Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition



1
Research Center on Animal Cognition (CRCA), Center for Integrative Biology (CBI), CNRS, University Paul Sabatier, 31062 Toulouse, France
2
Laboratoire Evolution et Diversité Biologique, UMR 5174 Centre National de la Recherche Scientifique, Université Paul Sabatier, ENSFEA, 31062 Toulouse, France
*
Author to whom correspondence should be addressed.
Genes 2018, 9(11), 534; https://0-doi-org.brum.beds.ac.uk/10.3390/genes9110534
Submission received: 29 September 2018
/
Revised: 29 October 2018
/
Accepted: 30 October 2018
/
Published: 6 November 2018
(This article belongs to the Special Issue Coevolution of Hosts and their Microbiome)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Microbes influence a wide range of host social behaviors and vice versa. So far, however, the mechanisms underpinning these complex interactions remain poorly understood. In social animals, where individuals share microbes and interact around foods, the gut microbiota may have considerable consequences on host social interactions by acting upon the nutritional behavior of individual animals. Here we illustrate how conceptual advances in nutritional ecology can help the study of these processes and allow the formulation of new empirically testable predictions. First, we review key evidence showing that gut microbes influence the nutrition of individual animals, through modifications of their nutritional state and feeding decisions. Next, we describe how these microbial influences and their social consequences can be studied by modelling populations of hosts and their gut microbiota into a single conceptual framework derived from nutritional geometry. Our approach raises new perspectives for the study of holobiont nutrition and will facilitate theoretical and experimental research on the role of the gut microbiota in the mechanisms and evolution of social behavior.
1. Introduction
Interactions between hosts and their microbiota, that together form the “holobiont” [1,2,3], influence various aspects of an animal’s biology, including nutrition [4] and behavior [5]. Growing evidence indicates that the microbiota can also have important consequences on the ways hosts interact with each other, for instance by triggering aggregations [6,7], guiding mate choice [8,9] or allowing kin discrimination [10]. How these complex host-microbe interactions are mediated is an open question. Recent studies point towards an effect of microbes on brain areas involved in the control of social interactions [11], or in the production of chemical signals mediating communication [12,13]. Here we argue that microbes that colonize the gut of animals may also influence a wide range of social behaviors through their impact on the nutritional needs and foraging decisions of animals.
Nutrition is central to all host-gut microbiota interactions [14]. On the one hand, gut microbes can influence the nutritional strategies of their hosts by expanding the host’s capability to digest and assimilate key nutrients [15,16], or by supplementing the host with nutrients that are difficult to find in the environment [17]. Termites, for instance, depend on complex microbial communities to exploit wood and soil nutrients that are otherwise not digestible by insects [18]. Microbes can also modify the feeding preferences of their hosts, for instance by attracting animals towards a diet that is suboptimal for the hosts but beneficial for the microbes [19]. On the other hand, nutrient intake by the hosts shapes the composition of their gut microbiota [18,20,21]. Variations in protein and carbohydrate intake modify the relative abundance of microbes in an animal’s gut [22], a change that can be observed within a few hours in mice [23], or a few days in humans [4].
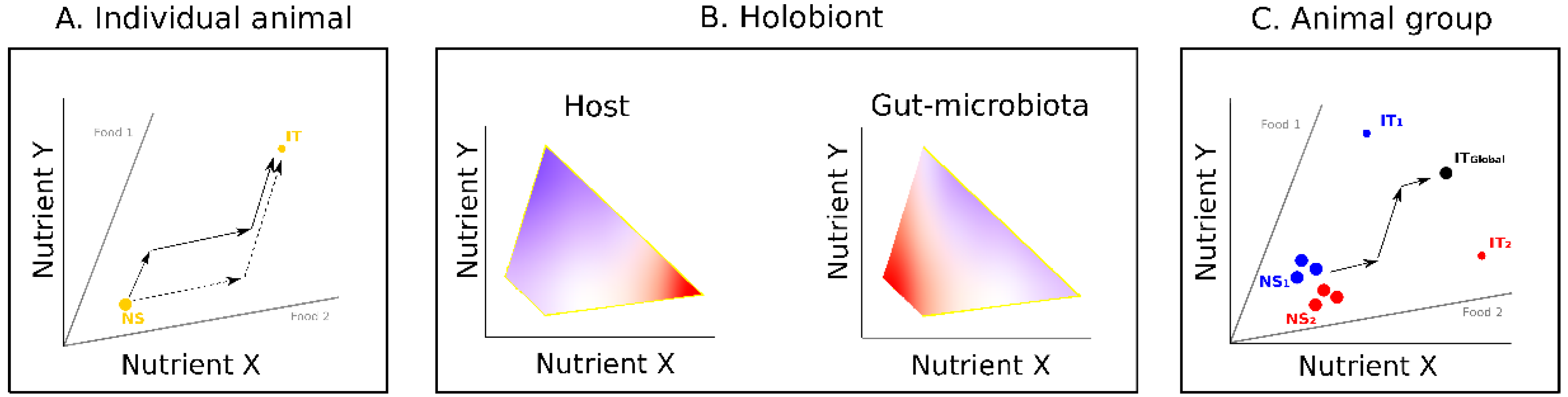
Over recent years, state space models of nutritional geometry [24,25] (Figure 1) have been increasingly used to study the nutritional interactions between gut microbes and their hosts, an approach known as “nutritional immunity” [14]. In these models, animals and foods are represented in a nutrient space defined by two or more key food components for the animal (typically, but not exclusively, the macronutrients carbohydrates, proteins and fats; for a recent review see [26]). Foods are radials defined by the ratio of the food components under consideration (nutritional rails). The animal is characterized by a nutritional state (NS), a snapshot of its nutritional condition, and by an intake target (IT), an optimal state at which fitness is maximized. The challenge for the animal is to consume available foods in amounts and balances allowing it to reach and maintain its IT [25]. Knowing the nutritional composition of foods, and the NS and the IT of the animal, it is thus possible to predict the feeding strategy that will most efficiently enable the animal to meet its nutritional needs (see examples in Figure 1A).
Nutritional immunology studies have extended this framework by considering microbes as trophic analogues of animals [27]. In this approach, an animal’s gut microbiota can be modelled in a nutrient space and characterized by a specific NS and IT [26]. Here, however, the NS of the gut microbiota varies with the nutritional behavior of its host. Using nutritional geometry designs, the performance responses of the host and the gut microbiota can be derived by experimentally mapping the expression of fitness traits of both holobiont components in separate nutrient spaces (see examples in Figure 1B). Therefore, in principle, it is possible to predict how the nutrient intake of a host will affect its fitness and the development of different microbe types (strains or species) in its gut [26].
Concepts of nutritional geometry have also been used to explore the effects of nutrition on social interactions, an approach called “social nutrition” [28,29,30]. Here, nutrition models are used to study how nutrition affects many social behaviors and structures, such as collective movements [31], cooperative foraging [32], or reproductive division of labor [33]. In social species, individuals must often trade-off between choosing foods that address their own nutritional needs and following others’ choices to maintain social cohesion, which can generate a variety of social responses [28]. These interacting effects between nutrition and social behaviors can be modelled by considering all individuals constituting a social group in a common nutrient space (Reference [28]; Figure 1C). Each individual is defined by its own NS and attempts to reach its IT. The ability of the different group members to do so depends on the nature and the frequency of their social interactions (e.g., attraction [34], competition [35,36], cooperation [37]). In social insects, for example, nutrient balance is achieved collectively by the foragers that collect nutrients to reach a colony IT composed of the different ITs of all colony members, including the other workers that need carbohydrates for energy, and the larvae that need proteins for growth [38].
In this review, we argue that concepts of nutritional geometry can bring new fundamental insights into emerging research on microbiota and social behaviors, by integrating nutritional interactions at different levels (between hosts and their gut microbes, and among hosts) in a common theoretical framework. We hypothesize that, by acting on the nutritional decisions of their hosts, gut microbes can influence a wide range of social behaviors, which in turn impact the composition of the gut microbiota communities. We develop this hypothesis, by first reviewing research demonstrating the links between the gut microbiota and the feeding behaviors of their hosts. We then describe how these multi-level interactions can be modelled in nutritional geometry to generate new empirical predictions about the influence of the gut microbiota on social behaviour. External microbes living on the surface of the animals may also influence a range of social behaviors [10,39,40,41,42,43]. However, it is unlikely that nutritional requirements of these microbes influence host nutritional decisions; at least before they are ingested by individuals, for instance through self-cleaning behaviors [44]. For these reasons, here we only focus on microbes that colonize the gut.
2. Influence of Gut Microbiota on Host Nutrition
The gut microbiota can modify the feeding preferences of their host. For instance, gnotobiotic fruit flies experimentally colonized with a specific bacterium species, and given a choice between several food sources with the same nutritional composition but seeded with different bacteria species, show an attraction towards the source seeded with the bacterium species present in their gut, whereas axenic (microbe free) flies show no preference [19]. This gut bacteria-mediated food preference is expressed by flies even if the food source is suboptimal (nutritionally imbalanced) and unseeded by bacteria, suggesting that the microbiota can cause important fitness costs by pushing the host away from its intake target [19].
How exactly gut microbes affect host nutritional behaviors is not clear. A major hypothesis is that microbes act through the gut-brain axis, a communication channel between the gut and the central nervous system [45,46], which involves regulatory processes that modify the gut microbial compositions and brain activity [46,47,48]. The gut-brain axis is involved in stress regulation, emotion, social bonding (reviewed by [49]), and eating disorders [50,51,52]. In mice, gut bacteria maintain the expression of receptors of fat that are altered in axenic individuals [53]. Axenic mice also show an increase in the number of sweet taste receptors in the gastro-intestinal tract compared to wild type non-axenic mice [54].
In principle, these complex interactions between gut microbiota composition and host feeding behaviors can be investigated using nutritional geometry. Previous studies have explored these effects by considering hosts and microbes in separate nutrient spaces (e.g., [26]; Figure 1B). Here we propose to integrate both components of the holobiont in a single nutrient space (Figure 2). In this approach, the host and the gut microbes have their own NS (animal host: NSA, microbes: NSM) defining a global holobiont nutritional state (NSH). The two components also have their own IT (animal host: ITA; microbes: ITM), defining a holobiont intake target (ITH). Every time the host ingests food, its microbial community is affected and can potentially change in composition (e.g., relative abundance of microbes or types of microbes [22]), which dynamically influences both the NSH and the ITH.
Several theoretical scenarios can then be considered. If the microbiota has no effect on the host feeding preferences, the host selects foods in order to reach and maintain its ITA (Figure 2A). In this case, the ITH cannot be reached. If the microbiota affects the host feeding preferences (e.g., [19]), the feeding behavior of the host mirrors the compromise between selective forces imposed by the host and the microbiota to reach their own intake targets. Nutrient intake by the host then depend on the relative influences of the two holobiont components in determining feeding decisions. If the nutritional interests of the host and its microbiota are in conflict any approach of the ITH towards the ITA moves the ITM away from its optimum, and vice versa. Depending on the composition of the microbial community, some microbe types may have more influence than others on the host behavior, and drag the nutrient intake towards their own ITM. Consequently, if only one microbe type colonizes the host (e.g., controlled experiments using gnotobiotic animals), host nutrient intake should follow nutritional rails leading somewhere between the ITA and the ITM and eventually ending on ITH (Figure 2B). In more natural conditions, where several microbe types colonize the host and compete (or cooperate) among themselves [55], the host nutrient intake should fall between the ITA and the different ITMs of all the microbe types, weighted for their relative influence on host feeding decisions (Figure 2C). In all cases, the microbiota can influence the NSA, by expanding the host’s capacity to assimilate nutrients [15,16], or by supplementing the host with new nutrients [17]. This may modify the host feeding choices, and lead the holobiont to reach the ITH. These interactions are expected to be highly dynamic since the composition of the gut microbiota can rapidly change as the host consumes different foods [4,22].
3. Integrating Gut Microbes, Host Nutrition and Social Behavior
In addition to influencing individual host behavior, microbes can affect the type and frequency of social interactions among hosts, for instance by triggering aggregations (e.g., drosophila [6]; cockroaches [7]), guiding sexual interactions (e.g., hyenas [9], drosophila [8]), or mediating kin recognition (e.g., ants [39]). Conversely, social behaviors can facilitate microbe transmission among hosts and shape their microbial communities by increasing the diversity of microbe types per individual [56] and by homogenizing communities across individuals [13]. Hosts that live together or interact frequently tend to have more similar microbiota than those that rarely encounter (e.g., baboons [57], meerkats [58], birds [59,60], bees [61]).
Based on these observations, we argue that gut microbes can play a major role in mediating host social interactions. By acting on the nutritional decisions of animals, microbes may affect the nature and frequency of social interactions among their hosts within groups or populations. Transfer of microbes between social partners may homogenize gut microbiota profiles within groups and further promote these interactions. Below we build on concepts of nutritional geometry to integrate host-microbiota interactions ([14,26]; Figure 1A,B and Figure 2) and host social behavior ([28,29]; Figure 1C) in a common framework.
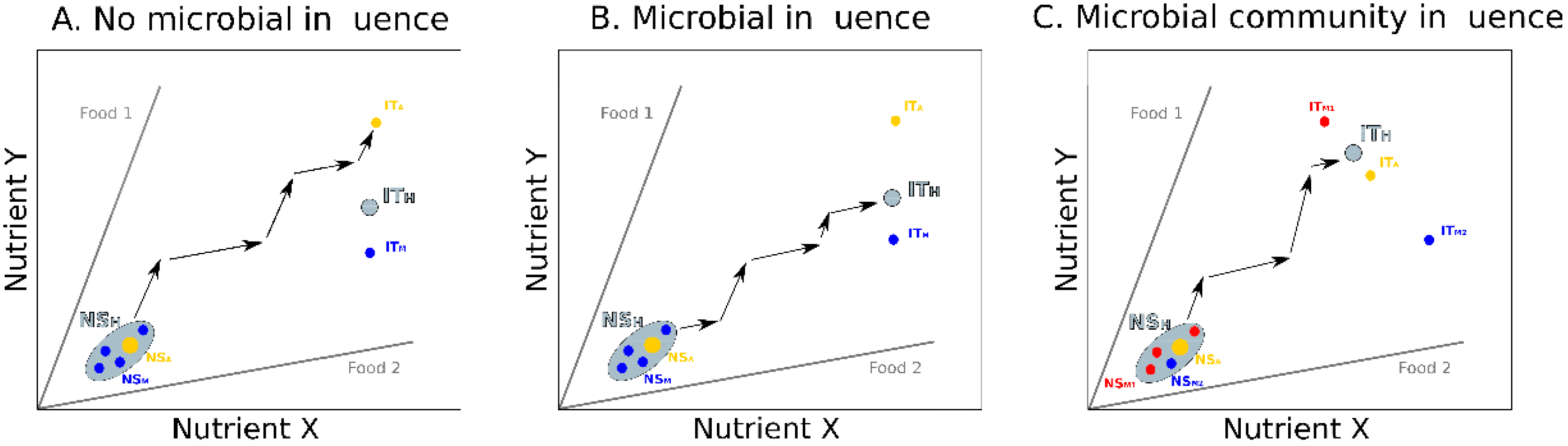
In this approach, multiple hosts and microbes are modelled in a single nutrient space, together forming a population of holobionts (Figure 3). As previously, holobionts have their own NSH and ITH. Each time an animal ingests food, its microbial community can be altered, thereby affecting both the NSH and the ITH. The microbiota composition can affect the nutritional decisions of each animal host depending on whether or not microbe types influence their feeding preferences. Here, however, the feeding decisions and microbiota composition of each holobiont also depend on the social interactions between holobionts. Social attraction, for instance, is expected to favor collective feeding and physical contacts leading to the homogenization of NSH and ITH among holobionts. In contrast, competition or social avoidance are expected to generate a high variability in NSH and ITH among holobionts, due to a reduced transmission of microbes.
Using this conceptual representation helps identify theoretical scenarios about how the gut microbiota may affect social interactions in groups exhibiting various levels of social complexity. In solitary species, where individuals rarely interact, the microbiota can be vertically transmitted across generations from mothers to offspring [62,63] or acquired from the environment [64]. In this case, we expect the NSH and the ITH of independently feeding individuals to be firstly influenced by vertical transmission and then primarily dependent on food intake, resulting in high variability across the nutrient space (Figure 3A). The degree of variability of the NSH and the ITH across the population of holobionts should mirror the diversity of the microbes available in the environment and the breadth of the hosts’ diet. This variability may be increased if microbes influence the nutritional decisions of the animal hosts, for instance by modifying their feeding preferences towards different food types [19]. In highly heterogeneous environments, such microbial influence on host feeding behaviors may further reduce the frequency of social interactions among animals searching for different food types, and reinforce the tendency for solitary behaviors.
In gregarious species, where individuals interact more frequently and often eat in groups, both vertical transmission and horizontal transmission (through social contacts) of microbes may occur, thereby reducing the variability in microbiota composition among hosts of the same group [65]. In this case, we expect a low variability of the NSH and the ITH across the holobionts (Figure 3B). If hosts do not interact evenly among each other, for instance following kinship or familiarity biases [66,67,68], the homogenization of the microbiota may give rise to sub-groups of interacting holobionts with more similar NSHs and ITHs, a mechanism further promoting social interactions within these sub-groups [7], and the emergence of a social structure.
In the most complex societies, characterized by strong social cohesion, frequent interactions and durable social structures, both vertical and horizontal transmissions of microbes occur. In such groups, microbes often need repeated inoculations to reach a mature functional form or assemblage useful to the host [69]. Here, continuous homogenization of gut microbiota through social contacts and differential social interactions among social partners may favor the emergence of specialized classes of highly interacting individuals with distinct NSHs and ITHs. This may be the case in social insects, where individuals at different developmental stages (e.g., larvae and adults) and belonging to different castes (breeders and workers) are characterized by highly divergent NSHs and ITHs [32,70,71]. Intra-class social interactions can lead hosts belonging to the same class towards situations where similar feeding choices strongly affect the microbiota composition, which in turn affect the hosts social behaviors. This feedback creates class specific NSHs from which the different holobionts cannot “escape”, since the specific condition of the hosts leads to an increase in the ITH distances (Figure 3C). At this stage, holobionts head on a nutritional rail that can be costly to change thus impeding movements of individuals from one class to another. A typical example is observed in ant colonies where individuals from different castes have different nutritional needs and transitions from one caste to another is accompanied by morphological, physiological and behavioral changes [72]. Alternately, intra-class variations of host nutritional choices can incur costs to the resident microbiota that can be detected by the loss of certain bacterial types, or the replacement by others. It can thus be predicted that associations between hosts and their gut microbiota are more specialized and stable in social hosts than in solitary hosts.
4. Conclusions
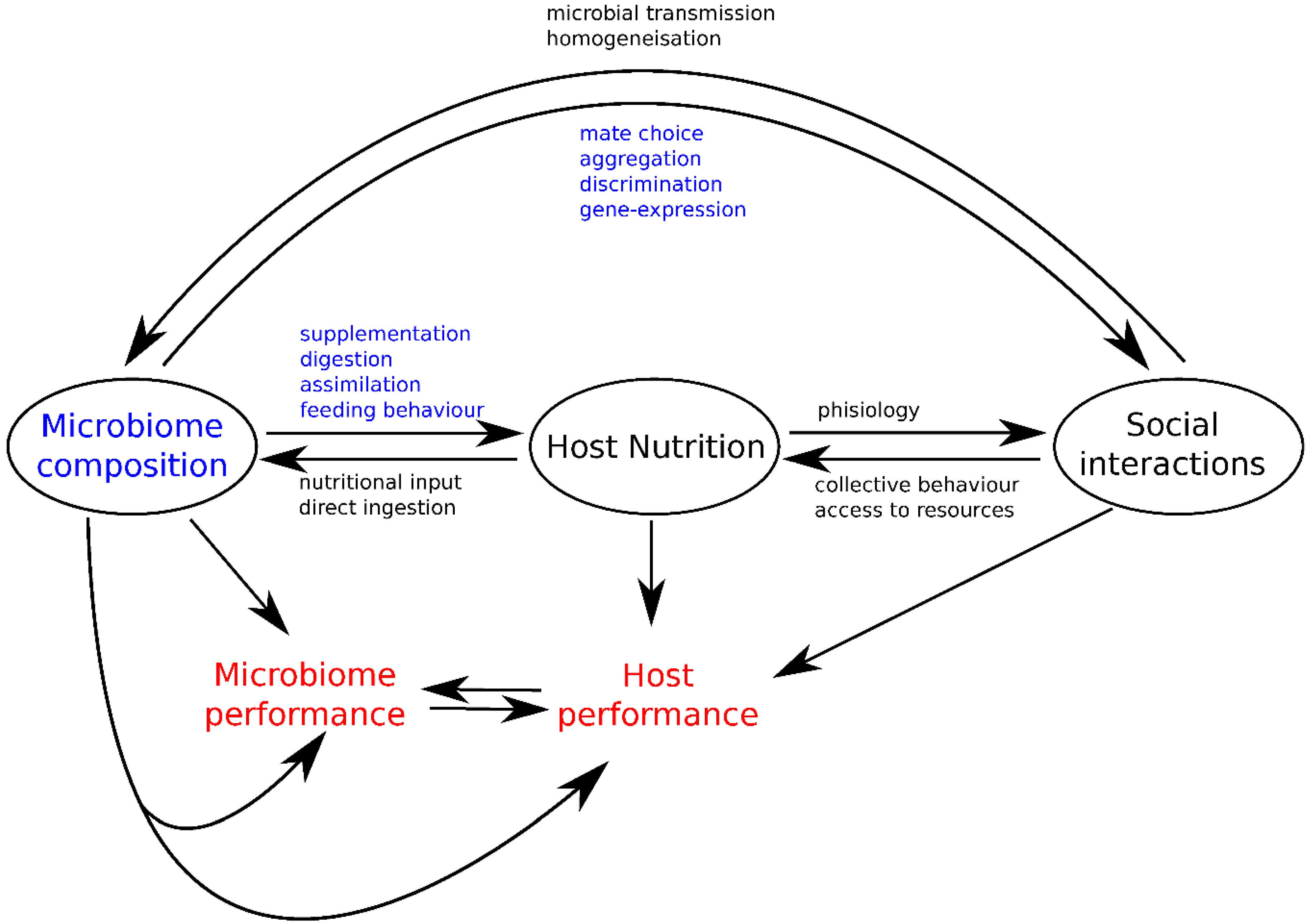
Nutritional geometry is a powerful approach in the study of nutritional interactions across organizational levels from molecules to individuals, groups and communities [28,73,74]. As we have described above, concepts of nutritional geometry can help in modelling how gut microbiota composition, host feeding behaviors and host social interactions integrate within a single theoretical framework. By identifying nutritional interactions and feedback loops between these three different components, this approach allows the formulation of new hypotheses to explore the role of the gut microbiota in the mechanisms and evolution of host social behaviors (Figure 4).
While nutritional studies have typically considered animal food intake and performance responses as a proxy of the intake target of an individual, growing evidence shows that these preferences vary among individuals, presumably because animal feeding decisions are influenced by several additional factors, including (but not only) its microbiota and social interactions [25]. Host nutritional choices can modify the relative abundance of microbe types in their gut by varying nutritional input available for microbial development [22] or through direct ingestion [18]. In turn, the gut microbiota composition can affect host nutrition by expanding [15], digesting and assimilating key nutrients for the host [17], or by dragging the host towards suboptimal diets [19]. Microbiota can intervene in gene-expression involved in social propensity [75], guiding mating choices [8,9], promoting aggregations [6,7], and mediating kin discrimination [39]. Conversely, social interactions favor the homogenization of the gut microbiota by increasing microbial transmission across individuals in the population [13,56]. Social interactions can also indirectly affect microbiota composition via the impact of collective behaviors during foraging [76] or differential access to resources [77].
The interplay between gut microbiota, host nutrition, and social interactions (Figure 4) emphasizes the need to study host-microbiota interactions using a more integrative approach, by considering populations of hosts and their feeding decisions. Using nutritional geometry, we derived speculative scenarios about how gut microbes may affect host social interactions and how host social interactions might constrain feeding decisions, and as a result, microbiota composition. The challenge for future studies will be to carry out experiments on specific components in order to examine the mechanisms underlying co-evolutionary feedback acting between hosts and their microbiota [55]. Deconstructing the nutritional needs and performance responses of the different components of host-microbe associations will help tackle important ongoing discussions about the relevance of the concept of the holobiont, whilst looking for potential evolutionary conflicts between microbes, host nutrition and social behaviors.
Computational models of nutritional geometry have already begun to explore nutritional interactions in populations of agents [73,78] and integrate multiple levels of interactions (e.g., host-pathogen interactions [37]). From an experimental point of view, it is possible to design manipulative experiments on host and microbiota nutrition, using behavioral assays or physiological measures on organisms provided with artificial diets [19]. Although it remains difficult to measure the intake target of microbe types in their natural host environment (where several microbes interact within assemblages), manipulating the microbiota composition under controlled conditions, using axenic [79,80] and gnotobiotic [19] animals, can help estimate the intake target of the animal host and how it is influenced by the microbiota. In the field, it is possible to reconstruct the gut microbiota composition of an animal and its nutrient intake by monitoring the animals’ food choices, and analyzing food samples and feces composition (see [81,82] for details). At the collective level, social network analyses constitute powerful tools to assess how gut microbiota influence social interactions and structures [83,84]. By quantifying social interactions and characterizing the nutritional needs of experimentally manipulated individuals, it is possible to picture the associations between microbiota-induced phenotypic changes (e.g., variation in holobiont nutritional needs) and the temporal dynamics of social interactions. This experimental approach might also be applied in species reared in their natural environments. For instance, using animal models that naturally show flexibility in their social behavior, either at different developmental stages or in different environments (i.e., social tipping points; see Reference [85] for details), it is possible to evaluate the variation of the onset of changes induced by the microbiota. Additionally, experimental manipulation of host nutrition and control of their social interactions, for instance using model organisms [19,86,87], can also help clarify the role of diet and social behavior in defining the effects of gut microbiota in hosts [44,60,88]. Since the basic principles of nutrition are shared by virtually all animal species [25], results from this approach may bring insights into the influence of the microbiota in the mechanisms and evolution of social life across a wide range of organisms.
Microbiota composition can be a key component shaping animal behaviors [5]. Thus, the interplay between microbiota and the host is central to understand multiple host interactions. The behavioral plasticity observed in the host can be seen as a combination of the interactions between environmental factors, and the genomic repertoire of the microbiota (i.e., microbiome) and the one of its host [89]. It is thus the holobiont itself that is plastic and able to cope with environmental and biological stressors (e.g., parasite infections [61,90]), possibly driving selection of both hosts and microbiota communities. Nutritional choice is one possible evolutionary channel of host diversity [91], shaping gut microbiota specialization [92], and driving the emergence of diverse social organizations across multiple taxa [65]. We hope that quantitative analyses of these interactions, as facilitated with nutritional geometry, will encourage future research in this direction.
Funding
This research was funded by a grant from the Agence Nationale de la Recherche to ML (ANR-16-CE02-0002-01). While writing, CP was a Postdoctoral Research Assistant on this grant. TGM was funded by a postdoctoral fellowship from the Fyssen Fondation. PH was funded by the ‘Laboratoire d’Excellence (LABEX)’ TULIP (ANR-10-LABX-41).
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Margulis, L.; Fester, R. Symbiosis as a Source of Evolutionary Innovation: Speciation and Morphogenesis; MIT Press: Cambridge, MA, USA, 1991; ISBN 0-262-13269-9. [Google Scholar]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Theis, K.R.; Dheilly, N.M.; Klassen, J.L.; Brucker, R.M.; Baines, J.F.; Bosch, T.C.; Cryan, J.F.; Gilbert, S.F.; Goodnight, C.J.; Lloyd, E.A. Getting the hologenome concept right: An eco-evolutionary framework for hosts and their microbiomes. Msystems 2016, 1. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezenwa, V.O.; Gerardo, N.M.; Inouye, D.W.; Medina, M.; Xavier, J.B. Animal behavior and the microbiome. Science 2012, 338, 198–199. [Google Scholar] [CrossRef] [PubMed]
- Venu, I.; Durisko, Z.; Xu, J.; Dukas, R. Social attraction mediated by fruit flies’ microbiome. J. Exp. Biol. 2014, 217, 1346–1352. [Google Scholar] [CrossRef] [PubMed]
- Wada-Katsumata, A.; Zurek, L.; Nalyanya, G.; Roelofs, W.L.; Zhang, A.; Schal, C. Gut bacteria mediate aggregation in the German cockroach. Proc. Natl. Acad. Sci. USA 2015, 112, 15678–15683. [Google Scholar] [CrossRef] [PubMed]
- Sharon, G.; Segal, D.; Ringo, J.M.; Hefetz, A.; Zilber-Rosenberg, I.; Rosenberg, E. Commensal bacteria play a role in mating preference of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2010, 107, 20051–20056. [Google Scholar] [CrossRef] [PubMed]
- Theis, K.R.; Venkataraman, A.; Dycus, J.A.; Koonter, K.D.; Schmitt-Matzen, E.N.; Wagner, A.P.; Holekamp, K.E.; Schmidt, T.M. Symbiotic bacteria appear to mediate hyena social odors. Proc. Natl. Acad. Sci. USA 2013, 110, 19832–19837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, K. Nestmate recognition mediated by intestinal bacteria in a termite, Reticulitermes speratus. Oikos 2001, 92, 20–26. [Google Scholar] [CrossRef]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Münger, E.; Montiel-Castro, A.J.; Langhans, W.; Pacheco-López, G. Reciprocal Interactions Between Gut Microbiota and Host Social Behavior. Front. Integr. Neurosci. 2018, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Lax, S.; Smith, D.P.; Hampton-Marcell, J.; Owens, S.M.; Handley, K.M.; Scott, N.M.; Gibbons, S.M.; Larsen, P.; Shogan, B.D.; Weiss, S. Longitudinal analysis of microbial interaction between humans and the indoor environment. Science 2014, 345, 1048–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponton, F.; Wilson, K.; Cotter, S.C.; Raubenheimer, D.; Simpson, S.J. Nutritional immunology: A multi-dimensional approach. PLoS Pathog. 2011, 7, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 2007, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D.; Stengel, A.; Dearing, M.D. Inoculation of tannin-degrading bacteria into novel hosts increases performance on tannin-rich diets. Environ. Microbiol. 2016, 18, 1720–1729. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Aphids and Their Symbiotic Bacteria Buchnera. Annu. Rev. Entomol. 1998, 17–37. [Google Scholar] [CrossRef] [PubMed]
- Nalepa, C.A.; Bignell, D.E.; Bandi, C. Detritivory, coprophagy, and the evolution of digestive mutualisms in Dictyoptera. Insectes Sociaux 2001, 48, 194–201. [Google Scholar] [CrossRef]
- Wong, A.C.N.; Wang, Q.P.; Morimoto, J.; Senior, A.M.; Lihoreau, M.; Neely, G.G.; Simpson, S.J.; Ponton, F. Gut Microbiota Modifies Olfactory-Guided Microbial Preferences and Foraging Decisions in Drosophila. Curr. Biol. 2017, 27, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.M.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Ravussin, Y.; Koren, O.; Spor, A.; LeDuc, C.; Gutman, R.; Stombaugh, J.; Knight, R.; Ley, R.E.; Leibel, R.L. Responses of gut microbiota to diet composition and weight loss in lean and obese mice. Obesity 2012, 20, 738–747. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.J.; Chew, Y.V.; Colakoglu, F.; Cliff, J.B.; Klaassens, E.; Read, M.N.; Solon-Biet, S.M.; McMahon, A.C.; Cogger, V.C.; Ruohonen, K.; et al. Diet-Microbiome Interactions in Health Are Controlled by Intestinal Nitrogen Source Constraints. Cell Metab. 2017, 25, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.; Ridaura, V.; Faith, J. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Raubenheimer, D. A multi-level analysis of feeding behaviour: The geometry of nutritional decisions. Phil. Trans. R. Soc. Lond. B 1993, 342, 381–402. [Google Scholar] [CrossRef]
- Simpson, S.J.; Raubenheimer, D. The Nature of Nutrition: A Unifying Framework from Animal Adaptation to Human Obesity; Princeton University Press: Princeton, NJ, USA, 2012; ISBN 1-4008-4280-8. [Google Scholar]
- Wong, A.C.N.; Holmes, A.; Ponton, F.; Lihoreau, M.; Wilson, K.; Raubenheimer, D.; Simpson, S.J. Behavioral microbiomics: A multi-dimensional approach to microbial influence on behavior. Front. Microbiol. 2015, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Steffan, S.A.; Chikaraishi, Y.; Currie, C.R.; Horn, H.; Gaines-Day, H.R.; Pauli, J.N.; Zalapa, J.E.; Ohkouchi, N. Microbes are trophic analogs of animals. Proc. Natl. Acad. Sci. USA 2015, 112, 15119–15124. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Buhl, J.; Charleston, M.A.; Sword, G.A.; Raubenheimer, D.; Simpson, S.J. Nutritional ecology beyond the individual: A conceptual framework for integrating nutrition and social interactions. Ecol. Lett. 2015, 18, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Buhl, J.; Charleston, M.A.; Sword, G.A.; Raubenheimer, D.; Simpson, S.J. Modelling nutrition across organizational levels: From individuals to superorganisms. J. Insect Physiol. 2014, 69, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Gómez-Moracho, T.; Pasquaretta, C.; Costa, J.T.; Buhl, J. Social nutrition: An emerging field in insect science. Curr. Opin. Insect Sci. 2018, 28, 73–80. [Google Scholar] [CrossRef]
- Buhl, J.; Sumpter, D.J.T.; Couzin, I.D.; Hale, J.J.; Despland, E.; Miller, E.R.; Simpson, S.J. From disorder to order in marching locusts. Science 2006, 312, 1402–1406. [Google Scholar] [CrossRef] [PubMed]
- Dussutour, A.; Simpson, S.J. Communal nutrition in ants. Curr. Biol. 2009, 19, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Salomon, M.; Mayntz, D.; Lubin, Y. Colony nutrition skews reproduction in a social spider. Behav. Ecol. 2008, 19, 605–611. [Google Scholar] [CrossRef] [Green Version]
- Senior, A.M.; Nakagawa, S.; Lihoreau, M.; Simpson, S.J.; Raubenheimer, D. An overlooked consequence of dietary mixing: A varied diet reduces interindividual variance in fitness. Am. Nat. 2015, 186, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.M.; Charleston, M.A.; Lihoreau, M.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Evolving nutritional strategies in the presence of competition: A geometric agent-based model. PLoS Comput. Biol. 2015, 11, e1004111. [Google Scholar] [CrossRef] [PubMed]
- Senior, A.M.; Lihoreau, M.; Charleston, M.A.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Adaptive collective foraging in groups with conflicting nutritional needs. R. Soc. Open Sci. 2016, 3, 150638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poissonnier, L.-A.; Lihoreau, M.; Gomez-Moracho, T.; Dussutour, A.; Buhl, J. A theoretical exploration of dietary collective medication in social insects. J. Insect Physiol. 2018, 106, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T. Animal behaviour: Feeding the superorganism. Curr. Biol. 2009, 19, R366–R368. [Google Scholar] [CrossRef] [PubMed]
- Dosmann, A.; Bahet, N.; Gordon, D.M. Experimental modulation of external microbiome affects nestmate recognition in harvester ants (Pogonomyrmex barbatus). PeerJ 2016, 4, e1566. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Tissot, M.; Cuevas, W.; Gordon, D.M. Harvester ants utilize cuticular hydrocarbons in nestmate recognition. J. Chem. Ecol. 2000, 26, 2245–2257. [Google Scholar] [CrossRef]
- Wagner, D.; Tissot, M.; Gordon, D. Task-related environment alters the cuticular hydrocarbon composition of harvester ants. J. Chem. Ecol. 2001, 27, 1805–1819. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.W.; Blomquist, G.J. Ecological, Behavioral, and Biochemical Aspects of Insect Hydrocarbons. Annu. Rev. Entomol. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Esponda, F.; Gordon, D.M. Distributed nestmate recognition in ants. Proc. R. Soc. B 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.; Heeb, P. Social and sexual behaviours aid transmission of bacteria in birds. Behav. Process. 2007, 74, 88–92. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306. [Google Scholar] [CrossRef] [PubMed]
- Mayer, E.A. Gut feelings: The emerging biology of gut-brain communication. Nat. Rev. Neurosci. 2011, 12, 453. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Kaelberer, M.M.; Buchanan, K.L.; Klein, M.E.; Barth, B.B.; Montoya, M.M.; Shen, X.; Bohórquez, D.V. A gut-brain neural circuit for nutrient sensory transduction. Science 2018, 361, eaat5236. [Google Scholar] [CrossRef] [PubMed]
- Montiel-Castro, A.J.; González-Cervantes, R.M.; Bravo-Ruiseco, G.; Pacheco-López, G. The microbiota-gut-brain axis: Neurobehavioral correlates, health and sociality. Front. Integr. Neurosci. 2013, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Konturek, S.; Konturek, P.; Pawlik, T.; Brzozowski, T. Brain-gut axis and its role in the control of food intake. J. Physiol. Pharmacol. 2004, 55, 137–154. [Google Scholar] [PubMed]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. BioEssays 2014, 36, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleiman, S.C.; Watson, H.J.; Bulik-Sullivan, E.C.; Huh, E.Y.; Tarantino, L.M.; Bulik, C.M.; Carroll, I.M. The intestinal microbiota in acute anorexia nervosa and during renourishment: Relationship to depression, anxiety, and eating disorder psychopathology. Psychosom. Med. 2015, 77, 969. [Google Scholar] [CrossRef] [PubMed]
- Duca, F.A.; Swartz, T.D.; Sakar, Y.; Covasa, M. Increased oral detection, but decreased intestinal signaling for fats in mice lacking gut microbiota. PLoS ONE 2012, 7, e39748. [Google Scholar] [CrossRef] [PubMed]
- Swartz, T.D.; Duca, F.; De Wouters, T.; Sakar, Y.; Covasa, M. Up-regulation of intestinal type 1 taste receptor 3 and sodium glucose luminal transporter-1 expression and increased sucrose intake in mice lacking gut microbiota. Br. J. Nutr. 2012, 107, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Foster, K.R.; Schluter, J.; Coyte, K.Z.; Rakoff-Nahoum, S. The evolution of the host microbiome as an ecosystem on a leash. Nature 2017, 548, 43. [Google Scholar] [CrossRef] [PubMed]
- Moeller, A.H.; Foerster, S.; Wilson, M.L.; Pusey, A.E.; Hahn, B.H.; Ochman, H. Social behavior shapes the chimpanzee pan-microbiome. Sci. Adv. 2016, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, J.; Barreiro, L.B.; Burns, M.B.; Grenier, J.C.; Lynch, J.; Grieneisen, L.E.; Altmann, J.; Alberts, S.C.; Blekhman, R.; Archie, E.A. Social networks predict gut microbiome composition in wild baboons. eLife 2015, 2015, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Leclaire, S.; Nielsen, J.F.; Drea, C.M. Bacterial communities in meerkat anal scent secretions vary with host sex, age, and group membership. Behav. Ecol. 2014, 25, 996–1004. [Google Scholar] [CrossRef] [Green Version]
- Lucas, F.S.; Heeb, P. Environmental factors shape cloacal bacterial assemblages in great tit Parus major and blue tit P. caeruleus nestlings. J. Avian Biol. 2005, 36, 510–516. [Google Scholar] [CrossRef]
- White, J.; Mirleau, P.; Danchin, E.; Mulard, H.; Hatch, S.A.; Heeb, P.; Wagner, R.H. Sexually transmitted bacteria affect female cloacal assemblages in a wild bird. Ecol. Lett. 2010, 13, 1515–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, H.; Schmid-Hempel, P. Socially transmitted gut microbiota protect bumble bees against an intestinal parasite. Proc. Natl. Acad. Sci. USA 2011, 108, 19288–19292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bright, M.; Bulgheresi, S. A complex journey: Transmission of microbial symbionts. Nat. Rev. Microbiol. 2010, 8, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Salem, H.; Florez, L.; Gerardo, N.; Kaltenpoth, M. An out-of-body experience: The extracellular dimension for the transmission of mutualistic bacteria in insects. Proc. R. Soc. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Caspi-Fluger, A.; Inbar, M.; Mozes-Daube, N.; Katzir, N.; Portnoy, V.; Belausov, E.; Hunter, M.S.; Zchori-Fein, E. Horizontal transmission of the insect symbiont Rickettsia is plant-mediated. Proc. R. Soc. B Biol. Sci. 2012, 279, 1791–1796. [Google Scholar] [CrossRef] [PubMed]
- Lombardo, M.P. Access to mutualistic endosymbiotic microbes: An underappreciated benefit of group living. Behav. Ecol. Sociobiol. 2008, 62, 479–497. [Google Scholar] [CrossRef]
- Barber, I.; Wright, H.A. How strong are familiarity preferences in shoaling fish? Anim. Behav. 2001, 61, 975–979. [Google Scholar] [CrossRef]
- Wiszniewski, J.; Lusseau, D.; Möller, L.M. Female bisexual kinship ties maintain social cohesion in a dolphin network. Anim. Behav. 2010, 80, 895–904. [Google Scholar] [CrossRef]
- Kurvers, R.H.J.M.; Adamczyk, V.M.A.P.; Kraus, R.H.S.; Hoffman, J.I.; van Wieren, S.E.; van der Jeugd, H.P.; Amos, W.; Prins, H.H.T.; Jonker, R.M. Contrasting context dependence of familiarity and kinship in animal social networks. Anim. Behav. 2013, 1–9. [Google Scholar] [CrossRef]
- Nalepa, C.A. Nourishment and the origin of termite eusociality. In Nourishment and Evolution in Insect Societies; Westview Press: Boulder, CO, USA, 1994; pp. 57–104. [Google Scholar]
- Moran, N.A.; Hansen, A.K.; Powell, J.E.; Sabree, Z.L. Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS ONE 2012, 7, e36393. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.C.; Fruciano, C.; Hildebrand, F.; Al Toufalilia, H.; Balfour, N.J.; Bork, P.; Engel, P.; Ratnieks, F.L.W.; Hughes, W.O.H. Gut microbiota composition is associated with environmental landscape in honey bees. Ecol. Evol. 2018, 8, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Wilson, E.O. The Insect Societies; Harvard University Press: Cambridge, MA, USA, 1971. [Google Scholar]
- Simpson, S.J.; Raubenheimer, D.; Charleston, M.A.; Clissold, F.J. Modelling nutritional interactions: From individuals to communities. Trends Ecol. Evol. 2010, 25, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.J.; Clissold, F.J.; Lihoreau, M.; Ponton, F.; Wilder, S.M.; Raubenheimer, D. Recent Advances in the Integrative Nutrition of Arthropods. Annu. Rev. Entomol. 2015, 60, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Stilling, R.M.; Moloney, G.M.; Ryan, F.J.; Hoban, A.E.; Bastiaanssen, T.F.; Shanahan, F.; Clarke, G.; Claesson, M.J.; Dinan, T.G.; Cryan, J.F. Social interaction-induced activation of RNA splicing in the amygdala of microbiome-deficient mice. eLife 2018, 7, e33070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elgar, M.A. Predator vigilance and group size in mammals and birds: A critical review of the empirical evidence. Biol. Rev. Camb. Philos. Soc. 1989, 64, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Kurvers, R.H.J.M.; van Oers, K.; Nolet, B.A.; Jonker, R.M.; van Wieren, S.E.; Prins, H.H.T.; Ydenberg, R.C. Personality predicts the use of social information. Ecol. Lett. 2010, 13, 829–837. [Google Scholar] [CrossRef] [PubMed]
- Lihoreau, M.; Charleston, M.A.; Senior, A.M.; Clissold, F.J.; Raubenheimer, D.; Simpson, S.J.; Buhl, J. Collective foraging in spatially complex nutritional environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160238. [Google Scholar] [CrossRef] [PubMed]
- Storelli, G.; Defaye, A.; Erkosar, B.; Hols, P.; Royet, J.; Leulier, F. Lactobacillus plantarum promotes Drosophila systemic growth by modulating hormonal signals through TOR-dependent nutrient sensing. Cell Metab. 2011, 14, 403–414. [Google Scholar] [CrossRef] [PubMed]
- Callens, M.; Macke, E.; Muylaert, K.; Bossier, P.; Lievens, B.; Waud, M.; Decaestecker, E. Food availability affects the strength of mutualistic host-microbiota interactions in Daphnia magna. ISME J. 2016, 10, 911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, C.A.; Raubenheimer, D.; Rothman, J.M.; Clarke, D.; Swedell, L. 30 Days in the life: Daily nutrient balancing in a wild chacma baboon. PLoS ONE 2013, 8, e70383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raubenheimer, D.; Machovsky-Capuska, G.E.; Chapman, C.A.; Rothman, J.M. Geometry of nutrition in field studies: An illustration using wild primates. Oecologia 2015, 177, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Vitousek, M.N.; Zonana, D.M.; Safran, R.J. An integrative view of the signaling phenotype: Dynamic links between signals, physiology, behavior and social context. Curr. Zool. 2014, 60, 739–754. [Google Scholar] [CrossRef] [Green Version]
- Senior, A.M.; Lihoreau, M.; Buhl, J.; Raubenheimer, D.; Simpson, S.J. Social network analysis and nutritional behavior: An integrated modeling approach. Front. Psychol. 2016, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Pruitt, J.N.; Berdahl, A.; Riehl, C.; Pinter-Wollman, N.; Moeller, H.V.; Pringle, E.G.; Aplin, L.M.; Robinson, E.J.; Grilli, J.; Yeh, P. Social tipping points in animal societies. Proc. R. Soc. B 2018, 285, 20181282. [Google Scholar] [CrossRef] [PubMed]
- Sokolowski, M.B. Foraging strategies of Drosophila melanogaster: A chromosomal analysis. Behav. Genet. 1980, 10, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Pasquaretta, C.; Battesti, M.; Klenschi, E.; Bousquet, C.A.; Sueur, C.; Mery, F. How social network structure affects decision-making in Drosophila melanogaster. Proc. Biol. Sci. 2016, 283. [Google Scholar] [CrossRef] [PubMed]
- Kort, R.; Caspers, M.; van de Graaf, A.; van Egmond, W.; Keijser, B.; Roeselers, G. Shaping the oral microbiota through intimate kissing. Microbiome 2014, 2, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Zepeda-Mendoza, M.L.; Gilbert, M.T.P. Do vertebrate gut metagenomes confer rapid ecological adaptation? Trends Ecol. Evol. 2016, 31, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Cho, I.; Yamanishi, S.; Cox, L.; Methé, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621. [Google Scholar] [CrossRef] [PubMed]
- Delsuc, F.; Metcalf, J.L.; Wegener Parfrey, L.; Song, S.J.; González, A.; Knight, R. Convergence of gut microbiomes in myrmecophagous mammals. Mol. Ecol. 2014, 23, 1301–1317. [Google Scholar] [CrossRef] [PubMed]
- Roggenbuck, M.; Schnell, I.B.; Blom, N.; Bælum, J.; Bertelsen, M.F.; Sicheritz-Pontén, T.; Sørensen, S.J.; Gilbert, M.T.P.; Graves, G.R.; Hansen, L.H. The microbiome of New World vultures. Nat. Commun. 2014, 5, 5498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Nutritional geometry models for individual animals, holobionts, and animal groups. (A) Model for a hypothetical animal (modified from [25]). The nutrient space is defined by two nutrients. Nutritional rails (grey lines) represent the ratio of nutrients X and Y in two foods. Yellow dots represent the nutritional state (NS) and the intake target (IT) of the animal. In this example, foods 1 and 2 are individually imbalanced (ratio different from the animal’s IT) but collectively complementary (fall in opposite sides of the animal’s IT): The animal can reach its IT by combining its intake from the two foods. Sequences of arrows show two possible strategies. (B) Model for a hypothetical holobiont (modified from [26]). The performance responses of holobiont components (host and gut microbes) to nutrient intake are shown in separate heatmap landscapes. The strength of response of each holobiont component is indicated by a color code (warmer colors show strongest responses). In this example, changes in nutrient intake have different influences on the fitness of the host (maximum for high X low Y intake) and the gut microbiota (maximum for low X low Y intake) indicating the potential for a conflict in food requirements. (C) Model for a hypothetical group of animals (modified from [28]). The group is composed of two sub-groups of highly interacting individuals (blue and red circles). In this example, each individual has its own NS but individuals of the same sub-group have the same IT (sub-group 1: IT1; sub-group 2: IT2). All individuals are socially attracted and feed together on the same foods. Animals must trade-off between choosing food to address their own nutritional needs and following others. This can be achieved by collectively reaching a group-level intake target (IT global) midway between the two sub-group ITs.
Figure 1.
Nutritional geometry models for individual animals, holobionts, and animal groups. (A) Model for a hypothetical animal (modified from [25]). The nutrient space is defined by two nutrients. Nutritional rails (grey lines) represent the ratio of nutrients X and Y in two foods. Yellow dots represent the nutritional state (NS) and the intake target (IT) of the animal. In this example, foods 1 and 2 are individually imbalanced (ratio different from the animal’s IT) but collectively complementary (fall in opposite sides of the animal’s IT): The animal can reach its IT by combining its intake from the two foods. Sequences of arrows show two possible strategies. (B) Model for a hypothetical holobiont (modified from [26]). The performance responses of holobiont components (host and gut microbes) to nutrient intake are shown in separate heatmap landscapes. The strength of response of each holobiont component is indicated by a color code (warmer colors show strongest responses). In this example, changes in nutrient intake have different influences on the fitness of the host (maximum for high X low Y intake) and the gut microbiota (maximum for low X low Y intake) indicating the potential for a conflict in food requirements. (C) Model for a hypothetical group of animals (modified from [28]). The group is composed of two sub-groups of highly interacting individuals (blue and red circles). In this example, each individual has its own NS but individuals of the same sub-group have the same IT (sub-group 1: IT1; sub-group 2: IT2). All individuals are socially attracted and feed together on the same foods. Animals must trade-off between choosing food to address their own nutritional needs and following others. This can be achieved by collectively reaching a group-level intake target (IT global) midway between the two sub-group ITs.

Figure 2.
Influence of the gut microbiota on host nutrition. In these hypothetical examples, hosts and microbes are represented in a common nutrient space defined by two nutrients. Food types (grey lines) are defined by their ratio in nutrients X and Y. The animal host (yellow dot) and the microbes (blue and red dots), together forming the holobiont (grey surface), are charatcerized by their own nutritional states (animal: NSA; microbe: NSM; holobiont: NSH) and intake targets (animal: ITA; microbe: ITM; holobiont: ITH). (A) The animal is colonized by a single microbe type that does not influence its feeding behavior. The animal can reach the ITA by combining its intake of the two complementary foods (black arrows). The optimal ITH for both holobiont components is never reached. (B) The animal is colonized by a single microbe type that influences its feeding behaviors. The animal can reach an ITH that falls between ITA and ITM depending on the relative influence of the host and the microbes on feeding decisions. (C) The animal is colonized by two microbe types (assemblage of microbes) that both influence its feeding behavior. The animal can reach an ITH that falls between ITA, ITM1 and ITM2 depending on the relative influence of the host and the two microbe types on feeding decisions. In this example, microbe type 1 has a stronger influence on the feeding decisions of the host than microbe type 2. The same logic can be extended to richer microbial communities where a global effect of the microbial community can affect the animal feeding behavior.
Figure 2.
Influence of the gut microbiota on host nutrition. In these hypothetical examples, hosts and microbes are represented in a common nutrient space defined by two nutrients. Food types (grey lines) are defined by their ratio in nutrients X and Y. The animal host (yellow dot) and the microbes (blue and red dots), together forming the holobiont (grey surface), are charatcerized by their own nutritional states (animal: NSA; microbe: NSM; holobiont: NSH) and intake targets (animal: ITA; microbe: ITM; holobiont: ITH). (A) The animal is colonized by a single microbe type that does not influence its feeding behavior. The animal can reach the ITA by combining its intake of the two complementary foods (black arrows). The optimal ITH for both holobiont components is never reached. (B) The animal is colonized by a single microbe type that influences its feeding behaviors. The animal can reach an ITH that falls between ITA and ITM depending on the relative influence of the host and the microbes on feeding decisions. (C) The animal is colonized by two microbe types (assemblage of microbes) that both influence its feeding behavior. The animal can reach an ITH that falls between ITA, ITM1 and ITM2 depending on the relative influence of the host and the two microbe types on feeding decisions. In this example, microbe type 1 has a stronger influence on the feeding decisions of the host than microbe type 2. The same logic can be extended to richer microbial communities where a global effect of the microbial community can affect the animal feeding behavior.

Figure 3.
Integrating microbiota, host nutrition and social behavior. Populations of holobionts are represented in a nutrient space defined by two nutrients. Foods (grey lines) are defined by their ratio in nutrients X and Y. Animal hosts (yellow dots), gut microbes (blue and red dots), and holobionts (grey surfaces) are characterized by their nutritional states (animal: NSA; microbe: NSM; holobiont: NSH) and intake targets (animal: ITA; microbe: ITM; holobiont: ITH). Black arrows illustrate the feeding decisions of each holobiont or sub-groups of interacting holobionts. Three hypothetical scenarios depict the expected nutritional interactions between gut microbes and hosts in groups of increasing social complexity. (A) In solitary species, where individuals feed independently and do not interact, the diversity of environmental microbes may generate a high variability in the NSH and the ITH of the different holobionts. Each holobiont can reach its ITH by alternating its intake of the two complementary foods. (B) In gregarious species, where individuals feed together and regularly interact, horizontal transmission of microbes may reduce inter-holobionts variability of NSHs and ITHs. In this example where individuals do not interact evenly, horizontal transmission favors the emergence of sub-groups (black circles) of highly interacting individuals with similar gut microbiota that can collectively reach their ITH. (C) In advanced societies, where individuals with different nutritional needs (e.g., young and adults, queens and workers) permanently co-occur, differential social interactions and continuous homogenization of gut microbiota through social contacts favor the emergence of classes of highly interacting individuals with distinct NSHs and ITHs (e.g., castes). Holobionts can cooperate with other holobionts from the same class to reach their common ITH.
Figure 3.
Integrating microbiota, host nutrition and social behavior. Populations of holobionts are represented in a nutrient space defined by two nutrients. Foods (grey lines) are defined by their ratio in nutrients X and Y. Animal hosts (yellow dots), gut microbes (blue and red dots), and holobionts (grey surfaces) are characterized by their nutritional states (animal: NSA; microbe: NSM; holobiont: NSH) and intake targets (animal: ITA; microbe: ITM; holobiont: ITH). Black arrows illustrate the feeding decisions of each holobiont or sub-groups of interacting holobionts. Three hypothetical scenarios depict the expected nutritional interactions between gut microbes and hosts in groups of increasing social complexity. (A) In solitary species, where individuals feed independently and do not interact, the diversity of environmental microbes may generate a high variability in the NSH and the ITH of the different holobionts. Each holobiont can reach its ITH by alternating its intake of the two complementary foods. (B) In gregarious species, where individuals feed together and regularly interact, horizontal transmission of microbes may reduce inter-holobionts variability of NSHs and ITHs. In this example where individuals do not interact evenly, horizontal transmission favors the emergence of sub-groups (black circles) of highly interacting individuals with similar gut microbiota that can collectively reach their ITH. (C) In advanced societies, where individuals with different nutritional needs (e.g., young and adults, queens and workers) permanently co-occur, differential social interactions and continuous homogenization of gut microbiota through social contacts favor the emergence of classes of highly interacting individuals with distinct NSHs and ITHs (e.g., castes). Holobionts can cooperate with other holobionts from the same class to reach their common ITH.

Figure 4.
Nutritional interactions between the gut microbiota, host feeding strategies and host social interactions.
Figure 4.
Nutritional interactions between the gut microbiota, host feeding strategies and host social interactions.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pasquaretta, C.; Gómez-Moracho, T.; Heeb, P.; Lihoreau, M. Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes 2018, 9, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9110534
AMA Style
Pasquaretta C, Gómez-Moracho T, Heeb P, Lihoreau M. Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes. 2018; 9(11):534. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9110534
Chicago/Turabian StylePasquaretta, Cristian, Tamara Gómez-Moracho, Philipp Heeb, and Mathieu Lihoreau. 2018. "Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition" Genes 9, no. 11: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/genes9110534
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.