
Development of Frozen Pulps and Powders from Carrot and Tomato by-Products: Impact of Processing and Storage Time on Bioactive and Biological Properties

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Processing
2.2.1. Freezing
2.2.2. Drying
2.3. Microbiological Evaluation
2.3.1. Fresh and Frozen Pulps
2.3.2. Dried Powders
2.4. Monitorization of Pulps Viscosity
2.5. Preparation of Tomato and Carrot Extracts
2.6. Antioxidant Activity
2.6.1. 2,2′-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid (ABTS) Assay
2.6.2. Oxygen Radical Absorbance Capacity (ORAC) Assay
2.6.3. 2-diphenyl-1-picrylhydrazyl (DPPH) Assay
2.7. Total Phenolic Content (TPC)
2.8. Identification and Quantification of Phenolics by High-Performance Liquid Chromatography (HPLC)
2.9. Monitorization of Carotenoid and Vitamin E Content
2.9.1. Carotenoids and Vitamin E Extraction
2.9.2. Identification and Quantification of Carotenoids by HPLC
2.9.3. Identification and Quantification of Vitamin E by HPLC
2.10. Statistical Analysis
3. Results and Discussion
3.1. Monitorization of Microbial Counts
3.2. Monitorization of Pulps Viscosity
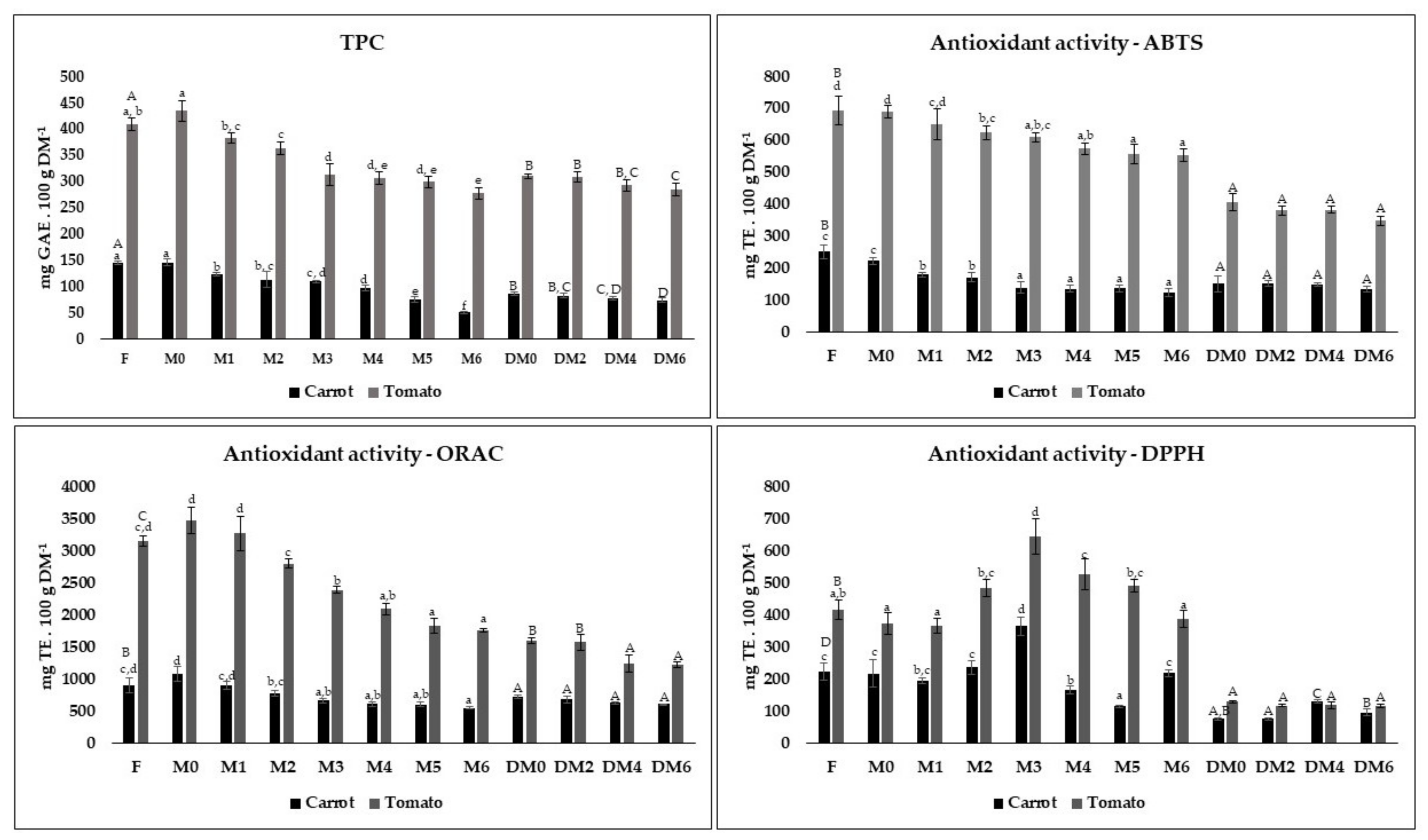
3.3. Variation of TPC and Antioxidant Activity
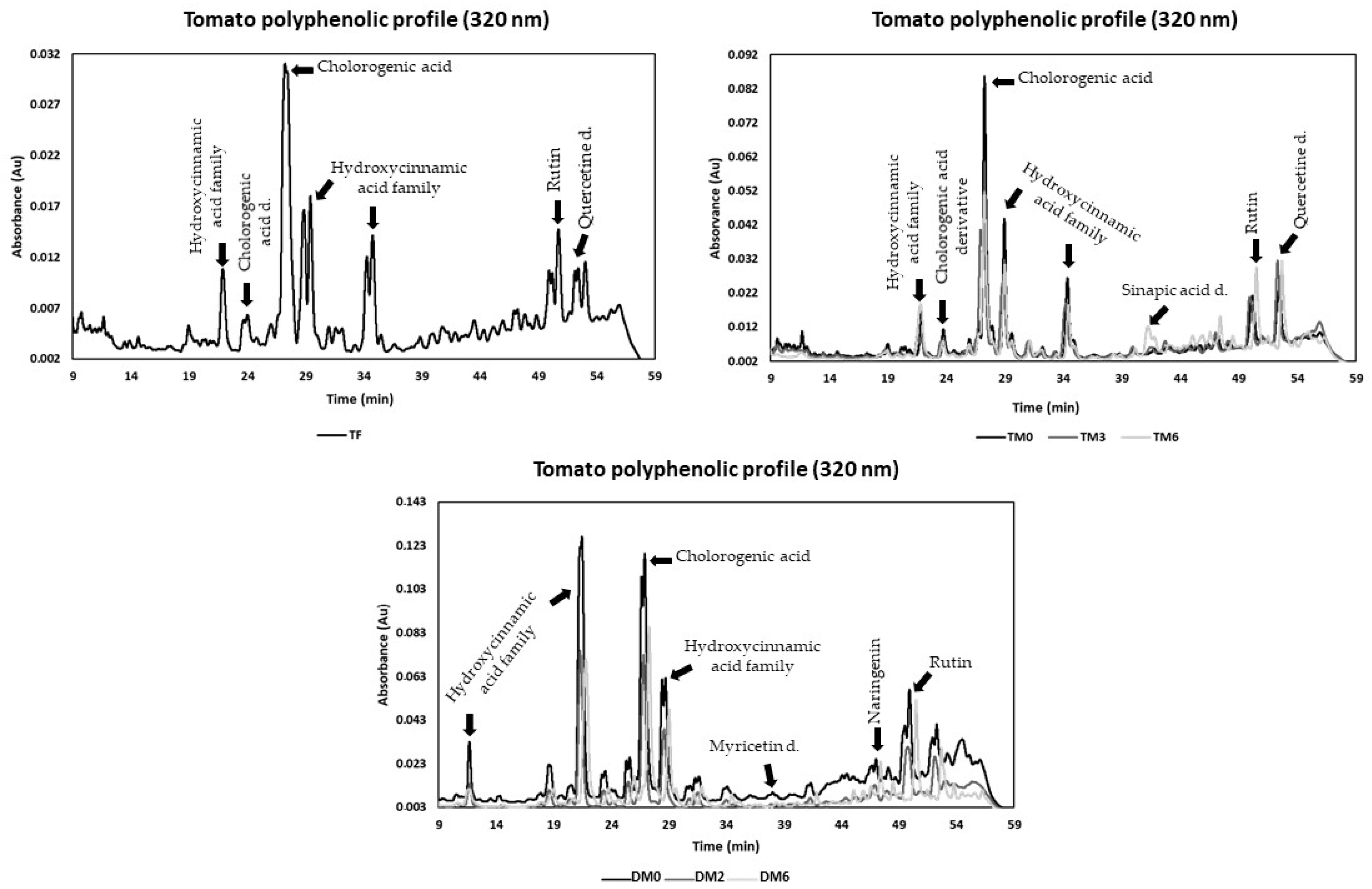
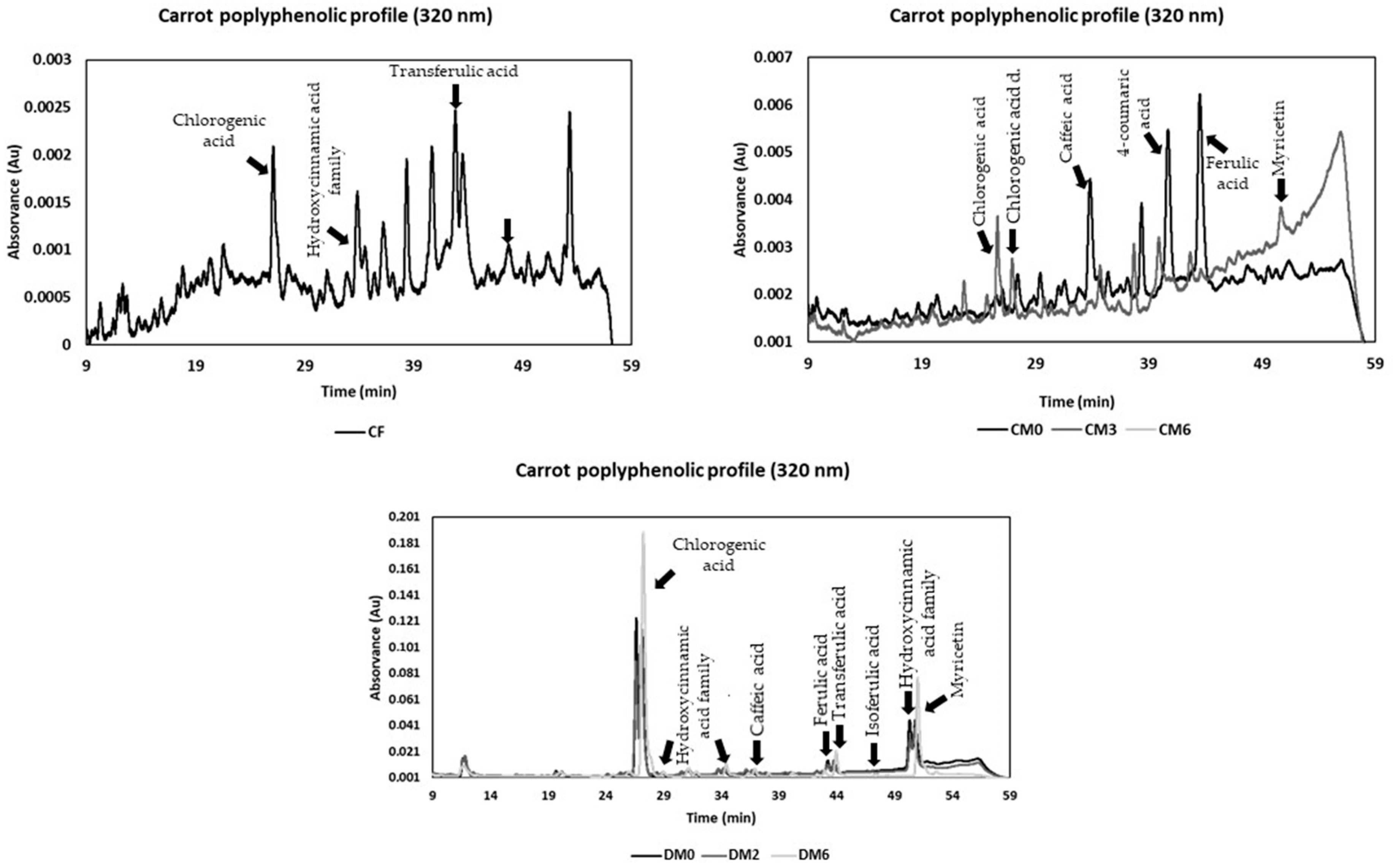
3.4. Phenolics Composition
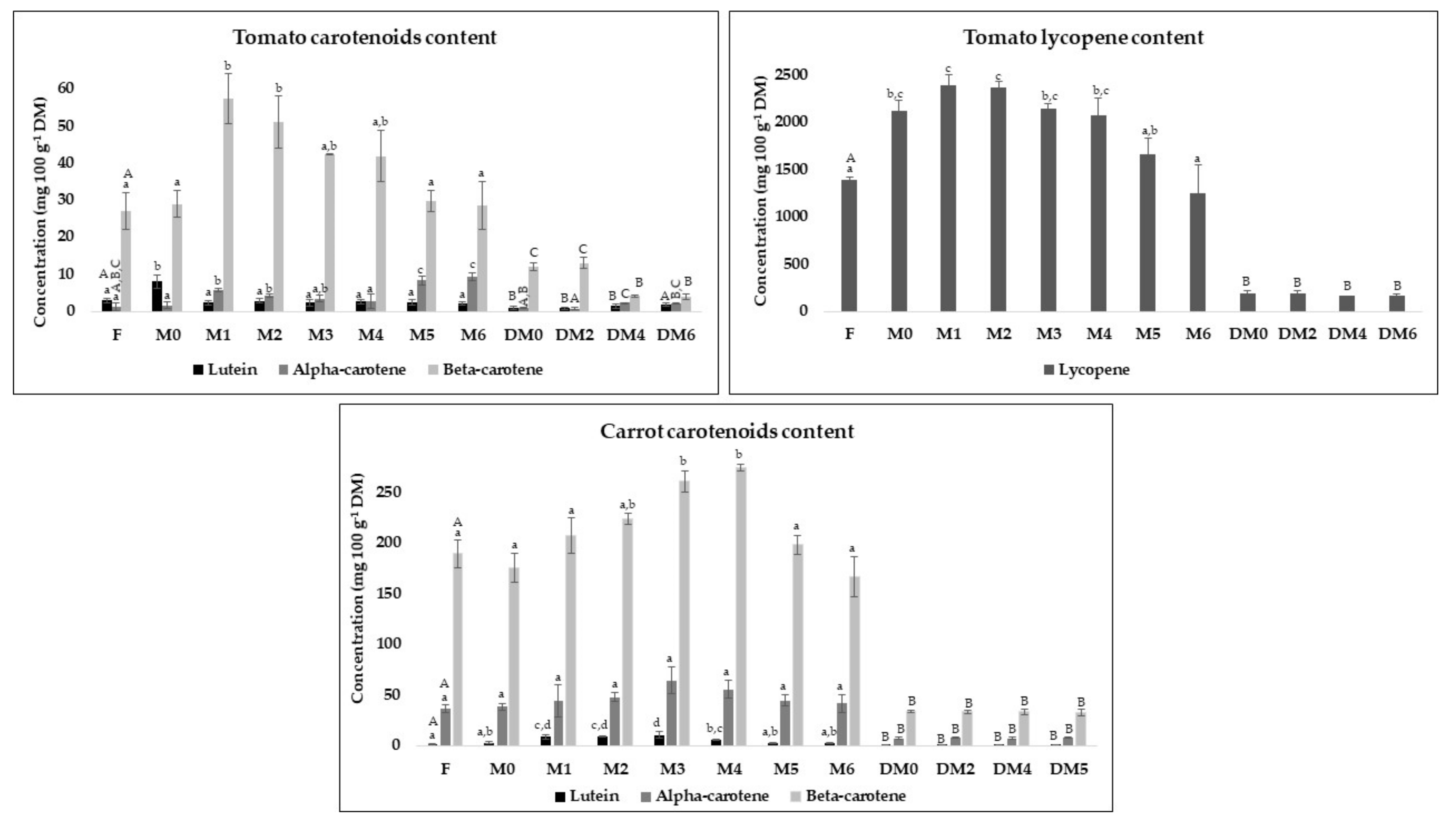
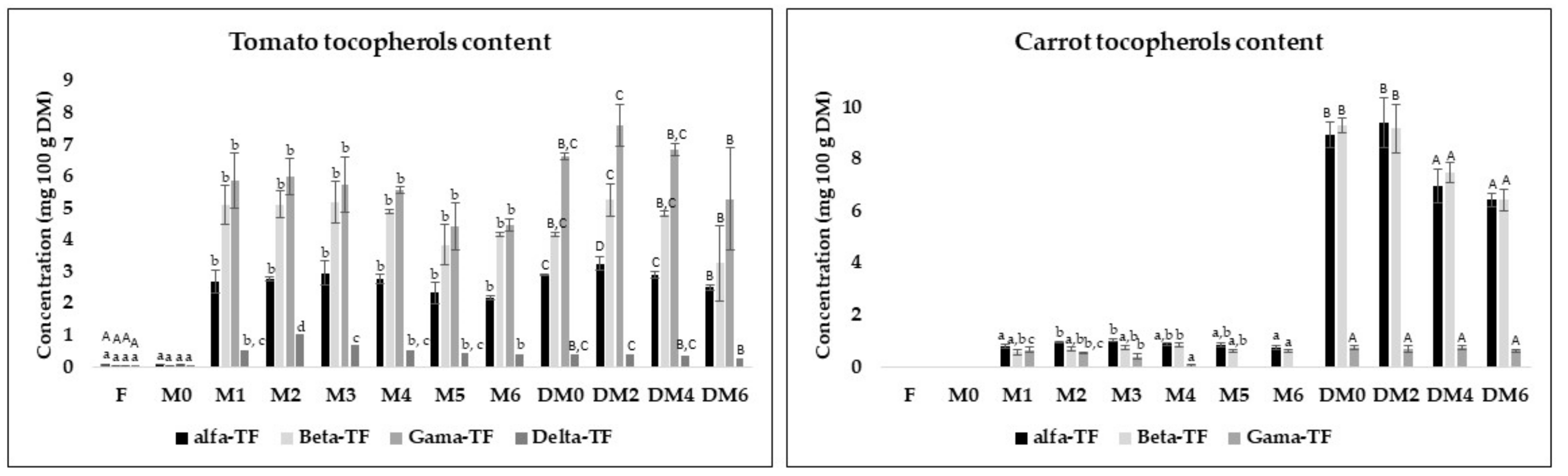
3.5. Carotenoids and Vitamine E Content
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Jenkins, W.; Tucker, M.E.; Grim, J. Routledge Handbook of Religion and Ecology; Routledge: London, UK, 2016. [Google Scholar] [CrossRef]
- Keser, D.; Guclu, G.; Kelebek, H.; Keskin, M.; Soysal, Y.; Sekerli, Y.E.; Selli, S. Characterization of aroma and phenolic composition of carrot (Daucus carota ‘Nantes’) powders obtained from intermittent microwave drying using GC–MS and LC–MS/MS. Food Bioprod. Process. 2020, 119, 350–359. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Toydemir, G.; Boyacioglu, D.; Beekwilder, J.; Hall, R.; Capanoglu, E. A review on the effect of drying on antioxidant potential of fruits and vegetables. Crit. Rev. Food Sci. Nutr. 2016, 56, S110–S129. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Aggarwal, P. Drying method affects bioactive compounds and antioxidant activity of carrot. Int. J. Veg. Sci. 2014, 21, 467–481. [Google Scholar] [CrossRef]
- Kyriacou, M.; Rouphael, Y. Towards a new definition of quality for fresh fruits and vegetables. Sci. Hortic. 2018, 234, 463–469. [Google Scholar] [CrossRef]
- Patras, A.; Tiwari, B.; Brunton, N. Influence of blanching and low temperature preservation strategies on antioxidant activity and phytochemical content of carrots, green beans and broccoli. LWT 2011, 44, 299–306. [Google Scholar] [CrossRef]
- Hung, P.V.; Duy, T.L. Effects of drying methods on bioactive compounds of vegetables and correlation between bioactive compounds and their antioxidantsfile. Int. Food Res. J. 2012, 19, 327–332. [Google Scholar]
- Nayak, B.; Liu, R.H.; Tang, J. Effect of processing on phenolic antioxidants of fruits, vegetables, and grains—A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 887–918. [Google Scholar] [CrossRef] [PubMed]
- Ribas-Agustí, A.; Martín-Belloso, O.; Soliva-Fortuny, R.; Elez-Martínez, P. Food processing strategies to enhance phenolic compounds bioaccessibility and bioavailability in plant-based foods. Crit. Rev. Food Sci. Nutr. 2018, 58, 2531–2548. [Google Scholar] [CrossRef] [Green Version]
- Neri, L.; Faieta, M.; Di Mattia, C.; Sacchetti, G.; Mastrocola, D.; Pittia, P. Antioxidant activity in frozen plant foods: Effect of cryoprotectants, freezing process and frozen storage. Foods 2020, 9, 1886. [Google Scholar] [CrossRef]
- Murcia, M.A.; Jiménez, A.M.; Martínez-Tomé, M. Vegetables antioxidant losses during industrial processing and refrigerated storage. Food Res. Int. 2009, 42, 1046–1052. [Google Scholar] [CrossRef]
- Celli, G.B.; Ghanem, A.; Brooks, M.S.-L. Influence of freezing process and frozen storage on the quality of fruits and fruit products. Food Rev. Int. 2015, 32, 280–304. [Google Scholar] [CrossRef]
- Ijabadeniyi, O.A.; Pillay, Y. Microbial safety of low water activity Foods: Study of simulated and durban household samples. J. Food Qual. 2017, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, B.; Falco, V.; Moutinho-Pereira, J.; Bacelar, E.; Peixoto, F.; Correia, C. Effects of elevated CO2 on grapevine (Vitis vinifera L.): Volatile composition, phenolic content, and in vitro antioxidant activity of red wine. J. Agric. Food Chem. 2009, 57, 265–273. [Google Scholar] [CrossRef]
- Coscueta, E.R.; Campos, D.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic soy protein hydrolysis: A tool for biofunctional food ingredient production. Food Chem. X 2019, 1, 100006. [Google Scholar] [CrossRef] [PubMed]
- Schaich, K.; Tian, X.; Xie, J. Reprint of “Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays”. J. Funct. Foods 2015, 18, 782–796. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin-Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef] [PubMed]
- Campos, D.A.; Ribeiro, T.B.; Teixeira, J.A.; Pastrana, L.; Pintado, M.M. Integral valorization of pineapple (Ananas comosus L.) By-products through a green chemistry approach towards Added Value Ingredients. Foods 2020, 9, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, A.; Pintado, M.M.; Almeida, D. Phytochemical composition and antioxidant activity of peach as affected by pasteurization and storage duration. LWT 2012, 49, 202–207. [Google Scholar] [CrossRef]
- Prates, J.A.M.; Quaresma, M.A.G.; Bessa, R.J.B.; Fontes, C.M.A.; Alfaia, C.M.M. Simultaneous HPLC quantification of total cholesterol, tocopherols and β-carotene in Barrosã-PDO veal. Food Chem. 2006, 94, 469–477. [Google Scholar] [CrossRef]
- Slavin, M.; Yu, L. A single extraction and HPLC procedure for simultaneous analysis of phytosterols, tocopherols and lutein in soybeans. Food Chem. 2012, 135, 2789–2795. [Google Scholar] [CrossRef]
- Vandekinderen, I.; Van Camp, J.; Devlieghere, F.; Veramme, K.; Denon, Q.; Ragaert, P.; De Meulenaer, B. Effect of Decontamination Agents on the Microbial Population, Sensorial Quality, and Nutrient Content of Grated Carrots (Daucus carota L.). J. Agric. Food Chem. 2008, 56, 5723–5731. [Google Scholar] [CrossRef]
- Saraiva, M.; Correia, C.B.; Cunha, I.C.; Maia, C.; Bonito, C.C.; Furtado, R.; Calhau, A. Interpretação de Resultados de Ensaios Microbiológicos em Alimentos Prontos Para Consumo e em Superfícies do Ambiente de Preparação e Distribuição Alimentar: Valores-guia; INSA IP—Instituto Nacional de Saúde Doutor Ricardo Jorge: Lisboa, Portugal, 2019. [Google Scholar]
- Lisiewska, Z.; Kmiecik, W. Effect of storage period and temperature on the chemical composition and organoleptic quality of frozen tomato cubes. Food Chem. 2000, 70, 167–173. [Google Scholar] [CrossRef]
- Bouzari, A.; Holstege, D.; Barrett, D.M. Mineral, fiber, and total phenolic retention in eight fruits and vegetables: A comparison of refrigerated and frozen storage. J. Agric. Food Chem. 2015, 63, 951–956. [Google Scholar] [CrossRef] [PubMed]
- Leja, M.; Kamińska, I.; Kramer, M.; Maksylewicz-Kaul, A.; Kammerer, D.; Carle, R.; Baranski, R. The content of phenolic compounds and radical scavenging activity varies with carrot origin and root color. Plant. Foods Hum. Nutr. 2013, 68, 163–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgé, S.; Tourniaire, F.; Gautier, H.; Goupy, P.; Rock, E.; Caris-Veyrat, C. Changes in the contents of carotenoids, phenolic compounds and vitamin C during technical processing and lyophilisation of red and yellow tomatoes. Food Chem. 2011, 124, 1603–1611. [Google Scholar] [CrossRef]
- Perea-Domínguez, X.P.; Hernández-Gastelum, L.Z.; Olivas-Olguin, H.R.; Espinosa-Alonso, L.G.; Valdez-Morales, M.; Medina-Godoy, S. Phenolic composition of tomato varieties and an industrial tomato by-product: Free, conjugated and bound phenolics and antioxidant activity. J. Food Sci. Technol. 2018, 55, 3453–3461. [Google Scholar] [CrossRef]
- Smith, J.C.; Biasi, W.V.; Holstege, D.; Mitcham, E.J. Effect of passive drying on ascorbic acid, α-tocopherol, and β-carotene in tomato and mango. J. Food Sci. 2018, 83, 1412–1421. [Google Scholar] [CrossRef]
- Char, C.D. Carrots (Daucus carota L.). Fruit Veg. Phytochem. 2017, 969–978. [Google Scholar]
- Kamiloglu, S.; Demirci, M.; Selen, S.; Toydemir, G.; Boyacioglu, D.; Capanoglu, E. Home processing of tomatoes (Solanum lycopersicum): Effects on in vitro bioaccessibility of total lycopene, phenolics, flavonoids, and antioxidant capacity. J. Sci. Food Agric. 2014, 94, 2225–2233. [Google Scholar] [CrossRef]
- Danesi, F.; Bordoni, A. Effect of home freezing and Italian style of cooking on antioxidant activity of edible vegetables. J. Food Sci. 2008, 73, H109–H112. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Le, M.D. Influence of various drying conditions on phytochemical compounds and antioxidant activity of carrot peel. Beverages 2018, 4, 80. [Google Scholar] [CrossRef] [Green Version]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Macheix, J.-J.; Fleuriet, A.; Billot, J. Fruit Phenolics; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar] [CrossRef]
- Bénard, C.; Gautier, H.; Bourgaud, F.; Grasselly, D.; Navez, B.; Caris-Veyrat, C.; Weiss, M.; Génard, M. Effects of low nitrogen supply on tomato (Solanum lycopersicum) fruit yield and quality with special emphasis on sugars, acids, ascorbate, carotenoids, and phenolic compounds. J. Agric. Food Chem. 2009, 57, 4112–4123. [Google Scholar] [CrossRef]
- Sun, T.; Simon, P.; Tanumihardjo, S.A. Antioxidant phytochemicals and antioxidant capacity of biofortified carrots (Daucus carota L.) of various colors. J. Agric. Food Chem. 2009, 57, 4142–4147. [Google Scholar] [CrossRef] [PubMed]
- Arscott, S.A.; Tanumihardjo, S.A. Carrots of many colors provide basic nutrition and bioavailable phytochemicals acting as a functional food. Compr. Rev. Food Sci. Food Saf. 2010, 9, 223–239. [Google Scholar] [CrossRef]
- Zhang, D.; Hamauzu, Y. Phenolic compounds and their antioxidant properties in different tissues of carrots (Daucus carota L.). Int. J. Food Agric. Environ. 2004, 2, 332. [Google Scholar]
- Gómez-García, R.; Campos, D.A.; Oliveira, A.; Aguilar, C.N.; Madureira, A.R.; Pintado, M. A chemical valorisation of melon peels towards functional food ingredients: Bioactives profile and antioxidant properties. Food Chem. 2021, 335, 127579. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, P.; Sharma, A.; Singh, B.; Nagpal, A.K. Bioactivities of phytochemicals present in tomato. J. Food Sci. Technol. 2018, 55, 2833–2849. [Google Scholar] [CrossRef]
- Martí, R.; Roselló, S.; Cebolla-Cornejo, J. Tomato as a source of carotenoids and polyphenols targeted to cancer prevention. Cancers 2016, 8, 58. [Google Scholar] [CrossRef]
- Hidalgo, A.; Di Prima, R.; Fongaro, L.; Cappa, C.; Lucisano, M. Tocols, carotenoids, heat damage and technological quality of diced tomatoes processed in different industrial lines. LWT 2017, 83, 254–261. [Google Scholar] [CrossRef]
- Saini, R.K.; Zamany, A.J.; Keum, Y.S. Ripening improves the content of carotenoid, α-tocopherol, and polyunsaturated fatty acids in tomato (Solanum lycopersicum L.) fruits. 3 Biotech 2017, 7, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Slavin, J.L.; Lloyd, B. Health benefits of fruits and vegetables. Adv. Nutr. 2012, 1, 506–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behsnilian, D.; Mayer-Miebach, E. Impact of blanching, freezing and frozen storage on the carotenoid profile of carrot slices (Daucus carota L. cv. Nutri Red). Food Control. 2017, 73, 761–767. [Google Scholar]
- Bermudez, L.F.; Del Pozo, T.; Lira, B.S.; De Godoy, F.; Boos, I.; Romanó, C.; Previtali, V.; Almeida, J.; Bréhélin, C.; Asis, R.; et al. A Tomato Tocopherol Binding Protein Sheds Light on Intracellular α-tocopherol Metabolism in Plants. Plant. Cell Physiol. 2018, 59, 2188–2203. [Google Scholar] [CrossRef] [Green Version]
- Ombódi, A.; Daood, H.G.; Helyes, L. Carotenoid and tocopherol composition of an orange-colored carrot as affected by water supply. HortScience 2014, 49, 729–733. [Google Scholar]
- Esposito, F.; Arlotti, G.; Bonifati, A.M.; Napolitano, A.; Vitale, D.; Fogliano, V. Antioxidant activity and dietary Wbre in durum wheat bran by-products. Food Res. Int. 2005, 38, 1167–1173. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetable | Sample | TAB | Enterobacteriacea | Yeasts and Molds | Bacillus cereus spp. | Aerobic Spore-Forms | Anaerobic Spore-Forms |
|---|---|---|---|---|---|---|---|
| Tomato | Non washed | 8.14 ± 0.15 | 6.28 ± 0.14 | 6.16 ± 0.12 | 4.63 ± 0.14 | - | - |
| Washed | 4.32 ± 0.14 | 3.82 ± 0.07 | 3.77 ± 0.07 | 2.57 ± 0.12 | - | - | |
| Fresh pulp | 4.54 ± 0.11 | 3.91 ± 0.08 | 3.89 ± 0.15 | 2.65 ± 0.17 | - | - | |
| M0 | 4.55 ± 0.15 | 3.98 ± 0.07 | 3.91 ± 0.10 | 2.57 ± 0.14 | - | - | |
| M1 | 4.52 ± 0.07 | 4.03 ± 0.06 | 3.85 ± 0.14 | 2.53 ± 0.14 | - | - | |
| M2 | 4.52 ± 0.15 | 4.00 ± 0.01 | 3.92 ± 0.11 | 2.56 ± 0.04 | - | - | |
| M3 | 4.56 ± 0.04 | 4.02 ± 0.11 | 3.87 ± 0.21 | 2.51 ± 0.09 | - | - | |
| M4 | 4.68 ± 0.10 | 4.13 ± 0.05 | 3.92 ± 0.20 | 2.63 ± 0.11 | - | - | |
| M5 | 4.61 ± 0.13 | 4.07 ± 0.07 | 3.89 ± 0.18 | 2.56 ± 0.09 | - | - | |
| M6 | 4.39 ± 0.01 | 3.80 ± 0.38 | 3.87 ± 0.27 | 2.62 ± 0.06 | - | - | |
| DM0 | 2.39 ± 0.12 | 2.74 ± 1.04 | - | - | 3.00 ± 0.00 | 3.00 ± 0.00 | |
| DM6 | 2.15 ± 0.21 | 2.15 ± 0.21 | - | - | 2.00 ± 0.00 | 2.39 ± 0.12 | |
| Carrot | Non washed | 9.18 ± 0.10 | 8.03 ± 0.10 | 5.96 ± 0.10 | 4.77 ± 0.08 | - | - |
| Washed | 5.29 ± 0.10 | 4.63 ± 0.13 | 3.65 ± 0.16 | 2.90 ± 0.09 | - | - | |
| Fresh pulp | 5.36 ± 0.09 | 4.82 ± 0.10 | 3.66 ± 0.11 | 2.93 ± 0.05 | - | - | |
| M0 | 5.34 ± 0.15 | 4.85 ± 0.13 | 3.69 ± 0.03 | 2.74 ± 0.15 | - | - | |
| M1 | 5.27 ± 0.07 | 4.82 ± 0.10 | 3.81 ± 0.08 | 2.69 ± 0.17 | - | - | |
| M2 | 5.26 ± 0.06 | 4.86 ± 0.07 | 3.74 ± 0.29 | 2.70 ± 0.05 | - | - | |
| M3 | 5.23 ± 0.01 | 4.83 ± 0.09 | 3.73 ± 0.03 | 2.67 ± 0.09 | - | - | |
| M4 | 5.31 ± 0.37 | 4.54 ± 0.21 | 3.76 ± 0.10 | 2.82 ± 0.16 | - | - | |
| M5 | 5.28 ± 0.02 | 4.81 ± 0.21 | 3.72 ± 0.12 | 2.65 ± 0.16 | - | - | |
| M6 | 5.22 ± 0.06 | 4.82 ± 0.24 | 3.73 ± 0.03 | 2.79 ± 0.06 | - | - | |
| DM0 | 4.49 ± 0.88 | 4.22 ± 1.10 | - | - | 2.67 ± 0.95 | 2.36 ± 0.51 | |
| DM6 | 4.61 ± 0.88 | 4.16 ± 1.00 | - | - | 2.85 ± 0.00 | 2.50 ± 0.28 |
| Vegetable | Sample | Viscosity (mPa s−1) |
|---|---|---|
| Tomato | Fresh pulp | 195.88 ± 3.73 a |
| M0 | 153.36 ± 1.27 b | |
| M1 | 142.42 ± 1.27 c | |
| M2 | 133.98 ± 2.15 d | |
| M3 | 122.04 ± 0.97 e | |
| M4 | 110.98 ± 3.66 f | |
| M5 | 105.08 ± 0.74 g | |
| M6 | 100.34 ± 1.39 h | |
| Carrot | Fresh pulp | 486.98 ± 1.40 a |
| M0 | 399.98 ± 4.35 b | |
| M1 | 390.58 ± 1.78 c | |
| M2 | 380.58 ± 3.43 d | |
| M3 | 368.94 ± 2.2 e | |
| M4 | 358.62 ± 2.41 f | |
| M5 | 351.28 ± 2.37 g | |
| M6 | 349.28 ± 2.92 g |
| Tomato Sample | Chlorogenic Acid | Sinapic Acid Derivative | Rutin | Quercetin Derivative | Naringenin |
|---|---|---|---|---|---|
| Fresh | 27.2 ± 2.6 b,A | 0.46 ± 0.28 a | 11.4 ± 0.8 b,c,d,A | 16.9 ± 0.3 c | 500.0 ± 27.4 a,b,* |
| M0 | 32.4 ± 0.7 c | 0.67 ± 0.09 a | 11.2 ± 0.4 b,c,d | 27.5 ± 2.0 d | 501.3 ± 10.7 a.b,* |
| M1 | 18.4 ± 0.9 a | 0.64 ± 0.28 a | 8.4 ± 2.0 a,b | 13.0 ± 0.9 a,b | 445.3 ± 5.9 b,c,* |
| M2 | 16.6 ± 0.5 a | 0.52 ± 0.16 a | 9.9 ± 1.4 a,b,c | 10.8 ± 0.7 a | 367.1 ± 48.1 c,e,* |
| M3 | 19.5 ± 2.7 a | 0.56 ± 0.30 a | 8.0 ± 2.2 a | 13.3 ± 1.0 b | 367.6 ± 52.4 c,e,* |
| M4 | 17.2 ± 2.4 a | 0.40 ± 0.12 a | 8.6 ± 0.3 a,b | 13.1 ± 0.8 b | 314.5 ± 93.8 e,* |
| M5 | 17.2 ± 0.3 a | 1.50 ± 0.02 b | 12.5 ± 0.3 c,d | 17.3 ± 0.4 c | 556.0 ± 7.4 a,* |
| M6 | 15.9 ± 0.1 a | 1.67 ± 0.09 b | 13.2 ± 1.3 d | 16.4 ± 0.4 c | 514.3 ± 3.1 a.b,* |
| DM0 | 35.6 ± 3.5 B | N.D. | 30.5 ± 3.1 B | N.D. | 1203.5 ± 142.3 A,B |
| DM2 | 34.2 ± 0.2 B | N.D. | 32.0 ± 1.0 B | N.D. | 1311.5 ± 29.3 B |
| DM4 | 35.4 ± 0.1 B | N.D. | 30.4 ± 0.2 B | N.D. | 1132.2 ± 16.5 A |
| DM6 | 38.0 ± 0.3 B | N.D. | 32.0 ± 0.2 B | N.D. | 1224.8 ± 22.7 A,B |
| Carrot Sample | Protocatechuic Acid | Chlorogenic Acid | Caffeic Acid | Trans-Ferulic Acid |
|---|---|---|---|---|
| Fresh | B.Q.L. | 2.48 ± 0.07 a,A | N.D. | 3.02 ± 0.03 |
| M0 | B.Q.L. | 2.85 ± 0.23 a | N.D. | B.Q.L. |
| M1 | 14.28 ± 1.67 | 5.36 ± 2.72 a | N.D. | B.Q.L. |
| M2 | B.Q.L. | 16.58 ± 0.43 b | B.Q.L. | N.D. |
| M3 | B.Q.L. | 39.32 ± 2.51 c | B.Q.L. | N.D. |
| M4 | B.Q.L. | 53.19 ± 1.88 d | B.Q.L. | N.D. |
| M5 | B.Q.L. | 62.13 ± 0.41 e | 0.25 ± 0.02 | N.D. |
| M6 | B.Q.L. | 61.51 ± 0.42 e | B.Q.L. | N.D. |
| DM0 | N.D. | 56.22 ± 3.15 D | 0.41 ± 0.03 | N.D. |
| DM2 | N.D. | 40.04 ± 0.31 C | 0.18 ± 0.01 | N.D. |
| DM4 | N.D. | 37.76 ± 0.08 B,C | B.Q.L. | N.D. |
| DM6 | N.D. | 36.18 ± 0.02 B | B.Q.L. | N.D. |
| Phenolic Acids | Chemical Formula | Retention Time (min) | λ Max (nm) |
|---|---|---|---|
| Hydroxybenzoic acid | |||
| Protocatechuic acid | C7H6O4 | 17.4 | 294 |
| Hydroxycinnamic acid | |||
| Chlorogenic acid | C16H18O9 | 27.2 | 326.0 |
| Caffeic acid | C9H8O4 | 31.0 | 323.6 |
| p-coumaric acid | C9H8O3 | 39.9 | 309.3 |
| Ferulic acid | C10H10O4 | 42.4 | 322.4 |
| Sinapic acid | C11H12O5 | 42.7 | 322.0 |
| Trans-ferulic acid | C10H10O4 | 42.6 | 322.4 |
| Iso-ferulic acid | C10H10O4 | 45.3 | 322.4 |
| Flavonols | |||
| Myricetin | C15H10O8 | 38.9 | 375.3 |
| Rutin | C27H30O16 | 49.8 | 354.7 |
| Quercetin | C15H10O7 | 49.9 | 343.9 |
| Flavanones | |||
| Naringenin | C15H12O5 | 48.8 | 283.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Araújo-Rodrigues, H.; Santos, D.; Campos, D.A.; Ratinho, M.; M. Rodrigues, I.; E. Pintado, M. Development of Frozen Pulps and Powders from Carrot and Tomato by-Products: Impact of Processing and Storage Time on Bioactive and Biological Properties. Horticulturae 2021, 7, 185. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070185
Araújo-Rodrigues H, Santos D, Campos DA, Ratinho M, M. Rodrigues I, E. Pintado M. Development of Frozen Pulps and Powders from Carrot and Tomato by-Products: Impact of Processing and Storage Time on Bioactive and Biological Properties. Horticulturae. 2021; 7(7):185. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070185
Chicago/Turabian StyleAraújo-Rodrigues, Helena, Diva Santos, Débora A. Campos, Modesta Ratinho, Ivo M. Rodrigues, and Manuela E. Pintado. 2021. "Development of Frozen Pulps and Powders from Carrot and Tomato by-Products: Impact of Processing and Storage Time on Bioactive and Biological Properties" Horticulturae 7, no. 7: 185. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7070185