- Hughes, E.; Tshiaba, P.; Gallagher, S.; Wagner, S.; Judkins, T.; Roa, B.; Rosenthal, E.; Domchek, S.; Garber, J.; Lancaster, J.; et al. Development and Validation of a Clinical Polygenic Risk Score to Predict Breast Cancer Risk. JCO Precis. Oncol. 2020, 4, 585–592. [Google Scholar] [CrossRef]
- Myriad—Home. 2020. Available online: https://www.myriadmyrisk.com (accessed on 10 November 2020).
- Treff, N.R.; Zimmerman, R.; Bechor, E.; Hsu, J.; Rana, B.; Jensen, J.; Li, J.; Samoilenko, A.; Mowrey, W.; Alstine, J.V.; et al. Validation of concurrent preimplantation genetic testing for polygenic and monogenic disorders, structural rearrangements and whole and segmental chromosome aneuploidy with a single universal platform. Eur. J. Med. Genet. 2019, 62, 103647. [Google Scholar] [CrossRef]
- Kumar, A.; Ryan, A.; Kitzman, J.O.; Wemmer, N.; Snyder, M.W.; Sigurjonsson, S.; Lee, C.; Banjevic, M.; Zarutskie, P.W.; Lewis, A.P.; et al. Whole genome prediction for preimplantation genetic diagnosis. Genome Med. 2015, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lencz, T.; Backenroth, D.; Granot-Hershkovitz, E.; Green, A.; Gettler, K.; Cho, J.H.; Weissbrod, O.; Zuk, O.; Carmi, S. Utility of polygenic embryo screening for disease depends on the selection strategy. bioRxiv 2021. [Google Scholar] [CrossRef]
- Turley, P.; Meyer, M.; Wang, N.; Cesarini, D.; Hammonds, E.; Martin, A.; Neale, B.; Rehm, H.; Wilkins-Haug, L.; Benjamin, D.; et al. Problems with Using Polygenic Scores to Select Embryos. N. Engl. J. Med. 2021, 385, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Ethics Committee of the American Society for Reproductive Medicine. Use of preimplantation genetic testing for monogenic defects (PGT-M) for adult-onset conditions: An Ethics Committee opinion. Fertil. Steril. 2018, 109, 989–992. [Google Scholar] [CrossRef]
- Singer, P. Parental choice and human development. In Human Enhancement; Savulescu, J., Bostrom, N., Eds.; Oxford University Press: Oxford, UK, 2009; Chapter 12; pp. 277–289. [Google Scholar]
- Physician Exercise of Conscience—Code of Medical Ethics Opinion 1.1.7. Available online: https://journalofethics.ama-assn.org/article/ama-code-medical-ethics-opinions-related-moral-distress/2017-06 (accessed on 7 July 2021).
- Timmers, P.R.; Mounier, N.; Läll, K.; Fischer, K.; Ning, Z.; Feng, X.; Bretherick, A.; Clark, D.W.; Consortium, E.; Shen, X.; et al. Genomic underpinnings of lifespan allow prediction and reveal basis in modern risks. bioRxiv. 2018. Available online: https://www.biorxiv.org/content/early/2018/07/06/363036.full.pdf (accessed on 7 July 2021).
- Meisner, A.; Kundu, P.; Zhang, Y.D.; Lan, L.V.; Kim, S.; Ghandwani, D.; Pal Choudhury, P.; Berndt, S.I.; Freedman, N.D.; Garcia-Closas, M.; et al. Combined Utility of 25 Disease and Risk Factor Polygenic Risk Scores for Stratifying Risk of All-Cause Mortality. Am. J. Hum. Genet. 2020, 107, 418–431. [Google Scholar] [CrossRef]
- Treff, N.R.; Eccles, J.; Marin, D.; Messick, E.; Lello, L.; Gerber, J.; Xu, J.; Tellier, L.C. Preimplantation genetic testing for polygenic disease relative risk reduction: Evaluation of genomic index performance in 11,883 adult sibling pairs. Genes 2020, 11, 648. [Google Scholar] [CrossRef]
- Yong, S.Y.; Raben, T.G.; Lello, L.; Hsu, S.D. Genetic Architecture of Complex Traits and Disease Risk Predictors. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tellier, L.C.A.M.; Eccles, J.; Treff, N.R.; Lello, L.; Fishel, S.; Hsu, S. Embryo Screening for Polygenic Disease Risk: Recent Advances and Ethical Considerations. Genes 2021, 12, 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081105
Tellier LCAM, Eccles J, Treff NR, Lello L, Fishel S, Hsu S. Embryo Screening for Polygenic Disease Risk: Recent Advances and Ethical Considerations. Genes. 2021; 12(8):1105. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081105
Chicago/Turabian StyleTellier, Laurent C. A. M., Jennifer Eccles, Nathan R. Treff, Louis Lello, Simon Fishel, and Stephen Hsu. 2021. "Embryo Screening for Polygenic Disease Risk: Recent Advances and Ethical Considerations" Genes 12, no. 8: 1105. https://0-doi-org.brum.beds.ac.uk/10.3390/genes12081105