
Genome-Wide Identification of NRT Gene Family and Expression Analysis of Nitrate Transporters in Response to Salt Stress in Poncirus trifoliata


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatment
2.2. Identification of NRTs in P. trifoliata and Amino Acid Characteristic Prediction
2.3. Phylogenetic Tree Construction and Synteny Correlation Analysis
2.4. Gene Structure and cis-Acting Elements Analysis
2.5. RNA Extract, cDNA Reverse Transcription, and Gene Expression Analysis via qRT-PCR
2.6. RNA-seq Data Analysis
2.7. Sequence Annotation, GO, and KEGG Enrichment Analysis
3. Result
3.1. Genome-Wide Identification of NRT Genes in P. trifoliata
3.2. Phylogenetic Relationship and Synteny Correlation Analysis
3.3. Motifs, Gene Structure, and cis-Acting Elements Analysis
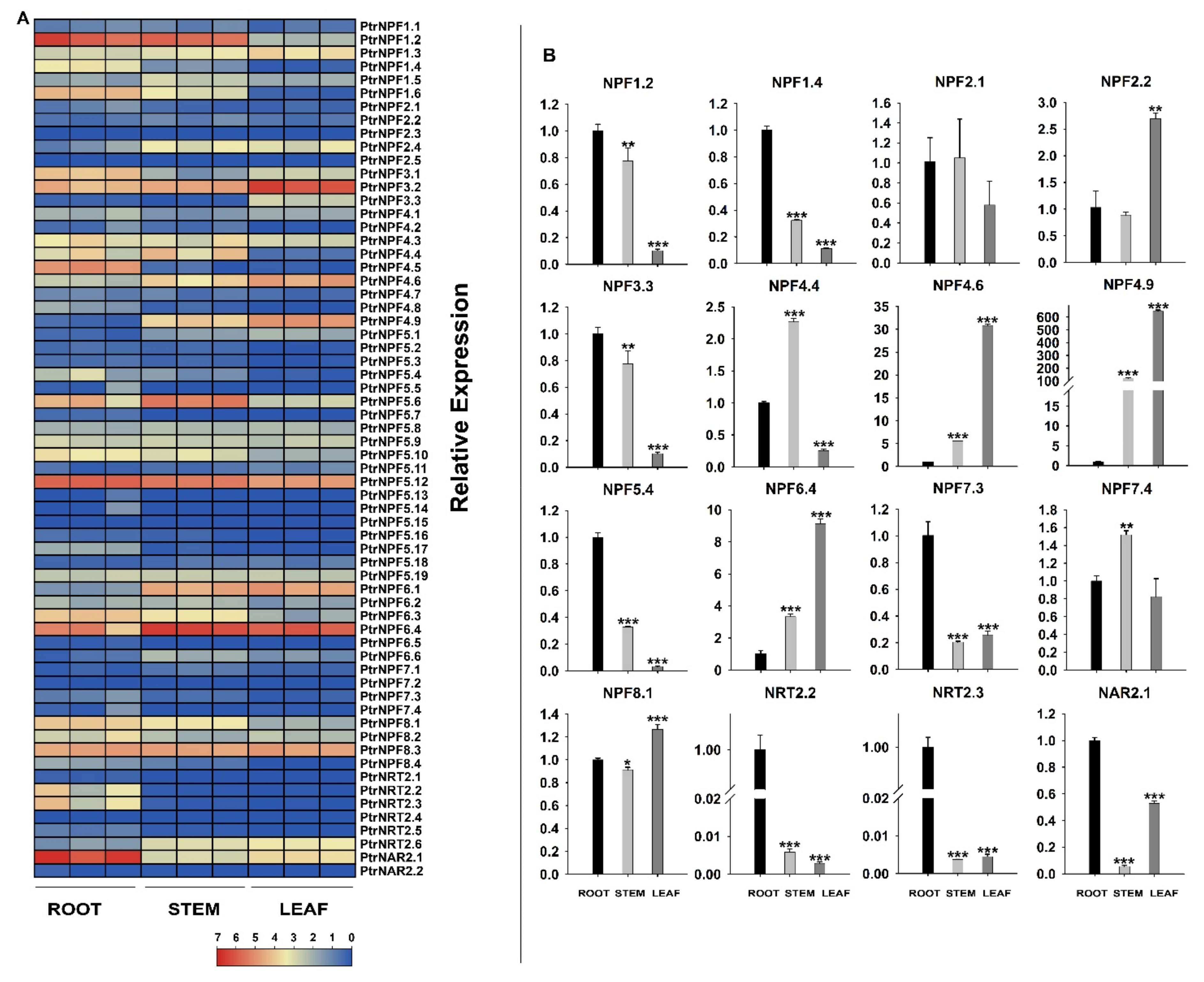
3.4. Expression Analysis of the PtrNRT Genes in Different Tissues
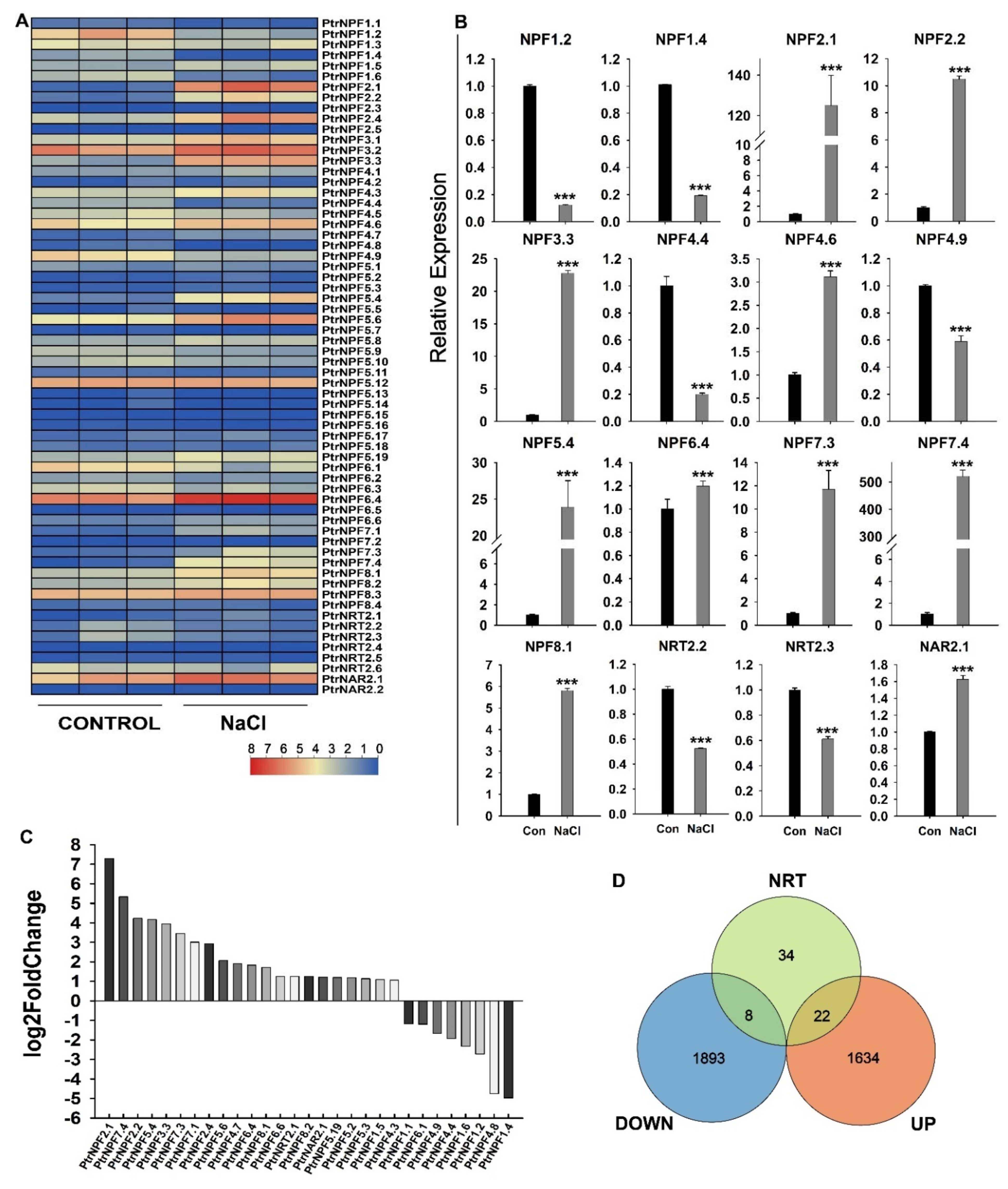
3.5. GO and KEGG Pathway Enrichment Analysis under Salt Treatment in P. trifoliata
3.6. Expression Analysis of the PtrNRT Genes in Response to Salt Stress
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y.Y.; Hsu, P.K.; Tsay, Y.F. Uptake, allocation and signaling of nitrate. Trends Plant Sci. 2012, 17, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, M.A.; Wang, L.M.; Jiao, Y.Y.; Chen, C.; Zhao, L.; Mei, M.J.; Yu, Y.L.; Bie, Z.L.; Huang, Y. Pumpkin rootstock improves nitrogen use efficiency of watermelon scion by enhancing nutrient uptake, cytokinin content, and expression of nitrate reductase genes. Plant Growth Regul. 2017, 82, 233–246. [Google Scholar] [CrossRef]
- Luo, J.; Li, H.; Liu, T.X.; Polle, A.; Peng, C.H.; Luo, Z.B. Nitrogen metabolism of two contrasting poplar species during acclimation to limiting nitrogen availability. J. Exp. Bot. 2013, 64, 4207–4224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konishi, M.; Yanagisawa, S. Emergence of a new step towards understanding the molecular mechanisms underlying nitrate-regulated gene expression. J. Exp. Bot. 2014, 65, 5589–5600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, F. Nitrate Assimilation: The role of in vitro nitrate reductase assay as nutritional predictor. In Applied Photosynthesis; Najapour, M., Ed.; InTech: Rijeka, Croatia, 2012; pp. 105–120. [Google Scholar]
- Crawford, N.M.; Glass, A.D.M. Molecular and physiological aspects of nitrate uptake in plants. Trends Plant Sci. 1998, 3, 389–395. [Google Scholar] [CrossRef]
- You, H.; Liu, Y.; Minh, T.N.; Lu, H.; Zhang, P.; Li, W.; Xiao, J.; Ding, X.; Li, Q. Genome-wide identification and expression analyses of nitrate transporter family genes in wild soybean (Glycine soja). J. Appl. Genet. 2020, 61, 489–501. [Google Scholar] [CrossRef]
- Zhang, J.; Han, Z.; Lu, Y.; Zhao, Y.; Wang, Y.; Zhang, J.; Ma, H.; Han, Y.Z. Genome-wide identification, structural and gene expression analysis of the nitrate transporters (NRTs) family in potato (Solanum tuberosum L.). PLoS ONE 2021, 16, e0257383. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutierrez, R.A. Nitrate Transport, Sensing, and Responses in Plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Leran, S.; Varala, K.; Boyer, J.C.; Chiurazzi, M.; Crawford, N.; Daniel-Vedele, F.; David, L.; Dickstein, R.; Fernandez, E.; Forde, B.; et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDE TRANSPORTER family members in plants. Trends Plant Sci. 2014, 19, 5–9. [Google Scholar] [CrossRef]
- Wang, X.; Cai, X.; Xu, C.; Wang, Q. Identification and characterization of the NPF, NRT2 and NRT3 in spinach. Plant Physiol. Biochem. 2021, 158, 297–307. [Google Scholar] [CrossRef]
- Bai, H.; Euring, D.; Volmer, K.; Janz, D.; Polle, A. The nitrate transporter (NRT) gene family in poplar. PLoS ONE 2013, 8, e72126. [Google Scholar] [CrossRef] [PubMed]
- Tahir, M.M.; Wang, H.; Ahmad, B.; Liu, Y.; Fan, S.; Li, K.; Lei, C.; Shah, K.; Li, S.H.; Zhang, D. Identification and characterization of NRT gene family reveals their critical response to nitrate regulation during adventitious root formation and development in apple rootstock. Sci. Hortic-Amst. 2021, 275, 109642. [Google Scholar] [CrossRef]
- Orsel, M.; Krapp, A.; Daniel-Vedele, F. Analysis of the NRT2 nitrate transporter family in Arabidopsis. Structure and gene expression. Plant Physiol. 2002, 129, 886–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, Z.; Kotur, Z.; Glass, A.D. Characterization of an intact two-component high-affinity nitrate transporter from Arabidopsis roots. Plant J. 2010, 63, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.H.; Kuo, H.F.; Canivenc, G.; Lin, C.S.; Lepetit, M.; Hsu, P.K.; Tillard, P.; Lin, H.L.; Wang, Y.Y.; Tsai, C.B.; et al. Mutation of the Arabidopsis NRT1.5 nitrate transporter causes defective root-to-shoot nitrate transport. Plant Cell 2008, 20, 2514–2528. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis Nitrate Transporter NRT1.9 Is Important in Phloem Nitrate Transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Hsu, P.K.; Tsay, Y.F. Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 2013, 163, 844–856. [Google Scholar] [CrossRef] [Green Version]
- Fang, Z.M.; Bai, G.X.; Huang, W.T.; Wang, Z.X.; Wang, X.L.; Zhang, M.Y. The Rice Peptide Transporter OsNPF7.3 Is Induced by Organic Nitrogen, and Contributes to Nitrogen Allocation and Grain Yield. Front. Plant Sci. 2017, 8, 1338. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.S.; Zhang, X.X.; Chen, S.M.; Li, T.; Chen, Y.; Jiang, J.F.; Chen, F.D. Isolation and characterization of the Chrysanthemum nitrate transporter CmNRT1. Genet. Mol. Res. 2016, 15, gmr.15017148. [Google Scholar] [CrossRef]
- Hu, R.; Qiu, D.; Chen, Y.; Miller, A.J.; Fan, X.; Pan, X.; Zhang, M. Knock-Down of a Tonoplast Localized Low-Affinity Nitrate Transporter OsNPF7.2 Affects Rice Growth under High Nitrate Supply. Front. Plant Sci. 2016, 7, 1529. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N.M.; Siddiqi, M.Y.; Glass, A.D. Dissection of the AtNRT2.1: AtNRT2.2 inducible high-affinity nitrate transporter gene cluster. Plant Physiol. 2007, 143, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Cerezo, M.; Tillard, P.; Filleur, S.; Munos, S.; Daniel-Vedele, F.; Gojon, A. Major alterations of the regulation of root NO3-uptake are associated with the mutation of Nrt2.1 and Nrt2.2 genes in arabidopsis. Plant Physiol. 2001, 127, 262–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; Lezhneva, L.; Boutet-Mercey, S.; Orsel, M.; Brehaut, V.; Miller, A.; Daniel-Vedele, F.; Sakakibara, H.; et al. The Arabidopsis nitrate transporter NRT2.4 plays a double role in roots and shoots of nitrogen-starved plants. Plant Cell 2012, 24, 245–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lezhneva, L.; Kiba, T.; Feria-Bourrellier, A.B.; Lafouge, F.; Boutet-Mercey, S.; Zoufan, P.; Sakakibara, H.; Daniel-Vedele, F.; Krapp, A. The Arabidopsis nitrate transporter NRT2.5 plays a role in nitrate acquisition and remobilization in nitrogen-starved plants. Plant J. 2014, 80, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Chopin, F.; Orsel, M.; Dorbe, M.F.; Chardon, F.; Truong, H.N.; Miller, A.J.; Krapp, A.; Daniel-Vedele, F. The Arabidopsis ATNRT2.7 nitrate transporter controls nitrate content in seeds. Plant Cell 2007, 19, 1590–1602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, C.; Song, A.; Zhang, X.; Wang, H.; Li, T.; Chen, Y.; Jiang, J.; Chen, F.; Chen, S. Cloning of chrysanthemum high-affinity nitrate transporter family (CmNRT2) and characterization of CmNRT2.1. Sci. Rep. 2016, 6, 23462. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Li, J.Q.; Yan, Y.; Liu, W.Q.; Zhang, W.N.; Gao, L.H.; Tian, Y.Q. Knock-Down of CsNRT2.1, a Cucumber Nitrate Transporter, Reduces Nitrate Uptake, Root length, and Lateral Root Number at Low External Nitrate Concentration. Front. Plant Sci. 2018, 9, 722. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Qi, D.; Sun, J.; Zheng, X.; Peng, M. Expression of the cassava nitrate transporter NRT2.1 enables Arabidopsis low nitrate tolerance. J. Genet. 2019, 98, 74. [Google Scholar] [CrossRef]
- Liu, T.K.; Dai, W.; Sun, F.F.; Yang, X.D.; Xiong, A.S.; Hou, X.L. Cloning and characterization of the nitrate transporter gene BraNRT2.1 in non-heading Chinese cabbage. Acta Physiol. Plant 2014, 36, 815–823. [Google Scholar] [CrossRef]
- Kotur, Z.; Mackenzie, N.; Ramesh, S.; Tyerman, S.D.; Kaiser, B.N.; Glass, A.D.M. Nitrate transport capacity of the Arabidopsis thaliana NRT2 family members and their interactions with AtNAR2.1. New Phytol. 2012, 194, 724–731. [Google Scholar] [CrossRef]
- Morton, M.J.L.; Awlia, M.; Al-Tamimi, N.; Saade, S.; Pailles, Y.; Negrao, S.; Tester, M. Salt stress under the scalpel—dissecting the genetics of salt tolerance. Plant J. 2019, 97, 148–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Yassin, A. Influence of salinity on citrus A review paper. J. Cent. Eur. Agric. 2004, 5, 263–272. [Google Scholar]
- Miranda, R.D.; Gomes, E.; Prisco, J.T.; Alvarez-Pizarro, J.C. Ammonium improves tolerance to salinity stress in Sorghum bicolor plants. Plant Growth Regul. 2016, 78, 121–131. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of photosynthesis, nitrogen and proline metabolism to salinity stress in Solanum lycopersicum under different levels of nitrogen supplementation. Plant Physiol. Bioch. 2016, 109, 72–83. [Google Scholar] [CrossRef]
- Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Khan, N.A. Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol. Plant. 2020, 168, 490–510. [Google Scholar]
- Xiao, Q.Y.; Chen, Y.; Liu, C.W.; Robson, F.; Roy, S.; Cheng, X.F.; Wen, J.Q.; Mysore, K.; Miller, A.J.; Murray, J.D. MtNPF6.5 mediates chloride uptake and nitrate preference in Medicago roots. EMBO J. 2021, 40, e106847. [Google Scholar] [CrossRef] [PubMed]
- Ehlting, B.; Dluzniewska, P.; Dietrich, H.; Selle, A.; Teuber, M.; Hansch, R.; Nehls, U.; Polle, A.; Schnitzler, J.P.; Rennenberg, H.; et al. Interaction of nitrogen nutrition and salinity in Grey poplar (Populus tremula × alba). Plant Cell Environ. 2007, 30, 796–811. [Google Scholar] [CrossRef]
- Lv, X.F.; Yu, P.; Deng, W.H.; Li, Y.C. Transcriptomic analysis reveals the molecular adaptation to NaCl stress in Zostera marina L. Plant Physiol. Bioch. 2018, 130, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Del Castello, F.; Nejamkin, A.; Cassia, R.; Correa-Aragunde, N.; Fernandez, B.; Foresi, N.; Lombardo, C.; Ramirez, L.; Lamattina, L. The era of nitric oxide in plant biology: Twenty years tying up loose ends. Nitric Oxide-Biol. Ch. 2019, 85, 17–27. [Google Scholar] [CrossRef]
- Sykes, S.R. Chloride and sodium excluding capacities of citrus rootstock germplasm introduced to Australia from the People’s Republic of China. Sci. Hortic-Amst. 2011, 128, 443–449. [Google Scholar] [CrossRef]
- Kaleem, F.; Shabir, G.; Aslam, K.; Rasul, S.; Manzoor, H.; Shah, S.M.; Khan, A.R. An Overview of the Genetics of Plant Response to Salt Stress: Present Status and the Way Forward. Appl. Biochem. Biotechnol. 2018, 186, 306–334. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [PubMed]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [Green Version]
- Ren, Z.J.; Bai, F.L.; Xu, J.W.; Wang, L.; Wang, X.H.; Zhang, Q.; Feng, C.X.; Niu, Q.; Zhang, L.Y.; Song, J.L.; et al. A chloride efflux transporter, BIG RICE GRAIN 1, is involved in mediating grain size and salt tolerance in rice. J. Integr. Plant Biol. 2021, 63, 2150–2163. [Google Scholar] [CrossRef]
- Rombauts, S.; Dehais, P.; Van Montagu, M.; Rouze, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Li, J.Y.; Fu, Y.L.; Pike, S.M.; Bao, J.; Tian, W.; Zhang, Y.; Chen, C.Z.; Zhang, Y.; Li, H.M.; Huang, J.; et al. The Arabidopsis nitrate transporter NRT1.8 functions in nitrate removal from the xylem sap and mediates cadmium tolerance. Plant Cell 2010, 22, 1633–1646. [Google Scholar] [CrossRef] [Green Version]
- Komarova, N.Y.; Thor, K.; Gubler, A.; Meier, S.; Dietrich, D.; Weichert, A.; Suter Grotemeyer, M.; Tegeder, M.; Rentsch, D. AtPTR1 and AtPTR5 transport dipeptides in planta. Plant Physiol. 2008, 148, 856–869. [Google Scholar] [CrossRef]
- Almagro, A.; Lin, S.H.; Tsay, Y.F. Characterization of the Arabidopsis nitrate transporter NRT1.6 reveals a role of nitrate in early embryo development. Plant Cell 2008, 20, 3289–3299. [Google Scholar] [CrossRef] [Green Version]
- Dietrich, D.; Hammes, U.; Thor, K.; Suter-Grotemeyer, M.; Fluckiger, R.; Slusarenko, A.J.; Ward, J.M.; Rentsch, D. AtPTR1, a plasma membrane peptide transporter expressed during seed germination and in vascular tissue of Arabidopsis. Plant J. 2004, 40, 488–499. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Lin, C.S.; Hsia, A.P.; Su, R.C.; Lin, H.L.; Tsay, Y.F. Mutation of a nitrate transporter, AtNRT1:4, results in a reduced petiole nitrate content and altered leaf development. Plant Cell Physiol. 2004, 45, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.S.; Stacey, G.; Tsay, Y.F. Mechanisms and functional properties of two peptide transporters, AtPTR2 and fPTR2. J. Biol. Chem. 2004, 279, 30150–30157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, N.C.; Liu, K.H.; Lo, H.J.; Tsay, Y.F. Cloning and functional characterization of an Arabidopsis nitrate transporter gene that encodes a constitutive component of low-affinity uptake. Plant Cell 1999, 11, 1381–1392. [Google Scholar] [CrossRef] [Green Version]
- Rentsch, D.; Laloi, M.; Rouhara, I.; Schmelzer, E.; Delrot, S.; Frommer, W.B. Ntr1 Encodes a High-Affinity Oligopeptide Transporter in Arabidopsis. FEBS Lett. 1995, 370, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Karim, S.; Lundh, D.; Holmstrom, K.O.; Mandal, A.; Pirhonen, M. Structural and functional characterization of AtPTR3, a stress-induced peptide transporter of Arabidopsis. J. Mol. Model. 2005, 11, 226–236. [Google Scholar] [CrossRef]
- Song, W.; Steiner, H.Y.; Zhang, L.; Naider, F.; Stacey, G.; Becker, J.M. Cloning of a second arabidopsis peptide transport gene. Plant Physiol. 1996, 110, 171–178. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.C.; Lin, C.S.; Hsu, P.K.; Lin, S.H.; Tsay, Y.F. The Arabidopsis Nitrate Transporter NRT1.7, Expressed in Phloem, Is Responsible for Source-to-Sink Remobilization of Nitrate. Plant Cell 2009, 21, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.H.; Huang, C.Y.; Tsay, Y.F. CHL1 is a dual-affinity nitrate transporter of arabidopsis involved in multiple phases of nitrate uptake. Plant Cell 1999, 11, 865–874. [Google Scholar] [CrossRef]
- Okamoto, M.; Kumar, A.; Li, W.B.; Wang, Y.; Siddiqi, M.Y.; Crawford, N.M.; Glass, A.D.M. High-affinity nitrate transport in roots of Arabidopsis depends on expression of the NAR2-like gene AtNRT3.1. Plant Physiol. 2006, 140, 1036–1046. [Google Scholar] [CrossRef] [Green Version]
- Orsel, M.; Chopin, F.; Leleu, O.; Smith, S.J.; Krapp, A.; Daniel-Vedele, F.; Miller, A.J. Characterization of a two-component high-affinity nitrate uptake system in Arabidopsis. Physiology and protein-protein interaction. Plant Physiol. 2006, 142, 1304–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Fan, X.; Fan, X.; Liu, X.; Miller, A.J.; Xu, G. Multiple roles of nitrate transport accessory protein NAR2 in plants. Plant Signal. Behav. 2011, 6, 1286–1289. [Google Scholar] [CrossRef] [PubMed]
- Laugier, E.; Bouguyon, E.; Mauries, A.; Tillard, P.; Gojon, A.; Lejay, L. Regulation of high-affinity nitrate uptake in roots of Arabidopsis depends predominantly on posttranscriptional control of the NRT2.1/NAR2.1 transport system. Plant Physiol. 2012, 158, 1067–1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anil, V.S.; Harmon, A.C.; Rao, K.S. Spatio-temporal accumulation and activity of calcium-dependent protein kinases during embryogenesis, seed development, and germination in sandalwood. Plant Physiol. 2000, 122, 1035–1043. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number 1 | Gene ID | No. of AA 2 | Start Site 3 | End Site 3 | MW (kDa) 4 | pI 5 | GRAVY 6 |
|---|---|---|---|---|---|---|---|
| Pt1g007360 | PtrNPF1.1 | 585 | 6701392 | 6706813 | 64.94 | 9.24 | 0.316 |
| Pt7g010530 | PtrNPF1.2 | 625 | 12932806 | 12936421 | 68.80 | 8.47 | 0.293 |
| Pt1g005100 | PtrNPF1.3 | 572 | 3120786 | 3128240 | 62.63 | 8.73 | 0.282 |
| Pt1g007370 | PtrNPF1.4 | 594 | 6707620 | 6709906 | 65.68 | 8.73 | 0.200 |
| Pt8g002750 | PtrNPF1.5 | 576 | 1842390 | 1858910 | 63.44 | 8.50 | 0.311 |
| Pt1g005080 | PtrNPF1.6 | 581 | 3134219 | 3137466 | 63.91 | 7.44 | 0.265 |
| Pt3g001010 | PtrNPF2.1 | 588 | 1262243 | 1264918 | 65.37 | 9.36 | 0.224 |
| Pt3g001000 | PtrNPF2.2 | 621 | 1247077 | 1258790 | 68.69 | 9.03 | 0.188 |
| Pt3g001020 | PtrNPF2.3 | 599 | 1271264 | 1273237 | 66.08 | 9.05 | 0.249 |
| Pt4g003350 | PtrNPF2.4 | 615 | 2043405 | 2047306 | 68.49 | 9.18 | 0.224 |
| Pt6g014330 | PtrNPF2.5 | 589 | 10127132 | 10129206 | 65.21 | 9.09 | 0.225 |
| Pt6g013450 | PtrNPF3.1 | 874 | 9410246 | 9415048 | 96.73 | 6.48 | 0.199 |
| Pt4g008540 | PtrNPF3.2 | 581 | 6165780 | 6169493 | 64.65 | 8.98 | 0.250 |
| Pt4g008550 | PtrNPF3.3 | 581 | 6155010 | 6158555 | 64.31 | 8.90 | 0.285 |
| Pt2g029890 | PtrNPF4.1 | 895 | 28694854 | 28703982 | 98.89 | 7.25 | 0.204 |
| Pt4g012130 | PtrNPF4.2 | 524 | 9062853 | 9065920 | 58.15 | 9.05 | 0.328 |
| PtUn015880 | PtrNPF4.3 | 605 | 31889734 | 31893237 | 67.43 | 9.15 | 0.140 |
| Pt3g020700 | PtrNPF4.4 | 614 | 25583942 | 25587711 | 68.26 | 8.94 | 0.283 |
| Pt5g003870 | PtrNPF4.5 | 588 | 2587369 | 2590412 | 65.06 | 8.70 | 0.329 |
| Pt4g003240 | PtrNPF4.6 | 579 | 1982178 | 1987204 | 63.48 | 8.17 | 0.428 |
| Pt3g001060 | PtrNPF4.7 | 577 | 1312638 | 1315969 | 63.72 | 9.13 | 0.383 |
| PtUn010670 | PtrNPF4.8 | 616 | 21086318 | 21089327 | 67.70 | 7.03 | 0.256 |
| PtUn010660 | PtrNPF4.9 | 597 | 21037830 | 21048570 | 65.82 | 8.14 | 0.263 |
| Pt4g020770 | PtrNPF5.1 | 584 | 20356772 | 20360906 | 65.32 | 9.16 | 0.200 |
| Pt9g008600 | PtrNPF5.2 | 633 | 6405423 | 6415253 | 70.97 | 9.13 | 0.262 |
| PtUn035060 | PtrNPF5.3 | 595 | 67666483 | 67683281 | 66.58 | 9.18 | 0.237 |
| PtUn007180 | PtrNPF5.4 | 610 | 11116991 | 11120542 | 67.74 | 9.00 | 0.326 |
| Pt6g013470 | PtrNPF5.5 | 488 | 9424535 | 9426524 | 55.00 | 8.26 | 0.433 |
| Pt6g014250 | PtrNPF5.6 | 592 | 10060119 | 10064466 | 65.62 | 8.88 | 0.332 |
| Pt6g013460 | PtrNPF5.7 | 570 | 9416116 | 9423292 | 63.61 | 5.56 | 0.430 |
| Pt5g016390 | PtrNPF5.8 | 777 | 12824326 | 12831040 | 86.71 | 9.18 | 0.295 |
| Pt2g008110 | PtrNPF5.9 | 535 | 4826115 | 4830066 | 59.83 | 8.75 | 0.354 |
| Pt5g005040 | PtrNPF5.10 | 558 | 3350358 | 3352696 | 61.58 | 8.01 | 0.412 |
| PtUn026920 | PtrNPF5.11 | 498 | 54785836 | 54789355 | 55.57 | 9.61 | 0.225 |
| Pt5g005050 | PtrNPF5.12 | 569 | 3357900 | 3361719 | 62.83 | 6.95 | 0.369 |
| Pt9g009730 | PtrNPF5.13 | 431 | 7757020 | 7758583 | 48.69 | 8.30 | 0.132 |
| Pt9g009670 | PtrNPF5.14 | 448 | 7703996 | 7706735 | 50.57 | 8.80 | 0.186 |
| PtUn005790 | PtrNPF5.15 | 567 | 7939272 | 7942114 | 64.07 | 9.13 | 0.451 |
| Pt9g009790 | PtrNPF5.16 | 788 | 7812307 | 7815445 | 87.66 | 8.56 | 0.231 |
| Pt9g008580 | PtrNPF5.17 | 595 | 6395222 | 6398328 | 66.54 | 9.11 | 0.208 |
| Pt3g030260 | PtrNPF5.18 | 606 | 35941239 | 35944759 | 67.20 | 9.20 | 0.243 |
| PtUn028140 | PtrNPF5.19 | 628 | 56454824 | 56461168 | 70.25 | 9.42 | 0.370 |
| PtUn008620 | PtrNPF6.1 | 634 | 14160194 | 14162884 | 70.31 | 8.83 | 0.238 |
| Pt1g006350 | PtrNPF6.2 | 581 | 2152543 | 2155882 | 63.49 | 9.08 | 0.339 |
| Pt3g008120 | PtrNPF6.3 | 553 | 6492850 | 6496785 | 60.95 | 9.10 | 0.263 |
| Pt2g013780 | PtrNPF6.4 | 588 | 17449466 | 17451952 | 65.33 | 9.13 | 0.226 |
| Pt5g002490 | PtrNPF6.5 | 515 | 1693500 | 1693532 | 56.93 | 5.60 | 0.239 |
| Pt1g003070 | PtrNPF6.6 | 579 | 4853921 | 4857900 | 63.50 | 9.20 | 0.409 |
| Pt2g001610 | PtrNPF7.1 | 504 | 1224993 | 1228021 | 55.40 | 9.00 | 0.339 |
| Pt3g039860 | PtrNPF7.2 | 602 | 42571348 | 42574088 | 66.47 | 7.90 | 0.199 |
| Pt4g015230 | PtrNPF7.3 | 596 | 13417384 | 13421889 | 66.00 | 6.39 | 0.265 |
| Pt3g041260 | PtrNPF7.4 | 613 | 43574897 | 43579506 | 68.08 | 8.40 | 0.213 |
| Pt6g013550 | PtrNPF8.1 | 568 | 9499282 | 9503587 | 63.40 | 8.82 | 0.214 |
| Pt8g011520 | PtrNPF8.2 | 574 | 9266295 | 9270180 | 64.15 | 6.76 | 0.182 |
| Pt2g017400 | PtrNPF8.3 | 584 | 20631552 | 20636512 | 64.40 | 5.96 | 0.303 |
| Pt6g013540 | PtrNPF8.4 | 501 | 9494530 | 9496538 | 55.98 | 8.58 | 0.246 |
| Pt8g007700 | PtrNRT2.1 | 535 | 5808930 | 5811379 | 57.99 | 9.14 | 0.343 |
| Pt8g007710 | PtrNRT2.2 | 530 | 3120817 | 3122747 | 57.43 | 9.35 | 0.374 |
| Pt8g007720 | PtrNRT2.3 | 530 | 6554577 | 6556169 | 57.39 | 9.35 | 0.382 |
| Pt6g017660 | PtrNRT2.4 | 530 | 6537447 | 6539366 | 57.48 | 9.22 | 0.392 |
| Pt4g006700 | PtrNRT2.5 | 419 | 6559711 | 6561547 | 45.27 | 9.18 | 0.392 |
| Pt2g007200 | PtrNRT2.6 | 471 | 15634625 | 15636904 | 50.91 | 8.96 | 0.456 |
| Pt3g004750 | PtrNAR2.1 | 173 | 4426662 | 4427403 | 19.23 | 9.37 | −0.479 |
| Pt4g006240 | PtrNAR2.2 | 170 | 3424356 | 3425077 | 19.14 | 6.83 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Z.; Li, M.; Xu, W.; Liu, J.-H.; Li, C. Genome-Wide Identification of NRT Gene Family and Expression Analysis of Nitrate Transporters in Response to Salt Stress in Poncirus trifoliata. Genes 2022, 13, 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13071115
Zhao Z, Li M, Xu W, Liu J-H, Li C. Genome-Wide Identification of NRT Gene Family and Expression Analysis of Nitrate Transporters in Response to Salt Stress in Poncirus trifoliata. Genes. 2022; 13(7):1115. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13071115
Chicago/Turabian StyleZhao, Zeqi, Mengdi Li, Weiwei Xu, Ji-Hong Liu, and Chunlong Li. 2022. "Genome-Wide Identification of NRT Gene Family and Expression Analysis of Nitrate Transporters in Response to Salt Stress in Poncirus trifoliata" Genes 13, no. 7: 1115. https://0-doi-org.brum.beds.ac.uk/10.3390/genes13071115