
In Vitro and In Vivo Antifungal Activities of Nine Commercial Essential Oils against Brown Rot in Apples


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Oil Extracts
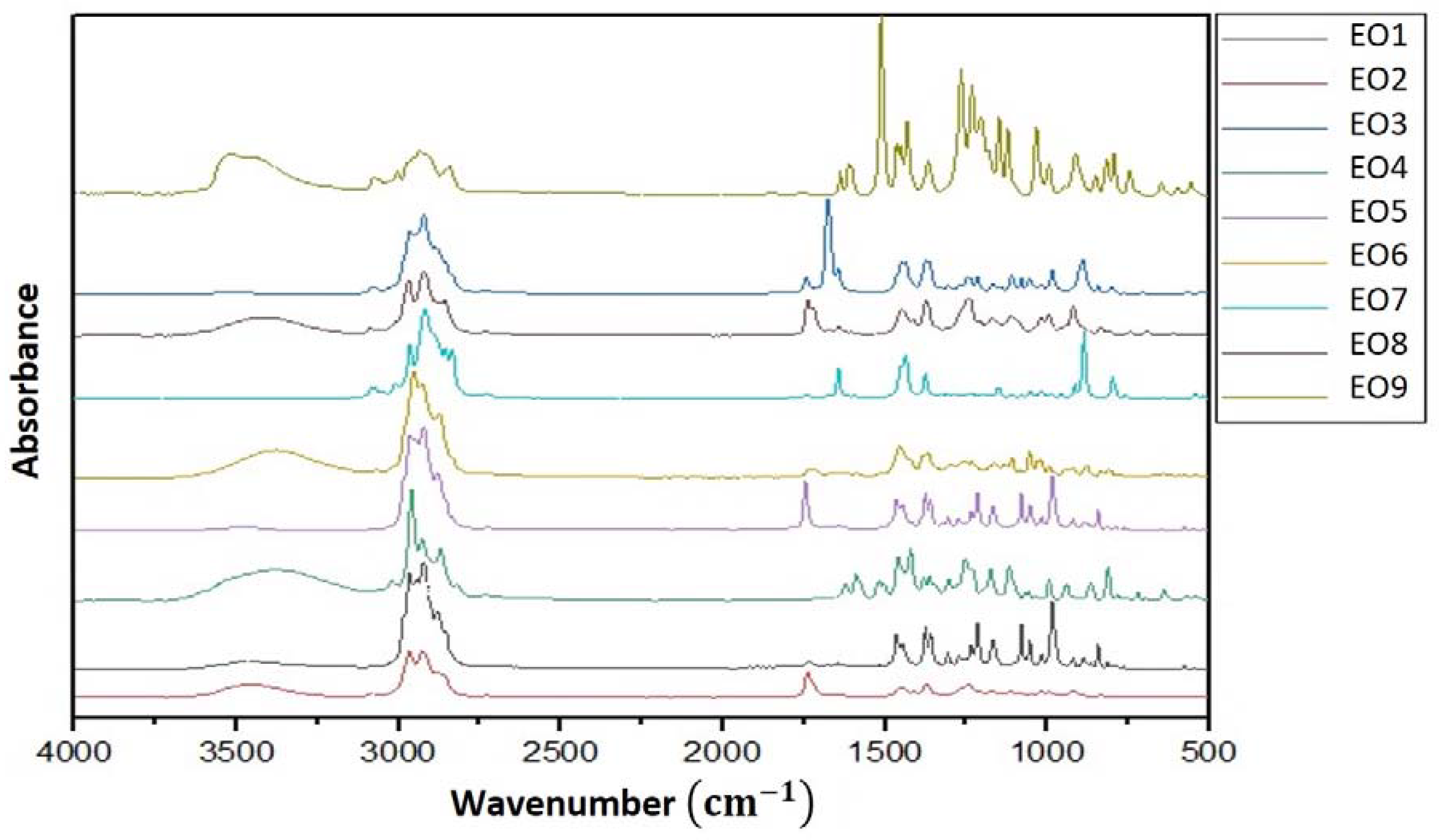
2.2. FT-IR Measurements
2.3. Fungal Material
2.4. Fruit Preparation
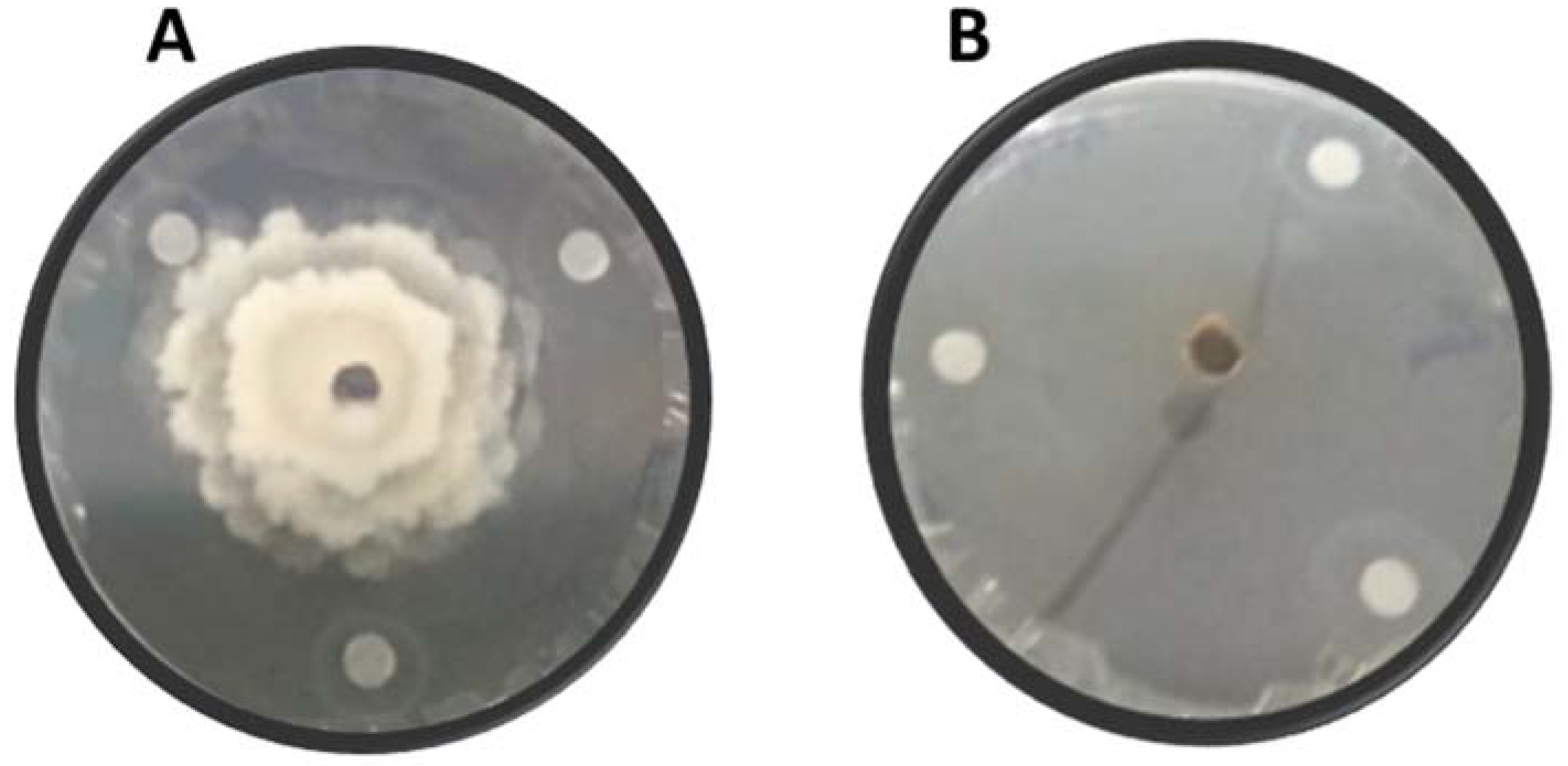
2.5. Antifungal Activity of Essential Oils on Mycelial Growth under In Vitro Conditions
2.6. Effect of Essential Oils on Conidial Germination
2.7. Effect of Essential Oils on Brown Rot Development in Artificially Wounded and Inoculated Fruit
2.8. Efficacy of Essential Oils under Semi-Commercial Conditions
2.9. Quantification of Quality Parameters
2.9.1. Weight Loss
2.9.2. Fruit Firmness and Maturity Index (MI)
2.9.3. Total Soluble Solids
2.9.4. Titratable Acidity
2.10. Statistical Analysis
3. Results
3.1. FT-IR Analysis of the Essential Oils
3.2. Antifungal Activity of Essential Oils on Mycelial Growth under In Vitro Conditions
3.3. Effect of Essential Oils on Spore Germination
3.4. Effect of Essential Oils on Brown Rot Disease Severity
3.5. Semi-Commercial Large-Scale Trial
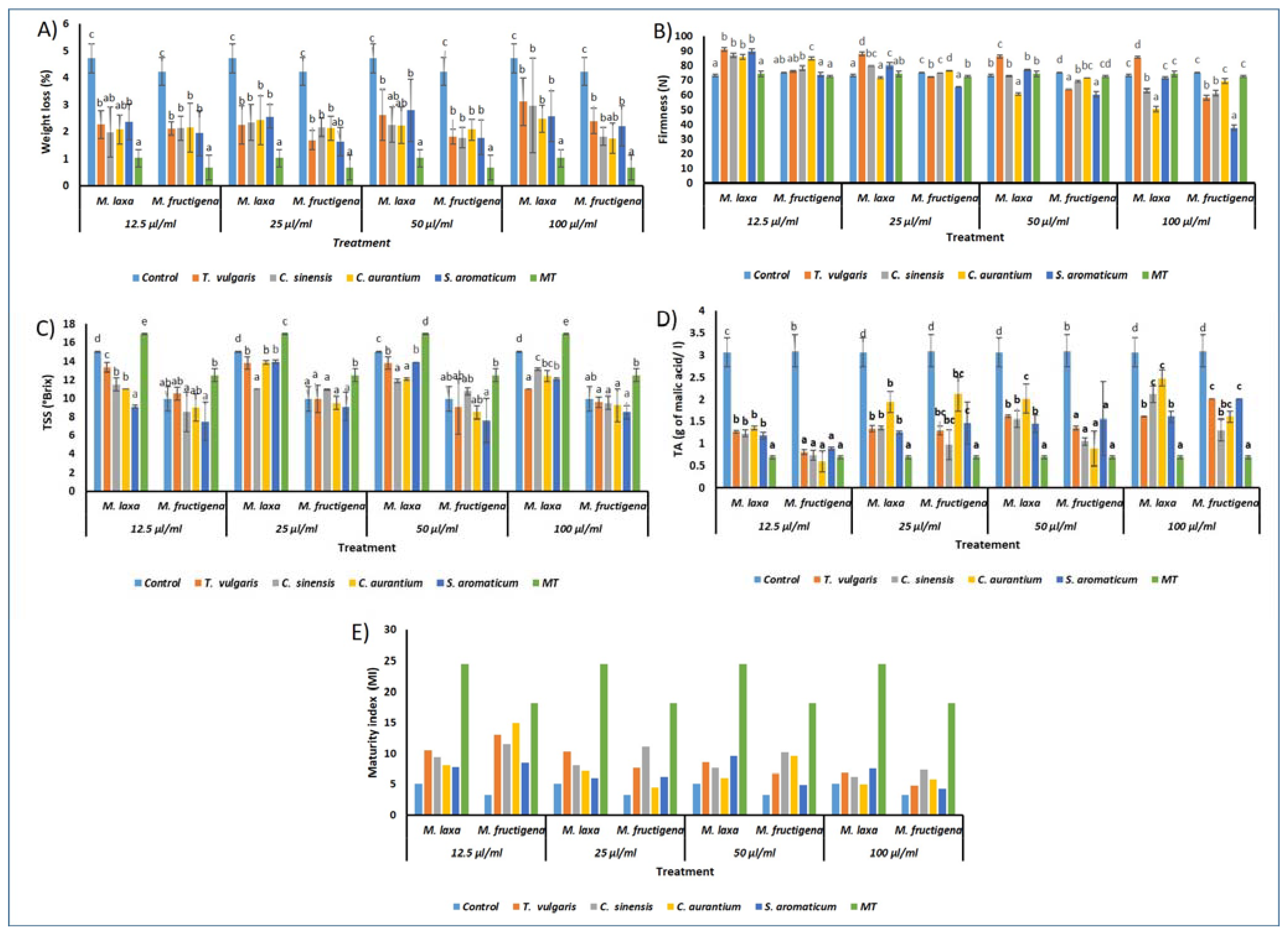
3.6. Effect of Treatments on Fruit Quality Parameters
3.6.1. Weight Loss
3.6.2. Fruit Firmness
3.6.3. Total Soluble Solids
3.6.4. Titratable Acidity
3.6.5. Maturity Index
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cornille, A.; Giraud, T.; Smulders, M.J.M.; Roldán-Ruiz, I.; Gladieux, P. The domestication and evolutionary ecology of apples. Trends Genet. 2014, 30, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Moinina, A.; Lahlali, R.; Boulif, M. Important pests, diseases and weather conditions affecting apple production in Morocco: Current state and perspectives. Rev. Maroc. Des Sci. Agron. Vét. 2019, 7, 71–87. [Google Scholar]
- Barkai-Golan, R. Postharvest Diseases of Fruits and Vegetables: Development and Control; Elsevier: Amsterdam, The Netherlands, 2001; ISBN 0080539297. [Google Scholar]
- Nabila, E.A.; Soufiyan, E.A. Use of Plant Extracts in the Control of Post-Harvest Fungal Rots in Apples. J. Bot. Res. 2019, 2, 31–58. [Google Scholar] [CrossRef]
- Hrustić, J.; Delibašić, G.; Stanković, I.; Grahovac, M.; Krstić, B.; Bulajić, A.; Tanović, B. Monilinia spp. causing brown rot of stone fruit in Serbia. Plant Dis. 2015, 99, 709–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Francesco, A.; Martini, C.; Mari, M. Biological control of postharvest diseases by microbial antagonists: How many mechanisms of action? Eur. J. Plant Pathol. 2016, 145, 711–717. [Google Scholar] [CrossRef]
- Rungjindamai, N.; Jeffries, P.; Xu, X.-M. Epidemiology and management of brown rot on stone fruit caused by Monilinia laxa. Eur. J. Plant Pathol. 2014, 140, 1–17. [Google Scholar] [CrossRef]
- Holb, I.J. Brown rot blossom blight of pome and stone fruits: Symptom, disease cycle, host resistance, and biological control. Int. J. Hortic. Sci. 2008, 14, 15–21. [Google Scholar] [CrossRef]
- Hrustić, J.; Mihajlović, M.; Grahovac, M.; Delibašić, G.; Bulajić, A.; Krstić, B.; Tanović, B. Genus Monilinia on pome and stone fruit species. Pestic. Fitomedicina 2012, 27, 283–297. [Google Scholar] [CrossRef]
- Martini, C.; Mari, M. Monilinia fructicola, Monilinia laxa (Monilinia rot, brown rot). In Postharvest Decay; Elsevier: Amsterdam, The Netherlands, 2014; pp. 233–265. [Google Scholar]
- Egüen, B.; Melgarejo, P.; De Cal, A. The effect of fungicide resistance on the structure of Monilinia laxa populations in Spanish peach and nectarine orchards. Eur. J. Plant Pathol. 2016, 145, 815–827. [Google Scholar] [CrossRef]
- Talibi, I.; Boubaker, H.; Boudyach, E.H.; Ait Ben Aoumar, A. Alternative methods for the control of postharvest citrus diseases. J. Appl. Microbiol. 2014, 117, 1–17. [Google Scholar] [CrossRef]
- Moraes Bazioli, J.; Belinato, J.R.; Costa, J.H.; Akiyama, D.Y.; Pontes, J.G.d.M.; Kupper, K.C.; Augusto, F.; de Carvalho, J.E.; Fill, T.P. Biological Control of Citrus Postharvest Phytopathogens. Toxins 2019, 11, 460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askarne, L.; Talibi, I.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Serghini, M.A.; Ait Ben Aoumar, A. In vitro and in vivo antifungal activity of several Moroccan plants against Penicillium italicum, the causal agent of citrus blue mold. Crop Prot. 2012, 40, 53–58. [Google Scholar] [CrossRef]
- González-Domínguez, E.; Armengol, J.; Rossi, V. Biology and epidemiology of Venturia species affecting fruit crops: A review. Front. Plant Sci. 2017, 8, 1496. [Google Scholar] [CrossRef] [PubMed]
- Elkhetabi, A.; Lahlali, R.; Askarne, L.; Ezrari, S.; El Ghadaroui, L.; Tahiri, A.; Hrustić, J.; Amiri, S. Efficacy assessment of pomegranate peel aqueous extract for brown rot (Monilinia spp.) disease control. Physiol. Mol. Plant Pathol. 2020, 110, 101482. [Google Scholar] [CrossRef]
- Jijakli, M.H.; Lepoivre, P. State of the art and challenges of post-harvest disease management in apples. In Fruit and Vegetable Diseases; Springer: Berlin/Heidelberg, Germany, 2004; pp. 59–94. [Google Scholar]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Turek, C.; Stintzing, F.C. Stability of essential oils: A review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 40–53. [Google Scholar] [CrossRef]
- Pontes, E.K.U.; Melo, H.M.; Nogueira, J.W.A.; Firmino, N.C.S.; de Carvalho, M.G.; Júnior, F.E.A.C.; Cavalcante, T.T.A. Antibiofilm activity of the essential oil of citronella (Cymbopogon nardus) and its major component, geraniol, on the bacterial biofilms of Staphylococcus aureus. Food Sci. Biotechnol. 2019, 28, 633–639. [Google Scholar] [CrossRef]
- El-Kalamouni, C.; Venskutonis, P.R.; Zebib, B.; Merah, O.; Raynaud, C.; Talou, T. Antioxidant and antimicrobial activities of the essential oil of Achillea millefolium L. grown in France. Medicines 2017, 4, 30. [Google Scholar] [CrossRef] [Green Version]
- Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Hijazi, A.; Cerny, M.; Kanaan, H.; Chokr, A.; Merah, O. Fennel oil and by-products seed characterization and their potential applications. Ind. Crops Prod. 2018, 111, 92–98. [Google Scholar] [CrossRef] [Green Version]
- Merah, O.; Sayed-Ahmad, B.; Talou, T.; Saad, Z.; Cerny, M.; Grivot, S.; Evon, P.; Hijazi, A. Biochemical composition of cumin seeds, and biorefining study. Biomolecules 2020, 10, 1054. [Google Scholar] [CrossRef] [PubMed]
- Hrustić, J.; Mihajlović, M.; Grahovac, M.; Delibašić, G.; Tanović, B. Fungicide sensitivity, growth rate, aggressiveness and frost hardiness of Monilinia fructicola and Monilinia laxa isolates. Eur. J. Plant Pathol. 2018, 151, 389–400. [Google Scholar] [CrossRef]
- Pascual, S. Induction of Conidia Production by Monilinia laxa on Agar Media by Acetone. Phytopathology 1990, 80, 494. [Google Scholar] [CrossRef]
- Talibi, I.; Karim, H.; Askarne, L.; Boubaker, H.; Boudyach, E.H.; Msanda, F.; Saadi, B.; Ait Ben Aoumar, A. Antifungal activity of aqueous and organic extracts of eight aromatic and medicinal plants against Geotrichum candidum, causal agent of citrus sour rot. Int. J. Agron. Plant Prod. 2013, 4, 3510–3521. [Google Scholar]
- Liu, Y.; Chen, Z.; Liu, Y.; Wang, X.; Luo, C.; Nie, Y.; Wang, K. Enhancing bioefficacy of Bacillus subtilis with sodium bicarbonate for the control of ring rot in pear during storage. Biol. Control 2011, 57, 110–117. [Google Scholar] [CrossRef]
- Soylu, E.M.; Kurt, Ş.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol activity and putative mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea agglomerans ACBP1 against brown rot disease of fruit. Microb. Pathog. 2020, 139, 103914. [Google Scholar] [CrossRef]
- Sangwanich, S.; Sangchote, S.; Leelasuphakul, W. Biocontrol of citrus green mould and postharvest quality parameters. Int. Food Res. J. 2013, 20, 3381. [Google Scholar]
- Hassani, A.; Fathi, Z.; Ghosta, Y.; Abdollahi, A.; Meshkatalsadat, M.H.; Marandi, R.J. Evaluation of plant essential oils for control of postharvest brown and gray mold rots on apricot. J. Food Saf. 2012, 32, 94–101. [Google Scholar] [CrossRef]
- Qin, X.; Xiao, H.; Xue, C.; Yu, Z.; Yang, R.; Cai, Z.; Si, L. Biocontrol of gray mold in grapes with the yeast Hanseniaspora uvarum alone and in combination with salicylic acid or sodium bicarbonate. Postharvest Biol. Technol. 2015, 100, 160–167. [Google Scholar] [CrossRef]
- Pobiega, K.; Igielska, M.; Włodarczyk, P.; Gniewosz, M. The use of pullulan coatings with propolis extract to extend the shelf life of blueberry (Vaccinium corymbosum) fruit. Int. J. Food Sci. Technol. 2021, 56, 1013–1020. [Google Scholar] [CrossRef]
- Li, Y.; Kong, D.; Lin, X.; Xie, Z.; Bai, M.; Huang, S.; Nian, H.; Wu, H. Quality evaluation for essential oil of Cinnamomum verum leaves at different growth stages based on GC–MS, FTIR and microscopy. Food Anal. Methods 2016, 9, 202–212. [Google Scholar] [CrossRef]
- Das, A.; Dhinesh Kannan, S.K.; Harshadha, K.; Hari Raj, K.; Bindhu, J. Molecular characterization and evaluation of antimicrobial activity of essential oils derived from Syzygium aromaticum. Int. J. Green Pharm. 2018, 12, 56–61. [Google Scholar]
- Catauro, M.; Bollino, F.; Tranquillo, E.; Sapio, L.; Illiano, M.; Caiafa, I.; Naviglio, S. Chemical analysis and anti-proliferative activity of Campania Thymus Vulgaris essential oil. J. Essent. Oil Res. 2017, 29, 461–470. [Google Scholar] [CrossRef]
- Ostolski, M.; Adamczak, M.; Brzozowski, B.; Stolarski, M.J. Screening of functional compounds in supercritical carbon dioxide extracts from perennial herbaceous crops. Agriculture 2021, 11, 488. [Google Scholar] [CrossRef]
- Boughendjioua, H. Fourier Transformed Infrared Spectroscopy Analysis of Constituents of Lemon Essential Oils from Algeria. Am. J. Opt. Photonics 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Bounaas, K.; Bouzidi, N.; Daghbouche, Y.; Garrigues, S.; de la Guardia, M.; Hattab, M. El Essential oil counterfeit identification through middle infrared spectroscopy K. Microchem. J. 2018, 139, 347–356. [Google Scholar] [CrossRef]
- Rodríguez, J.W.; Peyron, S.; Rigou, P.; Chalier, P. Rapid quantification of clove (Syzygium aromaticum) and spearmint (Mentha spicata) essential oils encapsulated in a complex organic matrix using an ATR-FTIR spectroscopic method. PLoS ONE 2018, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cebi, N.; Taylan, O.; Abusurrah, M.; Sagdic, O. Detection of Orange Essential Oil, Isopropyl Myristate, and Benzyl Alcohol in Lemon Essential Oil by FTIR Spectroscopy Combined with Chemometrics. Foods 2020, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Pateiro, M.; Munekata, P.E.S.; Sant’Ana, A.S.; Domínguez, R.; Rodríguez-Lázaro, D.; Lorenzo, J.M. Application of essential oils as antimicrobial agents against spoilage and pathogenic microorganisms in meat products. Int. J. Food Microbiol. 2021, 337, 108966. [Google Scholar] [CrossRef] [PubMed]
- Koum, O.; Walla, S.; DhaliwalL, G.S. Essential Oils as Green Pesticides: Potential and Constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Plant essential oils for pest and disease management. Crop Prot. 2000, 19, 603–608. [Google Scholar] [CrossRef]
- Basaid, K.; Chebli, B.; Mayad, E.H.; Furze, J.N.; Krier, F.; Barakate, M.; Paulitz, T.; Basaid, K.; Chebli, B.; Mayad, E.H.; et al. Biological activities of essential oils and lipopeptides applied to control plant pests and diseases: A review. Int. J. Pest Manag. 2020, 67, 1–23. [Google Scholar] [CrossRef]
- Reddy, S.G.E.; Dolma, S.K. Acaricidal activities of essential oils against two-spotted spider mite, Tetranychus urticae Koch. Toxin Rev. 2018, 37, 62–66. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Glowniak-Lipa, A.; Ludwiczuk, A.; Baj, T.; Malm, A. The in vitro activity of essential oils against Helicobacter pylori growth and urease activity. Molecules 2020, 25, 586. [Google Scholar] [CrossRef] [Green Version]
- Serra, E.; Hidalgo-Bastida, L.A.; Verran, J.; Williams, D.; Malic, S. Antifungal activity of commercial essential oils and biocides against Candida albicans. Pathogens 2018, 7, 15. [Google Scholar] [CrossRef] [Green Version]
- Palfi, M.; Konjevoda, P.; Vrandečić, K. Antifungal activity of essential oils on mycelial growth of Fusarium oxysporum and Bortytis cinerea. Emir. J. Food Agric. 2019, 31, 544–554. [Google Scholar] [CrossRef]
- Tzortzakis, N.G. Impact of cinnamon oil-enrichment on microbial spoilage of fresh produce. Innov. Food Sci. Emerg. Technol. 2009, 10, 97–102. [Google Scholar] [CrossRef]
- Lawson, S.K.; Sharp, L.G.; Powers, C.N.; McFeeters, R.L.; Satyal, P.; Setzer, W.N. Volatile compositions and antifungal activities of native american medicinal plants: Focus on the asteraceae. Plants 2020, 9, 126. [Google Scholar] [CrossRef] [Green Version]
- Rezgui, M.; Majdoub, N.; Mabrouk, B.; Baldisserotto, A.; Bino, A.; Ben Kaab, L.B.; Manfredini, S. Antioxidant and antifungal activities of marrubiin, extracts and essential oil from Marrubium vulgare L. against pathogenic dermatophyte strains. J. Mycol. Med. 2020, 30, 100927. [Google Scholar] [CrossRef]
- Neri, F.; Mari, M.; Brigati, S.; Bertolini, P. Fungicidal activity of plant volatile compounds for controlling Monilinia laxa in stone fruit. Plant Dis. 2007, 91, 30–35. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Reyes, J.G.; Spadaro, D.; Prelle, A.; Garibaldi, A.; Gullino, M.L. Efficacy of plant essential oils on postharvest control of rots caused by fungi on different stone fruits in vivo. J. Food Prot. 2013, 76, 631–639. [Google Scholar] [CrossRef]
- Carovic-Stanko, K.; Fruk, G.; Satovic, Z.; Ivic, D.; Politeo, O.; Sever, Z.; Grdiša, M.; Strikic, F.; Jemric, T. Effects of Ocimum spp. essential oil on Monilinia laxa in vitro. J. Essent. Oil Res. 2013, 25, 143–148. [Google Scholar] [CrossRef]
- Yılar, M.; Bayan, Y. Antifungal Activity of Thymbra spicata L. and Rosmarinus officinalis L. Essential Oils against Monilinia fructigena Honey in Whetze. Türk Tarım Doğa Bilim. Derg. 2018, 5, 121–126. [Google Scholar] [CrossRef] [Green Version]
- Santoro, K.; Maghenzani, M.; Chiabrando, V.; Bosio, P.; Gullino, M.L.; Spadaro, D.; Giacalone, G. Thyme and savory essential oil vapor treatments control brown rot and improve the storage quality of peaches and nectarines, but could favor gray mold. Foods 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moghaddam, M.; Mehdizadeh, L. Chemical Composition and Antifungal Activity of Essential Oil of Thymus vulgaris Grown in Iran against Some Plant Pathogenic Fungi. J. Essent. Oil-Bear. Plants 2020, 23, 1072–1083. [Google Scholar] [CrossRef]
- Fathi, Z.; Hassani, A.; Ghosta, Y.; Abdollahi, A.; Meshkatalsadat, M.H. The potential of thyme, clove, cinnamon and ajowan essential oils in inhibiting the growth of Botrytis cinerea and Monilinia fructicola. J. Essent. Oil-Bear. Plants 2012, 15, 38–47. [Google Scholar] [CrossRef]
- Sameza, M.L.; Nguemnang Mabou, L.C.; Tchameni, S.N.; Boat Bedine, M.A.; Tchoumbougnang, F.; Jazet Dongmo, P.M.; Boyom Fekam, F. Evaluation of clove essential oil as a mycobiocide against Rhizopus stolonifer and Fusarium solani, tuber rot causing fungi in yam (Dioscorea rotundata Poir.). J. Phytopathol. 2016, 164, 433–440. [Google Scholar] [CrossRef]
- Hashem, M.; Alamri, S.A.M.; Alqahtani, M.S.A.; Alshehri, S.R.Z. A multiple volatile oil blend prolongs the shelf life of peach fruit and suppresses postharvest spoilage. Sci. Hortic. 2019, 251, 48–58. [Google Scholar] [CrossRef]
- Amiri, A.; Dugas, R.; Pichot, A.L.; Bompeix, G. In vitro and in vivo activity of eugenol oil (Eugenia caryophylata) against four important postharvest apple pathogens. Int. J. Food Microbiol. 2008, 126, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, J.; Chen, H.; Fan, Y.; Shi, Z. Antifungal activity of eugenol against Botrytis cinerea. Trop. Plant Pathol. 2010, 35, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Trabelsi, D.; Hamdane, A.M.; Ben Said, M.; Abdrrabba, M. Chemical Composition and Antifungal Activity of Essential Oils from Flowers, Leaves and Peels of Tunisian Citrus aurantium Against Penicillium digitatum and Penicillium italicum. J. Essent. Oil-Bear. Plants 2016, 19, 1660–1674. [Google Scholar] [CrossRef]
- Regnier, T.; Combrinck, S.; Veldman, W.; Du Plooy, W. Application of essential oils as multi-target fungicides for the control of Geotrichum citri-aurantii and other postharvest pathogens of citrus. Ind. Crops Prod. 2014, 61, 151–159. [Google Scholar] [CrossRef]
- Kačániová, M.; Terentjeva, M.; Galovičová, L.; Ivanišová, E.; Štefániková, J.; Valková, V.; Borotová, P.; Kowalczewski, P.Ł.; Kunová, S.; Felšöciová, S. Biological activity and antibiofilm molecular profile of Citrus aurantium essential oil and its application in a food model. Molecules 2020, 25, 3956. [Google Scholar] [CrossRef]
- Casquete, R.; Castro, S.M.; Martín, A.; Ruiz-Moyano, S.; Saraiva, J.A.; Córdoba, M.G.; Teixeira, P. Evaluation of the effect of high pressure on total phenolic content, antioxidant and antimicrobial activity of citrus peels. Innov. Food Sci. Emerg. Technol. 2015, 31, 37–44. [Google Scholar] [CrossRef]
- Viuda, M.; Ruiz, Y.; Fernandez-Lopez, J.; Perez, A. Antifungal activity of lemon (Citrus lemon L.), mandarin (C. reticulata L.), grapefruit (C. paridisi L) and orange (C. sinensis) vs. essential oils. Food Saf. 2008, 28, 567–576. [Google Scholar] [CrossRef]
- Rezende, J.L.; Fernandes, C.C.; Costa, A.O.M.; Santos, L.S.; Vicente, F.; Sperandio, E.M.; Souchie, E.L.; Colli, A.C.; Crotti, A.E.M.; Miranda, M.L.D. Antifungal potential of essential oils from two varieties of Citrus sinensis (lima orange and bahia navel orange) in postharvest control of Rhizopus stolonifer (Ehrenb.: Fr.) Vuill. Food Sci. Technol. 2020, 40, 405–409. [Google Scholar] [CrossRef]
- De Clerck, C.; Maso, S.D.; Parisi, O.; Dresen, F.; Zhiri, A.; Haissam Jijakli, M. Screening of antifungal and antibacterial activity of 90 commercial essential oils against 10 pathogens of agronomical importance. Foods 2020, 9, 1418. [Google Scholar] [CrossRef]
- Ammad, F.; Moumen, O.; Gasem, A.; Othmane, S.; Hisashi, K.-N.; Zebib, B.; Merah, O. The potency of lemon (Citrus limon L.) essential oil to control some fungal diseases of grapevine wood. C. R. Biol. 2018, 341, 97–101. [Google Scholar] [CrossRef]
- Lazar-Baker, E.E.; Hetherington, S.D.; Ku, V.V.; Newman, S.M. Evaluation of commercial essential oil samples on the growth of postharvest pathogen Monilinia fructicola (G. Winter) Honey. Lett. Appl. Microbiol. 2011, 52, 227–232. [Google Scholar] [CrossRef]
- Badaway, M.E.I.; Abdelgaleil, S.A.M. Composition and antimicrobial activity of essential oils isolated from Egyptian plants against plan pathogenic bacteria and fungi. Ind. Crops Prod. 2014, 52, 776–782. [Google Scholar] [CrossRef]
- Phillips, C.A.; Laird, K.; Allen, S.C. The use of Citri-VTM®—An antimicrobial citrus essential oil vapour for the control of Penicillium chrysogenum, Aspergillus niger and Alternaria alternata in vitro and on food. Food Res. Int. 2012, 47, 310–314. [Google Scholar] [CrossRef]
- Sharma, N.; Prakash, S.M. Integrated management of soybean (Glycine× max L. Merr.) By essential oil of Citrus× sinensis L. cv. ’osbeck’ epicarp in postharvest. JOP 2012, 2, 33–44. [Google Scholar]
- Elshafie, H.S.; Mancini, E.; Camele, I.; De Martino, L.; De Feo, V. In vivo antifungal activity of two essential oils from Mediterranean plants against postharvest brown rot disease of peach fruit. Ind. Crops Prod. 2015, 66, 11–15. [Google Scholar] [CrossRef]
- Gebel, M.P.; Magurno, F. Assessment of the antifungal potential of the essential oil from Thymus vulgaris against Botrytis cinerea causative agent of postharvest grey mould on strawberry fruits. Columella J. Agric. Environ. Sci. 2014, 2, 17–24. [Google Scholar] [CrossRef]
- Vitoratos, A.; Bilalis, D.; Karkanis, A.; Efthimiadou, A. Antifungal Activity of Plant Essential Oils Against Botrytis cinerea, Penicillium italicum and Penicillium digitatum. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Barrera-Necha, L.L.; Bautista-Baños, S.; Flores-Moctezuma, H.E.; Estudillo, A.R. Efficacy of essential oils on the conidial germination, growth of Colletotrichum gloeosporioides (Penz.) Penz. and Sacc and control of postharvest diseases in papaya (Carica papaya L.). Plant Pathol. J. 2008, 5, 174–178. [Google Scholar] [CrossRef] [Green Version]
- Abdolahi, A.; Hassani, A.; Ghosta, Y.; Bernousi, I.; Meshkatalsadat, M. Study on the potential use of essential oils for decay control and quality preservation of tabarzeh table grape. J. Plant Prot. Res. 2010, 50, 45–52. [Google Scholar] [CrossRef]
- Ranasinghe, L.; Jayawardena, B.; Abeywickrama, K. An integrated strategy to control post-harvest decay of Embul banana by combining essential oils with modified atmosphere packaging. Int. J. Food Sci. Technol. 2005, 40, 97–103. [Google Scholar] [CrossRef]
- Azarakhsh, N.; Osman, A.; Ghazali, H.M.; Tan, C.P.; Mohd Adzahan, N. Lemongrass essential oil incorporated into alginate-based edible coating for shelf-life extension and quality retention of fresh-cut pineapple. Postharvest Biol. Technol. 2014, 88, 1–7. [Google Scholar] [CrossRef]
- Rojas-Graü, M.A.; Raybaudi-Massilia, R.M.; Soliva-Fortuny, R.C.; Avena-Bustillos, R.J.; McHugh, T.H.; Martín-Belloso, O. Apple puree-alginate edible coating as carrier of antimicrobial agents to prolong shelf-life of fresh-cut apples. Postharvest Biol. Technol. 2007, 45, 254–264. [Google Scholar] [CrossRef]
- Zarrad, K.; Ben Hamouda, A.; Chaieb, I.; Laarif, A.; Jemâa, J.M. Ben Chemical composition, fumigant and anti-acetylcholinesterase activity of the Tunisian Citrus aurantium L. essential oils. Ind. Crops Prod. 2015, 76, 121–127. [Google Scholar] [CrossRef]
- Moreno, J.R.A.; Urena, F.P.; Gonzalez, J.J.L. Conformational landscape in chiral terpenes from vibrational spectroscopy and quantum chemical calculations: S-(+)-carvone. Vib. Spectrosc. 2009, 51, 318–325. [Google Scholar] [CrossRef]
- Hayat, J.; Akodad, M.; Moumen, A.; Baghour, M.; Skalli, A.; Ezrari, S.; Belmalha, S. Phytochemical screening, polyphenols, flavonoids and tannin content, antioxidant activities and FTIR characterization of Marrubium vulgare L. from 2 different localities of Northeast of Morocco. Heliyon 2020, 6, e05609. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Plant Species | Inhibition of Mycelial Growth (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Essential Oil Concentration (μL/mL) | ||||||||
| 12.5 | 25 | 50 | 100 | |||||
| M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | M. Laxa | M. fructigena | |
| E. radiata | 45.14 ± 1.14 a | 48.96 ± 0.78 d | 49.94 ± 1.60 a | 54.36 ± 0.60 b | 54.27 ± 1.71 a | 56.06 ± 0.72 a | 71.91 ± 1.06 d | 63.69 ± 0.59 a |
| L. angustifolia | 46.91 ± 0.99 b | 44.92 ± 0.95 a | 50.51 ± 1.56 a | 51.74 ± 0.67 a | 54.44 ± 1.13 a | 56.53 ± 0.74 a | 66.92 ± 1.50 b | 71.19 ± 0.71 c |
| M. pulegium | 45.28 ± 0.78 a | 45.91 ± 1.28 b | 49.49 ± 1.29 a | 54.13 ± 0.81 b | 54.34 ± 0.84 b | 58.96 ± 0.45 b | 80.06 ± 0.78 e | 80.33 ± 0.46 d |
| O. compactum | 44.64 ± 0.63 a | 47.51 ± 1.36 c | 51.95 ± 0.57 b | 51.53 ± 1.89 a | 60.79 ± 0.74 c | 61.53 ± 0.38 c | 68.81 ± 0.69 c | 68.67 ± 0.63 b |
| R. officinalis | 48.69 ± 1.77 e | 47.02 ± 0.44 c | 51.77 ± 0.67 b | 54.15 ± 0.62 b | 56.45 ± 0.85 b | 58.66 ± 0.72 b | 63.89 ± 1.07 a | 63.76 ± 0.23 a |
| T. vulgaris | 50.92 ± 1.94 d | 52.14 ± 0.52 e | 61.38 ± 1.76 c | 64.00 ± 0.34 c | 72.34 ± 1.14 d | 77.85 ± 0.50 e | 94.81 ± 1.49 g | 92.51 ± 0.32 f |
| C. sinensis | 53.96 ± 1.12 e | 54.29 ± 0.32 f | 65.12 ± 0.91 d | 66.79 ± 1.06 d | 75.9 ± 1.11 e | 75.69 ± 0.41 d | 91.78 ± 1.49 f | 92.53 ± 0.49 f |
| C. aurantium | 53.72 ± 1.28 e | 54.53 ± 0.28 f | 68.56 ± 1.27 e | 68.77 ± 0.21 e | 75.63 ± 0.67 e | 75.56 ± 0.29 d | 92.35 ± 1.34 f | 91.4 ± 1.19 e |
| S. aromaticum | 64.02 ± 0.47 f | 63.98 ± 0.33 g | 73.05 ± 0.98 f | 73.21 ± 0.79 f | 82.59 ± 1.09 f | 82.91 ± 0.56 f | 94.69 ± 0.54 g | 94.73 ± 0.52 g |
| MT | 100 ± 0.00 g | 100 ± 0.00 h | 100 ± 0.00 g | 100 ± 0.00 g | 100 ± 0.00 g | 100 ± 0.00 g | 100 ± 0.00 h | 100 ± 0.00 h |
| Plant Species | EC50 (µL/mL) | |||
|---|---|---|---|---|
| Mycelial Growth | Spore Germination | |||
| M. laxa | M. fructigena | M. laxa | M. fructigena | |
| E. radiate | 20.80 | 15.34 | 73.52 | 69.62 |
| L. angustifolia | 21.23 | 21.28 | 63.36 | 53.03 |
| M. pulegium | 21.43 | 18.87 | 37.15 | 36.19 |
| O. compactum | 20.20 | 17.75 | 58.91 | 43.09 |
| R. officinalis | 17.30 | 16.79 | 23.98 | 40.85 |
| T. vulgaris | 14.38 | 12.52 | 13.55 | 15.98 |
| C. sinensis | 11.52 | 11.19 | 17.64 | 15.11 |
| C. aurantium | 10.96 | 10.36 | 14.12 | 9.99 |
| S. aromaticum | 6.74 | 10.09 | 7.80 | 6.00 |
| Plant Species | Inhibition of Spore Germination (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Essential Oil Concentrations (μL/mL) | ||||||||
| 12.5 | 25 | 50 | 100 | |||||
| M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | |
| E. radiata | 11.5 ± 2.12 a | 20 ± 1.41 a | 21.5 ± 0.71 a | 28.5 ± 2.12 a | 40.5 ± 3.54 a | 44 ± 1.41 a | 58.5 ± 2.12 a | 57.5 ± 0.71 a |
| L. angustifolia | 18 ± 1.41 ab | 23.5 ± 2.12 a | 33.5 ± 4.95 b | 34.5 ± 2.12 ab | 44.5 ± 2.12 a | 45.5 ± 3.54 ab | 59 ± 0.00 a | 65 ± 1.41 ab |
| M. pulegium | 25 ± 0.00 bc | 28 ± 0.00 ab | 39 ± 1.41 bc | 43.5 ± 2.12 b | 58.5 ± 2.12 b | 54.5 ± 3.54 b | 73 ± 2.83 b | 72 ± 4.24 b |
| O. compactum | 29.5 ± 0.71 c | 33.5 ± 0.71 b | 38 ± 0.00 bc | 43.5 ± 2.12 b | 49 ± 4.24 ab | 53 ± 1.41 ab | 56.5 ± 0.71 a | 60 ± 4.24 ab |
| R. officinalis | 23 ± 1.41 bc | 28 ± 4.24 ab | 44.5 ± 2.12 c | 42.5 ± 4.95 b | 50 ± 1.41 ab | 51.5 ± 3.54 ab | 65 ± 2.83 ab | 67.5 ± 2.12 ab |
| T. vulgaris | 51.60 ± 3.38 e | 52 ± 0.00 c | 65 ± 2.83 d | 59 ± 1.41 c | 73 ± 2.83 c | 70.5 ± 2.12 c | 96 ± 5.66 c | 100 ± 0.00 d |
| C. sinensis | 42 ± 1.14 d | 47.5 ± 2.12 c | 61.5 ± 2.12 d | 63.5 ± 3.54 cd | 77.5 ± 2.12 c | 74 ± 1.41 c | 100 ± 0.00 c | 95.5 ± 6.37 c |
| C. aurantium | 45.5 ± 3.54 de | 54.5 ± 4.95 c | 66.5 ± 4.95 de | 74.5 ± 6.37 de | 88 ± 1.41 d | 89.5 ± 2.12 d | 96 ± 5.66 c | 96.5 ± 4.95 c |
| S. aromaticum | 68.5 ± 2.12 f | 72.5 ± 0.71 d | 76.5 ± 2.12 e | 81.5 ± 4.54 e | 91 ± 1.41 d | 95 ± 1.41 de | 100 ± 0.00 c | 100 ± 0.00 c |
| MT | 100 ± 0.00 g | 100 ± 0.00 e | 100 ± 0.00 f | 100 ± 0.00 f | 100 ± 0.00 e | 100 ± 0.00 e | 100 ± 0.00 d | 100 ± 0.00 c |
| Plant Species | Disease Severity (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Essential Oil Concentrations (μL/mL) | ||||||||
| 12.5 | 25 | 50 | 100 | |||||
| M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | |
| T. vulgaris | 56.39 ± 0.38 b | 31.81 ± 0.33 b | 32.48 ± 0.61 b | 21.30 ± 0.27 b | 11.45 ± 0.42 c | 9.77 ± 0.39 e | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| C. sinensis | 76.25 ± 0.83 d | 60.41 ± 0.32 e | 49.47 ± 0.79 c | 32.14 ± 0.74 d | 8.91 ± 0.46 b | 5.86 ± 0.14 c | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| C. aurantium | 74.73 ± 0.95 d | 54.43 ± 0.03 d | 57.03 ± 0.74 d | 35.67 ± 0.51 e | 12.77 ± 0.04 d | 7.28 ± 0.24 d | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| S. aromaticum | 71.88 ± 1.14 c | 50.69 ± 0.55 c | 48.25 ± 0.88 c | 27.75 ± 0.60 c | 8.39 ± 0.35 b | 3.44 ± 0.09 b | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| MT | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a | 0.00 ± 0.00 a |
| Plant Species | Incidence (%) | |||||||
|---|---|---|---|---|---|---|---|---|
| Essential Oils Concentrations (μL/mL) | ||||||||
| 12.5 | 25 | 50 | 100 | |||||
| M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | M. laxa | M. fructigena | |
| Control | 100 ± 0.00 b | 97.5 ± 3.54 b | 100 ± 0.00 d | 97.5 ± 3.54 e | 100 ± 0.00 d | 97.5 ± 3.54 c | 100 ± 0.00 d | 97.5 ± 3.54 d |
| T. vulgaris | 100 ± 0.00 b | 100 ± 0.00 b | 67.5 ± 3.54 b | 72.5 ± 0.3.54 bc | 27.5 ± 3.54 a | 37.5 ± 3.54 a | 2.5 ± 3.54 a | 0.00 ± 0.00 a |
| C. sinensis | 92.5 ± 3.54 b | 100 ± 0.00 b | 87.5 ± 3.54 cd | 90 ± 0.00 de | 80 ± 0.00 c | 62.5 ± 3.54 b | 42.5 ± 3.54 c | 57.5 ± 3.54 c |
| C. aurantium | 100 ± 0.00 b | 95 ± 0.00 b | 62.5 ± 3.54 b | 60 ± 0.00 b | 35 ± 0.00 ab | 47.5 ± 3.54 ab | 7.5 ± 3.54 a | 30 ± 0.00 b |
| S. aromaticum | 100 ± 0.00 b | 97.5 ± 3.54 b | 82.5 ± 3.54 c | 77.5 ± 3.54 cd | 42.5 ± 3.54 b | 42.5 ± 3.54 ab | 2.5 ± 3.54 a | 27.5 ± 3.54 b |
| MT | 27.5 ± 3.54 a | 32.5 ± 3.54 a | 27.5 ± 3.54 a | 32.5 ± 3.54 a | 27.5 ± 3.54 a | 32.5 ± 3.54 a | 27.5 ± 3.54 b | 32.5 ± 3.54 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El Khetabi, A.; Ezrari, S.; El Ghadraoui, L.; Tahiri, A.; Ait Haddou, L.; Belabess, Z.; Merah, O.; Lahlali, R. In Vitro and In Vivo Antifungal Activities of Nine Commercial Essential Oils against Brown Rot in Apples. Horticulturae 2021, 7, 545. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7120545
El Khetabi A, Ezrari S, El Ghadraoui L, Tahiri A, Ait Haddou L, Belabess Z, Merah O, Lahlali R. In Vitro and In Vivo Antifungal Activities of Nine Commercial Essential Oils against Brown Rot in Apples. Horticulturae. 2021; 7(12):545. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7120545
Chicago/Turabian StyleEl Khetabi, Assia, Said Ezrari, Lahsen El Ghadraoui, Abdessalem Tahiri, Lhoussain Ait Haddou, Zineb Belabess, Othmane Merah, and Rachid Lahlali. 2021. "In Vitro and In Vivo Antifungal Activities of Nine Commercial Essential Oils against Brown Rot in Apples" Horticulturae 7, no. 12: 545. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae7120545