
Interactions of Gibberellins with Phytohormones and Their Role in Stress Responses


Instituto de Investigaciones Agrobiológicas de Galicia, Consejo Superior de Investigaciones Científicas, 15780 Santiago de Compostela, Spain
*
Author to whom correspondence should be addressed.
Horticulturae 2022, 8(3), 241; https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8030241
Submission received: 9 February 2022
/
Revised: 7 March 2022
/
Accepted: 8 March 2022
/
Published: 10 March 2022
(This article belongs to the Topic Plant Breeding, Genetics and Genomics)
Abstract
:Gibberellins are amongst the main plant growth regulators. Discovered over a century ago, the interest in gibberellins research is growing due to their current and potential applications in crop production and their role in the responses to environmental stresses. In the present review, the current knowledge on gibberellins’ homeostasis and modes of action is outlined. Besides this, the complex interrelations between gibberellins and other plant growth regulators are also described, providing an intricate network of interactions that ultimately drives towards precise and specific gene expression. Thus, genes and proteins identified as being involved in gibberellin responses in model and non-model species are highlighted. Furthermore, the molecular mechanisms governing the gibberellins’ relation to stress responses are also depicted. This review aims to provide a comprehensive picture of the state-of-the-art of the current perceptions of the interactions of gibberellins with other phytohormones, and their responses to plant stresses, thus allowing for the identification of the specific mechanisms involved. This knowledge will help us to improve our understanding of gibberellins’ biology, and might help increase the biotechnological toolbox needed to refine plant resilience, particularly under a climate change scenario.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Plant development, from the embryo stage to senescence, is under the control of intricate genetic programs that precisely unfold over the life of the plant. However, as sessile organisms, plants need to adapt to the constant changes in environmental conditions to assure their survival. This need has led to the development of a highly complex repertoire of molecular mechanisms that help them to adapt to and overcome different types of stress or unfavorable conditions, thus increasing plant resilience. In fact, under the current climate change scenario, the ability of plants to survive limiting or harsh conditions, or biological threats, will be more determinant than ever before for agricultural practices. The improvement of our understanding of the mechanisms underlying plant stress responses will help in the development of optimized crop production.
Plant growth regulators (PGRs), classically referred to as phytohormones, are a group of chemically diverse compounds that govern or influence both plant developmental programs and responses to inner and outer cues at minute concentrations. Hormone homeostasis (synthesis, transport, degradation) is tightly regulated, as minor variations in PGRs levels in the different tissues have drastic effects on plant responses, because of their role in controlling gene expression.
Research over the last few years has provided a clear picture of the different processes influenced by every PGR family, with the so-called classical hormones (auxins, gibberellins, cytokinins, abscisic acid, ethylene) attracting most efforts because of their direct impact on many relevant plant traits. However, it is becoming increasingly clear that PGRs’ interactions and specific ratios are key factors controlling plant responses in precise moments of development to distinct types of stress, at both the tissue and the whole plant level.
Gibberellins (GAs) are a set of PGRs that were discovered more than a century ago. Since then, more than a hundred different gibberellins have been described in plants, fungi and bacteria (reviewed in [1]). GAs have been classically implicated in a few crucial physiological responses, such as the regulation of plant stature and seed dormancy. Nonetheless, their relevance extends beyond their effect on stem elongation, germination and flowering, as they play a major role in the regulation of stress tolerance.
In the present review, an up-to-date and comprehensive overview of GAs biology is given. This focuses on the crosstalk with other PGRs and the molecular mechanisms involved in the signaling pathways of these interactions. Besides this, the distinct effects of GAs in response to stress are shown, paying attention to the identity of the genes and proteins involved in such processes.
2. Biochemical and Molecular Aspects
2.1. Biosynthesis and Homeostasis
GAs are acid diterpenoids synthesized from geranylgeranyl diphosphate via the terpenes route, with the formation of ent-kaurene as an intermediate step. There are 136 structurally characterized GAs, but only a few have been shown to be biologically active, mainly GA1, GA3, GA4 and GA7 [2,3]. However, as research advances, new biologically active GAs are being discovered [4]. Based on the number of carbon atoms, GAs are classified into two groups. In general terms, GAs with 20 carbon atoms (C20) lack biological activity, but they can be metabolized to C19, which are usually bioactive [5]. Other chemical modifications in their structure lead to changes in their activity rate, i.e., 3β-hydroxylation improves their bioactivity [6,7], while the 2β-hydroxylation inactivates them [8].
Although a significant part of the GAs synthesis pathway is shared between plants, fungi and bacteria [7], these are not identical, and the differences suggest an independent evolutionary origin [9]. The starting point of GAs synthesis is geranylgeranyl diphosphate (GGDP), which, by means of the enzyme ent-copalyl diphosphate synthase (CPS), is transformed into ent-copalyl diphosphate (ent-CPD), and then into ent-Kaurene by ent-kaurene synthase (KS). The ent-kaurene oxidase (KO) acts in the following steps to obtain ent-Kaurenoic acid, which after two oxidation steps by ent-kaurenoic acid oxidase (KAO) is transformed into GA12, the first GA in the synthesis route. GA12 is not biologically active, but, through the activities of different C20GA-oxidases (i.e., GA-20ox and GA3ox), it can be transformed into other intermediary compounds or active GAs [2,6,10]. Non-biologically active GAs act as precursors or intermediate compounds in the synthesis routes.
The synthesis processes are developed in different cellular compartments. Both GGDP and ent-kaurene are synthesized in the plastid, while the enzyme KAO is located in the endoplasmic reticulum, and the oxidases mentioned above convert GA12 into other GAs in the cytosol [11,12]. Moreover, different organs accumulate different GAs. For example, ovaries present high levels of GA12, GA15 or GA9, but sepals and petals have higher GA4 levels [13].
Different pathways of GAs catabolism regulate the levels of active hormone and avoid over-accumulation. The main form of GA deactivation is through oxidation reactions carried out by specific oxidases (GAox), such as the GA 13-oxidases [14], the oxidases codified by CYP714A1, CYP714A2 and CYP72A [15,16], and the C19-GA 2-oxidase [17]. An alternative route for GA degradation described in rice involves the 16α,17-epoxidation by the cytochrome P450 monooxygenase encoded by Elongated Uppermost Internode (EUI) [18]. A different way to regulate GAs concentration is through the formation of GA conjugates, particularly glucosyl derivatives, in what appears to be a form of GAs storage [19,20]. However, the formation of methyl esters in Arabidopsis was found to be an inactivation method [21].
2.2. Transport
GA location and synthesis varies at the organ and tissue levels. GAs are maintained inside the cells by means of the ion trap mechanism [22]. Weak acids such as GAs can enter the cytoplasm from the extracellular space due to their protonated state in that environment, and therefore have limited traveling ability on their own. Research has shown the relevance of GAs transport through plant tissues, but the involved mechanisms have not been characterized so far. No GA efflux proteins have been identified yet, although some proteins have been shown to import GAs into the cells. The abscisic acid (ABA) transporter ABA-IMPORTER TRANSPORTER 3 (AIT3) is able to import GA3, but it cannot import Jasmonic acid (JA) or Indole-3 acetic acid (IAA) [23]. The Jasmonate-Isoleucine (JA-Ile) importer GLUCOSINOLATE TRANSPORTER 1 (GTR1) imports glucosinolates and GAs [24]. The expression of the GAs importer protein, NITRATE TRANSPORTER 3/NITRATE PEPTIDE FAMILY (NPF3), is promoted by ABA and a lack of nitrogen, and is inhibited by GAs. Other NPF proteins are also able to import different GAs [25]. Besides this, some data suggest that the sugar transporters BIDIRECTIONAL SUGAR TRANSPORTER SWEET (SWEET13/14) have GA transporter abilities [26]. Due to the diversity of GAs in plants, it seems unlikely that a unique transport protein exists. Indeed, the current results suggest that bi- or multi-functional transporters are in charge of the transport across membranes.
The GAs biosynthesis rate varies according to the organ under analysis, and therefore the regulation of plant development requires GAs movement throughout the plant [11]. GAs can be carried in both acropetal and basipetal directions, though the former is believed to be more efficient [11,27,28]. GAs accumulate in the elongation zones during root growth, in an apparently ATP-dependent manner [29]. Nonetheless, combined mutant research and mathematical models in Arabidopsis suggest that GA gradients are tightly coupled with biosynthesis and catabolism to properly drive root growth [30]. In Arabidopsis shoots, GAs are stored in the basal zone of the hypocotyl [31]. It is noteworthy that several studies suggest that precursors [28] and conjugates [19] can also be transported. The GA precursor GA12 can move from the aerial organs to the root through the phloem and vice versa through the xylem [28], in order to modulate plant growth according to environmental temperature [32].
2.3. Signaling and Mode of Action
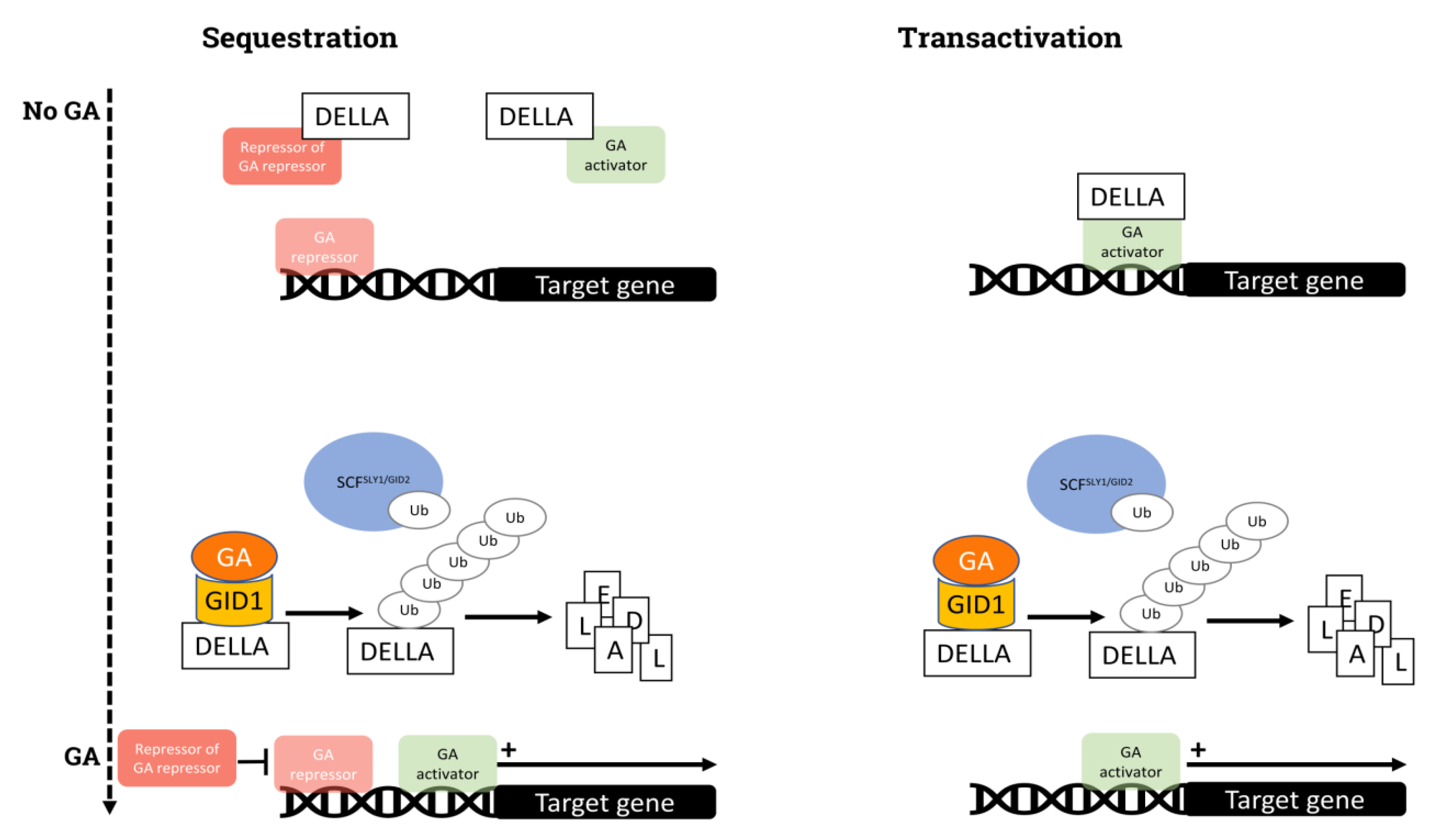
The GAs signaling mechanism and the identity of the involved proteins has been described in model species [33]. Briefly, the DELLA proteins are at the core of GA signaling. DELLA proteins, repressors of GAs signaling characterized by the presence of the DELLA motif (aspartic acid–glutamic acid–leucine–leucine–alanine), are members of the GRAS family (based on the designation of GIBBERELLIC-ACID INSENSITIVE, GAI, REPRESSOR OF GA, RGA, AND SCARECROW, SCR). These proteins bear a C-terminal GRAS domain, a highly conserved N-terminal domain and VHYNP motifs [34]. DELLA proteins function as transcriptional repressors of GA signaling via two main mechanisms: sequestration and transactivation (Figure 1). In the former, DELLA proteins bind to either transcription factors (TFs) or repressors, preventing their activity, while the latter involves the modulation of the expression of downstream genes by the complex formed by the union of DELLA proteins and TFs. Despite their influence in transcriptional processes, DELLA proteins lack DNA-binding domains. Overall, these proteins block the growth-promoting activity of GAs, and many outer and inner factors are integrated in the regulation of their activity.
The presence of GA is perceived by the nuclear receptor GIBBERELLIN INSENSITIVE DWARF 1 (GID1) [35,36,37]. Once the GA–GID complex is formed, the N-terminal domains of the DELLA proteins facilitate the formation of the GA–GID1–DELLA complex [35]. The SLEEPY1 (SLY) proteins [38], or GID2 [39], are F-box subunits of the SCF (Skp1–Cullin 1–F-box) E3 Ubiquitin ligase complex, which mediates the ubiquitination of proteins intended for degradation by 26S proteasome. Once DELLA proteins are bound to GID1 and GAs, the SCFSLY1/GID2 E3 Ubiquitin ligase promotes DELLA polyubiquitination and degradation through the activity of the 26S proteasome [33,40]. The reduction in the levels of DELLAs allows GAs to exert transcriptional modulation.
The activity of the DELLA proteins is subject to post-transcriptional modulations. One of these post-transcriptional modulations is sumoylation, which is a mechanism of protein stabilization. The presence of sumoylated DELLA proteins leads to a sequestration of GID1 receptors, preventing the degradation of non-sumoylated DELLA proteins and, therefore, their accumulation, restraining GA-promoted responses despite the presence of the hormone [41]. However, if the SLY F-box protein is stabilized by sumoylation, then the REPRESSOR OF GA (RGA) DELLA proteins are degraded, enhancing the activity of GAs [42]. These complex interactions portray GA-independent but closely related mechanisms controlling plant responses. Moreover, phosphorylation is important in maintaining DELLA stability and activity [43]. This phosphorylation precludes the formation of the DELLA–GA–GID1 complex, prevents its ubiquitination, and therefore favors the repression of the genes modulated by GAs [44]. The dephosphorylation of DELLA via TYPE-ONE PROTEIN PHOSPHATASE 4 (TOPP4) enhances the DELLA degradation activity driven by GAs [45].
The responses to GAs are influenced by several TFs, which modulate the activity of the DELLA proteins. SCARECROW-LIKE 3 (SCL3), a member of the GRAS family, competes with DELLA proteins for binding to INDETERMINATE DOMAIN (IDD) proteins, and modulates GAs levels through a feedback regulation between the DELLA/IDD and SCL3/IDD complexes [46,47]. GAI-ASSOCIATED FACTOR (GAF1) is a TF that binds DELLA proteins, enabling the expression of the GA biosynthesis genes GA20ox2 and GA3ox1. However, when DELLAs are degraded in the presence of GAs, GAF1 binds to TOPLESS RELATED (TPR) and represses the expression of those same genes [48]. This process depends on GA content, establishing a feedback regulatory mechanism [49]. Nonetheless, other DELLA-independent homeostasis mechanism have been described. The zinc finger TF AXIAL REGULATOR YABBY1 (YAB1), which is expressed in response to GAs, binds to the GA-RESPONSIVE ELEMENT (GARE) domain in the promoter of the GA3ox2 gene, apparently blocking its expression and taking part in the negative feedback regulation of GA synthesis [50]. In addition, OsYABBY4 interacts physically with the DELLA protein SLENDER RICE 1 (SLR1) and binds to the promoter region of GA20ox2, contributing to the modulation of GAs homeostasis [51].
Two different N-acetyl glucosamination processes have been shown to influence GA-related responses. SECRET AGENT (SEC), a positive regulator of the GA response, induces a-GlcNAcylation (O-linked N-acetylglucosamine) of the RGA DELLA protein, preventing its union with several light- and hormone-related proteins and thus maintaining the effect of GAs [52]. On the other hand, the interaction between SWITCH/SUCROSE NONFERMENTING (SWI/SNF), a chromatin-remodeling complex, SPINDLY (SPY), an O-GlNAc transferase, and DELLA negatively regulates GAs signaling [53,54]. The SWIC3 core subunit of SWITCH/SUCROSE NONFERMENTIG has been shown to modulate GA responses by means of chromatin structure regulation [55]. Indeed, epigenetic regulation has been shown to play a major role in GAs transcriptional activity. BRAHMA (BRM), a chromatin remodeling ATPase, induces the expression of genes related to GAs biosynthesis and signaling [56], while PICKLE (PKL), another major chromatin remodeler, modulates the expressions of 80% of the GA-responsive genes [57]. PKL seems to integrate outer and inner cues and hormone signaling pathways (such as GAs and Brassinosteroids (BRs)) to modulate gene expression [58]. Therefore, the growth-promoting activity of GAs is subject to the control of many factors at different levels, ensuring that the plant invests energy in development only under the most favorable conditions.
3. Interactions with PGRs
The relevance of the crosstalk between GAs and other hormones in the control of plant developmental processes has been reported in several species. The responses of tissues to PGRs depend on both the concentration of every growth regulator and the specific ratios between each set of hormones. This framework induces a highly complex scenario influenced by many factors, and particularly by genes involved in the homeostasis of every hormone (synthesis, transport…), which can be negatively or positively modulated by other hormones. Moreover, growth regulators can have synergistic or antagonistic effects on the activity of the same specific genes or proteins, thus adding extra levels of complexity to the processes under study. In the following section, we highlight what is known of the GAs’ interaction with other hormones (Figure 2) and with other potential plant growth regulators.
3.1. Auxins
Auxins, mainly Indole Acetic acid (IAA), govern many aspects of plant growth and development, from the embryo stage to senescence. IAA promotes GA synthesis by activating GA3ox and GA20ox and deactivating GA2ox, as seen in Arabidopsis, rice and pea [59,60,61]. The positive influence of auxins on GA content seems to occur in tissue-specific responses, such as those taking place in the roots of pea plants [62] or during fruit set in tomato [63]. On the other hand, GAs modulate auxin-related genes, although the outcomes of these responses depend on the specific set of GA-induced Auxin Response Factors (ARFs). Whilst they promote hypocotyl elongation via ARF6 and ARF8 [64], during grape parthenocarpy ARF10/16/17 are negative regulated by GA3 through the activity of the microRNAs miR160a/b/c [65].
Furthermore, GAs modify the expressions of several auxin transporters [66]. GA biosynthesis and signaling-deficient mutants in Arabidopsis show a reduced activity of PIN-FORMED (PIN) protein auxin transporters (PIN1, PIN2, PIN3), indicating that GAs are needed for the proper function of PIN proteins, as the wild-type phenotype is restored upon GA treatment [67]. This GA-dependent regulation has biological effects, such as gravitropism modulation in Arabidopsis roots via PIN2 stabilization [67,68] or xylogenesis promotion in Populus by means of PIN1 upregulation [69]. In Eucalyptus roots and stems, exogenous GA3 treatment promotes xylogenesis and alters the expression of genes not only related to GA biosynthesis, but also to auxin and secondary cell wall formation [70]. Xylem differentiation from cambium cells was also reported in hypocotyl cuttings of Pinus radiata seedlings treated with indole-butyric acid (IBA) and GA3 [71].
To further complicate their interplay, auxin content and signaling are also influenced by GAs. In Eucalyptus roots, GAs upregulate SUPERROOT2 (SUR2), which is involved in auxin homeostasis [70], while in Arabidopsis GA3 improves root responses to exogenous IAA through the modulation of auxin transporters (AUX1 and PIN1, PIN2 and PIN3) and signaling, as these effects are not detected in Arabidopsis signaling mutants (tir1-1 and axr1-3) [72].
The interaction between auxins and GAs occurs at many levels, including signaling, metabolism or gene expression, and in many cases in a tissue-specific manner. Although a clear relationship between both hormones cannot be stated, as they share a positive influence on some aspects of development, they seem to present a synergistic character.
3.2. Brassinosteroids
BRs and GAs appear to play a cooperative role in their physiological effects, as well as in GA biosynthesis regulation. The TF BRASSINAZOLE RESISTANT 1 (BZR1) controls BRs responses in plants, and is activated by GAs after DELLA degradation. Similarly, BRs take part in plant responses to light by enhancing the transcriptional activity of PHYTOCHROME INTERACTING FACTORS (PIF) TFs [64,73]. This is a potential intersection between BRs and GAs, since GAs allow the activity of PIF4 [74] and, conversely, PIF4 modulates the expression of GA3ox and GA20ox [75]. The over-expression of the BRs biosynthesis gene DWARF 4 (DWF4) and of GA20ox leads to an increment in the GA levels [76], as well as in the expression of BZR1 and BRI1 EMS SUPPRESSOR 1 (BES1) [77]. BRs cooperate with GAs to modulate plant height in rice through the enhancement of GA synthesis [78]. At physiological levels, BRs collaborate with GAs to promote cell elongation, but at high BRs levels GA biosynthesis is inhibited in rice [79]. Moreover, the inhibitory effect of high BRs levels on GAs biosynthesis is a strategy of some oomycetes to suppress immune responses in rice [80]. Therefore, there seems to be a molecular mechanism by which BRs and GAs exert mutual control over each other’s activity. However, in sunflower, Arabidopsis [81] and pea [82], this BRs-GAs interaction has not been found, suggesting the species- or stage-specific conditioning of these relations.
3.3. Ethylene
The gaseous hormone ethylene and GAs act antagonistically on root growth, as primary root growth is enhanced by GAs and repressed by ethylene. This is an example of their contrary effects on development. This opposite behavior might rely on the ability of ethylene to modulate GAs homeostasis. Several studies have shown the capacity of ethylene to negatively modulate or alter GAs biosynthesis genes in Arabidopsis [83,84]. Moreover, it has been suggested that ethylene regulates both biosynthetic and catabolic GA genes in this species [85]. This type of modulation has also been found in tomato and pea [86,87].
However, in Arabidopsis, GAs–ethylene interaction seems to be crucial for the development of the apical hook. After DELLA degradation, the activation of ETHYLENE INSENSITIVE 3/EIN3-LIKE 1 (EIN3/EIL1) by GAs promotes HOOKLESS1 (HLS1) expression. HSL1 modulates apical hook formation and prevents its premature opening in etiolated seedlings, showing once again the close interaction between light and GAs [88,89]. Besides this, GAs enhance the correct development of the apical hook by modulating PIN3 and PIN7, and promoting ethylene biosynthesis by activating ACC SYNTHASE5/ETO2 (ACS5/ETO2) and ACC SYNTHASE 8 (ACS8) [90]. The interaction of DELLA proteins GIBBERELLIC ACID INSENSITIVE (GAI), RGA and RGA52 with the ethylene response factors RELATED TO APETALA (RAP) RAP2.3 and RAP2.12 blocks their transcriptional activity, thus repressing their own action and enhancing the apical hook opening [91]. Overall, there seems to be a complex relation between GAs and ethylene, which can have contrasting characteristics according to the specific process under study.
3.4. Abscisic Acid
ABA and GAs have a reciprocal antagonist effect in biosynthesis modulation. ABA and GAs balance is critical in different biological processes, but it is particularly relevant for seed dormancy and germination. ABA-INSENSITIVE4 (ABI4) promotes ABA synthesis and GA2ox7 expression, inhibiting seed dormancy [92,93]. In addition, ABA, by means of CHOTTO1 (CHO1), inhibits GA20ox2 and reduces GA levels during seed germination. An inhibitory effect of GA on ABA synthesis has also been reported [94]. Several external cues, such as temperature, water stress and light, are also integrated within the ABA/GA balance to ensure the fine-tuning of seed germination.
3.5. Other PGRs
Jasmonic Acid (JA) plays important roles in several biological processes, but particularly in plant defense and responses to environmental conditions. Due to the different processes GAs and JA govern, they usually show an antagonistic relationship mediated by JASMONATE-ZIM DOMAIN (JAZ) and SLR1 [95]. DELLA proteins interact with the JA signaling machinery, enabling the enhanced activity of one hormone or the other, although some synergistic effects have also been found (reviewed in [96]). In tobacco, Jasmonates reduce GA20ox2 levels, thus lowering GA content [97]. DEFECTIVE ANTHER DEHISCENCE 1 (DAD1), which intervenes in jasmonate metabolism, is upregulated by GAs in Eucalyptus [70]. Interestingly, the interaction of JA/GA seems to present a cooperative nature in the formation of flower tissues, at least in model species such as Arabidopsis and rice (Castro-Camba et al., under review).
The putative interactions between cytokinins and GAs are not clearly defined, mainly due to the lack of precise data, although they seem to act antagonistically in several processes such as phase change [98]. GAs activate different cytokinin catabolic genes in Medicago truncatula [99] and tall fescue [100], resulting in the inhibition of tillering in the latter. On the other hand, the activation of cytokinin receptor CYTOKININ RESPONSE 1 (CRE1) by cytokinins reduces GA levels in M. truncatula [101], whereas SPY, which represses GAs signaling, acts as a positive regulator of cytokinins signaling [102]. However, in Eucalyptus, GAs have been shown to activate ISOPENTENYL TRANSFERASE 3 (IPT3), which intervenes in the metabolism of cytokinin, [70]. Moreover, CKs and GAs act synergistically in the regulation of morphological and physiological traits in Polygonum cuspidatum in response to nitrogen availability, with GAs also integrating light cues. Under high nitrogen conditions, the levels of endogenous CKs and GAs are increased, indicating that both hormones are involved in biomass allocation in response to nitrogen availability [103].
Strigolactones (SLs) were first identified as molecules synthesized by parasitic plants, but were later recognized as endogenous phytohormones. SLs and GAs have similar perception and signaling mechanisms [104,105]. Shoot elongation and tiller bud outgrowth seem to be under the control of the crosstalk between SLs and GAs [106]. However, the outcome of this crosstalk might depend on the process under analysis. SLs are required for the interaction between DWARF 14 (D14) and SLR1 proteins, negatively regulating GA signaling [107]. On the other hand, GA signaling represses SLs biosynthesis genes [108]. Bud outgrowth in Jatropha curcas is promoted by GAs but inhibited by SLs [109]. However, SLs modulate ABA/GA ratio, lowering ABA synthesis and increasing GA accumulation during the germination of thermo-inhibited seeds [110], while SLs regulate shoot elongation in rice, influencing GA metabolism and signaling [111].
3.6. Interactions with Potential PGRs or Relevant Compounds
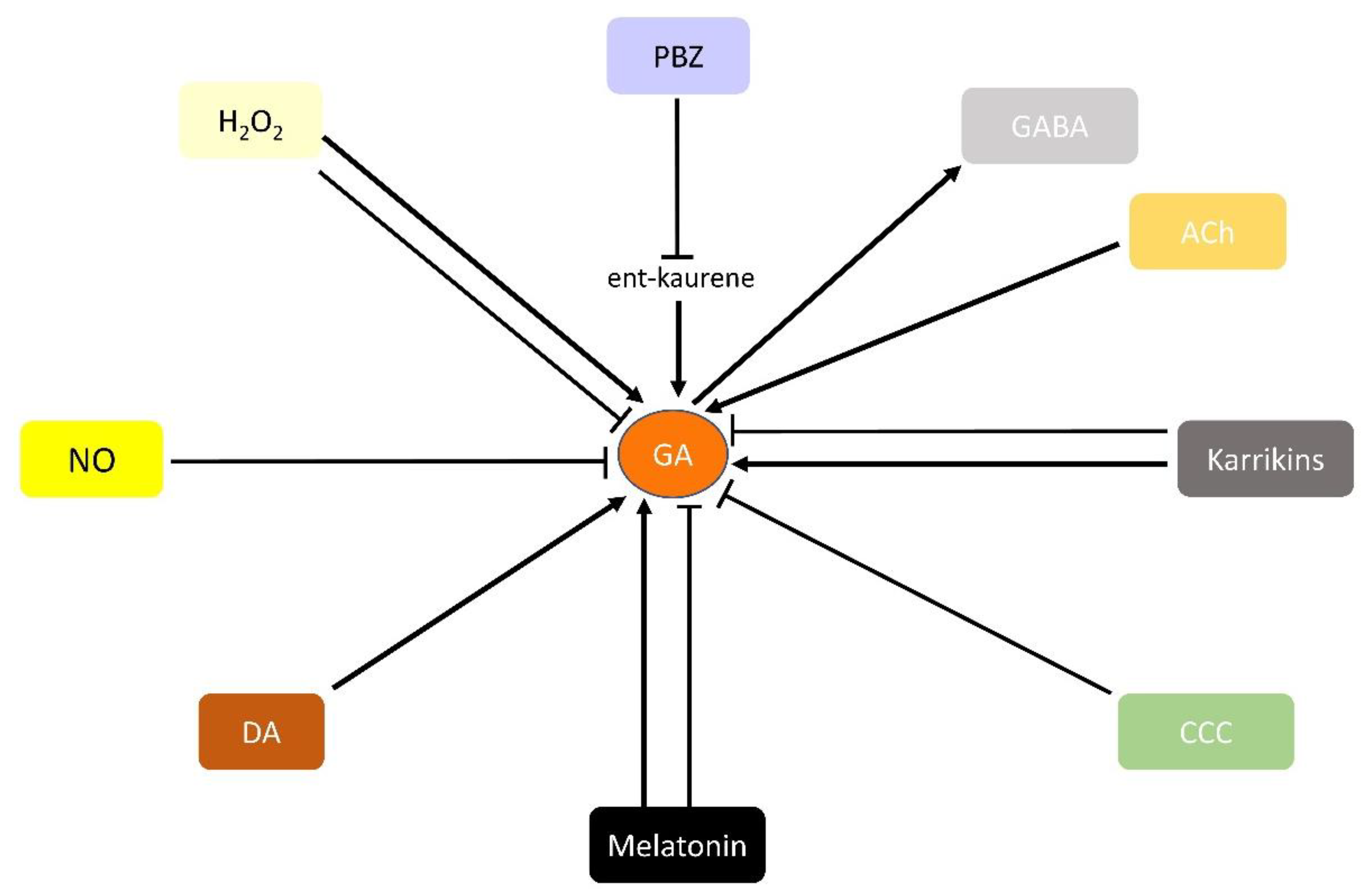
Several compounds not properly established as PGRs have been shown to modulate plant responses in a similar fashion to the classical hormones. Although the amount of available data is limited, GAs seem to interact with these potential PGRs (Figure 3). Oxygen and nitrogen reactive species are amongst these potential new PGRs. It has been shown that H2O2 treatment inhibits GAs biosynthesis genes in mung bean during adventitious rooting [112]. By contrast, these genes are upregulated by H2O2 and promote the germination of seeds in different species [113] or under hypoxia in pea [114]. The signaling molecule nitric oxide (NO) mediates auxin-induced processes, and eventually participates in adventitious root formation or in plant responses to different stresses, such as drought, salinity and heavy metal stress [115]. NO inhibits GAs signaling via the accumulation of DELLA proteins, and regulates endogenous levels of GA [116].
Melatonin is a well-known compound whose effects in plant development, including its influence on biomass production, seed germination and photosynthesis performance, have recently begun to be unraveled [117]. Exogenous melatonin treatment enhances seed germination in cotton by means of simultaneously increasing GA content and lowering ABA levels [118]. In cucumber, melatonin enhances seed germination via GA20ox and GA3ox activation, even under high salinity conditions [119]. Melatonin improves stress tolerance in plants through the modification of GA levels. Under heat stress, a melatonin pre-treatment enhances GA20ox expression and tolerance in tomato [120]. However, in cold-stressed watermelon plants, melatonin treatment reduces GA levels, and also induces a tolerance response [121]. In rice, melatonin deficiency promotes tolerance to cadmium, salt, cold and heat stress, reducing BRs content, which leads to a GA content drop [122]. Therefore, melatonin-induced stress responses seem to rely on the modulation of GA homeostasis, at least in the examples outlined.
Karrikins are a set of biomolecules found in the smoke derived from burnt plant material. Although their precise origin is not fully understood, these compounds seem to promote the seed germination of several species after fire. It is known that karrikins promote germination by stimulating GAs synthesis genes such as GA3ox, although surprisingly GA content does not change after karrikin application [123]. This implies a GA-independent response to karrikins [124]. However, this relationship between karrikins, GAs synthesis and germination is not found in all scenarios. In soybean, under dark conditions, karrikins delay seed germination, reducing GAs synthesis [125]. Moreover, karrikins can inhibit germination under unfavorable conditions, as a means of protection against abiotic stress [126].
Several compounds with a well-established role as neurotransmitters in animal biology have been found to present potential activity as PGRs. γ-aminobutyric acid (GABA) is a non-proteinogenic aminoacid that functions as an inhibitory neurotransmitter in mammals. GABA has been found in plants in recent years, and its activity has been related to several development processes. Despite the limited data available, GABA seems to have a relationship with GAs. Exogenous GA application increases GABA content in rice seeds [127], and controls GABA accumulation in grapevine depending on the time of application [128]. Moreover, GAs and GABA synthesis genes are upregulated after drought in Auricularia fibrillifera, suggesting potential synergy under specific conditions [129]. Dopamine (DA), another animal neurotransmitter also present in plants, can decrease ABA levels and increase GA levels in DA-treated maize seeds, stimulating their germination [130]. DA can also boosts GA3 action in lettuce hypocotyls [131]. Finally, Acetylcholine (ACh), usually characterized as an animal hormone, plays a role in plant development, raising the content of free GAs in Scots pine seedlings [132] and upregulating the GAs signaling pathway of tobacco plants under salt stress [133]. Future research on the activity of these neurotransmitters in plants will probably increase the number of interactions with GAs.
A few synthetic compounds, such as Paclobutrazol (PBZ) and chlorocholine chloride (CCC), have been shown to modulate GAs homeostasis. CCC inhibits GAs synthesis, and it is used in agriculture to increase nitrogen use efficiency and grain yield in maize [134], or to improve Sequoia sempervivens tolerance to cooling and drought [135]. On the other hand, PBZ is also an inhibitor of GAs synthesis. It acts by preventing the oxidation of ent-kaurene, and therefore inhibiting GA synthesis. Through these inhibitions, several plant traits are affected by PBZ treatment, including growth, water status, membrane stability, photosynthesis, etc. (reviewed in [136]). These effects on plant growth have made PBZ one of the main compounds used to modulate GA content, and therefore plant development, in agriculture.
4. Stress Responses
4.1. Abiotic Stress
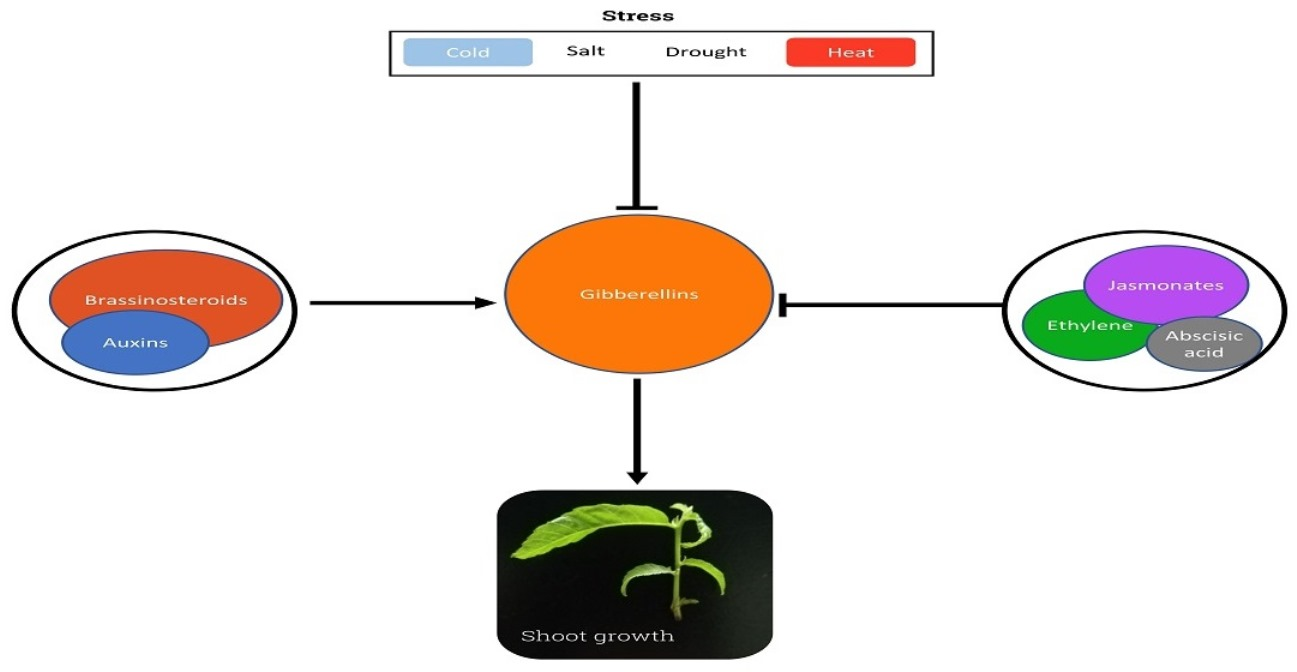
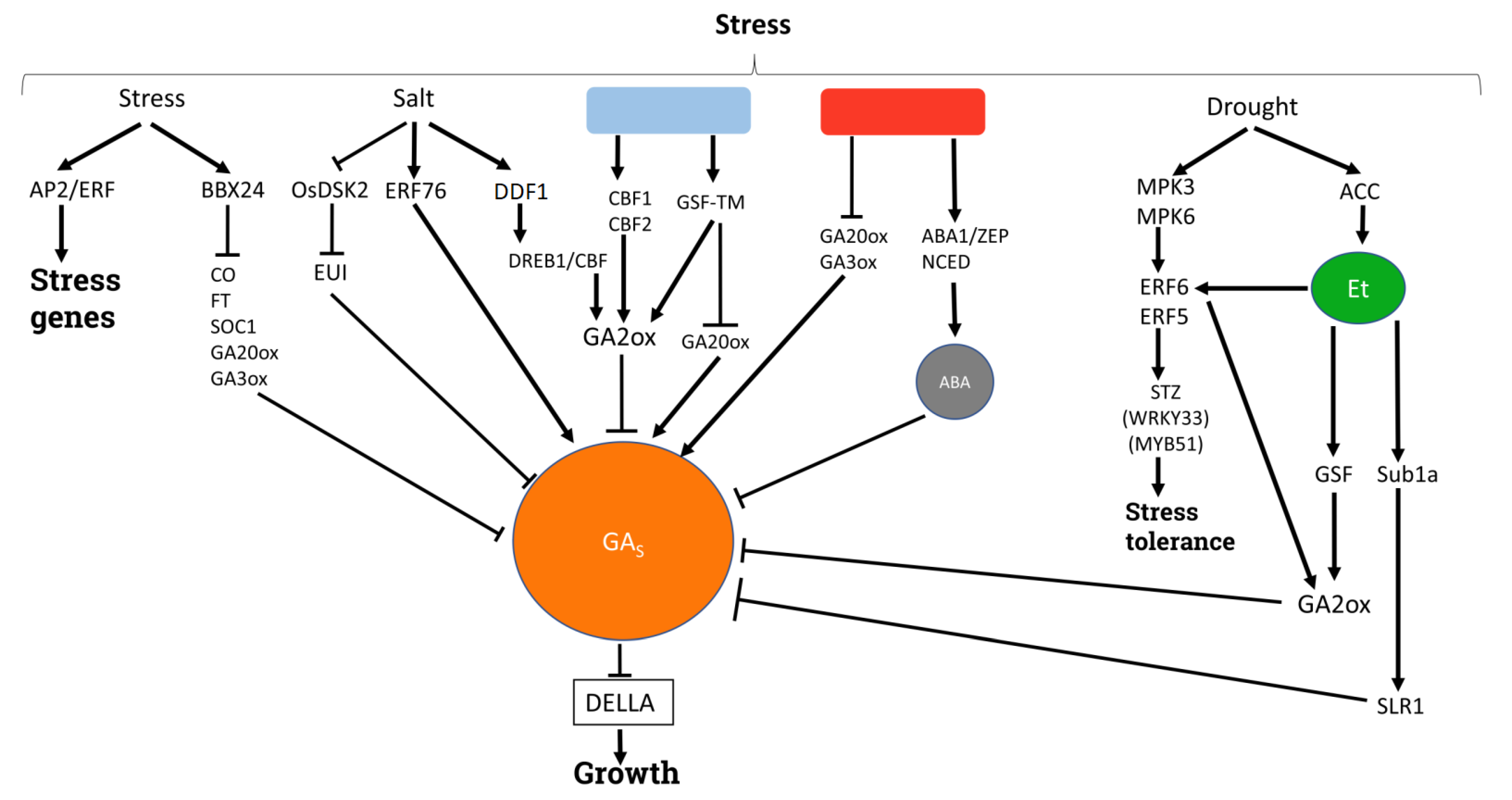
Phytohormones play essential roles in the adaptation of plants to the shifting conditions of the environment and the adjustment of their growth to nutrient availability. The activity of GAs seems to relate to several stress responses, particularly those of an abiotic nature (Figure 4). According to the results described in rice, responses to abiotic stress involve the regulation of GA homeostasis, where low levels are associated with an increased tolerance to stress [137]. Lowering bioactive GAs levels allows DELLA proteins to function as repressors of the growth factor RGA-like protein 3 (RGL3), or as activators of the stress-responsive genes APETALA2/ETHYLENERRESPONSEFACTOR (AP2/ERF) [138]. Plants establish a balanced trade-off between growth and defense responses, investing energy in one process or the other according to the ongoing conditions. Under abiotic stress, several biological processes, such as flowering, are modified or repressed as a strategy to save energy, which can be used to improve resistance [139]. GAs control flowering in many species. The B-BOX 24 zinc finger is a TF encoded by BBX24 that represses the transcription of the enhancer flowering genes CONSTANS (CO), FLOWERING LOCUS T (FT) and SUPPRESSOR OF OVEREXPRESSION OF CO1 (SOC1), and simultaneously reduces GA levels by repressing GA20ox and GA3ox. This activity inhibits flowering and improves the resistance of plants to cold, drought and salinity [140].
4.1.1. Mineral Stress
Plants often have to deal with reduced or excessive contents of minerals in the soil, which can severely impact their growth or have toxic effects. GAs mediate several mineral-related processes to adapt plant growth to the ongoing conditions. In wheat, the tolerance to K-deprivation was improved in plants with reduced GA sensitivity, suggesting GA mediation in the process [141]. In maize, the significant role of GAs has been shown in the response to low levels of phosphate (Pi) and nitrogen [142,143]. The reduction in bioactive GAs level and the subsequent accumulation of DELLAs caused by Pi starvation drives the inhibition of shoot growth and promotes root hair elongation to improve plant adaptation [144]. Furthermore, the normal growth of apple transgenic plants with decreased GA biosynthesis in response to Pi starvation is rescued by the application of GA [145]. Similarly, in maize, low N availability reduces the level of bioactive GAs. However, the application of GAs up-regulates the expression of transporters (NRTs) through the activity of several TFs [143]. The possible relationship between the decrease in GA content and the reduction in root elongation in maize plants treated with copper has been suggested by Matayoshi et al. [146]. GA signaling is also involved in the adaptation of plant responses to iron deficiency. Under low iron conditions, DELLAs are excluded from the epidermal cells of the root differentiation zone, the site of iron acquisition, which allows the expression of iron uptake genes activated by the FER-like IRON-DEFICIENCY INDUCED TRANSCRIPTION FACTOR (FIT) [147]. Thus, GAs are able to modulate the mineral uptake machinery and root development, providing a plastic framework under these growth-limiting conditions.
The positive effects of GA on the improvement of biomass production and photosynthesis parameters in response to the presence of ZnO nanoparticles have been reported in wheat [148]. Nonetheless, the relation of GA content and mineral stress is not always straightforward, as biosynthetic GA genes are upregulated in soybean under cadmium stress [149]. Indeed, GA accumulation leads to a rise in putrescine levels in rice plants, and reduces tolerance to ammonium stress [150].
4.1.2. Thermal Stress
Under low temperatures, plants activate the TFs encoded by COLD INDUCIBLE FACTOR (CBF) CBF1 and CBF3 genes, whose activity restrains plant growth. These TFs promote the expression of GA2ox and GA2ox7, which reduces GAs content. Consequently, the accumulation of DELLA proteins inhibits growth and enhances cold stress tolerance [151,152]. Similarly, under cold treatments, GIBBERELLIN SUPPRESSOR FACTOR-TRANSMEMBRANE DOMAIN (GSF-TM) activates GA2ox and represses GA20ox genes, reducing bioactive GA content [153]. Moreover, the coordinated effect of temperature and light in improving cold stress responses by means of CBF TFs modulation has been reported in tomato [154].
On the other hand, the effect of heat stress in GA synthesis and activity has been described during seed development. Heat inhibits the expression of GA20ox and GA3ox genes, and therefore, GA synthesis is blocked. In addition, the activation of ABA1/ZEAXANTHIN EPOXIDASE (ABA1/ZEP) and NINE-CIS-EPOXYCAROTENOID DIOXYGENASE 2 (NCED) promotes ABA synthesis and increases endogenous ABA levels. The drop in GA content allows SPY activation, which acts by means of DELLA proteins blocking seed germination at supraoptimal temperatures [155].
4.1.3. Saline Stress
Under saline stress, growth is restrained through DELLA accumulation and GAs content reduction [156]. In rice plants grown under normal conditions, DOMINANT SUPPRESSOR OF KAR2 (OsDSK2) enhances EUI poly-ubiquitination and degradation, but under saline stress OsDSK2 levels decrease and EUI promotes GAs inactivation [157]. In Arabidopsis, high salinity levels induce the expression of DWARF AND DELAYED FLOWERING 1 (DDF1), an AP2 transcription factor that belongs to the DEHYDRATION RESPONSIVE ELEMENT BINDING PROTEIN (DREB1)/C-REPEAT BINDING FACTOR (CBF) family. The DDF1 induction of the GA3ox7 gene lowers GA levels, and represses plant growth to adapt it to saline stress [158]. However, the opposite trend has been detected in tree species. In poplar, the response to saline stress has been improved by the increase in GA biosynthesis mediated by ERF76 [159,160]. Moreover, MdBZR1 and MdBZR1-like TFs enhance GA biosynthesis genes and improve salt tolerance in apple [161]. In addition, this trend is confirmed in Scots pine, in which acetylcholine upregulates GAs biosynthetic genes under salt stress conditions [133].
4.1.4. Water Stress
Plants’ responses to drought conditions are particularly influenced by ethylene and ABA. It is expected that ABA and GAs present an antagonistic behavior in the response to drought, although the molecular mechanisms of their interaction in this context have not been elucidated [162]. Ethylene modulates the levels of other phytohormones (including GAs) to cope with this growth-limiting situation. Water stress and weak osmotic stress in leaves induce the accumulation of the ethylene precursor 1-Aminocycloropane (ACC). In the first step, the protein kinases MPK3 and MPK6 phosphorylate and activate ERF5 and ERF6 ethylene response factors. In turn, these ERFs induce the stress tolerance genes SALT TOLERANCE ZINC-FINGER (STZ), WRKY33 and MYB51. Besides this, the expression of GA2ox6 induced by ERF5 and ERF6 degrades GAs and stabilizes the DELLA proteins, thus repressing plant growth [163]. Nonetheless, GA biosynthesis genes are highly expressed in drought-tolerant soybean cultivars when compared to drought-sensitive ones [149]. Under drought stress, GSF-TM represses GA20ox and stimulates GA2ox, lowering bioactive GAs levels [153]. Moreover, the suppression of GA signaling and activity enhances water stress responses in tomato plants [164]. When GA receptors are mutated or GA activity is suppressed, water loss is reduced through different strategies, including leaf area reduction, the repression of xylem formation or stomatal closure [165,166]. Indeed, high CO2 levels induce GAs biosynthesis genes in Masson pine, leading to stomatal density, conductance, and transpiration rate modulation to cope with the stress [167]. As in the case of drought, there is also a link between ethylene and GAs in the case of waterlogged plants. The ethylene-induced expression of SUBMERGENCE-1A (Sub1A) activates SLR1, which reduces GA response and represses the growth of rice plants [168]. Therefore, the proposed antagonistic relationship between GAs and ethylene stated above seems clear in this type of stress.
4.2. Biotic Stress
The main effects of GAs on the pathogenic responses of plants are a consequence of GA interaction with other phytohormones. Moreover, a balance between growth and defense responses must be met because plants invest energy in one process or the other. In this sense, GAs with growth-promoting activity must be shut down to improve defense responses. As previously mentioned, GAs induce the ubiquitination of DELLA proteins, inactivating JAZ1 and repressing MYC2 [169], which inhibits growth and activates the response to pathogens [170]. NON-RACE-SPECIFIC DISEASE RESISTANCE (NDR), a pathogen defense gene, interacts with GAs, balancing the response against pathogens with the flowering and growth processes [171]. In Arabidopsis, DELLA proteins’ repression reduces resistance to necrotrophic pathogens without interacting with salicylic acid (SA) or JA. On the other hand, in rice, SA and JA induce the expression of the DELLA protein SLR1 to enhance resistance to hemibiotrophic and necrotrophic pathogens, and at the same time repress GA signaling, which leads to SLR1 stabilization [172].
Plant responses to viral diseases are also mediated by GAs [173]. Different viruses disturb the development of plants by interfering with the GAs signaling pathway [174] or by altering GA synthesis, reducing their concentrations in species such as potato [175] and rice [176]. GAs reduce plant resistance against diseases caused by bacteria, such as the bacterial blight (Xanthomonas oryzae) in cassava [177] or in rice [178]. Studies carried out in rice in which SLR1 proteins were stabilized through the overexpression of IDEAL PLANT ARCHITECTURE 1 (IPA1), disturbing the GAs signaling pathway, [179] or in which GA synthesis was altered [180], support this hypothesis.
In the case of diseases caused by the attack of nematodes, the role of GAs is still obscure. Studies performed in rice showed a worsening of symptoms caused by Magnaporthe oryzae [180] and Meloidogyne graminicola due to the antagonistic GA–JA crosstalk [181]. However, the exogenous application of GAs seems to improve the resistance of soybean plants to the soybean cyst nematode (Heterodera glycines) [182]. GAs restrain JA-related defense responses, thus shifting the balance between JA and salicylic acid, resulting in the enhancement of salicylic acid signaling and biotroph resistance [183,184].
Root Symbiosis
Roots are continuously exposed to a vast range of microbes within the so-called rhizosphere. GAs seem to be related to the establishment of symbiotic associations of roots with soil microorganisms and fungi. In pea, GAs reduce the number of nodules, but increase their growth in a way independent of ethylene, a key growth regulator of nodulation [185]. In Pisum sativum and Medicago truncatula, DELLA proteins regulate nodule formation by modulating the activities of KNOX and BEL1-LIKE HOMEODOMAIN (BELL) TFs [186,187]. In roots, DELLA proteins, in conjunction with other GRAS proteins, are necessary for mycorrhizal colonization. Therefore, the antagonistic relation between GAs and DELLA proteins suggests that the GAs negatively affect mycorrhizal symbiosis [188]. Nevertheless, GAs can positively and negatively regulate the development of mycorrhizae [189]. A proper understanding of the effects of GAs on arbuscular mycorrhizae is essential due to their role in the regulation of the productivity of horticultural crops [190].
However, many symbiotic and non-symbiotic plant growth-promoting rhizobacteria (PGPR) have been found to synthesize GAs, therefore influencing GA levels in plants and consequently growth and stress responses, among other processes [191]. This has important practical effects, since they can be added to fertilizers to improve crop production [192] in species such as banana [193] or tobacco [194].
5. Conclusions
Interest in GAs research is a continuously increasing subject due to the significant impacts of these compounds on plant growth, responses to stress, and overall performance. The nature of the interaction with other PGRs is considerably complex, although in general terms, it presents a cooperative nature with other growth-inducing phytohormones (auxins, brassinosteroids), while it is mostly antagonistic towards stress-related PGRs (ethylene, jasmonates, ABA). However, this might be an oversimplification as, for specific responses, the nature of these relationships might shift, as shown above. The identification of genes and proteins at the core of these interactions enables the modulation of specific plant responses. Moreover, the interactions of GAs with new potential PGRs and other compounds provide the means for the precise articulation of plant performance, particularly in relevant crops.
Interestingly, the link between GAs signaling and the ability of plants to adapt to different types of stress facilitates the attainment of higher degrees of resilience, which might be crucial for plant adaptation to the conditions derived from a climate change scenario. The knowledge gathered in the present review concerning interactions with PGRs and responses to stress, together with the identification of genes involved in those processes, opens the pathway for the development of specific and accurate breeding programs that might help increase plant production and resilience.
Gibberellins have led to a Green Revolution that has greatly increased crop yield in the second half of the last century. However, the main challenge nowadays is how to maintain or increase that production with a continuously growing world population and under the context of a climate change, in which plants will be threatened by increasing biotic and abiotic stresses.
Author Contributions
Conceptualization, J.M.V.; investigation, R.C.-C.; writing—original draft preparation, R.C.-C.; visualization, R.C.-C.; visualization—editing, J.M.V.; writing—review and editing, J.M.V. and C.S.; supervision, C.S.; funding, C.S. and N.V. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Xunta de Galicia (Spain) through the projects IN607A and “Contrato Programa” 2021 (AGI/CSIC I+D+I 2021, Ref- ACAM 20210200033).
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- MacMillan, J. Occurrence of Gibberellins in Vascular Plants, Fungi, and Bacteria. J. Plant Growth Regul. 2002, 20, 387–442. [Google Scholar] [CrossRef] [PubMed]
- Bon, D.J.Y.D.; Mander, L.N.; Lan, P. Syntheses of Gibberellins A15 and A24, the Key Metabolites in Gibberellin Biosynthesis. J. Org. Chem. 2018, 83, 6566–6572. [Google Scholar] [CrossRef] [PubMed]
- Camara, M.C.; Vandenberghe, L.P.S.; Rodrigues, C.; de Oliveira, J.; Faulds, C.; Bertrand, E.; Soccol, C.R. Current advances in gibberellic acid (GA3) production, patented technologies and potential applications. Planta 2018, 248, 1049–1062. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. A novel gibberellin promotes seedling establishment. Nat. Plants 2019, 5, 459–460. [Google Scholar] [CrossRef] [PubMed]
- Sponsel, V.M.; Hedden, P. Gibberellin Biosynthesis and Inactivation. In Plant Hormones; Springer: Dordrecht, The Netherlands, 2010; pp. 63–94. [Google Scholar]
- Bömke, C.; Tudzynski, B. Diversity, regulation, and evolution of the gibberellin biosynthetic pathway in fungi compared to plants and bacteria. Phytochemistry 2009, 70, 1876–1893. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Cerezo, S.; Martínez-Montiel, N.; García-Sánchez, J.; Pérez-y-Terrón, R.; Martínez-Contreras, R.D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef]
- Thomas, S.G.; Phillips, A.L.; Hedden, P. Molecular cloning and functional expression of gibberellin 2- oxidases, multifunctional enzymes involved in gibberellin deactivation. Proc. Natl. Acad. Sci. USA 1999, 96, 4698–4703. [Google Scholar] [CrossRef] [Green Version]
- Kawaide, H. Biochemical and Molecular Analyses of Gibberellin Biosynthesis in Fungi. Biosci. Biotechnol. Biochem. 2006, 70, 583–590. [Google Scholar] [CrossRef]
- Hedden, P.; Kamiya, Y. GIBBERELLIN BIOSYNTHESIS: Enzymes, Genes and Their Regulation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1997, 48, 431–460. [Google Scholar] [CrossRef] [Green Version]
- Binenbaum, J.; Weinstain, R.; Shani, E. Gibberellin Localization and Transport in Plants. Trends Plant Sci. 2018, 23, 410–421. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Lange, M.J.P.; Lange, T. Ovary-derived precursor gibberellin A9 essential for cucumber female flower development. Development 2016, 143, 4425–4429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magome, H.; Nomura, T.; Hanada, A.; Takeda-Kamiya, N.; Ohnishi, T.; Shinma, Y.; Katsumata, T.; Kawaide, H.; Kamiya, Y.; Yamaguchi, S. CYP714B1 and CYP714B2 encode gibberellin 13-oxidases that reduce gibberellin activity in rice. Proc. Natl. Acad. Sci. USA 2013, 110, 1947–1952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nomura, T.; Magome, H.; Hanada, A.; Takeda-Kamiya, N.; Mander, L.N.; Kamiya, Y.; Yamaguchi, S. Functional analysis of arabidopsis CYP714A1 and CYP714A2 reveals that they are distinct gibberellin modification enzymes. Plant Cell Physiol. 2013, 54, 1837–1851. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Chen, Q.; Xin, P.; Yuan, J.; Ma, Y.; Wang, X.; Xu, M.; Chu, J.; Peters, R.J.; Wang, G. CYP72A enzymes catalyse 13-hydrolyzation of gibberellins. Nat. Plants 2019, 5, 1057–1065. [Google Scholar] [CrossRef] [Green Version]
- Rieu, I.; Eriksson, S.; Powers, S.J.; Gong, F.; Griffiths, J.; Woolley, L.; Benlloch, R.; Nilsson, O.; Thomas, S.G.; Hedden, P.; et al. Genetic analysis reveals that C19-GA 2-oxidation is a major gibberellin inactivation pathway in Arabidopsis. Plant Cell 2008, 20, 2420–2436. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Nomura, T.; Xu, Y.; Zhang, Y.; Peng, Y.; Mao, B.; Hanada, A.; Zhou, H.; Wang, R.; Li, P.; et al. ELONGATED UPPERMOST INTERNODE Encodes a Cytochrome P450 Monooxygenase That Epoxidizes Gibberellins in a Novel Deactivation Reaction in Rice. Plant Cell 2006, 18, 442–456. [Google Scholar] [CrossRef] [Green Version]
- Piotrowska, A.; Bajguz, A. Conjugates of abscisic acid, brassinosteroids, ethylene, gibberellins, and jasmonates. Phytochemistry 2011, 72, 2097–2112. [Google Scholar] [CrossRef]
- Schneider, G.; Schliemann, W. Gibberellin conjugates: An overview. Plant Growth Regul. 1994, 15, 247–260. [Google Scholar] [CrossRef]
- Varbanova, M.; Yamaguchi, S.; Yang, Y.; McKelvey, K.; Hanada, A.; Borochov, R.; Yu, F.; Jikumaru, Y.; Rosa, J.; Cortea, D.; et al. Methylation of gibberellins by Arabidopsis GAMT1 and GAMT2. Plant Cell 2007, 19, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Kramer, E.M. How far can a molecule of weak acid travel in the apoplast or xylem? Plant Physiol. 2006, 141, 1233–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanno, Y.; Hanada, A.; Chiba, Y.; Ichikawa, T.; Nakazawa, M.; Matsui, M.; Koshiba, T.; Kamiya, Y.; Seo, M. Identification of an abscisic acid transporter by functional screening using the receptor complex as a sensor. Proc. Natl. Acad. Sci. USA 2012, 109, 9653–9658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, H.; Oikawa, T.; Hamamoto, S.; Ishimaru, Y.; Kanamori-Sato, M.; Sasaki-Sekimoto, Y.; Utsumi, T.; Chen, J.; Kanno, Y.; Masuda, S.; et al. The jasmonate-responsive GTR1 transporter is required for gibberellin-mediated stamen development in Arabidopsis. Nat. Commun. 2015, 6, 6095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tal, I.; Zhang, Y.; Jørgensen, M.E.; Pisanty, O.; Barbosa, I.C.R.; Zourelidou, M.; Regnault, T.; Crocoll, C.; Erik Olsen, C.; Weinstain, R.; et al. The Arabidopsis NPF3 protein is a GA transporter. Nat. Commun. 2016, 7, 11486. [Google Scholar] [CrossRef] [PubMed]
- Kanno, Y.; Oikawa, T.; Chiba, Y.; Ishimaru, Y.; Shimizu, T.; Sano, N.; Koshiba, T.; Kamiya, Y.; Ueda, M.; Seo, M. AtSWEET13 and AtSWEET14 regulate gibberellin-mediated physiological processes. Nat. Commun. 2016, 7, 13245. [Google Scholar] [CrossRef] [PubMed]
- Ragni, L.; Nieminen, K.; Pacheco-Villalobos, D.; Sibout, R.; Schwechheimer, C.; Hardtke, C.S. Mobile gibberellin directly stimulates Arabidopsis hypocotyl xylem expansion. Plant Cell 2011, 23, 1322–1326. [Google Scholar] [CrossRef] [Green Version]
- Regnault, T.; Davière, J.-M.; Wild, M.; Sakvarelidze-Achard, L.; Heintz, D.; Carrera Bergua, E.; Lopez Diaz, I.; Gong, F.; Hedden, P.; Achard, P. The gibberellin precursor GA12 acts as a long-distance growth signal in Arabidopsis. Nat. Plants 2015, 1, 15073. [Google Scholar] [CrossRef]
- Shani, E.; Weinstain, R.; Zhang, Y.; Castillejo, C.; Kaiserli, E.; Chory, J.; Tsien, R.Y.; Estelle, M. Gibberellins accumulate in the elongating endodermal cells of Arabidopsis root. Proc. Natl. Acad. Sci. USA 2013, 110, 4834–4839. [Google Scholar] [CrossRef] [Green Version]
- Rizza, A.; Tang, B.; Stanley, C.E.; Grossmann, G.; Owen, M.R.; Band, L.R.; Jones, A.M. Differential biosynthesis and cellular permeability explain longitudinal gibberellin gradients in growing roots. Proc. Natl. Acad. Sci. USA 2021, 118. [Google Scholar] [CrossRef]
- Rizza, A.; Jones, A.M. The makings of a gradient: Spatiotemporal distribution of gibberellins in plant development. Curr. Opin. Plant Biol. 2019, 47, 9–15. [Google Scholar] [CrossRef]
- Camut, L.; Regnault, T.; Sirlin-Josserand, M.; Sakvarelidze-Achard, L.; Carrera, E.; Zumsteg, J.; Heintz, D.; Leonhardt, N.; Lange, M.J.P.; Lange, T.; et al. Root-derived GA12 contributes to temperature-induced shoot growth in Arabidopsis. Nat. Plants 2019, 5, 1216–1221. [Google Scholar] [CrossRef] [PubMed]
- Davière, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.L.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.P.; et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell 2006, 18, 3399–3414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueguchi-Tanaka, M.; Nakajima, M.; Katoh, E.; Ohmiya, H.; Asano, K.; Saji, S.; Hongyu, X.; Ashikari, M.; Kitano, H.; Yamaguchi, I.; et al. Molecular interactions of a soluble gibberellin receptor, GID1, with a rice DELLA protein, SLR1, and gibberellin. Plant Cell 2007, 19, 2140–2155. [Google Scholar] [CrossRef] [Green Version]
- Murase, K.; Hirano, Y.; Sun, T.P.; Hakoshima, T. Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 2008, 456, 459–463. [Google Scholar] [CrossRef]
- Shimada, A.; Ueguchi-Tanaka, M.; Nakatsu, T.; Nakajima, M.; Naoe, Y.; Ohmiya, H.; Kato, H.; Matsuoka, M. Structural basis for gibberellin recognition by its receptor GID1. Nature 2008, 456, 520–523. [Google Scholar] [CrossRef]
- McGinnis, K.M.; Thomas, S.G.; Soule, J.D.; Strader, L.C.; Zale, J.M.; Sun, T.P.; Steber, C.M. The Arabidopsis SLEEPY1 gene encodes a putative F-box subunit of an SCF E3 ubiquitin ligase. Plant Cell 2003, 15, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Gomi, K.; Sasaki, A.; Itoh, H.; Ueguchi-Tanaka, M.; Ashikari, M.; Kitano, H.; Matsuoka, M. GID2, an F-box subunit of the SCF E3 complex, specifically interacts with phosphorylated SLR1 protein and regulates the gibberellin-dependent degradation of SLR1 in rice. Plant J. 2004, 37, 626–634. [Google Scholar] [CrossRef]
- Van De Velde, K.; Ruelens, P.; Geuten, K.; Rohde, A.; Van Der Straeten, D. Exploiting DELLA Signaling in Cereals. Trends Plant Sci. 2017, 22, 880–893. [Google Scholar] [CrossRef]
- Conti, L.; Nelis, S.; Zhang, C.; Woodcock, A.; Swarup, R.; Galbiati, M.; Tonelli, C.; Napier, R.; Hedden, P.; Bennett, M.; et al. Small Ubiquitin-like Modifier Protein SUMO Enables Plants to Control Growth Independently of the Phytohormone Gibberellin. Dev. Cell 2014, 28, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Park, B.S.; Kim, D.Y.; Yeu, S.Y.; Song, S.I.; Song, J.T.; Seo, H.S. E3 SUMO ligase AtSIZ1 positively regulates SLY1-mediated GA signalling and plant development. Biochem. J. 2015, 469, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Xue, H.W. Rice early flowering1, a CKI, phosphorylates della protein SLR1 to negatively regulate gibberellin signalling. EMBO J. 2010, 29, 1916–1927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sasaki, A.; Itoh, H.; Gomi, K.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Jeong, D.H.; An, G.; Kitano, H.; Ashikari, M.; et al. Accumulation of phosphorylated repressor for Gibberellin signaling in an F-box mutant. Science 2003, 299, 1896–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Q.; Wang, W.; Guo, X.; Yue, J.; Huang, Y.; Xu, X.; Li, J.; Hou, S. Arabidopsis DELLA Protein Degradation Is Controlled by a Type-One Protein Phosphatase, TOPP4. PLoS Genet. 2014, 10, e1004464. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Hirano, K.; Sato, T.; Mitsuda, N.; Nomoto, M.; Maeo, K.; Koketsu, E.; Mitani, R.; Kawamura, M.; Ishiguro, S.; et al. DELLA protein functions as a transcriptional activator through the DNA binding of the INDETERMINATE DOMAIN family proteins. Proc. Natl. Acad. Sci. USA 2014, 111, 7861–7866. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.L.; Ogawa, M.; Fleet, C.M.; Zentella, R.; Hu, J.; Heo, J.O.; Lim, J.; Kamiya, Y.; Yamaguchi, S.; Sun, T.P. SCARECROW-LIKE 3 promotes gibberellin signaling by antagonizing master growth repressor DELLA in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 2160–2165. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, J.; Ito, T.; Kamiya, Y.; Yamaguchi, S.; Takahashi, Y. Binding of GID1 to DELLAs promotes dissociation of GAF1 from DELLA in GA dependent manner. Plant Signal. Behav. 2015, 10, e1052923. [Google Scholar] [CrossRef] [Green Version]
- Fukazawa, J.; Mori, M.; Watanabe, S.; Miyamoto, C.; Ito, T.; Takahashi, Y. DELLA-GAF1 complex is a main component in gibberellin feedback regulation of GA20 oxidase 2. Plant Physiol. 2017, 175, 1395–1406. [Google Scholar] [CrossRef] [Green Version]
- Dai, M.; Zhao, Y.; Ma, Q.; Hu, Y.; Hedden, P.; Zhang, Q.; Zhou, D.X. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism. Plant Physiol. 2007, 144, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Ma, Y.; Li, J. The rice YABBY4 gene regulates plant growth and development through modulating the gibberellin pathway. J. Exp. Bot. 2016, 67, 5545–5556. [Google Scholar] [CrossRef] [Green Version]
- Zentella, R.; Hu, J.; Hsieh, W.-P.; Matsumoto, P.A.; Dawdy, A.; Barnhill, B.; Oldenhof, H.; Hartweck, L.M.; Maitra, S.; Thomas, S.G.; et al. O-GlcNAcylation of master growth repressor DELLA by SECRET AGENT modulates multiple signaling pathways in Arabidopsis. Genes Dev. 2016, 30, 164–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimada, A.; Ueguchi-Tanaka, M.; Sakamoto, T.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Sazuka, T.; Ashikari, M.; Matsuoka, M. The rice SPINDLY gene functions as a negative regulator of gibberellin signaling by controlling the suppressive function of the DELLA protein, SLR1, and modulating brassinosteroid synthesis. Plant J. 2006, 48, 390–402. [Google Scholar] [CrossRef] [PubMed]
- Filardo, F.; Robertson, M.; Singh, D.P.; Parish, R.W.; Swain, S.M. Functional analysis of HvSPY, a negative regulator of GA response, in barley aleurone cells and Arabidopsis. Planta 2009, 229, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Sarnowska, E.A.; Rolicka, A.T.; Bucior, E.; Cwiek, P.; Tohge, T.; Fernie, A.R.; Jikumaru, Y.; Kamiya, Y.; Franzen, R.; Schmelzer, E.; et al. DELLA-interacting SWI3C core subunit of switch/sucrose nonfermenting chromatin remodeling complex modulates gibberellin responses and hormonal cross talk in Arabidopsis. Plant Physiol. 2013, 163, 305–317. [Google Scholar] [CrossRef] [Green Version]
- Archacki, R.; Buszewicz, D.; Sarnowski, T.J.; Sarnowska, E.; Rolicka, A.T.; Tohge, T.; Fernie, A.R.; Jikumaru, Y.; Kotlinski, M.; Iwanicka-Nowicka, R.; et al. BRAHMA ATPase of the SWI/SNF Chromatin Remodeling Complex Acts as a Positive Regulator of Gibberellin-Mediated Responses in Arabidopsis. PLoS ONE 2013, 8, e58588. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Oh, D.H.; Dassanayake, M.; Nguyen, K.T.; Ogas, J.; Choi, G.; Sun, T.P. Gibberellin signaling requires chromatin remodeler PICKLE to promote vegetative growth and phase transitions. Plant Physiol. 2017, 173, 1463–1474. [Google Scholar] [CrossRef] [Green Version]
- Yamamuro, C.; Zhu, J.-K.; Yang, Z. Epigenetic Modifications and Plant Hormone Action. Mol. Plant 2016, 9, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Frigerio, M.; Alabadí, D.; Pérez-Gómez, J.; García-Cárcel, L.; Phillips, A.L.; Hedden, P.; Blázquez, M.A. Transcriptional regulation of gibberellin metabolism genes by auxin signaling in Arabidopsis. Plant Physiol. 2006, 142, 553–563. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, D.P.; Davidson, S.E.; Clarke, V.C.; Yamauchi, Y.; Yamaguchi, S.; Kamiya, Y.; Reid, J.B.; Ross, J.J. Regulation of the gibberellin pathway by auxin and DELLA proteins. Planta 2010, 232, 1141–1149. [Google Scholar] [CrossRef]
- Yin, C.; Gan, L.; Ng, D.; Zhou, X.; Xia, K. Decreased panicle-derived indole-3-acetic acid reduces gibberellin A 1 level in the uppermost internode, causing panicle enclosure in male sterile rice Zhenshan 97A. J. Exp. Bot. 2007, 58, 2441–2449. [Google Scholar] [CrossRef] [Green Version]
- Weston, D.E.; Reid, J.B.; Ross, J.J. Auxin regulation of gibberellin biosynthesis in the roots of pea (Pisum sativum). Funct. Plant Biol. 2009, 36, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Serrani, J.C.; Ruiz-Rivero, O.; Fos, M.; García-Martínez, J.L. Auxin-induced fruit-set in tomato is mediated in part by gibberellins. Plant J. 2008, 56, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, E.; Zhu, J.Y.; Bai, M.Y.; Arenhart, R.A.; Sun, Y.; Wang, Z.Y. Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl. Elife 2014, 3, e03031. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Abdelrahman, M.; Jiu, S.; Guan, L.; Han, J.; Zheng, T.; Jia, H.; Song, C.; Fang, J.; Wang, C. VvmiR160s/VvARFs interaction and their spatio-temporal expression/cleavage products during GA-induced grape parthenocarpy. BMC Plant Biol. 2019, 19, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanenka, Y.; Verstraeten, I.; Löfke, C.; Tabata, K.; Naramoto, S.; Glanc, M.; Friml, J. Gibberellin DELLA signaling targets the retromer complex to redirect protein trafficking to the plasma membrane. Proc. Natl. Acad. Sci. USA 2018, 115, 3716–3721. [Google Scholar] [CrossRef] [Green Version]
- Willige, B.C.; Isono, E.; Richter, R.; Zourelidou, M.; Schwechheimer, C. Gibberellin regulates PIN-FORMED abundance and is required for auxin transport-dependent growth and development in Arabidopsis thaliana. Plant Cell 2011, 23, 2184–2195. [Google Scholar] [CrossRef] [Green Version]
- Löfke, C.; Zwiewka, M.; Heilmann, I.; Van Montagu, M.C.E.; Teichmann, T.; Friml, J. Asymmetric gibberellin signaling regulates vacuolar trafficking of PIN auxin transporters during root gravitropism. Proc. Natl. Acad. Sci. USA 2013, 110, 3627–3632. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhao, L.; Guo, W.; Yu, Y.; Tao, L.; Zhang, L.; Song, X.; Huang, W.; Cheng, L.; Chen, J.; et al. Exogenous application of phytohormones promotes growth and regulates expression of wood formation-related genes in Populus simonii × P. nigra. Int. J. Mol. Sci. 2019, 20, 792. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.Y.; Guo, G.S.; Qiu, Z.F.; Li, X.D.; Zeng, B.S.; Fan, C.J. Exogenous GA3 application altered morphology, anatomic and transcriptional regulatory networks of hormones in Eucalyptus grandis. Protoplasma 2018, 255, 1107–1119. [Google Scholar] [CrossRef]
- Pizarro, A.; Díaz-Sala, C. Effect of polar auxin transport and gibberellins on xylem formation in pine cuttings under adventitious rooting conditions. Isr. J. Plant Sci. 2020, 67, 27–39. [Google Scholar] [CrossRef]
- Li, G.; Zhu, C.; Gan, L.; Ng, D.; Xia, K. GA3 enhances root responsiveness to exogenous IAA by modulating auxin transport and signalling in Arabidopsis. Plant Cell Rep. 2015, 34, 483–494. [Google Scholar] [CrossRef] [PubMed]
- Gallego-Bartolome, J.; Minguet, E.G.; Grau-Enguix, F.; Abbas, M.; Locascio, A.; Thomas, S.G.; Alabadi, D.; Blazquez, M.A. Molecular mechanism for the interaction between gibberellin and brassinosteroid signaling pathways in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 13446–13451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lucas, M.; Davière, J.M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, L.V.; Viola, I.L.; Ariel, F.D.; Gonzalez, D.H. Class I TCP Transcription Factors Target the Gibberellin Biosynthesis Gene GA20ox1 and the Growth-Promoting Genes HBI1 and PRE6 during Thermomorphogenic Growth in Arabidopsis. Plant Cell Physiol. 2019, 60, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Stewart Lilley, J.L.; Gan, Y.; Graham, I.A.; Nemhauser, J.L. The effects of DELLAs on growth change with developmental stage and brassinosteroid levels. Plant J. 2013, 76, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Unterholzner, S.J.; Rozhon, W.; Papacek, M.; Ciomas, J.; Lange, T.; Kugler, K.G.; Mayer, K.F.; Sieberer, T.; Poppenberger, B. Brassinosteroids are master regulators of gibberellin biosynthesis in Arabidopsis. Plant Cell 2015, 27, 2261–2272. [Google Scholar] [CrossRef] [Green Version]
- Castorina, G.; Consonni, G. The role of brassinosteroids in controlling plant height in poaceae: A genetic perspective. Int. J. Mol. Sci. 2020, 21, 1191. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Xiao, Y.; Liu, D.; Gao, S.; Liu, L.; Yin, Y.; Jin, Y.; Qian, Q.; Chu, C. Brassinosteroid regulates cell elongation by modulating gibberellin metabolism in ricec w open. Plant Cell 2014, 26, 4376–4393. [Google Scholar] [CrossRef] [Green Version]
- de Vleesschauwer, D.; van Buyten, E.; Satoh, K.; Balidion, J.; Mauleon, R.; Choi, I.R.; Vera-Cruz, C.; Kikuchi, S.; Höfte, M. Brassinosteroids antagonize gibberellin- and salicylate-mediated root immunity in rice. Plant Physiol. 2012, 158, 1833–1846. [Google Scholar] [CrossRef] [Green Version]
- Kurepin, L.V.; Joo, S.H.; Kim, S.K.; Pharis, R.P.; Back, T.G. Interaction of Brassinosteroids with Light Quality and Plant Hormones in Regulating Shoot Growth of Young Sunflower and Arabidopsis Seedlings. J. Plant Growth Regul. 2012, 31, 156–164. [Google Scholar] [CrossRef]
- Jager, C.E.; Symons, G.M.; Ross, J.J.; Smith, J.J.; Reid, J.B. The brassinosteroid growth response in pea is not mediated by changes in gibberellin content. Planta 2005, 221, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Baghour, M.; Chapple, A.; Hedden, P.; Van Der Straeten, D.; Genschik, P.; Moritz, T.; Harberd, N.P. The plant stress hormone ethylene controls floral transition via DELLA-dependent regulation of floral meristem-identity genes. Proc. Natl. Acad. Sci. USA 2007, 104, 6484–6489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Grauwe, L.; Chaerle, L.; Dugardeyn, J.; Decat, J.; Rieu, I.; Vriezen, W.H.; Moritz, T.; Beemster, G.T.S.; Phillips, A.L.; Harberd, N.P.; et al. Reduced gibberellin response affects ethylene biosynthesis and responsiveness in the Arabidopsis gai eto2-1 double mutant. New Phytol. 2008, 177, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; He, L.; Huang, R. The coordination of ethylene and other hormones in primary root development. Front. Plant Sci. 2019, 10, 874. [Google Scholar] [CrossRef]
- Shinozaki, Y.; Hao, S.; Kojima, M.; Sakakibara, H.; Ozeki-Iida, Y.; Zheng, Y.; Fei, Z.; Zhong, S.; Giovannoni, J.J.; Rose, J.K.C.; et al. Ethylene suppresses tomato (Solanum lycopersicum) fruit set through modification of gibberellin metabolism. Plant J. 2015, 83, 237–251. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Foo, E.; Ross, J.J.; Reid, J.B. Relationship between gibberellin, ethylene and nodulation in Pisum sativum. New Phytol. 2011, 189, 829–842. [Google Scholar] [CrossRef]
- An, F.; Zhang, X.; Zhu, Z.; Ji, Y.; He, W.; Jiang, Z.; Li, M.; Guo, H. Coordinated regulation of apical hook development by gibberellins and ethylene in etiolated Arabidopsis seedlings. Cell Res. 2012, 22, 915–927. [Google Scholar] [CrossRef] [Green Version]
- Zhao, N.; Zhao, M.; Tian, Y.; Wang, Y.; Han, C.; Fan, M.; Guo, H.; Bai, M.Y. Interaction between BZR1 and EIN3 mediates signalling crosstalk between brassinosteroids and ethylene. New Phytol. 2021, 232, 2308–2323. [Google Scholar] [CrossRef]
- Gallego-Bartolomé, J.; Arana, M.V.; Vandenbussche, F.; Žádníková, P.; Minguet, E.G.; Guardiola, V.; Van Der Straeten, D.; Benkova, E.; Alabadí, D.; Blázquez, M.A. Hierarchy of hormone action controlling apical hook development in Arabidopsis. Plant J. 2011, 67, 622–634. [Google Scholar] [CrossRef]
- Marín-de La Rosa, N.; Sotillo, B.; Miskolczi, P.; Gibbs, D.J.; Vicente, J.; Carbonero, P.; Oñate-Sánchez, L.; Holdsworth, M.J.; Bhalerao, R.; Alabadí, D.; et al. Large-scale identification of gibberellin-related transcription factors defines group VII ETHYLENE RESPONSE FACTORS as functional DELLA partners. Plant Physiol. 2014, 166, 1022–1032. [Google Scholar] [CrossRef] [Green Version]
- Shu, K.; Zhang, H.; Wang, S.; Chen, M.; Wu, Y.; Tang, S.; Liu, C.; Feng, Y.; Cao, X.; Xie, Q. ABI4 Regulates Primary Seed Dormancy by Regulating the Biogenesis of Abscisic Acid and Gibberellins in Arabidopsis. PLoS Genet. 2013, 9, e1003577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shu, K.; Chen, Q.; Wu, Y.; Liu, R.; Zhang, H.; Wang, P.; Li, Y.; Wang, S.; Tang, S.; Liu, C.; et al. ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J. 2016, 85, 348–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, R.; Kanno, Y.; Jikumaru, Y.; Nakabayashi, K.; Kamiya, Y.; Nambara, E. CHOTTO1, a putative double APETALA2 repeat transcription factor, is involved in abscisic acid-mediated repression of gibberellin Biosynthesis during seed germination in Arabidopsis. Plant Physiol. 2009, 151, 641–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Um, T.Y.; Lee, H.Y.; Lee, S.; Chang, S.H.; Chung, P.J.; Oh, K.-B.; Kim, J.-K.; Jang, G.; Choi, Y. Do Jasmonate Zim-Domain Protein 9 Interacts With Slender Rice 1 to Mediate the Antagonistic Interaction Between Jasmonic and Gibberellic Acid Signals in Rice. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Liu, H.; Timko, M.P. Jasmonic Acid Signaling and Molecular Crosstalk with Other Phytohormones. Int. J. Mol. Sci. 2021, 22, 2914. [Google Scholar] [CrossRef]
- Heinrich, M.; Hettenhausen, C.; Lange, T.; Wünsche, H.; Fang, J.; Baldwin, I.T.; Wu, J. High levels of jasmonic acid antagonize the biosynthesis of gibberellins and inhibit the growth of Nicotiana attenuata stems. Plant J. 2013, 73, 591–606. [Google Scholar] [CrossRef]
- Barbosa, N.C.S.; Dornelas, M.C. The Roles of Gibberellins and Cytokinins in Plant Phase Transitions. Trop. Plant Biol. 2021, 14, 11–21. [Google Scholar] [CrossRef]
- Fonouni-Farde, C.; Kisiala, A.; Brault, M.; Emery, R.J.N.; Diet, A.; Frugier, F. DELLA1-mediated gibberellin signaling regulates cytokinin-dependent symbiotic nodulation. Plant Physiol. 2017, 175, 1795–1806. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, L.; Ge, Y.; Wang, J.; Yu, J.; Yang, Z.; Huang, B. Gibberellic acid inhibition of tillering in tall fescue involving crosstalks with cytokinins and transcriptional regulation of genes controlling axillary bud outgrowth. Plant Sci. 2019, 287, 110168. [Google Scholar] [CrossRef]
- Fonouni-Farde, C.; McAdam, E.; Nichols, D.; Diet, A.; Foo, E.; Frugier, F. Cytokinins and the CRE1 receptor influence endogenous gibberellin levels in Medicago truncatula. Plant Signal. Behav. 2018, 13, e1428513. [Google Scholar] [CrossRef] [Green Version]
- Greenboim-Wainberg, Y.; Maymon, I.; Borochov, R.; Alvarez, J.; Olszewski, N.; Ori, N.; Eshed, Y.; Weiss, D. Cross Talk between Gibberellin and Cytokinin: The Arabidopsis GA Response Inhibitor SPINDLY Plays a Positive Role in Cytokinin Signaling. Plant Cell 2005, 17, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugiura, D.; Sawakami, K.; Kojima, M.; Sakakibara, H.; Terashima, I.; Tateno, M. Roles of gibberellins and cytokinins in regulation of morphological and physiological traits in Polygonum cuspidatum responding to light and nitrogen availabilities. Funct. Plant Biol. 2015, 42, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzec, M. Strigolactones and Gibberellins: A New Couple in the Phytohormone World? Trends Plant Sci. 2017, 22, 813–815. [Google Scholar] [CrossRef] [PubMed]
- Wallner, E.-S.; López-Salmerón, V.; Greb, T. Strigolactone versus gibberellin signaling: Reemerging concepts? Planta 2016, 243, 1339–1350. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Yamagami, D.; Asami, T. Effects of gibberellin and strigolactone on rice tiller bud growth. J. Pestic. Sci. 2018, 43, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, H.; Xue, Y.-L.; Miyakawa, T.; Hou, F.; Qin, H.-M.; Fukui, K.; Shi, X.; Ito, E.; Ito, S.; Park, S.-H.; et al. Molecular mechanism of strigolactone perception by DWARF14. Nat. Commun. 2013, 4, 2613. [Google Scholar] [CrossRef] [Green Version]
- Ito, S.; Yamagami, D.; Umehara, M.; Hanada, A.; Yoshida, S.; Sasaki, Y.; Yajima, S.; Kyozuka, J.; Ueguchi-Tanaka, M.; Matsuoka, M.; et al. Regulation of Strigolactone Biosynthesis by Gibberellin Signaling. Plant Physiol. 2017, 174, 1250–1259. [Google Scholar] [CrossRef]
- Ni, J.; Gao, C.; Chen, M.-S.; Pan, B.-Z.; Ye, K.; Xu, Z.-F. Gibberellin Promotes Shoot Branching in the Perennial Woody Plant Jatropha curcas. Plant Cell Physiol. 2015, 56, 1655–1666. [Google Scholar] [CrossRef] [Green Version]
- Toh, S.; Kamiya, Y.; Kawakami, N.; Nambara, E.; McCourt, P.; Tsuchiya, Y. Thermoinhibition uncovers a role for strigolactones in arabidopsis seed germination. Plant Cell Physiol. 2012, 53, 107–117. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.; Wang, Q.; Chen, P.; Yin, C.; Lin, Y. Strigolactones regulate shoot elongation by mediating gibberellin metabolism and signaling in rice (Oryza sativa L.). J. Plant Physiol. 2019, 237, 72–79. [Google Scholar] [CrossRef]
- Li, S.W.; Leng, Y.; Shi, R.F. Transcriptomic profiling provides molecular insights into hydrogen peroxide-induced adventitious rooting in mung bean seedlings. BMC Genom. 2017, 18, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazir, F.; Fariduddin, Q.; Khan, T.A. Hydrogen peroxide as a signalling molecule in plants and its crosstalk with other plant growth regulators under heavy metal stress. Chemosphere 2020, 252, 126486. [Google Scholar] [CrossRef] [PubMed]
- Zdunek-Zastocka, E.; Grabowska, A. The interplay of PsABAUGT1 with other abscisic acid metabolic genes in the regulation of ABA homeostasis during the development of pea seeds and germination in the presence of H2O2. Plant Sci. 2019, 285, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Nabi, R.B.S.; Tayade, R.; Hussain, A.; Kulkarni, K.P.; Imran, Q.M.; Mun, B.-G.; Yun, B.-W. Nitric oxide regulates plant responses to drought, salinity, and heavy metal stress. Environ. Exp. Bot. 2019, 161, 120–133. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin and its relationship to plant hormones. Ann. Bot. 2018, 121, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Sharif, R.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P.; et al. Melatonin and its effects on plant systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Liu, L.; Wang, H.; Li, D.; Bai, Z.; Zhang, Y.; Sun, H.; Zhang, K.; Li, C. Exogenous melatonin accelerates seed germination in cotton (Gossypium hirsutum L.). PLoS ONE 2019, 14, e0216575. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.J.; Zhang, N.; Yang, R.C.; Wang, L.; Sun, Q.Q.; Li, D.B.; Cao, Y.Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Jahan, M.S.; Shu, S.; Wang, Y.; Hasan, M.M.; El-Yazied, A.A.; Alabdallah, N.M.; Hajjar, D.; Altaf, M.A.; Sun, J.; Guo, S. Melatonin Pretreatment Confers Heat Tolerance and Repression of Heat-Induced Senescence in Tomato Through the Modulation of ABA- and GA-Mediated Pathways. Front. Plant Sci. 2021, 12, 1–14. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Zheng, J.; Dong, Y.; Liu, Q.; Yang, X.; Wei, C.; Zhang, Y.; Ma, J.; Zhang, X. Local melatonin application induces cold tolerance in distant organs of Citrullus lanatus L. via long distance transport. Sci. Rep. 2017, 7, 40858. [Google Scholar] [CrossRef] [Green Version]
- Hwang, O.J.; Back, K. Melatonin deficiency confers tolerance to multiple abiotic stresses in rice via decreased brassinosteroid levels. Int. J. Mol. Sci. 2019, 20, 5173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.C.; Riseborough, J.-A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins Discovered in Smoke Trigger Arabidopsis Seed Germination by a Mechanism Requiring Gibberellic Acid Synthesis and Light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.C.; Flematti, G.R.; Riseborough, J.-A.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins enhance light responses during germination and seedling development in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 7095–7100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Y.; Chen, F.; Shuai, H.; Luo, X.; Ding, J.; Tang, S.; Xu, S.; Liu, J.; Liu, W.; Du, J.; et al. Karrikins delay soybean seed germination by mediating abscisic acid and gibberellin biogenesis under shaded conditions. Sci. Rep. 2016, 6, 22073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Waters, M.T.; Smith, S.M. Karrikin-KAI2 signalling provides Arabidopsis seeds with tolerance to abiotic stress and inhibits germination under conditions unfavourable to seedling establishment. New Phytol. 2018, 219, 605–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Xiang, J.; Zhang, L.; Zhu, X.; Evers, J.; van der Werf, W.; Duan, L. Optimizing soaking and germination conditions to improve gamma-aminobutyric acid content in japonica and indica germinated brown rice. J. Funct. Foods 2014, 10, 283–291. [Google Scholar] [CrossRef]
- Jung, C.J.; Hur, Y.Y.; Moon, J.S.; Jung, S.-M. Pre-bloom application of gibberellin in ‘Tamnara’ grape increases γ-aminobutyric acid (GABA) production at full bloom. Hortic. Environ. Biotechnol. 2017, 58, 568–575. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, Z.; Shi, L.; Yang, R.; Guo, H.; Zhang, S.; Geng, G. Transcriptome analysis of Auricularia fibrillifera fruit-body responses to drought stress and rehydration. BMC Genomics 2022, 23, 58. [Google Scholar] [CrossRef]
- CHENG, H.; WANG, X.; FENG, T.; PENG, C.; WANG, W.; YANG, M.; ZHOU, Y. Giving maize an excited start—Effects of dopamine on maize germination. J. Integr. Agric. 2020, 19, 2690–2698. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, T.; Liu, W.; Liu, Y.; Zhao, Y.; Liu, Y.; Li, W.; Ding, K.; Ma, F.; Li, C. Functions of dopamine in plants: A review. Plant Signal. Behav. 2020, 15, 1827782. [Google Scholar] [CrossRef]
- Tretyn, A.; Kendrick, R.E. Acetylcholine in plants: Presence, metabolism and mechanism of action. Bot. Rev. 1991, 57, 33–73. [Google Scholar] [CrossRef]
- Qin, C.; Ahanger, M.A.; Lin, B.; Huang, Z.; Zhou, J.; Ahmed, N.; Ai, S.; Mustafa, N.S.A.; Ashraf, M.; Zhang, L. Comparative transcriptome analysis reveals the regulatory effects of acetylcholine on salt tolerance of Nicotiana benthamiana. Phytochemistry 2021, 181, 112582. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.-B.; Dong, X.-R.; Fang, M.-Y.; Wang, Q.; Yan, P.; Wang, Q.-Y.; Lu, L.; Dong, Z.-Q. Effects of basic application of chlorocholine chloride combined with nitrogen fertilizer on nitrogen use of summer maize in North China Plain. J. Appl. Ecol. 2021, 32, 931–941. [Google Scholar] [CrossRef]
- Ju, S.; Xu, D.; Zhang, C.; Lu, J.; Jiang, X.; Ji, L. Induction of tolerance by chlorocholine chloride in Sequoia sempervirens seedlings under natural cooling and drought. J. For. Sci. 2020, 66, 236–243. [Google Scholar] [CrossRef]
- Soumya, P.R.; Kumar, P.; Pal, M. Paclobutrazol: A novel plant growth regulator and multi-stress ameliorant. Indian J. Plant Physiol. 2017, 22, 267–278. [Google Scholar] [CrossRef]
- Zhou, J.; Li, Z.; Xiao, G.; Zhai, M.; Pan, X.; Huang, R.; Zhang, H. OsCYP71D8L as a key regulator involved in growth and stress response by mediating gibberellins homeostasis in rice. J. Exp. Bot. 2019, 71, 1160–1170. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and Waterlogging Stress in Plants: A Review Highlighting Research Opportunities and Understudied Aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef]
- Yang, Y.; Ma, C.; Xu, Y.; Wei, Q.; Imtiaz, M.; Lan, H.; Gao, S.; Cheng, L.; Wang, M.; Fei, Z.; et al. A zinc finger protein regulates flowering time and abiotic stress tolerance in chrysanthemum by modulating gibberellin biosynthesis. Plant Cell 2014, 26, 2038–2054. [Google Scholar] [CrossRef] [Green Version]
- Oliferuk, S.; Ródenas, R.; Pérez, A.; Martínez, V.; Rubio, F.; Santa-María, G.E. DELLAs Contribute to Set the Growth and Mineral Composition of Arabidopsis thaliana Plants Grown Under Conditions of Potassium Deprivation. J. Plant Growth Regul. 2017, 36, 487–501. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, B.; Zhao, Y.; Zhang, J.; Li, Z. Auxin and GA signaling play important roles in the maize response to phosphate deficiency. Plant Sci. 2019, 283, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yao, Q.; Zhang, Y.; Zhang, Y.; Xing, J.; Yang, B.; Mi, G.; Li, Z.; Zhang, M. The role of gibberellins in regulation of nitrogen uptake and physiological traits in maize responding to nitrogen availability. Int. J. Mol. Sci. 2020, 21, 1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, C.; Gao, X.; Liao, L.; Harberd, N.P.; Fu, X. Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. Plant Physiol. 2007, 145, 1460–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Y.; Ren, Y.R.; Zheng, P.F.; Qu, F.J.; Song, L.Q.; You, C.X.; Wang, X.F.; Hao, Y.J. Functional identification of apple MdMYB2 gene in phosphate-starvation response. J. Plant Physiol. 2020, 244, 153089. [Google Scholar] [CrossRef]
- Matayoshi, C.L.; Pena, L.B.; Arbona, V.; Gómez-Cadenas, A.; Gallego, S.M. Early responses of maize seedlings to Cu stress include sharp decreases in gibberellins and jasmonates in the root apex. Protoplasma 2020, 257, 1243–1256. [Google Scholar] [CrossRef]
- Wild, M.; Davière, J.M.; Regnault, T.; Sakvarelidze-Achard, L.; Carrera, E.; Lopez Diaz, I.; Cayrel, A.; Dubeaux, G.; Vert, G.; Achard, P. Tissue-Specific Regulation of Gibberellin Signaling Fine-Tunes Arabidopsis Iron-Deficiency Responses. Dev. Cell 2016, 37, 190–200. [Google Scholar] [CrossRef] [Green Version]
- Iftikhar, A.; Ali, S.; Yasmeen, T.; Arif, M.S.; Zubair, M.; Rizwan, M.; Alhaithloul, H.A.S.; Alayafi, A.A.M.; Soliman, M.H. Effect of gibberellic acid on growth, photosynthesis and antioxidant defense system of wheat under zinc oxide nanoparticle stress. Environ. Pollut. 2019, 254, 113109. [Google Scholar] [CrossRef]
- Bashir, W.; Anwar, S.; Zhao, Q.; Hussain, I.; Xie, F. Interactive effect of drought and cadmium stress on soybean root morphology and gene expression. Ecotoxicol. Environ. Saf. 2019, 175, 90–101. [Google Scholar] [CrossRef]
- Wang, B.; Wei, H.; Zhang, H.; Zhang, W.-H. Enhanced accumulation of gibberellins rendered rice seedlings sensitive to ammonium toxicity. J. Exp. Bot. 2020, 71, 1514–1526. [Google Scholar] [CrossRef]
- Achard, P.; Gong, F.; Cheminant, S.; Alioua, M.; Hedden, P.; Genschika, P. The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell 2008, 20, 2117–2129. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Chen, H.; Wei, D.; Ma, H.; Lin, J. Arabidopsis CBF3 and DELLAs positively regulate each other in response to low temperature. Sci. Rep. 2017, 7, 39819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.I.; Li, P.F.; Yang, C.H. NAC-Like Gene GIBBERELLIN SUPPRESSING FACTOR Regulates the Gibberellin Metabolic Pathway in Response to Cold and Drought Stresses in Arabidopsis. Sci. Rep. 2019, 9, 19226. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, X.; Dong, S.; Jiang, X.; Wang, L.; Yu, J.; Zhou, Y. Crosstalk of PIF4 and DELLA modulates CBF transcript and hormone homeostasis in cold response in tomato. Plant Biotechnol. J. 2020, 18, 1041–1055. [Google Scholar] [CrossRef]
- Toh, S.; Imamura, A.; Watanabe, A.; Nakabayashi, K.; Okamoto, M.; Jikumaru, Y.; Hanada, A.; Aso, Y.; Ishiyama, K.; Tamura, N.; et al. High temperature-induced abscisic acid biosynthesis and its role in the inhibition of gibberellin action in Arabidopsis seeds. Plant Physiol. 2008, 146, 1368–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of Plant Responses to Environmentally Activated Phytohormonal Signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Wang, J.; Qin, H.; Zhou, S.; Wei, P.; Zhang, H.; Zhou, Y.; Miao, Y.; Huang, R. The Ubiquitin-Binding Protein OsDSK2a Mediates Seedling Growth and Salt Responses by Regulating Gibberellin Metabolism in Rice. Plant Cell 2020, 32, 414–428. [Google Scholar] [CrossRef] [Green Version]
- Magome, H.; Yamaguchi, S.; Hanada, A.; Kamiya, Y.; Oda, K. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J. 2008, 56, 613–626. [Google Scholar] [CrossRef]
- Yao, W.; Wang, S.; Zhou, B.; Jiang, T. Transgenic poplar overexpressing the endogenous transcription factor ERF76 gene improves salinity tolerance. Tree Physiol. 2016, 36, 896–908. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Liang, X.; Lv, S.; Guan, T.; Jiang, T.; Cheng, Y. Histone deacetylase gene PtHDT902 modifies adventitious root formation and negatively regulates salt stress tolerance in poplar. Plant Sci. 2020, 290, 110301. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Wang, Q.; Chen, M.; Liu, X.; Gao, D.; Li, D.; Li, L. MdBZR1 and MdBZR1-2like Transcription Factors Improves Salt Tolerance by Regulating Gibberellin Biosynthesis in Apple. Front. Plant Sci. 2019, 10, 1043. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic Acid-Mediated Drought Stress Responses in Plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef] [PubMed]
- Dubois, M.; Skirycz, A.; Claeys, H.; Maleux, K.; Dhondt, S.; De Bodt, S.; Vanden Bossche, R.; De Milde, L.; Yoshizumi, T.; Matsui, M.; et al. ETHYLENE RESPONSE FACTOR6 acts as a central regulator of leaf growth under water-limiting conditions in Arabidopsis. Plant Physiol. 2013, 162, 319–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shohat, H.; Cheriker, H.; Kilambi, H.V.; Illouz Eliaz, N.; Blum, S.; Amsellem, Z.; Tarkowská, D.; Aharoni, A.; Eshed, Y.; Weiss, D. Inhibition of Gibberellin Accumulation by Water Deficiency Promotes Fast and Long-Term ‘Drought Avoidance’ Responses in Tomato. New Phytol. 2021, 232, 1985–1998. [Google Scholar] [CrossRef] [PubMed]
- Nir, I.; Moshelion, M.; Weiss, D. The Arabidopsis GIBBERELLIN METHYL TRANSFERASE 1 suppresses gibberellin activity, reduces whole-plant transpiration and promotes drought tolerance in transgenic tomato. Plant. Cell Environ. 2014, 37, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Illouz-Eliaz, N.; Nissan, I.; Nir, I.; Ramon, U.; Shohat, H.; Weiss, D. Mutations in the tomato gibberellin receptors suppress xylem proliferation and reduce water loss under water-deficit conditions. J. Exp. Bot. 2020, 71, 3603–3612. [Google Scholar] [CrossRef]
- Wu, F.; Sun, X.; Zou, B.; Zhu, P.; Lin, N.; Lin, J.; Ji, K. Transcriptional analysis of masson pine (Pinus massoniana) under high CO2 stress. Genes 2019, 10, 804. [Google Scholar] [CrossRef] [Green Version]
- Fukao, T.; Bailey-Serres, J. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci. USA 2008, 105, 16814–16819. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Lee, L.Y.C.; Xia, K.; Yan, Y.; Yu, H. DELLAs Modulate Jasmonate Signaling via Competitive Binding to JAZs. Dev. Cell 2010, 19, 884–894. [Google Scholar] [CrossRef] [Green Version]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Dhar, N.; Short, D.P.G.; Mamo, B.E.; Corrion, A.J.; Wai, C.M.; Anchieta, A.; VanBuren, R.; Day, B.; Ajwa, H.; Subbarao, K.V.; et al. Arabidopsis defense mutant ndr1-1 displays accelerated development and early flowering mediated by the hormone gibberellic acid. Plant Sci. 2019, 285, 200–213. [Google Scholar] [CrossRef]
- de Vleesschauwer, D.; Seifi, H.S.; Filipe, O.; Haeck, A.; Huu, S.N.; Demeestere, K.; Höfte, M. The DELLA protein SLR1 integrates and amplifies salicylic acid- and jasmonic acid-dependent innate immunity in rice. Plant Physiol. 2016, 170, 1831–1847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Li, Y. Current understanding of the interplays between host hormones and plant viral infections. PLoS Pathog. 2021, 17, e1009242. [Google Scholar] [CrossRef] [PubMed]
- Pan, L.L.; Miao, H.; Wang, Q.; Walling, L.L.; Liu, S.S. Virus-induced phytohormone dynamics and their effects on plant–insect interactions. New Phytol. 2021, 230, 1305–1320. [Google Scholar] [CrossRef]
- Križnik, M.; Petek, M.; Dobnik, D.; Ramšak, Ž.; Baebler, Š.; Pollmann, S.; Kreuze, J.F.; Žel, J.; Gruden, K. Salicylic acid perturbs srna-gibberellin regulatory network in immune response of potato to potato virus y infection. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Shimizu, T.; Kondoh, H.; Hiraguri, A.; Sasaya, T.; Choi, I.R.; Omura, T.; Kikuchi, S. Relationship between symptoms and gene expression induced by the infection of three strains of Rice dwarf virus. PLoS ONE 2011, 6, e18094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, W.; Li, B.; Liu, G.; Wei, Y.; He, C.; Shi, H. Identification and functional analysis of cassava DELLA proteins in plant disease resistance against cassava bacterial blight. Plant Physiol. Biochem. 2018, 124, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.L.; Li, Q.; Deng, Y.W.; Lou, Y.G.; Wang, M.Y.; Zhou, G.X.; Zhang, Y.Y.; He, Z.H. Altered disease development in the eui mutants and Eui overexpressors indicates that gibberellins negatively regulate rice basal disease resistance. Mol. Plant 2008, 1, 528–537. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Shi, Z.; Zhang, X.; Wang, M.; Zhang, L.; Zheng, K.; Liu, J.; Hu, X.; Di, C.; Qian, Q.; et al. Inducible overexpression of Ideal Plant Architecture1 improves both yield and disease resistance in rice. Nat. Plants 2019, 5, 389–400. [Google Scholar] [CrossRef]
- Qin, X.; Liu, J.H.; Zhao, W.S.; Chen, X.J.; Guo, Z.J.; Peng, Y.L. Gibberellin 20-Oxidase Gene OsGA20ox3 regulates plant stature and disease development in rice. Mol. Plant-Microbe Interact. 2013, 26, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Yimer, H.Z.; Nahar, K.; Kyndt, T.; Haeck, A.; Van Meulebroek, L.; Vanhaecke, L.; Demeestere, K.; Höfte, M.; Gheysen, G. Gibberellin antagonizes jasmonate-induced defense against Meloidogyne graminicola in rice. New Phytol. 2018, 218, 646–660. [Google Scholar] [CrossRef] [Green Version]
- Lei, P.; Qi, N.; Zhou, Y.; Wang, Y.; Zhu, X.; Xuan, Y.; Liu, X.; Fan, H.; Chen, L.; Duan, Y. Soybean miR159-GmMYB33 Regulatory Network Involved in Gibberellin-Modulated Resistance to Heterodera glycines. Int. J. Mol. Sci. 2021, 22, 13172. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C.M. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robert-Seilaniantz, A.; Navarro, L.; Bari, R.; Jones, J.D. Pathological hormone imbalances. Curr. Opin. Plant Biol. 2007, 10, 372–379. [Google Scholar] [CrossRef] [PubMed]
- McAdam, E.L.; Reid, J.B.; Foo, E. Gibberellins promote nodule organogenesis but inhibit the infection stages of nodulation. J. Exp. Bot. 2018, 69, 2117–2130. [Google Scholar] [CrossRef]
- Dolgikh, A.V.; Kirienko, A.N.; Tikhonovich, I.A.; Foo, E.; Dolgikh, E.A. The DELLA proteins influence the expression of cytokinin biosynthesis and response genes during nodulation. Front. Plant Sci. 2019, 10, 432. [Google Scholar] [CrossRef]
- Dolgikh, A.V.; Rudaya, E.S.; Dolgikh, E.A. Identification of bell transcription factors involved in nodule initiation and development in the legumes pisum sativum and medicago truncatula. Plants 2020, 9, 1808. [Google Scholar] [CrossRef]
- Yu, N.; Luo, D.; Zhang, X.; Liu, J.; Wang, W.; Jin, Y.; Dong, W.; Liu, J.; Liu, H.; Yang, W.; et al. A della protein complex controls the arbuscular mycorrhizal symbiosis in plants. Cell Res. 2014, 24, 130–133. [Google Scholar] [CrossRef]
- Takeda, N.; Handa, Y.; Tsuzuki, S.; Kojima, M.; Sakakibara, H.; Kawaguchi, M. Gibberellins interfere with symbiosis signaling and gene expression and alter colonization by Arbuscular Mycorrhizal fungi in Lotus Japonicus. Plant Physiol. 2015, 167, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiol. Res. 2020, 239, 126569. [Google Scholar] [CrossRef]
- Tsukanova, K.A.; Chebotar, V.K.; Meyer, J.J.M.; Bibikova, T.N. Effect of plant growth-promoting Rhizobacteria on plant hormone homeostasis. S. Afr. J. Bot. 2017, 113, 91–102. [Google Scholar] [CrossRef]
- Kang, S.M.; Khan, A.L.; You, Y.H.; Kim, J.G.; Kamran, M.; Lee, I.J. Gibberellin production by newly isolated strain Leifsonia soli SE134 and Its potential to promote plant growth. J. Microbiol. Biotechnol. 2014, 24, 106–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, J.; Ruan, Y.; Wang, B.; Zhang, J.; Waseem, R.; Huang, Q.; Shen, Q. Plant growth-promoting rhizobacteria strain bacillus amyloliquefaciens njn-6-enriched bio-organic fertilizer suppressed fusarium wilt and promoted the growth of banana plants. J. Agric. Food Chem. 2013, 61, 3774–3780. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, J.; Sun, H.; Han, X.; Peng, Y.; Liu, J.; Liu, K.; Ding, Y.; Wang, C.; Du, B. Transcriptome Profiles Reveal the Growth-Promoting Mechanisms of Paenibacillus polymyxa YC0136 on Tobacco (Nicotiana tabacum L.). Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Simplified schematic representation of the GAs signaling mechanism. See text for details and references. Adapted from [33].
Figure 1.
Simplified schematic representation of the GAs signaling mechanism. See text for details and references. Adapted from [33].

Figure 2.
Schematic representation of the relations between GAs and other major PGRs. Arrows indicate activation and blunt-end lines indicate repression or inhibition. See text for details and references. ABA: abscisic acid, ACS5/ETO2: ACC synthase 5/ETO2, ACS8: ACC synthase 8, ARF: Auxin Response Factor, BRs: Brassinosteroids, BZR1: Brassinazole Resistant1, CHO1: Chotto1, CKs: Cytokinins, CKX3: Cytokinin dehydrogenase 3, DAD1: Defective anther dehiscence 1, DELLA: Della proteins, DWF4: Dwarf 4, EIN/EIL: ethylene insensitive 3/EIN3-like 1, Et: ethylene, GA: gibberellins, GA2ox: GA2-oxidase, GA3ox: GA3-oxidase, GA20ox: GA20-oxidase, HLS1: Hookless 1, IAA: indole-acetic acid, IBA: indole-butyric acid, IPT3: Isopentenyl transferase 3, JAs: Jasmonates, miRNA160: microRNA160, PIF: Phytochrome interacting factors, PIN: pin-formed, SLs: Strigolactones, SLR1: Slender Rice 1, SUR2: superroot 2.
Figure 2.
Schematic representation of the relations between GAs and other major PGRs. Arrows indicate activation and blunt-end lines indicate repression or inhibition. See text for details and references. ABA: abscisic acid, ACS5/ETO2: ACC synthase 5/ETO2, ACS8: ACC synthase 8, ARF: Auxin Response Factor, BRs: Brassinosteroids, BZR1: Brassinazole Resistant1, CHO1: Chotto1, CKs: Cytokinins, CKX3: Cytokinin dehydrogenase 3, DAD1: Defective anther dehiscence 1, DELLA: Della proteins, DWF4: Dwarf 4, EIN/EIL: ethylene insensitive 3/EIN3-like 1, Et: ethylene, GA: gibberellins, GA2ox: GA2-oxidase, GA3ox: GA3-oxidase, GA20ox: GA20-oxidase, HLS1: Hookless 1, IAA: indole-acetic acid, IBA: indole-butyric acid, IPT3: Isopentenyl transferase 3, JAs: Jasmonates, miRNA160: microRNA160, PIF: Phytochrome interacting factors, PIN: pin-formed, SLs: Strigolactones, SLR1: Slender Rice 1, SUR2: superroot 2.

Figure 3.
Schematic representation of the relations between GAs and potential PGRs. Arrows indicate activation and blunt-end lines indicate repression or inhibition. See text for details and references. ACh: acetylcholine, CCC: clorocholine chloride, DA: dopamine, GA: gibberellins, GABA: γ-aminobutyric acid, H2O2: hydrogen peroxide, NO: nitric oxide, PBZ: paclobutrazol.
Figure 3.
Schematic representation of the relations between GAs and potential PGRs. Arrows indicate activation and blunt-end lines indicate repression or inhibition. See text for details and references. ACh: acetylcholine, CCC: clorocholine chloride, DA: dopamine, GA: gibberellins, GABA: γ-aminobutyric acid, H2O2: hydrogen peroxide, NO: nitric oxide, PBZ: paclobutrazol.

Figure 4.
Schematic representation of the GA-related signaling involved in abiotic stress responses. Arrows indicate activation and blunt-end lines indicate repression or inhibition. Blue-box indicates cold stress, red-box indicates heat stress. See text for details and references. ABA: abscisic acid, ABA1/ZEP: aba1/zeaxanthin epoxidase, ACC: 1-aminociclopropane, AP2/ERF: Apetala2/erf, BBX24: B-box domain protein 24, CBF1/CBF2: cold inducible factor, CO: constans, DDF1: dwarf and delay flowering 2, DELLAs: different DELLA proteins, DREB1/CBF: AP2 dehydration responsive element binding protein, ERF5/6/76: ethylene response factors 5,6 and 76, Et: ethylene, EUI: elongated uppermost internode, FT: flowering locus T, GA2ox: GA2-oxidase, GA3ox: GA3-oxidase, GA20ox: GA20-oxidase, GAs: Gibberellins, GSF: gibberellin suppressor factor, GSF-TM: gibberellin-suppressor factor–transmembrane domain, MPK3/6: mitogen-activated protein kinases 3 and 6, MYB51: MYB domain protein 51, NCED: nine-cis-epoxycarotenoid dioxygenase 2, OsDSK2: Oryza sativa dominant suppressor of kar2, SLR1: slender rice 1, SOC1: suppressor of overexpression of CO1, STZ: salt tolerance zinc-finger, Sub1a: Submergence-1a, WRKY33: WRKY family protein 33.
Figure 4.
Schematic representation of the GA-related signaling involved in abiotic stress responses. Arrows indicate activation and blunt-end lines indicate repression or inhibition. Blue-box indicates cold stress, red-box indicates heat stress. See text for details and references. ABA: abscisic acid, ABA1/ZEP: aba1/zeaxanthin epoxidase, ACC: 1-aminociclopropane, AP2/ERF: Apetala2/erf, BBX24: B-box domain protein 24, CBF1/CBF2: cold inducible factor, CO: constans, DDF1: dwarf and delay flowering 2, DELLAs: different DELLA proteins, DREB1/CBF: AP2 dehydration responsive element binding protein, ERF5/6/76: ethylene response factors 5,6 and 76, Et: ethylene, EUI: elongated uppermost internode, FT: flowering locus T, GA2ox: GA2-oxidase, GA3ox: GA3-oxidase, GA20ox: GA20-oxidase, GAs: Gibberellins, GSF: gibberellin suppressor factor, GSF-TM: gibberellin-suppressor factor–transmembrane domain, MPK3/6: mitogen-activated protein kinases 3 and 6, MYB51: MYB domain protein 51, NCED: nine-cis-epoxycarotenoid dioxygenase 2, OsDSK2: Oryza sativa dominant suppressor of kar2, SLR1: slender rice 1, SOC1: suppressor of overexpression of CO1, STZ: salt tolerance zinc-finger, Sub1a: Submergence-1a, WRKY33: WRKY family protein 33.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Castro-Camba, R.; Sánchez, C.; Vidal, N.; Vielba, J.M. Interactions of Gibberellins with Phytohormones and Their Role in Stress Responses. Horticulturae 2022, 8, 241. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8030241
AMA Style
Castro-Camba R, Sánchez C, Vidal N, Vielba JM. Interactions of Gibberellins with Phytohormones and Their Role in Stress Responses. Horticulturae. 2022; 8(3):241. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8030241
Chicago/Turabian StyleCastro-Camba, Ricardo, Conchi Sánchez, Nieves Vidal, and Jesús Mª Vielba. 2022. "Interactions of Gibberellins with Phytohormones and Their Role in Stress Responses" Horticulturae 8, no. 3: 241. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8030241
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.