
Holocene Vegetation Dynamics, Landscape Change and Human Impact in Western Ireland as Revealed by Multidisciplinary, Palaeoecological Investigations of Peat Deposits and Bog-Pine in Lowland Connemara


Abstract
:
1. Introduction
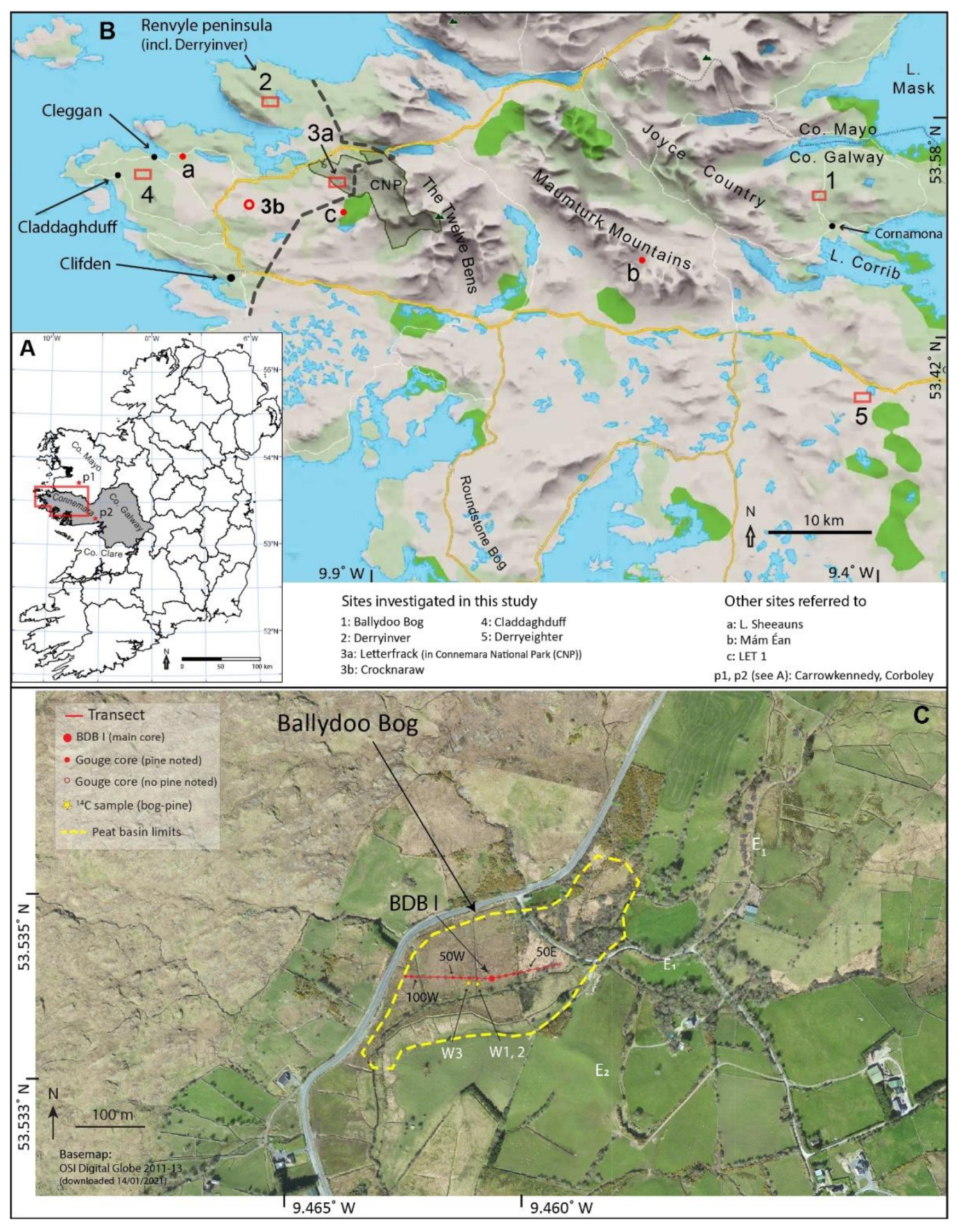
2. Study Sites
2.1. Ballydoo Bog, North-East Connemara
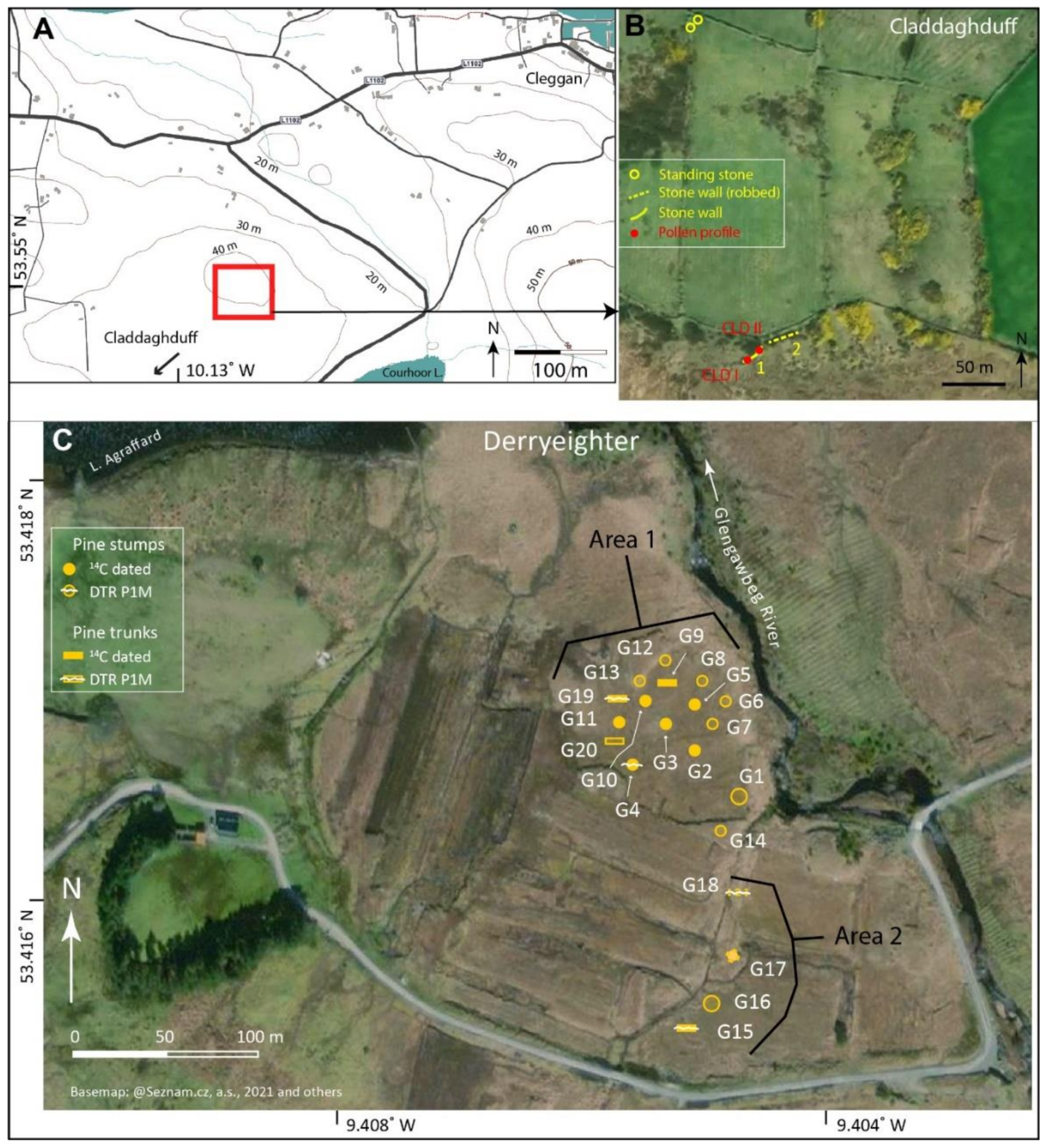
2.2. Derryinver, Letterfrack, Crocknaraw and Claddaghduff, North-West Connemara
2.3. Derryeighter, Eastern Connemara
3. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site (Name, Location, Altitude, etc.) ¶ | Publications, Theses, etc. * |
|---|---|
| Ballydoo Bog, north of Cornamona, north-east Connemara; basin size: 350 × 150 m; 4.5 ha (pollen) | |
| BDB I (peat): ps: 88 + 4 sps; 14C: 12 (4); span: 11.7–0.23 ka 53.53383, −9.46091; 32 m asl; Figure 1C | * [11,30], * [31]; presented in detail |
| Derryinver hill, Renvyle, north-west Connemara (stone alignment; pre-bog stone walls; henge-like enclosure) (pollen) | |
| DYR I: mineral soil beneath stone wall. ps: 7; 14C: 1; top: 2.43 ka 53.58369, −9.98697; 74 m asl; Figure 2A | * [24]; [10] |
| DYR II: mineral soil, overlying bk-peat. ps: 14; 14C: 3; span: 2.2–0.11 ka 53.58363, −9.9869; 73.6 m asl; Figure 2A | * [24]; [10] |
| DYR III: mineral soil beneath stone wall. ps: 6; 14C: 1; top: 1.47 ka 53.5857, −9.9865; 66 m asl; Figure 2A | * [24]; [10] |
| DYR VI: peat-filled ditch in enclosure; ps: 26; 14C: 2; span: 3.2–0.09 ka 53.58471, −9.98664; 78 m asl; Figure 2A | * [24]; [10] |
| Derryinver basin bog, Renvyle (extensive blanket bog with pine timbers; cores from marginal location near hill) (pollen) | |
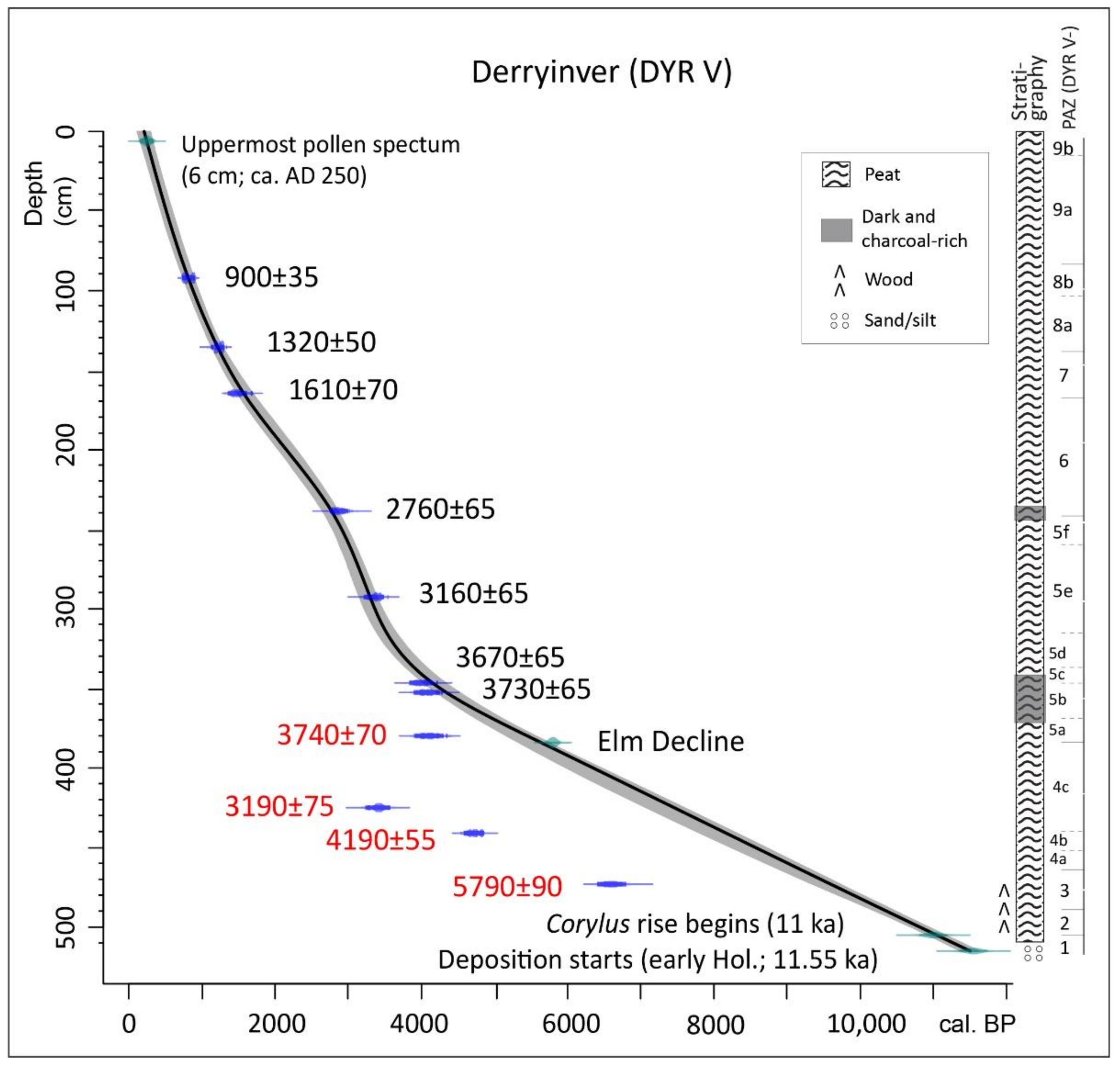
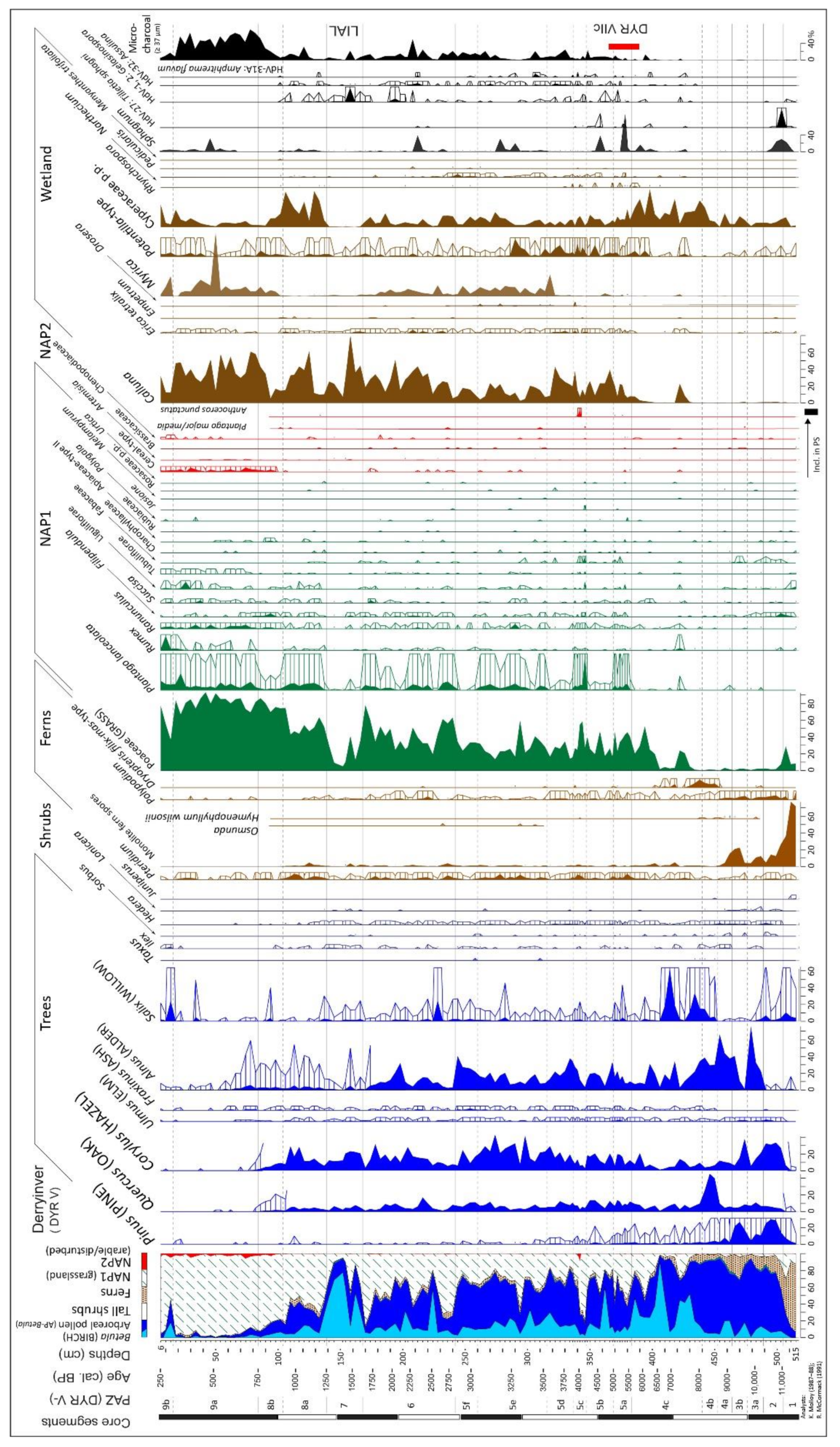
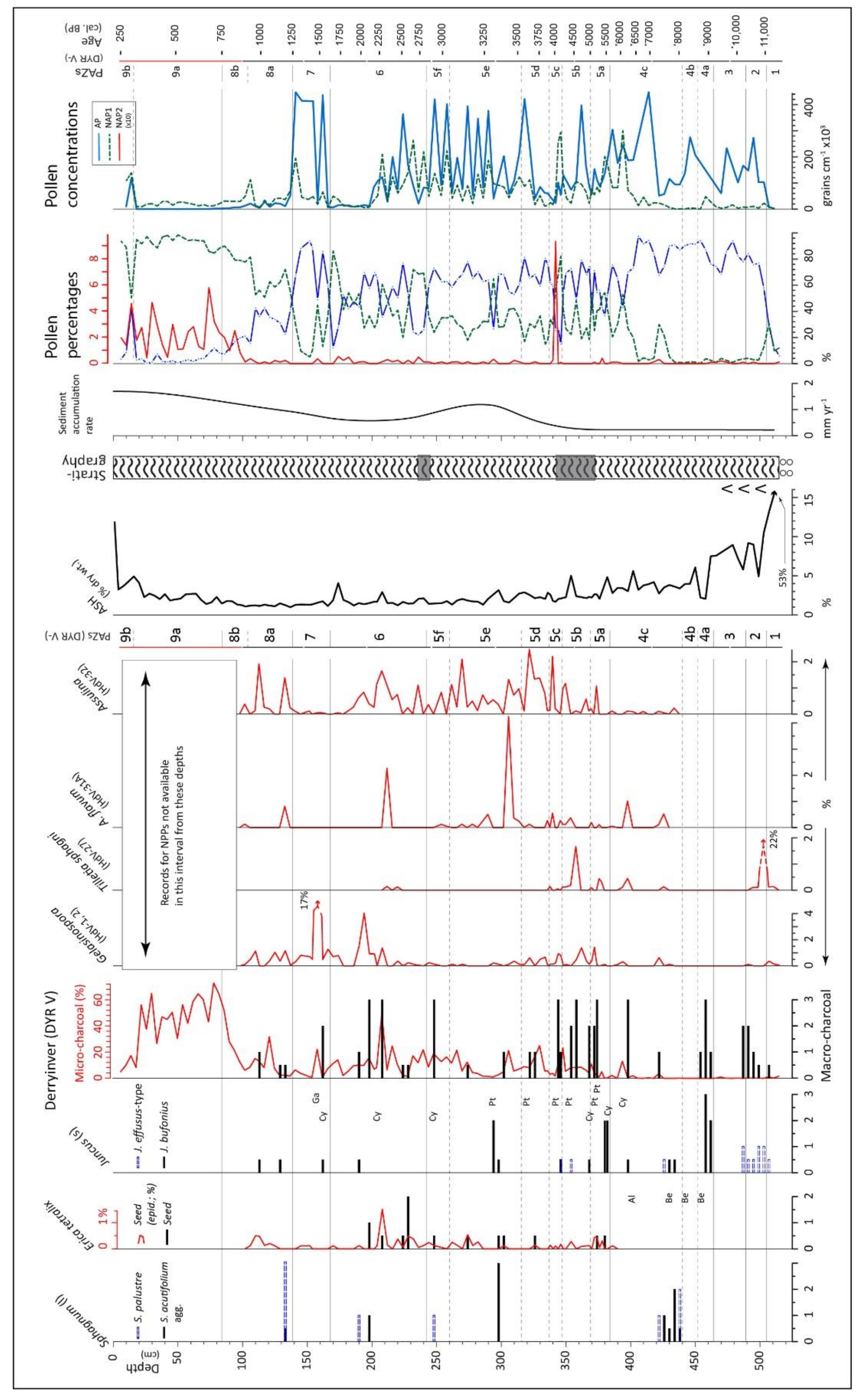
| DYR V: peat profile beside hill; ps: 136; 14C: 11 (4); span: 11.5–0.25 ka 53.58407, −9.97903; 30 m asl; Figure 2A | * [24,25]; [9]; presented in detail |
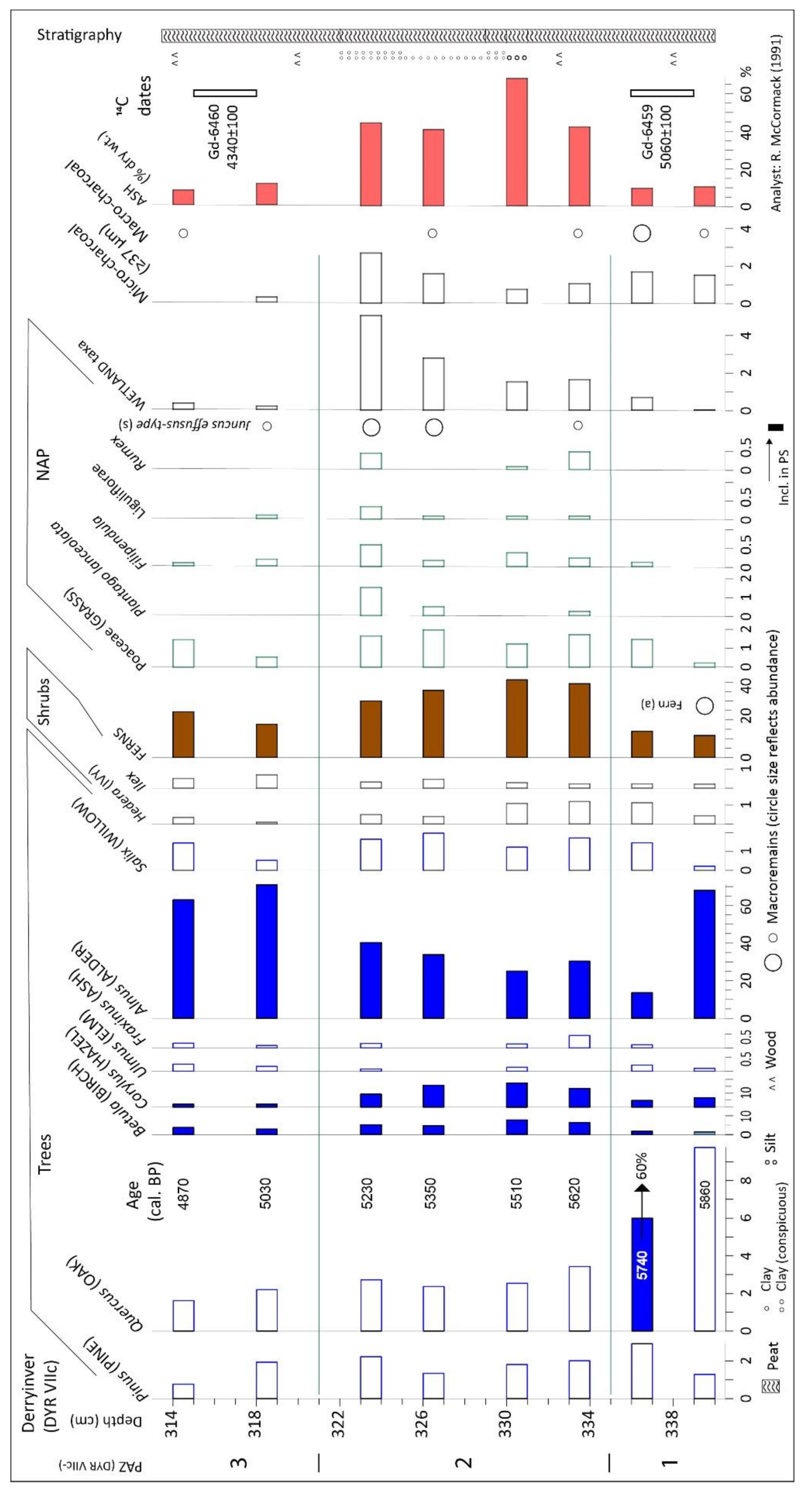
| DYR VIIc: 15 m from DYR V, nearer hill; ps: 8; 14C: 2; span: 5.86–4.87 ka 53.58419, −9.97908; 30 m asl; Figure 2A | * [25]; presented in detail |
| Letterfrack, Connemara National Park, north-west Connemara (pollen and pine dendrochronology) | |
| FRK II: basin peat; ps: 90; 14C: 11; span: 10.3–0.22 ka 53.54375, −9.93593; 97 m asl; Figure 2B | * [26]; [8] |
| FRK III: blanket peat, mineral soil beneath; ps: 8; 14C: 2; span: 3.7–2.35 ka 53.54477, −9.9395; 91 m asl; Figure 2B | [27] |
| FRK IV: blanket peat, mineral soil beneath; ps: 32; 14C: 4; span: 4.36–0.1 ka 53.54556, −9.94644; 70 m asl; Figure 2B | [27] (in part) |
| Dendrochronology: pines from CNP, Letterfrack, and from Carrowkennedy (Co. Mayo) and Corboley (Barna, S.E. Connemara) (at VC, CNP); Figure 1A,B and Figure 2B | * [11]; presented in detail |
| Crocknaraw (hilltop site) and Claddaghduff, north-west Connemara (stone walls at bog edge) (pollen) | |
| CNR I: blanket peat, underlying mineral soil; ps: 10; 14C: 3; span: 6.3–3.26 ka 53.53502, −10.02508; 105 m asl; Figure 1B | * [11] |
| CLD I: blanket peat and underlying mineral soil, beneath stone wall; ps: 17; 14C: 2; span: 4.9–2.9 ka 53.54895, −10.12698; 38 m asl; Figure 3A,B | * [24] |
| Derryeighter, eastern Connemara (14C dating and pine dendrochronology;Figure 3C) | |
| Derryeighter, Area 1 (centred on 53.4166853, −9.4052939; 42 m asl) Derryeighter, Area 2 (centred on 53.4153475, −9.4050792; 42 m asl) | * [11]; presented in detail |
4. Results
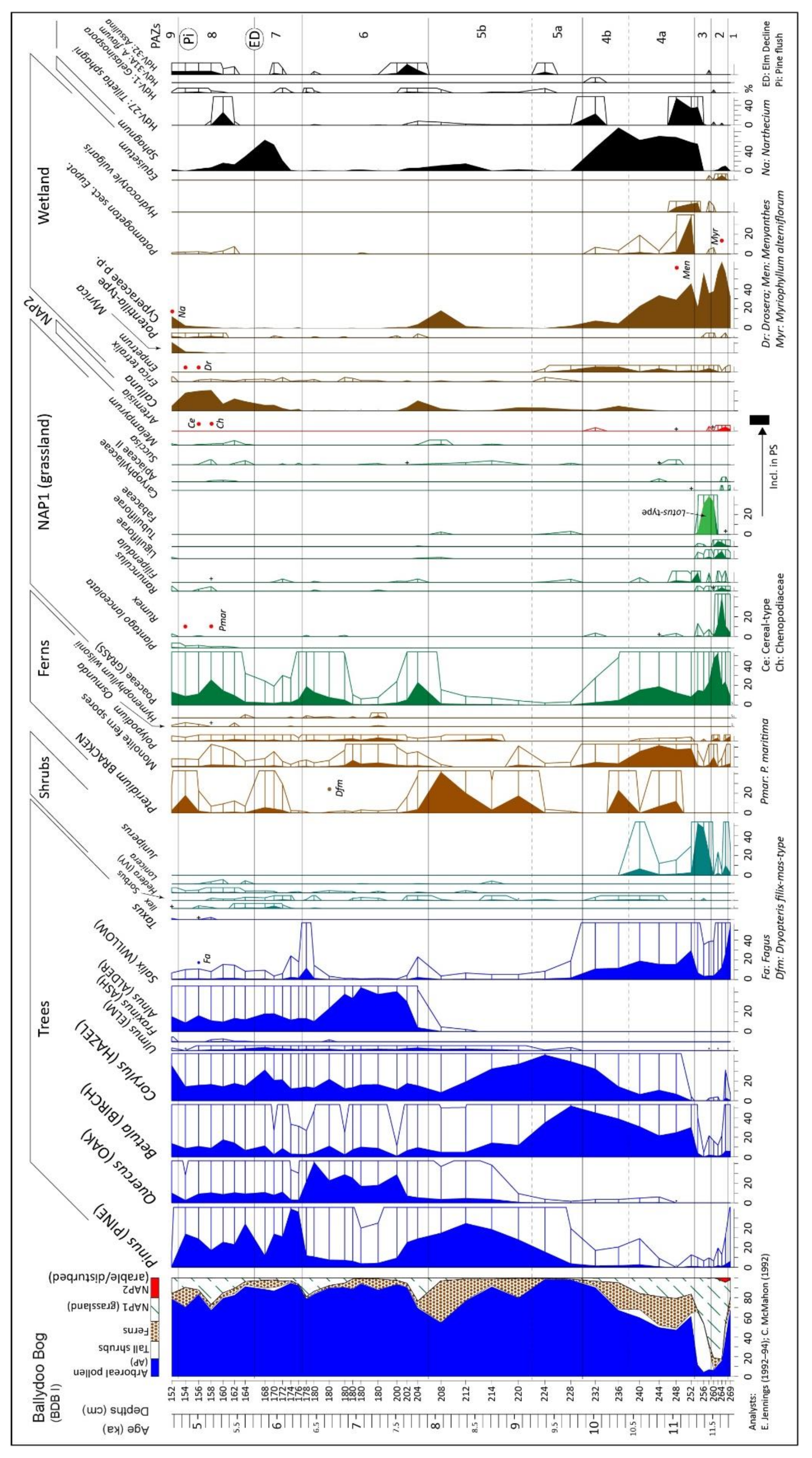
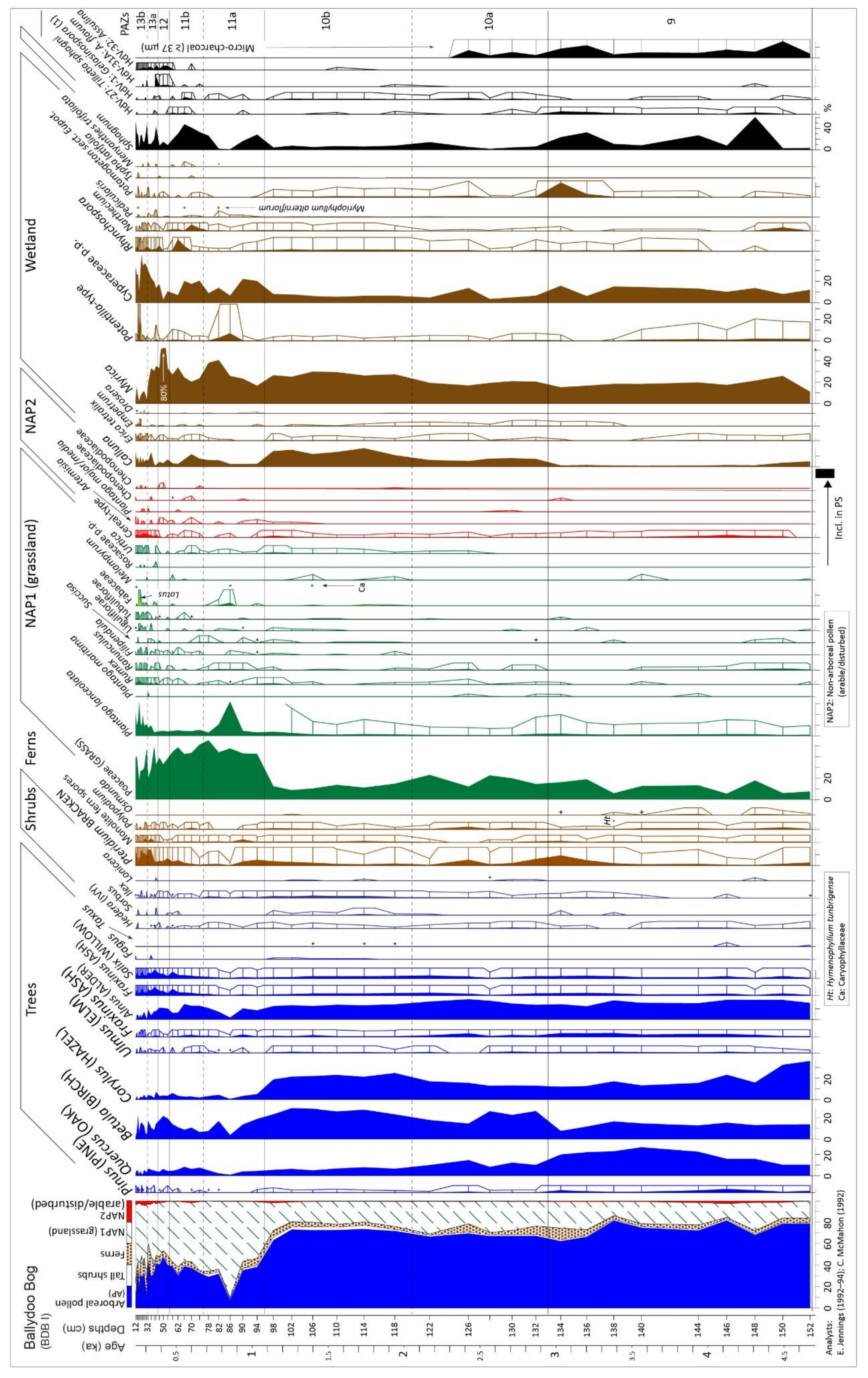
4.1. Ballydoo Bog, Cornamona
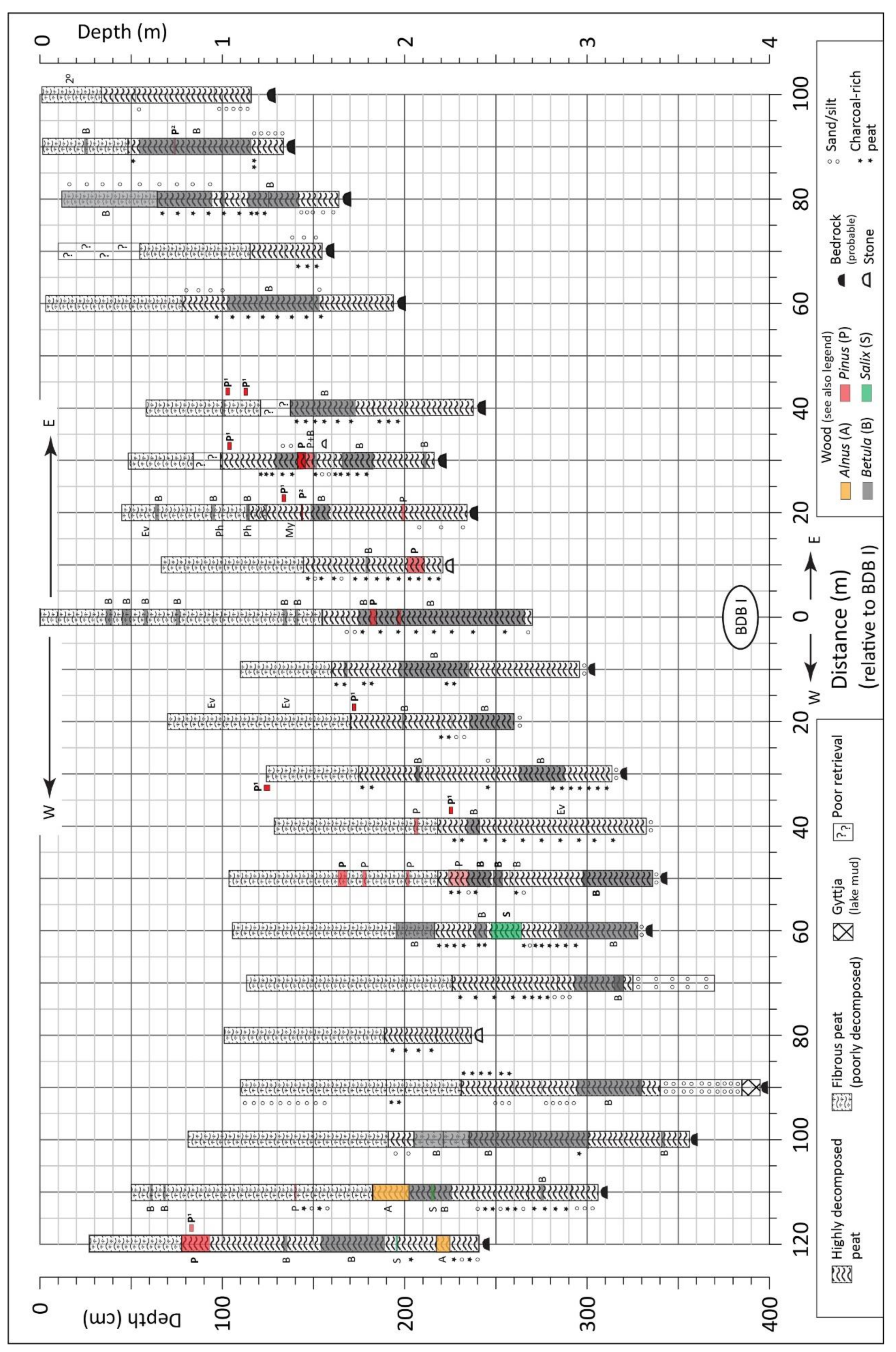
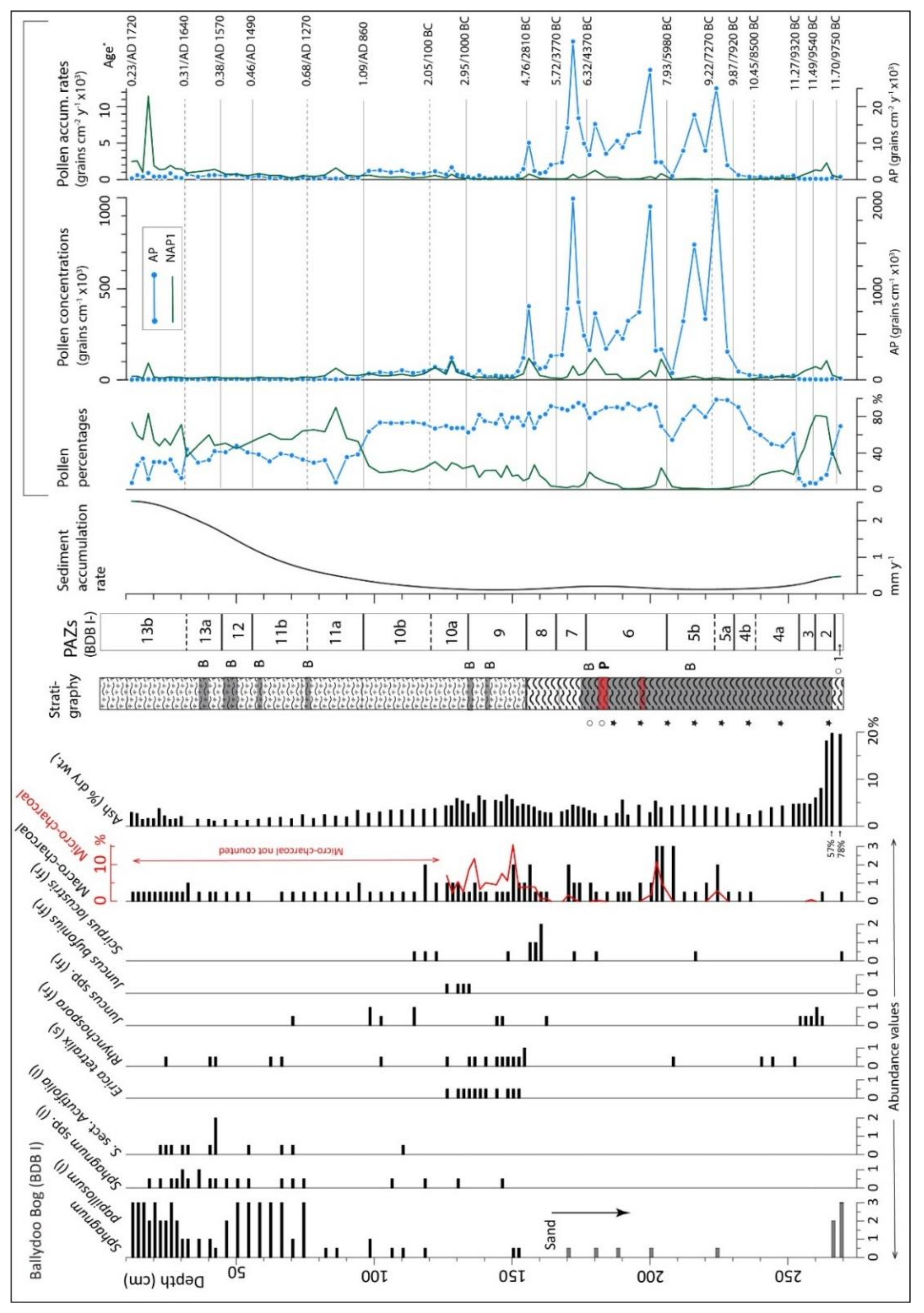
4.1.1. Peat Stratigraphy (in Basin and Core BDB I)
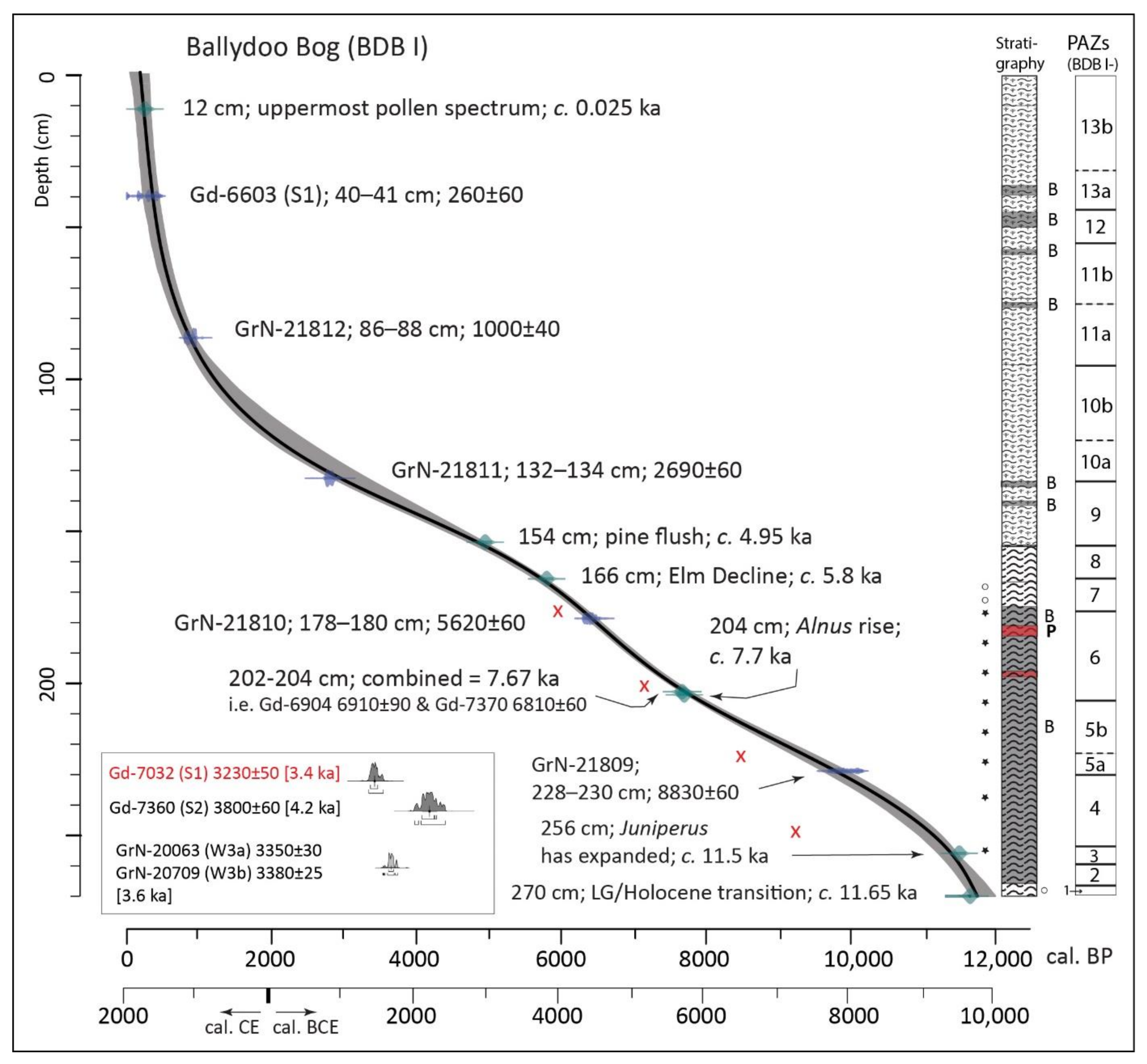
4.1.2. Radiocarbon Dating (Ballydoo)
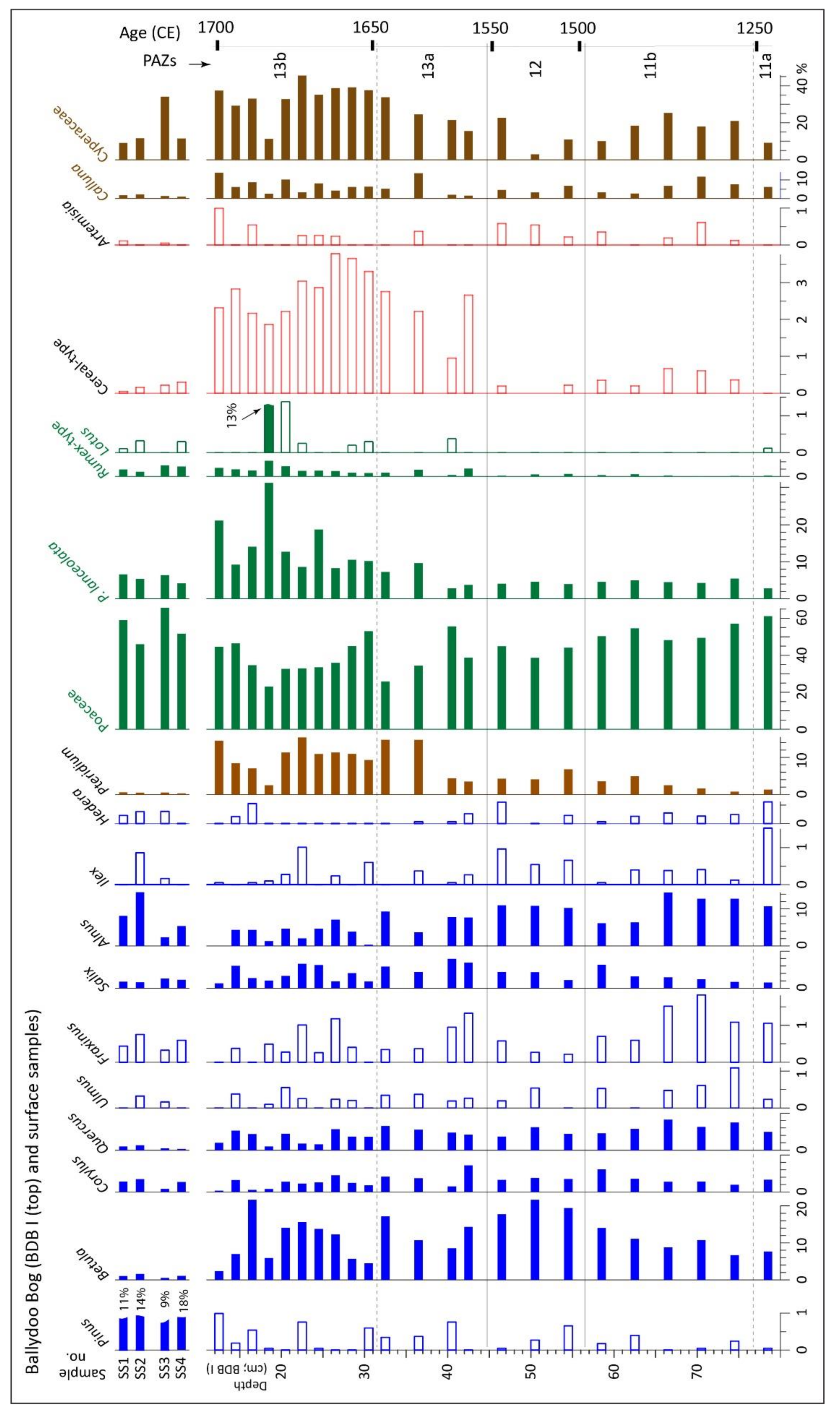
4.1.3. Pollen and Related Data, Core BDB I
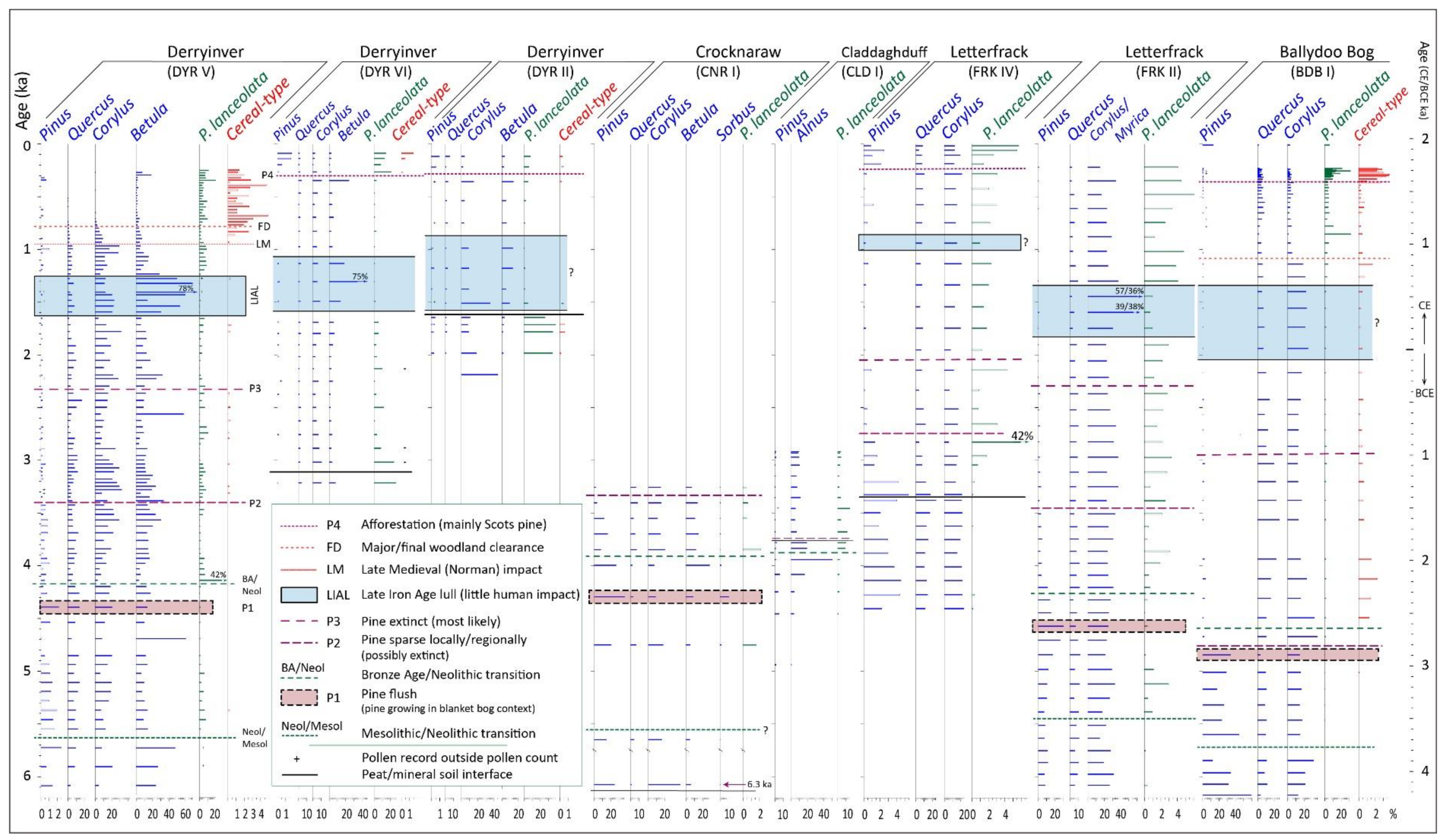
4.2. Derryinver, Renvyle
4.3. Other Investigations in North-West Connemara
4.3.1. Crocknaraw (Pollen Profile)
4.3.2. Claddaghduff (Pollen Profiles)
4.3.3. Letterfrack, Connemara National Park (Pine Dendrochronology and Pollen Profiles)
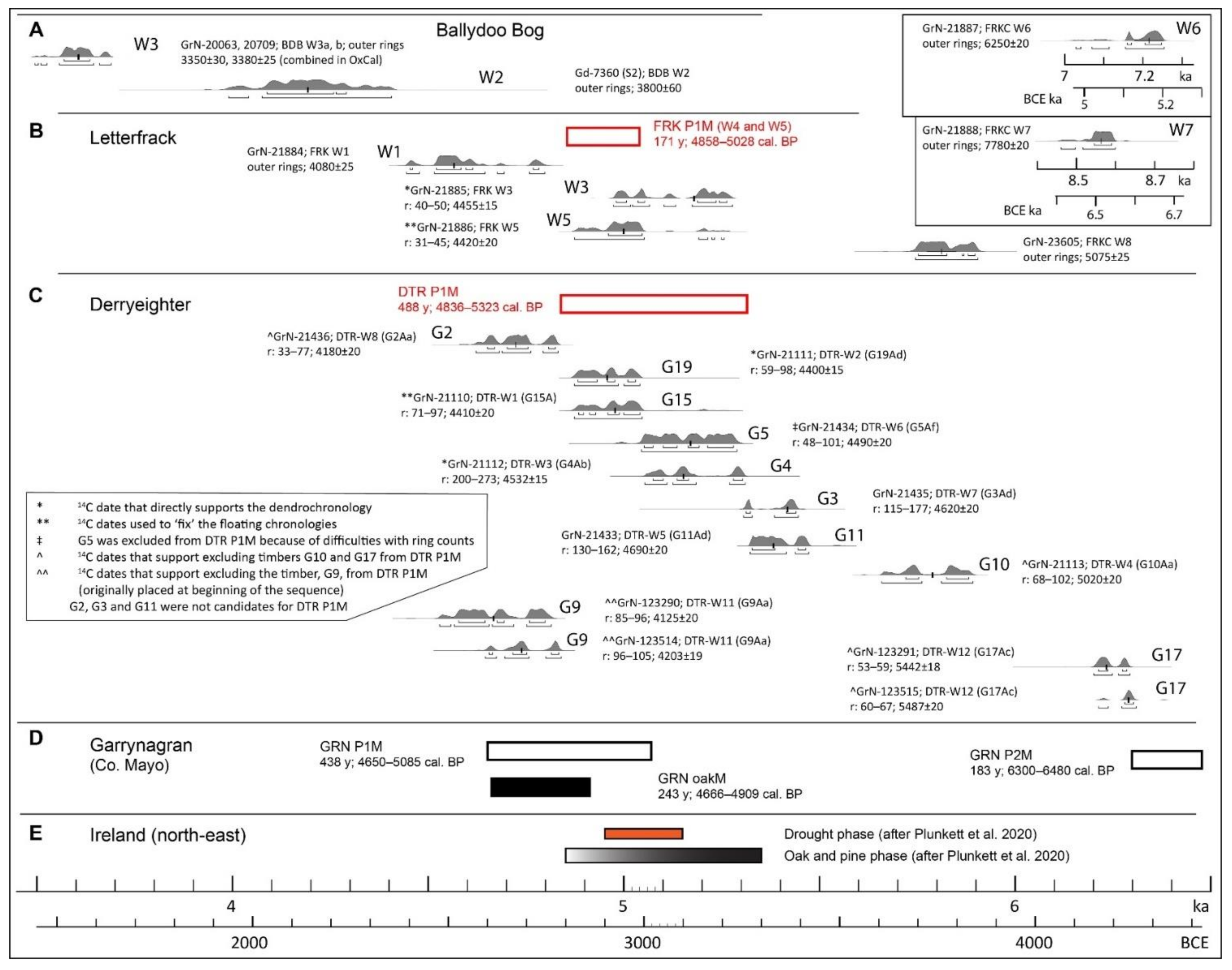
FRK W1, i.e., a stump on ~40 cm of peat located on elevated ground to the south of the Visitor Centre (VC) where shallow blanket bog has been largely cut away. Timber from the outer rings yielded the date 4080 ± 15 BP, which suggests that the tree died at c. 4.6 ka;
FRK W5, i.e., the large stump close to core FRK II. Rings 31–45 yielded the date 4420 ± 20 BP, which indicates that the pine tree began life before/at c. 5 ka. It died some 180 y later;
FRK W3, i.e., the largest stump and longest-lived tree (>180 y) in the basin and ~80 m distant from FRK II and FRK W5, yielded the date 4455 ± 15 BP (rings 40–50). This suggests that the tree probably began life before 5 ka and hence was contemporaneous with FRK W5 (the prefixes FRK and FRKC are generally omitted from the timber samples that derive from Letterfrack from here on).
4.4. Results of Investigations of Pine Timbers at Derryeighter, Eastern Connemara
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leake, B.E.; Tanner, P.W.G.; Senior, A. The Geology of Connemara [1: 63,360 Geological Map; 1″ = 1 Mile]; University of Glasgow: Glasgow, UK, 1981. [Google Scholar]
- Morris, J.H.; Long, C.B.; McConnell, B.; Archer, J.B. Geology of Connemara; Geological Survey of Ireland: Dublin, Ireland, 1995. [Google Scholar]
- Webb, D.A.; Scannell, M.J.P. Flora of Connemara and the Burren; Royal Dublin Society and Cambridge University Press: Dublin, Ireland; Cambridge, UK, 1983. [Google Scholar]
- Gibbons, M.; Higgins, J. Connemara’s emerging prehistory. Archaeol. Irel. 1988, 2, 63–66. [Google Scholar]
- Gosling, P. Archaeological Inventory of County Galway. Volume 1: West Galway; Government Stationery Office: Dublin, Ireland, 1993. [Google Scholar]
- Robinson, T. Twilight on old stones. Field Day Rev. 2007, 3, 43–51. [Google Scholar]
- Molloy, K.; O’Connell, M. The nature of the vegetational changes at about 5000 B.P. with particular reference to the elm decline: Fresh evidence from Connemara, western Ireland. New Phytol. 1987, 106, 203–220. [Google Scholar] [CrossRef]
- O’Connell, M.; Molloy, K.; Bowler, M. Post-glacial landscape evolution in Connemara, western Ireland with particular reference to woodland history. In The Cultural Landscape—Past, Present and Future; Birks, H.H., Birks, H.J.B., Kaland, P.E., Moe, D., Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 487–514. [Google Scholar]
- Molloy, K.; O’Connell, M. Palaeoecological investigations towards the reconstruction of woodland and land-use history at Lough Sheeauns, Connemara, western Ireland. Rev. Palaeobot. Palynol. 1991, 67, 75–113. [Google Scholar] [CrossRef]
- Molloy, K.; O’Connell, M. Early land use and vegetation history at Derryinver Hill, Renvyle Peninsula, Co. Galway, Ireland. In Climate Change and Human Impact on the Landscape; Chambers, F.M., Ed.; Chapman and Hall: London, UK, 1993; pp. 185–199. [Google Scholar]
- Jennings, E. Palaeoecological Studies towards the Reconstruction of the Holocene History of Pinus sylvestris L. in Western Ireland. Ph.D. Thesis, National University of Ireland Galway (NUIG), Galway, Ireland, 1997, unpublished. [Google Scholar]
- Huang, C.C.; O’Connell, M. Recent land-use and soil-erosion history within a small catchment in Connemara, western Ireland: Evidence from lake sediments and documentary sources. Catena 2000, 41, 293–335. [Google Scholar] [CrossRef]
- Huang, C.C.; O’Connell, M. Erratum to “Recent land-use and soil-erosion history within a small catchment in Connemara, western Ireland: Evidence from lake sediments and documentary sources”. Catena 2001, 43, 349–350. [Google Scholar] [CrossRef]
- Cunningham, J.D.; MacNeill, G.; Pollard, D. Chernobyl Its Effect on Ireland; Nuclear Energy Board: Dublin, Ireland, 1987; Available online: https://inis.iaea.org/search/search.aspx?orig_q=RN:19058507 (accessed on 24 September 2021).
- Fairlie, I. Dispersal, deposition and collective doses after the Chernobyl disaster. Med. Confl. Surviv. 2007, 23, 10–30. [Google Scholar] [CrossRef]
- O’Connell, M.; Molloy, K. Aran Islands, western Ireland: Farming history and environmental change reconstructed from field surveys, historical sources, and pollen analyses. J. N. Atl. 2019, 38, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Leake, B.E. A new map and interpretation of the geology of part of Joyces Country, Counties Galway and Mayo. Ir. J. Earth Sci. 2014, 32, 1–21. [Google Scholar] [CrossRef]
- Atherton, I.; Bosanquet, S.D.S.; Lawley, M. (Eds.) Mosses and Liverworts of Britain and Ireland. A Field Guide; British Bryological Society: Plymouth, UK, 2010. [Google Scholar]
- Parnell, J.; Curtis, T. Webb’s an Irish Flora, 8th ed.; Cork University Press: Cork, Ireland, 2012. [Google Scholar]
- Legg, R.; Ludlow, F.; Travis, C. Mapping the Irish rath (ringfort): Landscape and settlement patterns in the early Medieval period. In Historical Geography, GIScience and Textual Analysis: Landscapes of Time and Place; Travis, C., Ludlow, F., Gyuris, F., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 197–210. [Google Scholar]
- O’Sullivan, A. Crannogs. Lake-Dwellings of Early Ireland; Country House: Dublin, Ireland, 2000. [Google Scholar]
- Fredengren, C. Crannogs. A Study of People’s Interaction with Lakes, with Particular Reference to Lough Gara in the North-West of Ireland; Wordwell: Bray, Ireland, 2002. [Google Scholar]
- Robinson, T. Connemara. Part 1: Introduction and Gazetteer. Part 2: A One-Inch Map; Folding Landscapes: Roundstone, Ireland, 1990. [Google Scholar]
- Molloy, K. Palaeoecological Investigations towards the Reconstruction of Prehistoric Human Impact in N.W. Connemara, Western Ireland. Ph.D. Thesis, University College Galway [UCG; now NUIG], Galway, Ireland, 1989, unpublished. [Google Scholar]
- McCormack, R.F. Further Palaeoecological Investigations towards the Reconstruction of Vegetation and Land-Use History at Derryinver, Rinvyle, N.W. Connemara. Bachelor’s Thesis, Department of Botany, UCG [NUIG], Galway, Ireland, 1991, unpublished. [Google Scholar]
- Bowler, M. A Vegetation History of Connemara National Park, Co. Galway. Master’s Thesis, Department of Botany, UCG [NUIG], Galway, Ireland, 1986, unpublished. [Google Scholar]
- O’Connell, M. Origins of Irish lowland blanket bog. In Ecology and Conservation of Irish Peatlands; Doyle, G.J., Ed.; Royal Irish Academy: Dublin, Ireland, 1990; pp. 49–71. [Google Scholar]
- Hawkes, A. The beginnings and evolution of the fulacht fia tradition in early prehistoric Ireland. Proc. R. Ir. Acad. 2014, 114C, 89–139. [Google Scholar] [CrossRef]
- Lundholm, A.; Corrigan, E.; Black, K.; Nieuwenhuis, M. Ecosystem services provision from alternative management options was modified to Ireland’s western peatland forests under future development scenarios. Ir. For. 2020, 77, 49–73. [Google Scholar]
- Jennings, E.M. Reconstruction of Vegetation and Land Use in N.E. Connemara during the Early and Mid Holocene Based on a Peat Profile from North of Ballydoo Lough, Cornamona, Co. Galway. Bachelor’s Thesis, Department of Botany, UCG [NUIG], Galway, Ireland, 1992, unpublished. [Google Scholar]
- McMahon, C. Reconstruction of Vegetation and Land Use in N.E. Connemara during the Later Holocene Based on a Peat Profile from North of Ballydoo Lough, Cornamona, Co. Galway. Bachelor’s Thesis, Department of Botany, UCG [NUIG], Galway, Ireland, 1992, unpublished. [Google Scholar]
- Beug, H.-J. Leitfaden der Pollenbestimmung für Mitteleuropa und Angrenzende Gebiete. Lieferung 1; Gustav Fischer: Stuttgart, Germany, 1961. [Google Scholar]
- Fægri, K.; Iversen, J. Textbook of Pollen Analysis, 3rd ed.; Munksgaard: Copenhagen, Denmark, 1975. [Google Scholar]
- Fægri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Fægri, K., Kaland, P.E., Kryzwinski, K., Eds.; Wiley: Chichester, UK, 1989. [Google Scholar]
- Moore, P.D.; Webb, J.A. An Illustrated Guide to Pollen Analysis, 1st ed.; Hodder and Stoughton: London, UK, 1978; p. 133. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. An Illustrated Guide to Pollen Analysis, 2nd ed.; Blackwell Scientific: London, UK, 1991. [Google Scholar]
- Punt, W.; Blackmore, s.; Hoen, P.P.; Stafford, P.J. The Northwest European Pollen Flora, Volumes I–VIII; Elsevier: Amsterdam, The Netherlands, 1976–2003. [Google Scholar]
- Mohr, R. Untersuchungen zur nacheiszeitlichen Vegetations- und Moorentwicklung im nordwestlichen Niedersachsen mit besonderer Berücksichtigung von Myrica gale L. VAG Vechtaer Arb. Geogr. Regionalwiss. 1990, 12, 1–144. [Google Scholar]
- Birks, H.J.B. Past and Present Vegetation of the Isle of Skye. A Palaeoecological Study; Cambridge University Press: Cambridge, UK, 1973. [Google Scholar]
- Beug, H.-J. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebiete, 2nd ed.; Friedrich Pfeil: München, Germany, 2015. [Google Scholar]
- Van Geel, B. A palaeoecological study of Holocene peat bog sections in Germany and the Netherlands. Rev. Palaeobot. Palynol. 1978, 25, 1–120. [Google Scholar] [CrossRef]
- Feeser, I. COUNTPOL. A Computer Program for Processing and Plotting Pollen Data; NUIG: Galway, Ireland; CAU: Kiel, Germany, 2007; unpublished. [Google Scholar]
- Beijerinck, W. Zadenatlas der Nederlandische Flora. Ten Behoeve van de Botanie, Palaeontologie, Bodemcultuur en Warenkennis; Backhuys and Meesters: Amsterdam, The Netherlands, 1947. [Google Scholar]
- Körber-Grohne, U. Bestimmungsschlüssel für subfossile Juncus-Samen und Gramineen-Früchte. Prob. Küstenforsch 1964, 7, 1–47. [Google Scholar]
- Greguss, P. Holzanatomie der Europäischen Laubhölzer und Sträucher, 1st ed.; Akadémiai Kiadó: Budapest, Hungary, 1959. [Google Scholar]
- Grosser, D. Die Hölzer Mitteleuropas; Springer: Berlin/Heidelberg, Germany, 1977. [Google Scholar]
- Schweingruber, F.H. Anatomie europäischer Hölzer. Anatomy of European Woods; Haupt: Bern, Switzerland, 1990. [Google Scholar]
- Blaauw, M. Methods and code for ‘classical’ age-modelling of radiocarbon sequences. Quat. Geochronol. 2010, 5, 512–518. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. Deposition models for chronological records. Quat. Sci. Rev. 2008, 27, 42–60. [Google Scholar] [CrossRef] [Green Version]
- Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef] [Green Version]
- Reimer, P.J.; Austin, W.E.N.; Bard, E.; Bayliss, A.; Blackwell, P.G.; Bronk Ramsey, C.; Butzin, M.; Cheng, H.; Edwards, R.L.; Friedrich, M.; et al. The IntCal20 Northern Hemisphere radiocarbon age calibration curve (0–55 cal kBP). Radiocarbon 2020, 62, 725–757. [Google Scholar] [CrossRef]
- Baillie, M.G.L.; Pilcher, J.R. A simple crossdating program for tree-ring research. Tree-Ring Bull. 1973, 33, 7–14. [Google Scholar]
- Munro, M.A. An improved algorithm for crossdating tree-ring series. Tree-Ring Bull. 1984, 44, 17–27. [Google Scholar]
- Graybill, D.A. Revised computer programs for tree-ring research. Tree-Ring Bull. 1979, 39, 77–82. [Google Scholar]
- Plunkett, G.; Brown, D.M.; Swindles, G.T. Siccitas magna ultra modum: Examining the occurrence and societal impact of droughts in prehistoric Ireland. Proc. R. Ir. Acad. 2020, 120C, 23–50. [Google Scholar] [CrossRef] [Green Version]
- Lowe, J.J.; Rasmussen, S.O.; Björck, S.; Hoek, W.Z.; Steffensen, J.P.; Walker, M.J.C.; Yu, Z.C. Synchronisation of palaeoenvironmental events in the North Atlantic region during the Last Termination: A revised protocol recommended by the INTIMATE group. Quat. Sci. Rev. 2008, 27, 6–17. [Google Scholar] [CrossRef]
- Walker, M.; Head, M.J.; Lowe, J.; Berkelhammer, M.; Björck, S.; Cheng, H.; Cwynar, L.C.; Fisher, D.; Gkinis, V.; Long, A.; et al. Subdividing the Holocene Series/Epoch: Formalization of stages/ages and subseries/subepochs, and designation of GSSPs and auxiliary stratotypes. J. Quat. Sci. 2019, 34, 173–186. [Google Scholar] [CrossRef]
- Molloy, K.; O’Connell, M. Post-glaciation plant colonisation of Ireland: Fresh insights from An Loch Mór, Inis Oírr, western Ireland. In Mind the Gap II: New Insights into the Irish Postglacial; Sleeman, D.P., Carlsson, J., Carlsson, J.E.L., Eds.; Irish Naturalists’ Journal: Belfast, Northern Ireland, 2014; pp. 66–88. [Google Scholar]
- O’Connell, M.; Molloy, K.; Jennings, E. Long-term human impact and environmental change in mid-western Ireland, with particular reference to Céide Fields—An overview. E G Quat. Sci. J. 2020, 70, 1–32. [Google Scholar] [CrossRef]
- O’Connell, M. Post-glacial vegetation and landscape change in upland Ireland with particular reference to Mám Éan, Connemara. Rev. Palaeobot. Palynol. 2021, 290, 104377. [Google Scholar] [CrossRef]
- Gaillard, M.-J.; Sugita, S.; Bunting, J.; Dearing, J.; Bittmann, F. Human impact on terrestrial ecosystems, pollen calibration and quantitative reconstruction of past land-cover. Veget. Hist. Archaeobot. 2008, 17, 415–418. [Google Scholar] [CrossRef]
- O’Connell, M.; Molloy, K. Farming and woodland dynamics in Ireland during the Neolithic. Biol. Environ. (Proc. R. Ir. Acad. Ser. B) 2001, 101, 99–128. [Google Scholar]
- Barber, K.E.; Chambers, F.M.; Maddy, D. Holocene palaeoclimates from peat stratigraphy: Macrofossil proxy climate records from three oceanic raised bogs in England and Ireland. Quat. Sci. Rev. 2003, 22, 521–539. [Google Scholar] [CrossRef]
- Swindles, G.T.; Lawson, I.T.; Matthews, I.P.; Blaauw, M.; Daley, T.J.; Charman, D.J.; Roland, T.P.; Plunkett, G.; Schettler, G.; Gearey, B.R.; et al. Centennial-scale climate change in Ireland during the Holocene. Earth-Sci. Rev. 2013, 126, 300–320. [Google Scholar] [CrossRef] [Green Version]
- McClatchie, M.; Potito, A. Tracing environmental, climatic and social change in Neolithic Ireland. Proc. R. Ir. Acad. 2020, 120C, 23–50. [Google Scholar] [CrossRef]
- Leuschner, H.H.; Bauerochse, A.; Metzler, A. Environmental change, bog history and human impact around 2900 B.C. in NW Germany—preliminary results from a dendroecological study of a sub-fossil pine woodland at Campemoor, Dümmer Basin. Veget. Hist. Archaeobot. 2007, 16, 183–195. [Google Scholar] [CrossRef]
- Tauber, H. Differential pollen dispersion and the interpretation of pollen diagrams. Dan. Geol. Unders. II Raekke 1965, 89, 1–69. [Google Scholar]
- Overland, A.; O’Connell, M. New insights into late Holocene farming and woodland dynamics in western Ireland with particular reference to the early medieval horizontal watermill at Kilbegly, Co. Roscommon. Rev. Palaeobot. Palynol. 2011, 163, 205–226. [Google Scholar] [CrossRef] [Green Version]
- Behre, K.-E. The interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 1981, 23, 225–245. [Google Scholar]
- O’Connell, M. Reconstruction of local landscape development in the post-Atlantic based on palaeoecological investigations at Carrownaglogh prehistoric field system, County Mayo, Ireland. Rev. Palaeobot. Palynol. 1986, 49, 117–176. [Google Scholar] [CrossRef]
- Hall, V.A. A study of the modern pollen rain from a reconstructed 19th century farm. Ir. Nat. J. 1989, 23, 82–92. [Google Scholar]
- Mitchell, G.F. Studies in Irish Quaternary deposits: No. 7. Proc. R. Ir. Acad. 1951, 53B, 111–206. [Google Scholar]
- Mitchell, F.J.G.; Cooney, T. Vegetation history in the Killarney valley. In Ross Island. Mining, Metal and Society in Early Ireland (Bronze Age Studies 6); O’Brien, W., Ed.; Department of Archaeology, NUIG: Galway, Ireland, 2004; pp. 481–493. [Google Scholar]
- Doyle, L. Giant Irish Deer and early alder: The significance of a fragment. Seanda 2010, 5, 3. [Google Scholar]
- Young, D.S.; Green, C.P.; Batchelor, C.R.; Austin, P.; Elias, S.A.; Athersuch, J.; Lincoln, P. Macrofossil evidence of alder (Alnus sp.) in Britain early in the Late Glacial Interstadial: Implications for the northern cryptic refugia debate. J. Quat. Sci. 2021, 36, 40–55. [Google Scholar] [CrossRef]
- O’Connell, M. Early cereal-type pollen records from Connemara, western Ireland and their possible significance. Pollen Spores 1987, 29, 207–224. [Google Scholar]
- Cooney, G.; Bayliss, A.; Healy, F.; Whittle, A.; Danaher, E.; Cayney, L.; Mallory, J.; Smyth, J.; Kador, T.; O’Sullivan, M. Ireland. In Gathering Time. Dating the Early Neolithic Enclosures of Southern Britain and Ireland; Whittle, A., Healy, F., Bayliss, A., Eds.; Oxbow Books: Oxford, UK, 2011; Volume 2, pp. 562–669. [Google Scholar]
- McClatchie, M.; Bogaard, A.; Colledge, S.; Whitehouse, N.J.; Schulting, R.J.; Barratt, P.; McLaughlin, T.R. Neolithic farming in north-western Europe: Archaeobotanical evidence from Ireland. J. Archaeol. Sci. 2014, 51, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Blackford, J.J.; Chambers, F.M. Proxy records of climate from blanket mires: Evidence for a Dark Age (1400 BP) climatic deterioration in the British Isles. Holocene 1991, 1, 63–67. [Google Scholar] [CrossRef]
- Blackford, J.J.; Chambers, F.M. Proxy climate record for the last 1000 years from Irish blanket peat and a possible link to solar variability. Earth Planet. Sci. Lett. 1991, 133, 145–150. [Google Scholar] [CrossRef]
- Chambers, F.M.; Blackford, J.J. Mid- and late-Holocene climatic changes: A test of periodicity and solar forcing in proxy-climate data from blanket peat bogs. J. Quat. Sci. 2001, 16, 329–338. [Google Scholar] [CrossRef]
- McGeever, A.H.; Mitchell, F.J.G. Pine stumps in Irish peats: Is their occurrence a valid proxy climate indicator? J. Quat. Sci. 2015, 30, 489–496. [Google Scholar] [CrossRef]
- Hannon, G.E.; Bradshaw, R.H.W. Recent vegetation dynamics on two Connemara lake islands, western Ireland. J. Biogeog. 1989, 16, 75–81. [Google Scholar] [CrossRef]
- Chambers, F.M.; Ogle, M.I.; Blackford, J.J. Palaeoenvironmental evidence for solar forcing of Holocene climate: Linkages to solar science. Prog. Phys. Geog. 1999, 23, 181–204. [Google Scholar] [CrossRef]
- O’Connell, M.; McDonnell, K. Holocene vegetation history of SW Connemara, Co. Galway with particular reference to Carna and Roundstone [Site A3.6]. In The Quaternary of Western Ireland. INQUA 2019 Field Guide Pre:GL-2; Coxon, P., Ed.; Irish Quaternary Association (IQUA): Dublin, Ireland, 2019; pp. 83–102. [Google Scholar]
- Stolze, S.; Monecke, T. Holocene history of ‘non-native’ trees in Ireland. Rev. Palaeobot. Palynol. 2017, 244, 347–355. [Google Scholar] [CrossRef]
- Campbell, B.M.S.; Ludlow, F. Climate, disease and society in late-medieval Ireland. Proc. R. Ir. Acad. 2020, 120C, 159–252. [Google Scholar] [CrossRef]
- Walsh, S. A Summary of Climate Averages for Ireland 1981–2010. Climatological Note No. 14; Met Éireann: Dublin, Ireland, 2012. [Google Scholar]
- O’Connell, M.; Molloy, K. Mid- and late-Holocene environmental change in western Ireland: New evidence from coastal peats and fossil timbers with particular reference to relative sea-level change. Holocene 2017, 27, 1825–1845. [Google Scholar] [CrossRef]
- Bennett, K.D. The post-glacial history of Pinus sylvestris in the British Isles. Quat. Sci. Rev. 1984, 3, 133–155. [Google Scholar] [CrossRef]
- Fossitt, J.A. Late-glacial and Holocene vegetation history of western Donegal, Ireland. Biol. Environ. (Proc. R. Ir. Acad.) 1994, 94B, 1–31. [Google Scholar]
- Caseldine, C.; Thompson, G.; Langdon, C.; Hendon, D. Evidence for an extreme climatic event on Achill Island, Co. Mayo, Ireland around 5200–5100 cal. yr BP. J. Quat. Sci. 2005, 20, 169–178. [Google Scholar] [CrossRef]
- McLaughlin, T.R.; Whitehouse, N.J.; Schulting, R.J.; McClatchie, M.; Barratt, P.; Bogaard, A. The changing face of Neolithic and Bronze Age Ireland: A big data approach to the settlement and burial records. J. World Prehist. 2016, 29, 117–153. [Google Scholar] [CrossRef] [Green Version]
- Ginn, V.; Plunkett, G. Filling the gaps: A chronology of Bronze Age settlement in Ireland. J. Ir. Archaeol. 2020, 29, 41–61. [Google Scholar]
- Feeser, I.; Dörfler, W.; Kneisel, J.; Hinz, M.; Dreibrodt, S. Human impact and population dynamics in the Neolithic and Bronze Age: Multi-proxy evidence from north-western Central Europe. Holocene 2019, 29, 1596–1606. [Google Scholar] [CrossRef] [Green Version]
- Lynch, A. Man and environment in south-west Ireland, 4000 B.C.–A.D. 800. BAR Br. Ser. 1981, 85, 1–175. [Google Scholar]
- Waddell, J. The Prehistoric Archaeology of Ireland, 3rd ed.; Wordwell: Bray, Ireland, 2010. [Google Scholar]
- Holmes, J.A.; Leuenberger, M.; Molloy, K.; O’Connell, M. Younger Dryas and Holocene environmental change at the Atlantic fringe of Europe derived from lake-sediment stable-isotope records from western Ireland. Boreas 2020, 49, 233–249. [Google Scholar] [CrossRef]
- Becker, K.; Ó Néill, J.; O’Flynn, L. Iron Age Ireland: Finding an Invisible People. Final Report to the Heritage Council. Project 16365. 2008 Archaeology Grant Scheme. 2008. Available online: https://www.ucd.ie/t4cms/iron_age_ireland_project_16365_pilotweb.pdf (accessed on 7 November 2021).
- Cahill Wilson, J.; Standish, C.D. Mobility and migration in late Iron Age and early Medieval Ireland. J. Archaeol. Sci. Rep. 2016, 6, 230–241. [Google Scholar] [CrossRef]
- Caulfield, S. Céide Fields and Belderrig valley: Eighty-four years of research. In The Céide Fields. Ireland; Boschiero, P., Latini, L., Caulfield, S., Eds.; Fondazione Benetton: Treviso, Italy, 2018; pp. 55–73. [Google Scholar]
- Stout, G.; Stout, M. Early landscapes: From prehistory to plantation. In Atlas of the Irish Rural Landscape, 2nd ed.; Aalen, F.H.A., Whelan, K., Stout, M., Eds.; Cork University Press: Cork, Ireland, 2011; pp. 31–65. [Google Scholar]















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
O’Connell, M.; Jennings, E.; Molloy, K. Holocene Vegetation Dynamics, Landscape Change and Human Impact in Western Ireland as Revealed by Multidisciplinary, Palaeoecological Investigations of Peat Deposits and Bog-Pine in Lowland Connemara. Geographies 2021, 1, 251-291. https://0-doi-org.brum.beds.ac.uk/10.3390/geographies1030015
O’Connell M, Jennings E, Molloy K. Holocene Vegetation Dynamics, Landscape Change and Human Impact in Western Ireland as Revealed by Multidisciplinary, Palaeoecological Investigations of Peat Deposits and Bog-Pine in Lowland Connemara. Geographies. 2021; 1(3):251-291. https://0-doi-org.brum.beds.ac.uk/10.3390/geographies1030015
Chicago/Turabian StyleO’Connell, Michael, Eneda Jennings, and Karen Molloy. 2021. "Holocene Vegetation Dynamics, Landscape Change and Human Impact in Western Ireland as Revealed by Multidisciplinary, Palaeoecological Investigations of Peat Deposits and Bog-Pine in Lowland Connemara" Geographies 1, no. 3: 251-291. https://0-doi-org.brum.beds.ac.uk/10.3390/geographies1030015