
Variability of Polyphenolic Compounds and Biological Activities among Perilla frutescens var. crispa Genotypes


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Extract Preparations
2.2. Determination of Phytochemical Contents
2.3. Analysis of Antioxidant Activities
2.4. Determination of Antimelanogenic Effect
2.5. Determination of Nitric Oxide (NO) Production
2.6. Cell Viability Assay
2.7. Quantitative Real-Time PCR (qRT-PCR)
2.8. Statistical Analysis
3. Results and Discussion
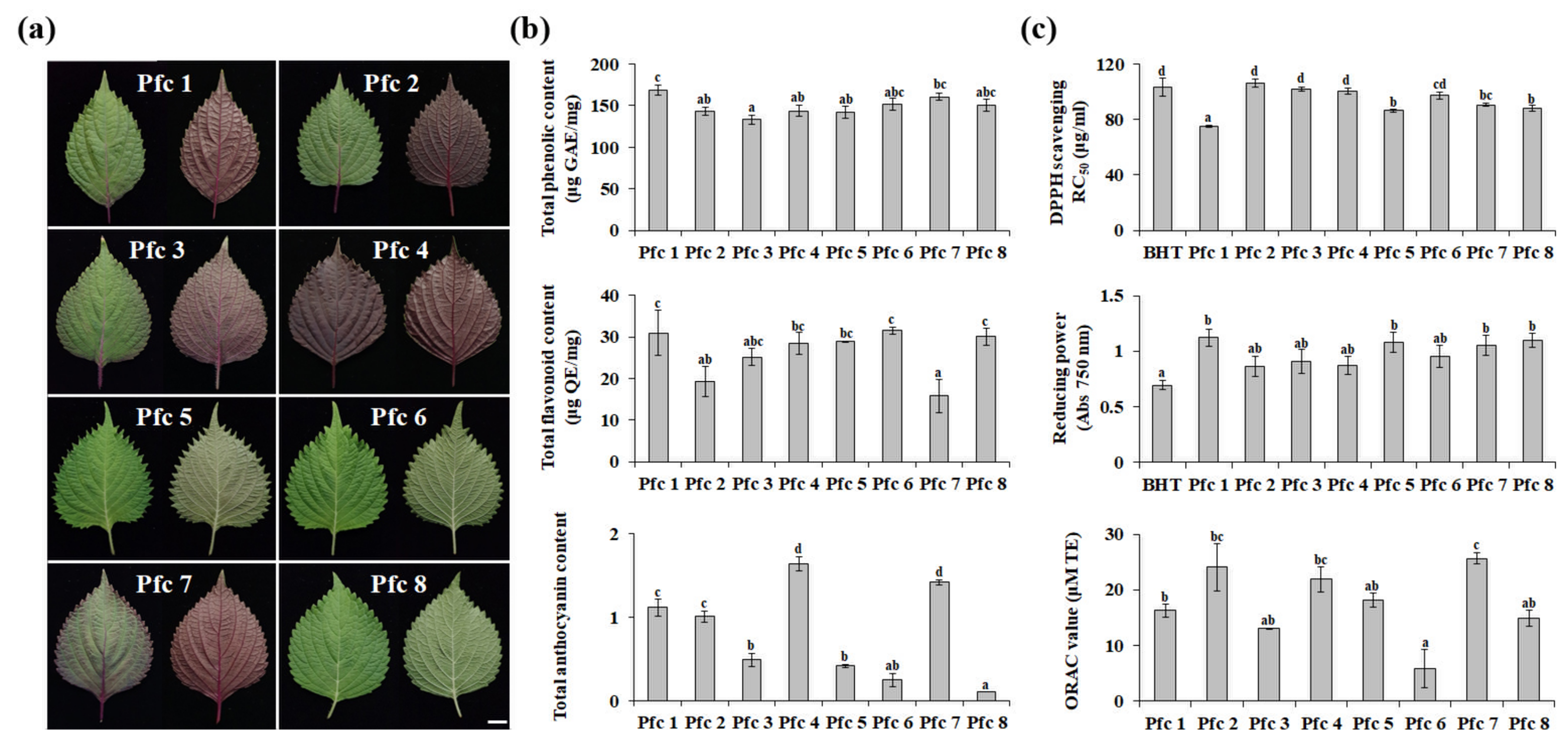
3.1. Phytochemical Compounds and Antioxidant Activities of Pfc Genotypes
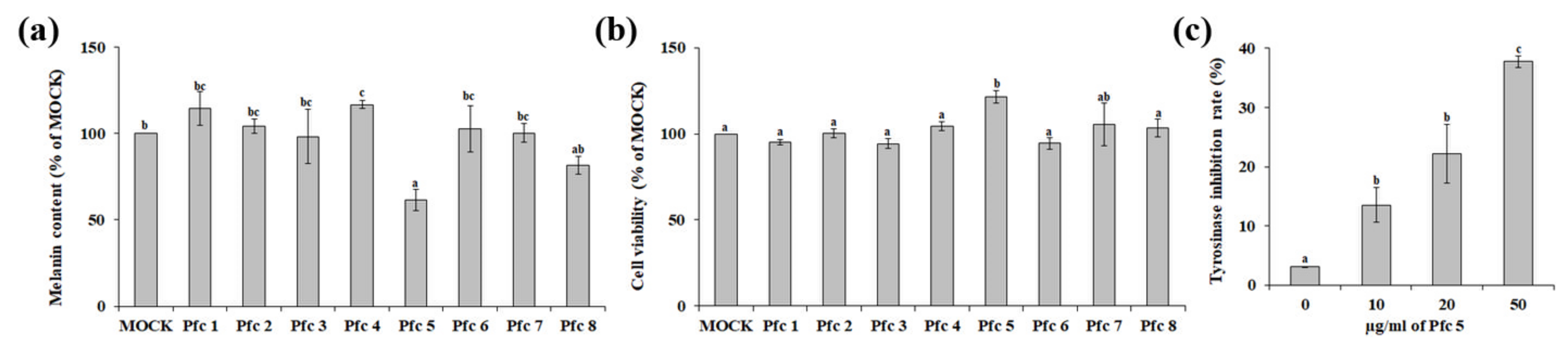
3.2. Variation in Antimelanogenic Principles of Pfc Genotypes
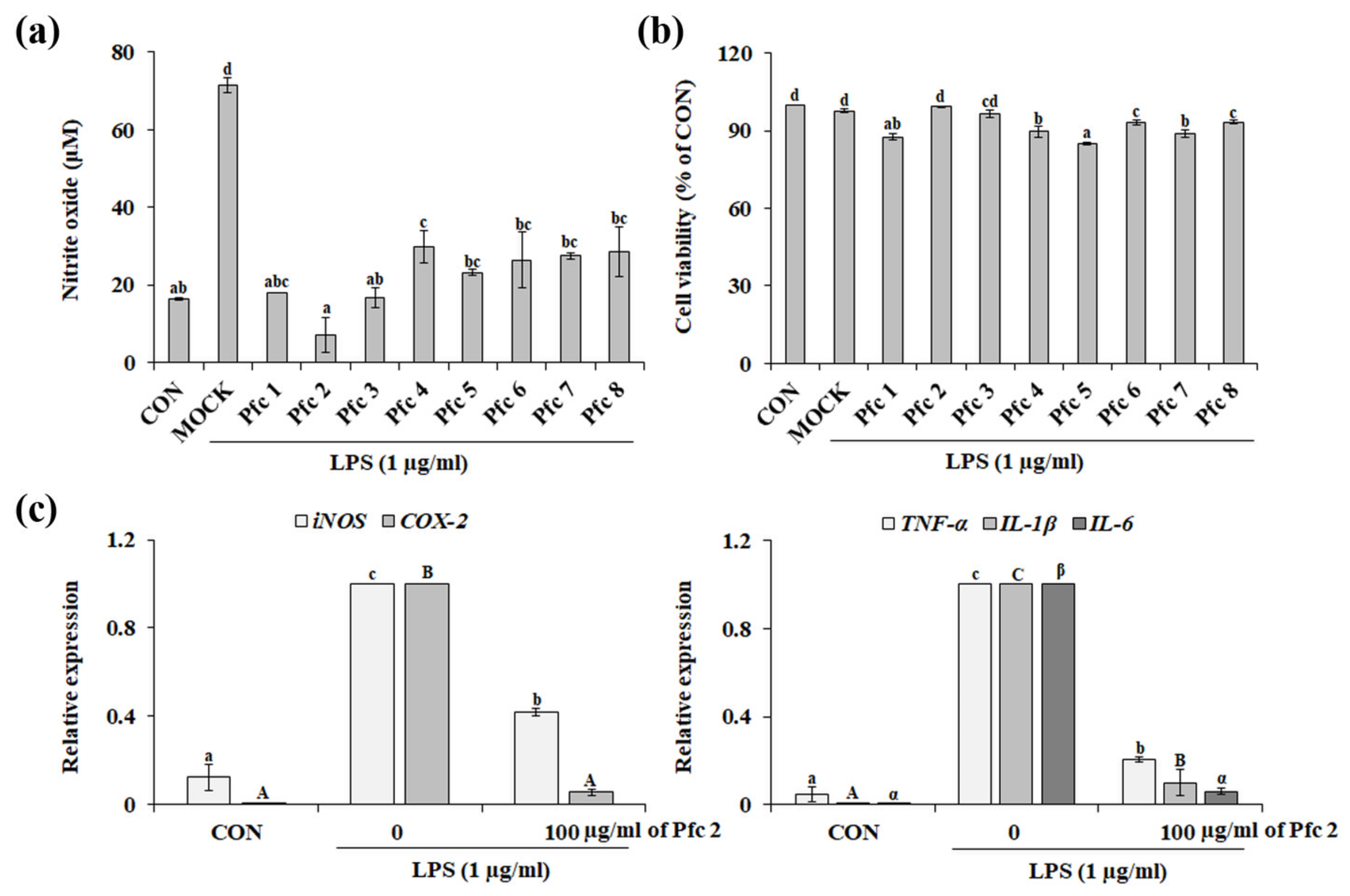
3.3. Variation in Anti-Inflammatory Effects among Genotypes of Pfc
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, P.; Penny, J.; Partington, D.L.; Panozzo, J.F. Genotype and environment effects on the chemical composition and rheological properties of field peas. J. Sci. Food Agric. 2019, 99, 5409–5416. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Bazargani, M.; Falahati-Anbaran, M.; Rohloff, J. Comparative analyses of phytochemical variation within and between congeneric species of willow herb, Epilobium hirsutum and E. parviflorum: Contribution of environmental factors. Front. Plant Sci. 2021, 11, 595190. [Google Scholar] [CrossRef] [PubMed]
- Balada, C.; Castro, M.; Fassio, C.; Zamora, A.; Marchant, M.J.; Acevedo, W.; Guzmán, L. Genetic diversity and biological activity of Curcuma longa ecotypes from Rapa Nui using molecular markers. Saudi J. Biol. Sci. 2021, 28, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Ju, H.J.; Bang, J.-H.; Chung, J.-W.; Hyun, T.K. Variation in essential oil composition and antimicrobial activity among different genotypes of Perilla frutescens var. crispa. J. Appl. Biol. Chem. 2021, 64, 127–131. [Google Scholar] [CrossRef]
- Ahmed, H.M. Ethnomedicinal, phytochemical and pharmacological investigations of Perilla frutescens (L.) Britt. Molecules 2019, 24, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saita, E.; Kishimoto, Y.; Tani, M.; Iizuka, M.; Toyozaki, M.; Sugihara, N.; Kondo, K. Antioxidant activities of Perilla frutescens against low-density lipoprotein oxidation in vitro and in human subjects. J. Oleo Sci. 2012, 61, 113–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.Y.; Leu, Y.L.; Fang, Y.; Lin, C.F.; Kuo, L.M.; Sung, W.C.; Tsai, Y.F.; Chung, P.J.; Lee, M.C.; Kuo, Y.T.; et al. Anti-inflammatory effects of Perilla frutescens in activated human neutrophils through two independent pathways: Src family kinases and calcium. Sci. Rep. 2015, 5, 18204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, C.H.; So, Y.; Kim, H.Y.; Han, S.N.; Kim, J.B. Anti-Arthritic activities of supercritical carbon dioxide extract derived from radiation mutant Perilla frutescens var. crispa in collagen antibody-induced arthritis. Nutrients 2019, 11, 2959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sa, K.J.; Choi, I.Y.; Park, K.C.; Lee, J.K. Genetic diversity and population structure among accessions of Perilla frutescens (L.) Britton in East Asia using new developed microsatellite markers. Genes Genom. 2018, 40, 1319–1329. [Google Scholar] [CrossRef]
- Ma, S.J.; Sa, K.J.; Hong, T.K.; Lee, J.K. Genetic diversity and population structure analysis in Perilla crop and their weedy types from northern and southern areas of China based on simple sequence repeat (SSRs). Genes Genom. 2019, 41, 267–281. [Google Scholar] [CrossRef]
- Lim, H.J.; Woo, K.W.; Lee, K.R.; Lee, S.K.; Kim, H.P. Inhibition of proinflammatory cytokine generation in lung inflammation by the leaves of Perilla frutescens and its constituents. Biomol. Ther. 2014, 22, 62–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.; Eom, S.H.; Kim, J.S.; Jo, I.H.; Hyun, T.K. Influence of ripening stages on phytochemical composition and bioavailability of ginseng berry (Panax ginseng C.A. Meyer). J. Appl. Bot. Food Qual. 2019, 92, 130–137. [Google Scholar]
- Yoo, T.K.; Kim, J.S.; Hyun, T.K. Polyphenolic composition and anti-melanoma activity of white forsythia (Abeliophyllum distichum Nakai) organ extracts. Plants 2020, 9, 757. [Google Scholar] [CrossRef]
- Ju, H.J.; Yoo, T.K.; Jin, S.; Kim, H.; Hyun, T.K. In vitro evaluation of the pharmacological properties of crude methanol extract and its fractions of Aconitum austrokoreense aerial parts. Rev. Mex. Ing. Quim. 2020, 19, 1341–1350. [Google Scholar] [CrossRef]
- Jin, S.; Hyun, T.K. Ectopic expression of production of anthocyanin pigment 1 (PAP1) improves the antioxidant and anti-melanogenic properties of ginseng (Panax ginseng C.A. Meyer) hairy roots. Antioxidants 2020, 9, 922. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Park, G.; Son, H.J.; Lee, S.J. Suppression of α-MSH and IBMX-induced melanogenesis by cordycepin via inhibition of CREB and MITF, and activation of PI3K/Akt and ERK-dependent mechanisms. Int. J. Mol. Med. 2012, 29, 119–124. [Google Scholar] [PubMed] [Green Version]
- Yoo, T.K.; Jeong, W.T.; Kim, J.G.; Ji, H.S.; Ahn, M.A.; Chung, J.W.; Lim, H.B.; Hyun, T.K. UPLC-ESI-Q-TOF-MS-Based metabolite profiling, antioxidant and anti-inflammatory properties of different organ extracts of Abeliophyllum distichum. Antioxidants 2021, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Leonov, A.; Arlia-Ciommo, A.; Piano, A.; Svistkova, V.; Lutchman, V.; Medkour, Y.; Titorenko, V.I. Longevity extension by phytochemicals. Molecules 2015, 20, 6544–6572. [Google Scholar] [CrossRef] [Green Version]
- Abbaszadeh, H.; Keikhaei, B.; Mottaghi, S. A review of molecular mechanisms involved in anticancer and antiangiogenic effects of natural polyphenolic compounds. Phytother. Res. 2019, 33, 2002–2014. [Google Scholar] [CrossRef]
- Fukushima, A.; Nakamura, M.; Suzuki, H.; Saito, K.; Yamazaki, M. High-throughput sequencing and de novo assembly of red and green forms of the Perilla frutescens var. crispa transcriptome. PLoS ONE 2015, 10, e0129154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nićiforović, N.; Mihailović, V.; Masković, P.; Solujić, S.; Stojković, A.; Pavlović Muratspahić, D. Antioxidant activity of selected plant species; potential new sources of natural antioxidants. Food Chem. Toxicol. 2010, 48, 3125–3130. [Google Scholar] [CrossRef] [PubMed]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanisms of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, W.; Liu, W.; Zhu, D.; Cao, Y.; Tang, A.; Gong, G.; Su, H. Natural skin-whitening compounds for the treatment of melanogenesis (Review). Exp. Ther. Med. 2020, 20, 173–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, J.-H.; Lee, B.M. Inhibitory effects of plant extracts on tyrosinase, L-DOPA oxidation, and melanin synthesis. J. Toxicol. Environ. Health Part A 2007, 70, 393–407. [Google Scholar] [CrossRef]
- Ha, T.-J.; Hwang, S.-W.; Jung, H.-J.; Park, K.-H.; Yang, M.-S. Apigenin, tyrosinase inhibitor isolated from the flowers of Hemisteptia lyrata Bunge. J. Appl. Biol. Chem. 2002, 445, 170–172. [Google Scholar]
- Yoon, H.S.; Ko, H.-C.; Kim, S.-J.; Kim, S.S. Stimulatory effects of a 5,6,7,3′,4′-pentamethoxyflavone, sinensetin, on melanogenesis in B16/F10 murine melanoma cells. Lat. Am. J. Pharm. 2015, 34, 1087–1092. [Google Scholar]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, T.Y.; Kim, S.H.; Kim, S.H.; Kim, Y.K.; Park, H.J.; Chae, B.S.; Jung, H.J.; Kim, H.M. Inhibitory effect of mast cell-mediated immediate-type allergic reactions in rats by Perilla frutescens. Immunopharmacol. Immunotoxicol. 2000, 22, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.P.; Lin, C.H.; Chen, Y.C.; Kao, S.H. Anti-inflammatory effects of Perilla frutescens leaf extract on lipopolysaccharide-stimulated RAW264.7 cells. Mol. Med. Rep. 2014, 10, 1077–1083. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Shin, D.; Lim, S.S.; Lee, J.Y.; Kang, Y.H. Purple perilla extracts allay ER stress in lipid-laden macrophages. PLoS ONE 2014, 9, e110581. [Google Scholar]
- Toriyabe, M.; Omote, K.; Kawamata, T.; Namiki, A. Contribution of interaction between nitric oxide and cyclooxygenases to the production of prostaglandins in carrageenan-induced inflammation. Anesthesiology 2004, 101, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D.; Zhang, H.; Lao, Y.Z.; Wu, R.; Xu, J.W.; Murad, F.; Bian, K.; Xu, H.X. Anti-inflammatory effect of 1,3,5,7-tetrahydroxy-8-isoprenylxanthone isolated from twigs of Garcinia esculenta on stimulated macrophage. Mediat. Inflamm. 2015, 2015, 350564. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.M.; An, J. Cytokines, inflammation, and pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Compound (µg/g of Extract) | Pfc 1 | Pfc 2 | Pfc 3 | Pfc 4 | Pfc 5 | Pfc 6 | Pfc 7 | Pfc 8 |
|---|---|---|---|---|---|---|---|---|
| Rutin | 2.33 ± 0.14 b,(1) | 0 a | 2.60 ± 0.02 bc | 8.31 ± 1.47 e | 0 a | 4.06 ± 0.10 cd | 5.23 ± 0.30 d | 2.23 ± 0.01 b |
| Taxifolin | 0.99 ± 0.03 d | 0 a | 0.62 ± 0.01 c | 0 a | 1.15 ± 0.02 d | 0.34 ± 0.17 b | 0.07 ± 0.01 a | 1.44 ± 0.01 e |
| Narirutin | 47.98 ± 0.11 c | 2.43 ± 0.03 a | 2.43 ± 0.10 a | 18.68 ± 8.77 b | 44.44 ± 0.07 c | 5.85 ± 0.18 a | 11.45 ± 6.32 ab | 45.56 ± 1.47 c |
| Naringin | 2.27 ± 0.06 a | 2.73 ± 0.01 a,b | 0.94 ± 0.06 a | 5.59 ± 2.67 b | 0.65 ± 0.0.004 a | 1.12 ± 0.24 a | 1.91 ± 0.02 a | 2.04 ± 1.10 a |
| Hesperidin | 0.86 ± 0.02 a | 1.36 ± 0.01 a | 9.01 ± 3.23 b | 1.99 ± 0.12 a | 1.22 ± 0.02 a | 13.78 ± 0.03 c | 2.81 ± 0.02 a | 15.78 ± 0.06 c |
| Neohesperidin | 4.92 ± 0.02 f | 0.87 ± 0.01 b | 0.76 ± 0.01 b | 1.32 ± 0.02 c | 1.59 ± 0.01 d | 0.51 ± 0.01 a | 2.07 ± 0.02 e | 1.20 ± 0.03 c |
| Myricetin | 1.24 ± 0.01 cd | 1.18 ± 0.01 c | 1.06 ± 0.07 b | 1.29 ± 0.02 d | 0 a | 1.71 ± 0.01 e | 1.21 ± 0.01 cd | 0 a |
| Quercetin | 0 a | 0 a | 0 a | 1.16 ± 0.01 b | 1.71 ± 0.01 d | 0 a | 1.55 ± 0.03 c | 0 a |
| Luteolin | 0 a | 0 a | 0 a | 1.02 ± 0.01 b | 0 a | 0 a | 0 a | 0 a |
| Naringenin | 0.49 ± 0.07 ab | 0.50 ± 0.05 ab | 0.48 ± 0.05 ab | 0.54 ± 0.01 ab | 0.36 ± 0.03 a | 0.62 ± 0.12 b | 0.485 ± 0.005 ab | 0.43 ± 0.02 ab |
| Hesperetin | 2.23 ± 0.01 b | 7.62 ± 1.39 c | 1.14 ± 0.57 ab | 0 a | 0 a | 0 a | 0.28 ± 0.003 a | 0 a |
| Apigenin | 0 a | 0 a | 0 a | 0.63 ± 0.003 b | 2.41 ± 0.11 d | 0.77 ± 0.39 b | 1.76 ± 0.01 c | 0.36 ± 0.004 ab |
| Kaempferol | 15.09 ± 0.07 b | 45.61 ± 9.13 c | 5.33 ± 2.19 ab | 3.35 ± 0.01 a | 13.26 ± 1.02 ab | 2.68 ± 0.10 a | 3.35 ± 0.05 a | 9.35 ± 0.69 ab |
| Isorhaemnetin | 2.09 ± 1.16 ab | 4.68 ± 0.54 b | 2.81 ± 1.62 ab | 4.59 ± 0.03 b | 2.30 ± 0.69 ab | 3.54 ± 0.27 ab | 3.71 ± 0.55 ab | 1.76 ± 0.37 a |
| Rhamnetin | 1.86 ± 0.08 a | 2.64 ± 0.20 a | 7.24 ± 2.76 b | 1.93 ± 0.02 a | 12.96 ± 0.28 c | 1.93 ± 0.97 a | 1.73 ± 0.13 a | 13.63 ± 0.06 c |
| Sinensetin | 1.01 ± 0.16 a | 1.50 ± 0.32 ab | 1.09 ± 0.01 a | 1.07 ± 0.06 a | 3.15 ± 0.15 c | 1.74 ± 0.04 b | 1.96 ± 0.16 b | 1.07 ± 0.04 a |
| Nobiletin | 0.39 ± 0.20 b | 1.14 ± 0.22 c | 0 a | 0 a | 1.82 ± 0.12 d | 0 a | 0 a | 0.71 ± 0.004 b |
| Tangeretin | 6.79 ± 0.12 d | 0.54 ± 0.09 b | 0 a | 0 a | 1.07 ± 0.14 c | 0 a | 0 a | 0 a |
| Gallic acid | 5.64 ± 0.15 bc | 4.10 ± 2.05 b | 5.77 ± 0.02 bc | 0 a | 6.65 ± 0.02 c | 0 a | 5.62 ± 0.04 bc | 0 a |
| Protocatecuic acid | 1.52 ± 0.76 a | 2.42 ± 0.02 a | 2.32 ± 0.05 a | 1.48 ± 0.74 a | 2.24 ± 0.01 a | 2.24 ± 0.03 a | 2.36 ± 0.05 a | 1.46 ± 0.73 a |
| p-Hydroxybenzoic acid | 0.69 ± 0.35 a | 1.07 ± 0.06 abc | 1.52 ± 0.08 cd | 1.55 ± 0.08 d | 0.98 ± 0.02 ab | 1.23 ± 0.09 bcd | 1.25 ± 0.08 bcd | 1.24 ± 0.06 bcd |
| Vanillic acid | 5.76 ± 2.89 a | 16.23 ± 0.25 c | 10.87 ± 0.13 b | 10.69 ± 0.10 b | 8.68 ± 0.11 ab | 12.65 ± 2.48 bc | 11.79 ± 0.37 b | 6.28 ± 0.07 a |
| Chlorogenic acid | 2.47 ± 1.24 a | 3.65 ± 0.17 a | 3.27 ± 0.03 a | 3.65 ± 0.11 a | 3.50 ± 0.04 a | 3.32 ± 0.02 a | 3.40 ± 0.03 a | 3.31 ± 0.02 a |
| Caffeic acid | 3.22 ± 1.61 a | 6.37 ± 0.13 b | 4.947 ± 0.26 ab | 5.35 ± 0.01 b | 5.54 ± 0.06 b | 5.08 ± 0.48 ab | 5.25 ± 0.10 b | 13.00 ± 0.28 c |
| Syringic acid | 1.86 ± 0.93 a | 2.61 ± 0.02 ab | 2.66 ± 0.39 ab | 2.75 ± 0.05 ab | 2.56 ± 0.01 ab | 3.78 ± 0.42 b | 7.76 ± 0.05 c | 3.02 ± 0.01 ab |
| p-Coumaric acid | 3.57 ± 1.78 a | 10.38 ± 0.05 b | 12.11 ± 0.11 bc | 17.29 ± 0.10 e | 17.86 ± 0.05 e | 4.08 ± 0.53 a | 16.11 ± 0.11 de | 13.87 ± 1.01 cd |
| Benzoic acid | 1.88 ± 0.94 b | 0 a | 1.75 ± 0.05 b | 2.51 ± 0.03 b | 0 a | 1.23 ± 0.61 ab | 2.30 ± 0.01 b | 2.41 ± 0.04 b |
| Ferullic acid | 2.14 ± 0.01 a | 2.04 ± 0.007 ab | 2.55 ± 0.2 b | 2.13 ± 0.02 ab | 4.14 ± 0.02 c | 2.26 ± 0.05 ab | 2.73 ± 0.03 b | 3.65 ± 0.005 c |
| Sinapinic acid | 4.94 ± 0.50 bc | 6.19 ± 0.02 d | 5.75 ± 0.02 cd | 6.04 ± 0.02 d | 3.86 ± 0.003 a | 5.70 ± 0.52 c,d | 7.36 ± 0.02 e | 4.13 ± 0.31 ab |
| Cinnamic acid | 1.21 ± 0.13 a | 1.44 ± 0.02 bc | 1.71 ± 0.12 d | 1.34 ± 0.005 ab | 1.74 ± 0.1 d | 1.28 ± 0.08 ab | 1.41 ± 0.004 abc | 1.63 ± 0.01 cd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ju, H.J.; Kim, K.C.; Kim, H.; Kim, J.-S.; Hyun, T.K. Variability of Polyphenolic Compounds and Biological Activities among Perilla frutescens var. crispa Genotypes. Horticulturae 2021, 7, 404. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7100404
Ju HJ, Kim KC, Kim H, Kim J-S, Hyun TK. Variability of Polyphenolic Compounds and Biological Activities among Perilla frutescens var. crispa Genotypes. Horticulturae. 2021; 7(10):404. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7100404
Chicago/Turabian StyleJu, Hyun Ju, Kyeoung Cheol Kim, Heekyu Kim, Ju-Sung Kim, and Tae Kyung Hyun. 2021. "Variability of Polyphenolic Compounds and Biological Activities among Perilla frutescens var. crispa Genotypes" Horticulturae 7, no. 10: 404. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7100404