
Metabolic Profiling of White and Green Radish Cultivars (Raphanus sativus)
by
 and
and
Chang Ha Park
1,†, 
Wooyeal Ki
2,†,
Nam Su Kim
3,
Soo-Yun Park
4,
Jae Kwang Kim
5,* and
Sang Un Park
2,6,* 1
Department of Biological Sciences, Keimyung University, Daegu 42601, Korea
2
Department of Crop Science, Chungnam National University, 99 Daehak-ro, Yuseong-gu, Daejeon 34134, Korea
3
Korea Research Institute of Bioscience and Biotechnology, 30 Yeongudanji-ro, Ochang-eup, Cheongju-si 28116, Korea
4
National Institute of Agricultural Sciences, Rural Development Administration, Wanju-gun 55365, Korea
5
Division of Life Sciences and Bio-Resource and Environmental Center, Incheon National University, Incheon 22012, Korea
6
Department of Smart Agriculture Systems, Chungnam National University, 99 Daehak-ro, Yuseong-gu, Daejeon 34134, Korea
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Horticulturae 2022, 8(4), 310; https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8040310
Submission received: 24 February 2022
/
Revised: 5 April 2022
/
Accepted: 5 April 2022
/
Published: 7 April 2022
(This article belongs to the Special Issue Physico-Chemical Characterization of Fruits and Vegetables)
Abstract
:Radish (Raphanus sativus) is a Brassica vegetable important for human nutrition and health because it is rich in diverse metabolites. Although previous studies have evaluated various metabolites, few studies have comprehensively profiled the primary and secondary metabolites in the roots of white- and green-colored radishes. Thus, this study aimed to provide information about the contents of metabolites beneficial for human health in both cultivars and to investigate the relationships between the various metabolites detected. In particular, among the 55 metabolites detected in radish roots, the levels of most amino acids and phenolic acids, vital to nutrition and health, were higher in green radish roots, while slightly higher levels of glucosinolates were observed in white radish roots—information which can be used to develop an effective strategy to promote vegetable consumption. Furthermore, glutamic acid, as a metabolic precursor of amino acids and chlorophylls, was positively correlated with other amino acids (cysteine, tryptophan, asparagine, alanine, serine, phenylalanine, valine, isoleucine, proline, leucine, beta-alanine, lysine, and GABA), and chlorophylls (chlorophyll a and chlorophyll b) detected in radish roots and phenylalanine, a metabolic precursor of phenolic compounds, were positively correlated with kaempferol, 4-hydroxybenzoate, and catechin. In addition, strong positive correlations between carbohydrates (sucrose and glucose) and phenolics were observed in this study, indicating that sucrose and glucose function as energy sources for phenolic compounds.
1. Introduction
Plant species of the Brassicaceae family have long been recognized as beneficial for human health. They are the most commonly grown vegetables worldwide because of their high contents of vitamins, carotenoids, tocopherol, minerals, folate, amino acids, carbohydrates, dietary fiber, and bioactive compounds, namely, glucosinolates and phenylpropanoids. They have commercial importance as components of the daily human diet and as a primary source for the vegetable oil industry [1,2,3].
The Brassicaceae family includes many important vegetable crops, including Brassica rapa (Chinese cabbage), Brassica juncea (mustard), Brassica oleracea (cabbage, kale, broccoli, cauliflower, kohlrabi, and brussels sprouts), and Raphanus sativus (radish). The intake of these cruciferous vegetables plays a crucial role in antioxidation, improving the immune system, and preventing aging-related and cardiovascular diseases, as well as diabetes and cancers. These health benefits are related to antitoxic, anticancer, antioxidant, antidiabetic, and anti-inflammatory effects, and are mainly associated with the properties of secondary metabolites, including phenylpropanoids, carotenoids, vitamins, and glucosinolates, present in the Brassicaceae family [1,2,3,4,5].
Plant metabolites can be divided into primary and secondary metabolites. However, their borders are ambiguous because intermediate metabolites generated in primary metabolism are crucial for secondary metabolism, and primary metabolites and secondary metabolites show functional overlap [6]. Generally, primary metabolites can be considered endogenous compounds, such as amino acids, organic acids, and sugars, which are directly involved in basic plant life [7,8,9]. Furthermore, plant sugars, amino acids, and proteins are crucial for human nutrition. Simple sugars, such as glucose, fructose, and sucrose, are commonly found in plants. Of these, glucose and sucrose play important roles in the regulation of gene expression associated with metabolism, stress responses, and the growth and development of many plants [10,11,12,13]. It has been demonstrated that the above two sugars are involved in the synthesis of polyphenols [14,15]. Furthermore, the carbon molecules in glucose and fructose are reported to be directed into diverse metabolic pathways as a result of the spatiotemporal activities of corresponding metabolizing enzymes [16,17,18]. The enzymes involved in sugar metabolism appear to play a critical role in carbon allocation and partitioning in plants [16,17]. Secondary metabolites can be biosynthesized from primary metabolites, such as phenylalanine, tyrosine, or tryptophan, and can provide competitiveness when plants are exposed to unfavorable conditions due to biotic or abiotic factors [8]. Glucosinolates and phenolic compounds are secondary metabolites commonly present in species of the family Brassicaceae. Glucosinolates are sulfur-containing compounds that can be subdivided into three groups: aliphatic, aromatic, and indolic glucosinolates, based on their precursors (aliphatic amino acids (methionine, isoleucine, leucine, or valine), aromatic amino acids (phenylalanine or tyrosine), and tryptophan) [19]. Furthermore, glucosinolates possess health-beneficial properties (e.g., antioxidant, anti-inflammatory, and anti-angiogenic effects) [20], and these sulfur compounds are responsible for the bitter taste and spicy flavor found in radish [21]. Most phenolic compounds are derived from amino acids (phenylalanine or tyrosine) and contribute to plant defense and reproduction by providing resistance against various stresses or attracting pollinators via pigmentation [22]. In addition, the intake of plant phenolics is recommended to prevent chronic diseases [23].
Radish (Raphanus sativus L.) has been continuously consumed as a root vegetable, and various radish cultivars have been developed since it was domesticated in Europe in pre-Roman times [24]. White- and green-colored radishes, otherwise called Mu, have been widely used in traditional Korean cooking for more than 1500 years [25]. Before cooking, Mu is often grated or cut into small pieces to ensure optimal preparation of this cruciferous vegetable. In Korea, people will often eat Mu with boiled fish, soy sauce, and Korean fermented soybean. Various organs of the radish, including the seeds, leaves, and roots, have been used for therapeutic purposes. The annual yield of radish in Korea is 4.5 million tons. Previous studies reported the presence of various metabolites, including amino acids, carbohydrates, organic acids, glucosinolates, phenolics, and carotenoids, in different radish cultivars [26,27,28,29]. Nonetheless, to date, there have been no studies on metabolite profiles in green and white radish cultivars (Figure 1) using high-performance liquid chromatography (HPLC) and gas chromatography–time-of-flight mass spectrometry (GC–TOFMS). Therefore, this study aimed to provide information on the health-beneficial metabolites present in these two radish cultivars using HPLC- and GC–TOFMS-based metabolite analysis and investigated the relationships between metabolites found in these cultivars.
2. Materials and Methods
2.1. Plant Materials
The seeds of white-colored (exhibiting white color at the roots) and green-colored (exhibiting green color at the roots) radishes were obtained from Asia Seed Co., Ltd., Seoul, Korea. Seeds were cultivated at an experimental farm of the National Institute of Horticultural and Herbal Science, Wanju-gun, Korea. The roots were harvested after 70 days. Analyses were performed for three biological replicates, each consisting of 3–8 white- and green-colored radish roots. The roots of each radish cultivar were frozen in liquid nitrogen and freeze-dried for phytochemical analysis.
2.2. HPLC Analysis of Desulfoglucosinolates
Desulfoglucosinolates were extracted and analyzed as described in our previous study [28]. Freeze-dried sample powders (100 mg) of white- and green-colored radish roots were extracted with 1.5 mL of methanol (70%, v/v) at 70 °C and then maintained at 70 °C for 5 min in a water bath, followed by centrifugation at 11,000 rpm for 15 min. The supernatants were transferred into fresh tubes. Afterward, the pellets were extracted twice more using the same extraction protocol, and then the collected extract of radish samples was loaded onto a column packed with DEAE-Sephadex A-25. After adding an arylsulfatase solution (75 µL) for desulfation, the desulfoglucosinolates were eluted with 1.5 mL of water and then syringe-filtered into a vial. The system, program, and conditions used to isolate and quantify individual glucosinolates were based on our previous study [28], and the detailed HPLC conditions for analyzing the desulfoglucosinolates are described in Table S1.
2.3. HPLC Analysis of Phenolic Compounds
Phenolic compounds were extracted and analyzed as described in our previous study [28]. Freeze-dried sample powders (100 mg) of white- and green-colored radish roots were extracted with 1 mL of methanol (70%, v/v) and then sonicated for 65 min, followed by centrifugation at 11,000 rpm for 15 min. The supernatants were transferred into fresh tubes. Afterward, the pellets were extracted twice using the same extraction protocol and then dried with nitrogen gas. The dried samples were re-extracted with 1 mL of methanol, and the resulting extracts were filtered through a syringe and collected in a vial. The system, program, and conditions used to isolate and quantify phenolic compounds were based on the method described in our previous study [28], and the detailed HPLC conditions for analyzing the phenolic compounds are described in Table S1.
2.4. Chlorophyll Analysis
Approximately 100 mg of ground powder root samples of white- and green-colored radishes were mixed with 2 mL of methanol. The mixtures were incubated in a water bath (HB-205SW, Hanbeak Scientific Co., Bucheon, Korea) for 30 min, followed by vigorous vortexing for 20 s. The resultant mixtures were centrifuged and filtered under sterilized conditions. The chlorophyll contents of the crude extracts were measured at 665 and 652 nm wavelengths, as described by Lichtenthaler and Buschmann [30].
2.5. GC–TOFMS Analysis
Freeze-dried sample powders (10 mg) of white- and green-colored radish roots were mixed with 0.45 mL of methanol with ribitol (0.2 g L−1) at 1300 rpm and 37 °C for 2 min and then centrifuged at 11,000 rpm for 20 min. The supernatants were added to 190 μL of chloroform and 480 μL of water and mixed vigorously for 30 s, followed by centrifugation at 11,000 rpm for 15 min. The polar phases (450 μL) were transferred to tubes and freeze-dried in a concentrator. After adding 40 μL of methoxyamine hydrochloride pyridine (30 g L−1), the extracts were shaken at 990 rpm and 37 °C for 120 min, subjected to the addition of N-Methyl-N-(trimethylsilyl) trifluoroacetamide (70 μL), then shaken at 990 rpm and 37 °C for 30 min. The extracts were transferred to vials for analysis of hydrophilic compounds. The system and conditions for analyzing and identifying hydrophilic compounds were determined using the protocol reported in our previous study [28], and the detailed GC–TOFMS conditions for analyzing the hydrophilic metabolites are described in Table S1.
2.6. Statistical Analysis
Student’s t-test and multivariate analysis (PCA, PLS-DA, and HCA with Pearson correlation) were performed using SAS software version 9.2 (SAS Institute Inc., Cary, NC, USA) and MetaboAnalyst 5.0 (http://www.metaboanalyst.ca/, accessed on 7 February 2022).
3. Results
3.1. Desulfoglucosinolate Analysis
A total of seven glucosinolates (glucoraphanin, glucoalyssin, gluconapin, 4-hydroxyglucobrassicin, glucoerucin, glucoraphasatin, and 4-methoxyglucobrassicin) were quantified in the white and green radish roots (Table 1), and the sum of these glucosinolates was found to be higher in white radish roots. Specifically, the levels of gluconapin, 4-hydroxyglucobrassicin, glucoerucin, and glucoraphasatin were slightly higher in white radish roots than in green radish roots. However, the levels of glucoalyssin and 4-methoxyglucobrassicin were slightly higher in green radish roots.
3.2. Phenolic Content
A total of four phenolics (gallic acid, catechin, 4-hydroxybenzoic acid, and kaempferol) were quantified in the white and green radish roots (Table 2). Specifically, the levels of gallic acid, catechin, 4-hydroxybenzoic acid, and kaempferol were slightly higher in green radish roots than in white radish roots (p < 0.05).
3.3. Chlorophyll Content
Chlorophyll a and chlorophyll b contents were measured in the white and green radish roots (Table 3). Green radish roots had much higher chlorophyll a and chlorophyll b levels than white radish roots, as shown in Figure S1. As chlorophyll reflects green light, this difference corresponded to the phenotypic differences.
3.4. GC–TOFMS Analysis
Forty-two metabolites (amino acids, carbohydrates, organic acids, and sugar alcohols) were identified in white- and green-colored radish roots using GC–TOFMS (Table S2). As shown in Figure S1, the levels of most amino acids (alanine, valine, leucine, isoleucine, proline, serine, β-alanine, 4-aminobutyric acid, cysteine, arginine, glutamic acid, phenylalanine, asparagine, glutamine, lysine, and tryptophan) were significantly higher in green radish roots, except pyroglutamic acid. Similarly, the green cultivar contained higher levels of carbohydrates (sucrose and raffinose), sugar alcohols (mannitol, inositol, and glycerol), organic acids (threonic acid and shikimic acid), and TCA intermediates (fumaric acid, malic acid, and citric acid). In contrast, the white cultivar had higher contents of organic acids (pyruvic acid, lactic acid, nicotinic acid, glyceric acid, and succinic acid).
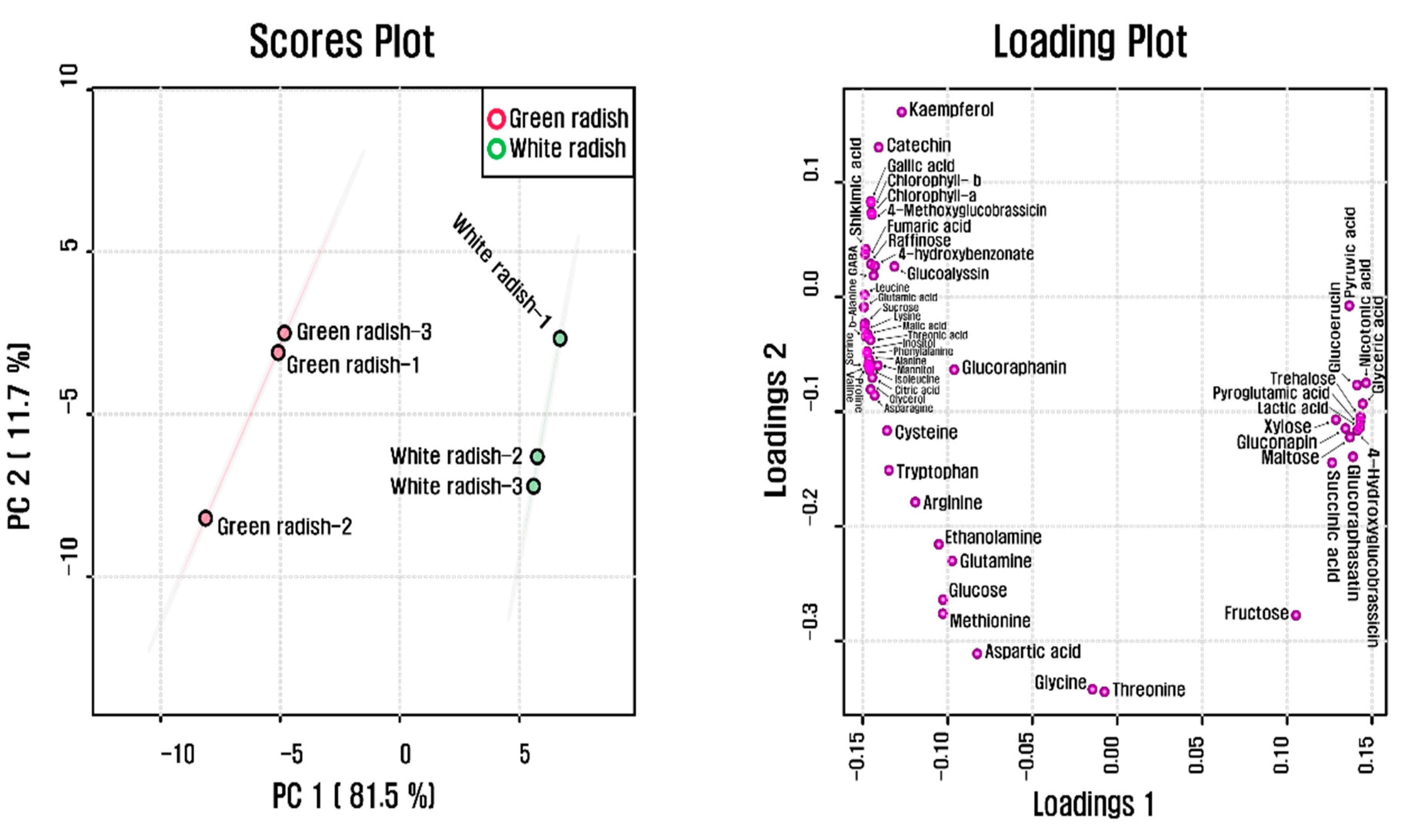
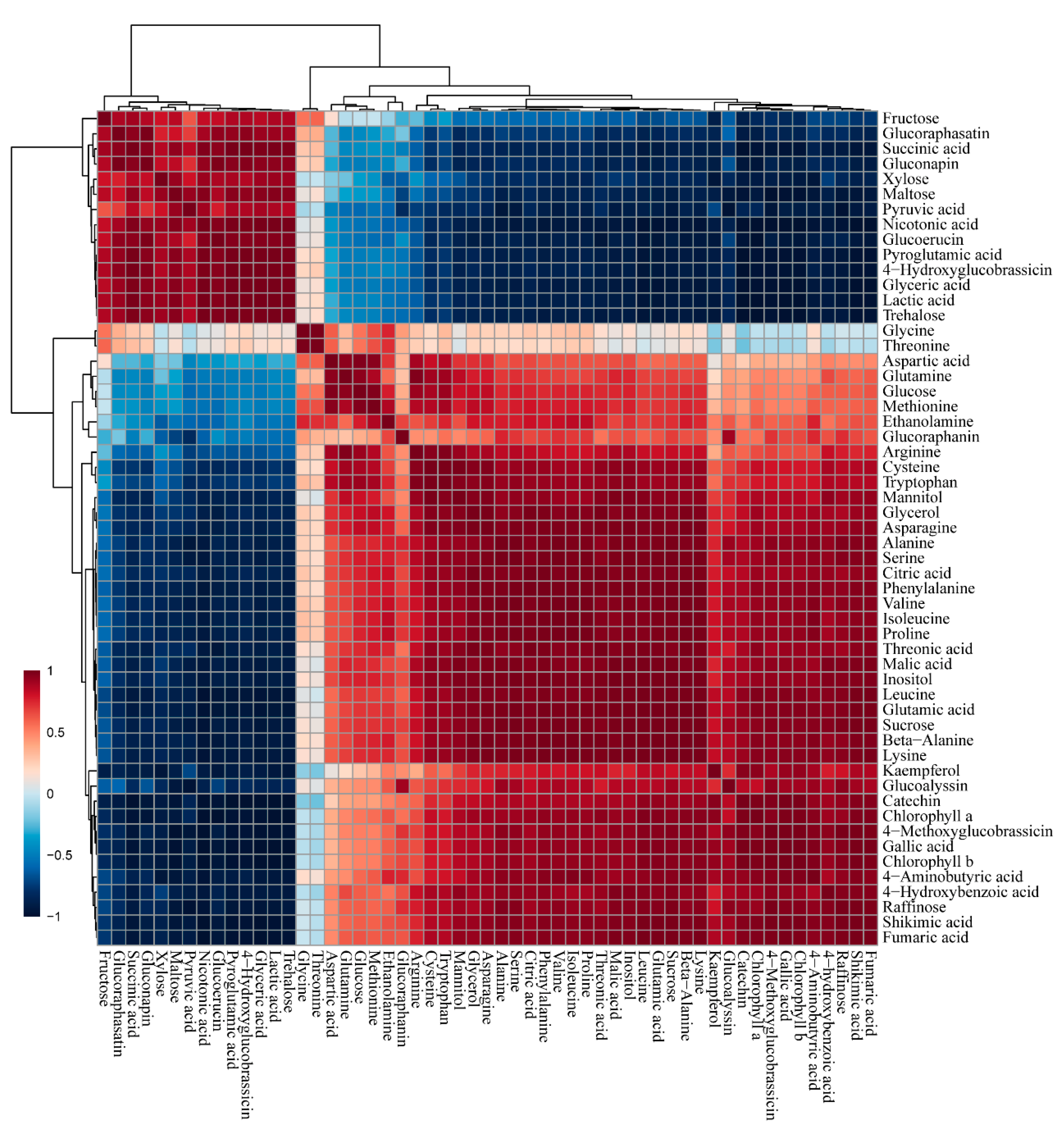
A total of 55 phytochemicals detected in both radish cultivars were subjected to PCA, which showed a clear separation between white and green radish cultivars in PC1 and PC2, explaining 93.2% of the total variance (Figure 2). The separation was due to amino acids, carbohydrates, organic acids, and TCA intermediates. According to the results from the Pearson correlation analysis (Figure 3), these metabolites were clustered into two groups, one of which comprised most organic acids, amino acids, and phenolic compounds, while the other included several carbohydrates and glucosinolates. Glutamine, glutamate, aspartate, and asparagine, consisting of metabolic networks responsible for amino acid metabolism, showed a strong positive correlation with the other amino acids. In particular, glutamic acid, which has a central position in amino acid metabolism, was highly correlated with cysteine (r = 0.91898, p = 0.0095806), tryptophan (r = 0.91056, p = 0.011641), asparagine (r = 0.97936, p = 0.0006349), alanine (r = 0.97812, p = 0.00071283), serine (r = 0.98507, p = 0.00033263), phenylalanine (r = 0.99026, p = 0.00014174), valine (r = 0.98204, p = 0.00048069), isoleucine (r = 0.97868, p = 0.00067718), proline (r = 0.97915, p = 0.00064742), leucine (r = 0.99783, p = 0.0000070689), beta-alanine (r = 0.99494, p = 0.000038297), lysine (r = 0.99886, p = 0.0000019387), and GABA (r = 0.95723, p = 0.0027053). Furthermore, glutamic acid was strongly positively correlated with chlorophyll a (r = 0.96751, p = 0.00157) and chlorophyll b (r = 0.96515, p = 0.0018). Phenylalanine, a precursor of phenolic compounds, and sucrose, known to promote flavonoid biosynthesis, were positively correlated with 4-hydroxybenzonate, catechin, and kaempferol (r > 0.8, p < 0.05).
4. Discussion
The differences in metabolites between white and green radish roots were investigated in this study. A total of 55 metabolites, including amino acids, sugars, sugar alcohols, organic acids, glucosinolates, chlorophylls, phenolics, and amines, were identified in both radish cultivars. According to the primary metabolite profiles, the levels of amino acids, chlorophylls, and organic acids were significantly higher in green radish roots than in white radish roots. In particular, green radish roots are richer in essential amino acids (lysine, leucine, isoleucine, valine, phenylalanine, and tryptophan) that are not biosynthesized in the human body and should be supplemented by food [31]; they contain higher levels of GABA, which reduces stress and promotes sleep in humans [32], and proline, which plays a role in wound healing [33]. Additionally, chlorophyll a and b levels were much higher in green radish roots, revealing that green coloration in the roots is due to chlorophyll pigments. Therefore, the intake of green radish roots can benefit human health and they are richer in amino acids, chlorophylls, and organic acids than white radish roots.
According to the secondary metabolite profiles, four phenolics (gallic acid, catechin, 4-hydroxybenzoic acid, and kaempferol) and seven glucosinolates (glucoraphanin, glucoalyssin, gluconapin, 4-methoxyglucobrassicin, 4-hydroxyglucobrassicin, glucoraphasatin, and glucoerucin) were detected in both cultivars. The levels of phenolic compounds (gallic acid, catechin, 4-hydroxybenzoic acid, and kaempferol) were significantly higher in green radish roots, whereas white radish roots contained higher levels of glucosinolates (4-hydroxyglucobrassicin, glucoraphasatin, gluconapin, and glucoerucin). These findings were consistent with previous studies that identified glucosinolates and phenolic compounds in various radish cultivars. Park et al. [28] detected glucosinolates (glucoraphasatin, glucoerucin, 4-methoxyglucobrassicin, and 4-hydroxyglucobrassicin) and flavonoids (catechin and kaempferol) in the roots of white radishes [16]. Wang et al. [34] identified glucoraphanin, glucoalyssin, and gluconapin in radish roots [23]. In addition, the analysis of glucosinolates in three radish cultivars (Seo Ho, Man Tang Hong, and Hong Feng No. 1) revealed that the content of glucoraphasatin was significantly higher in all the cultivars examined compared to other glucosinolates [35]. Moreover, the analysis of aliphatic glucosinolates in the Raphanus species (R. sativus, R. raphanistrum, R. sativus L. var. raphanistroides) revealed that the content of glucoraphasatin was much higher, while the lowest content was detected for gluconapin [36]. Similar to the previous report, the content of glucoraphasatin was highest, whereas the content of gluconapin was lowest in both cultivars. Bajkacz et al. [37] reported the presence of 4-hydroxybenzoic acid in radish roots; Goyeneche et al. [38] identified gallic acid in radish roots. In line with the previous study [39], catechin was found to be the most abundant phenolic compound in R. sativus root extracts. Furthermore, the current and previous studies found that secondary metabolite contents varied between the roots of both cultivars. This may be due to a combination of specificity, environmental factors, or climatic factors that affect the quality and quantity of phytochemicals.
In this study, the levels of most amino acids were significantly higher in the green radish roots, and glutamic acid was positively correlated with amino acids (cysteine, tryptophan, asparagine, alanine, serine, phenylalanine, valine, isoleucine, proline, leucine, beta-alanine, lysine, and GABA), revealing that glutamic acid plays a central role in plant amino acid metabolism. Similarly, previous studies have reported a strong positive correlation between glutamic acid and other amino acids in Morus alba fruits [40], Lycoris radiata flowers [22], and Brassica rapa L. var. japonica plants [41]. Furthermore, the larger pools of glutamic acid and phenylalanine, which are initial precursors for chlorophyll metabolism and phenolic compound metabolism, respectively, reflected the higher levels of chlorophyll a and chlorophyll b and phenolic compounds detected in green radish roots, supported by the strong positive correlations between glutamic acid and chlorophylls and between phenylalanine and phenolics, respectively. These findings are in agreement with previous studies describing the increased pool of endogenous phenylalanine, derived from the cessation of cell division, enhanced anthocyanin biosynthesis in suspension cultures of Vitis sp. via upregulation of phenylalanine ammonia-lyase (PAL) and chalcone synthase (CHS) expression [42], and exogenous supply of phenylalanine improved the production of phenolic compounds in Pisum sativum leaves via an increase in PAL activity [43]. Furthermore, the endogenous abundance of phenylalanine has been reported to be positively correlated with phenolics in the roots, leaves, and stems of Lavandula pubescens [44], the roots of xBrassicoraphanus, Chinese cabbage and radish [28], and the flowers of Lycoris radiata [22].
Sucrose and glucose function as energy sources for phenolic compounds. Strong correlations between carbohydrates (sucrose and glucose) and phenolic compounds were observed in this study, suggesting that sucrose and glucose positively impact phenolic compounds. Our findings are in agreement with those of Shin et al. [45], who reported that an increase in endogenous sucrose abundance, derived from calcium signaling, enhanced anthocyanin content with upregulation of flavonoid biosynthesis gene expression, and those of Zakhleniuk et al. [46], who showed that the increased abundance of glucose and sucrose, induced by a sucrose transporter 2 (SUC2) mutation, was associated with increased accumulation of anthocyanins. Park et al. [47] reported that the enhanced pool of sucrose, caused by AtMYB75 overexpression, was positively correlated with flavones in Scutellaria baicalensis hairy roots. The exogenous application of sucrose induced the upregulation of anthocyanin biosynthesis in radish hypocotyls [48], while sucrose-dependent enhancement of anthocyanin production was observed in Arabidopsis expressing petunia CHS-A [49], and an increase in anthocyanin accumulation by sucrose was recorded in yellow lupine cells [50]. Furthermore, a positive correlation between sucrose and phenolic compounds has been reported in several Brassica vegetables, such as Chinese cabbage (Brassica rapa ssp. pekinensis) [51] and mizuna (Brassica rapa L. var. japonica) [41].
5. Conclusions
In conclusion, this is the first study to profile metabolites in the roots of white- and green-colored radishes and investigate the metabolic relationship between diverse metabolites detected in both cultivars. Among the 55 metabolites detected in green- and white-colored radishes, the levels of most amino acids, organic acids, phenolic compounds, and chlorophylls, which are important for nutrition and health, were higher in green radishes. In contrast, the white radish contained slightly higher levels of glucosinolates. Furthermore, metabolic precursors (glutamic acid and phenylalanine) were positively correlated with the levels of other amino acids, chlorophylls, and phenolics, and carbohydrates (sucrose and glucose), acting as energy sources, were positively correlated with phenolics. The metabolic profiles of Korean white- and green-colored radish roots associated with primary metabolites (amino acids, carbohydrates, and organic acids) and secondary metabolites (chlorophyll, desulfoglucosinolate, and phenolics) will provide valuable information and help to lay the groundwork for human health.
Supplementary Materials
The following supporting information can be downloaded at: https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/horticulturae8040310/s1, Figure S1: Heatmap representing changes in metabolite composition of green and white radish roots, Table S1: Conditions for phytochemical analysis of metabolites detected in this study, Table S2: Metabolites identified in Korean white- and green-colored radish roots using GC–TOFMS.
Author Contributions
Conceptualization, J.K.K. and S.U.P.; methodology, J.K.K. and S.-Y.P.; formal analysis, J.K.K., C.H.P. and N.S.K.; investigation, C.H.P., N.S.K. and W.K.; resources, S.-Y.P.; data curation, C.H.P. and W.K.; writing—original draft preparation, C.H.P. and W.K.; writing—review and editing, C.H.P. and W.K.; visualization, C.H.P.; supervision, S.U.P.; project administration, S.U.P.; funding acquisition, S.U.P. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the Next-generation BioGreen 21 Program (PJ015665), Korea.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data reported are available in the Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Björkman, M.; Klingen, I.; Birch, A.N.; Bones, A.M.; Bruce, T.J.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health–Influences of climate, environment and agronomic practice. Phytochemistry 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Perles, R.; Mena, P.; Garcia-Viguera, C.; Moreno, D. Brassica foods as a dietary source of vitamin C: A review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1076–1091. [Google Scholar] [CrossRef] [PubMed]
- Jahangir, M.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Health-affecting compounds in Brassicaceae. Compr. Rev. Food Sci. Food Saf. 2009, 8, 31–43. [Google Scholar] [CrossRef]
- Podsędek, A. Natural antioxidants and antioxidant capacity of Brassica vegetables: A review. LWT Food Sci. Technol. 2007, 40, 1–11. [Google Scholar] [CrossRef]
- Soengas, P.; Sotelo, T.; Velasco, P.; Cartea, M.E. Antioxidant properties of Brassica vegetables. Funct. Plant Sci. Biotechnol. 2011, 5, 43–55. [Google Scholar]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an emerging tool for the study of plant–pathogen interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef] [Green Version]
- Hussein, R.A.; El-Anssary, A.A. Plants secondary metabolites: The key drivers of the pharmacological actions of medicinal plants. Herb. Med. 2019, 1, 13. [Google Scholar]
- Park, C.H.; Yeo, H.J.; Kim, N.S.; Eun, P.Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Park, S.-Y.; Kim, J.K.; Park, S.U. Metabolic profiling of pale green and purple kohlrabi (Brassica oleracea vargongylodes). Appl. Biol. Chem. 2017, 60, 249–257. [Google Scholar] [CrossRef]
- Bolouri-Moghaddam, M.R.; Roy, K.L.; Xiang, L.; Rolland, F.; Ende, W.V.D. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rolland, F.; Winderickx, J.; Thevelein, J.M. Glucose-sensing mechanisms in eukaryotic cells. Trends Biochem. Sci. 2001, 26, 310–317. [Google Scholar] [CrossRef]
- Feduraev, P.; Chupakhina, G.; Maslennikov, P.; Tacenko, N.; Skrypnik, L. Variation in phenolic compounds content and antioxidant activity of different plant organs from Rumex crispus L. and Rumex obtusifolius L. at different growth stages. Antioxidants 2019, 8, 237. [Google Scholar] [CrossRef] [Green Version]
- Nešović, M.; Gašić, U.; Tosti, T.; Horvacki, N.; Nedić, N.; Sredojević, M.; Blagojević, S.; Ignjatović, L.; Tešić, Ž. Distribution of polyphenolic and sugar compounds in different buckwheat plant parts. RSC Adv. 2021, 11, 25816–25829. [Google Scholar] [CrossRef]
- Bar-Peled, M.; O’Neill, M.A. Plant nucleotide sugar formation, interconversion, and salvage by sugar recycling. Annu. Rev. Plant Biol. 2011, 62, 127–155. [Google Scholar] [CrossRef] [Green Version]
- Gibeaut, D.M. Nucleotide sugars and glycosyltransferases for synthesis of cell wall matrix polysaccharides. Plant Physiol. Biochem. 2000, 38, 69–80. [Google Scholar] [CrossRef]
- Tsai, C.J.; Harding, S.A.; Tschaplinski, T.J.; Lindroth, R.L.; Yuan, Y. Genome-wide analysis of the structural genes regulating defense phenylpropanoid metabolism in Populus. New Phytol. 2006, 172, 47–62. [Google Scholar] [CrossRef]
- Miękus, N.; Marszałek, K.; Podlacha, M.; Iqbal, A.; Puchalski, C.; Świergiel, A.H. Health benefits of plant-derived sulfur compounds, glucosinolates, and organosulfur compounds. Molecules 2020, 25, 3804. [Google Scholar] [CrossRef]
- Esteve, M. Mechanisms underlying biological effects of cruciferous glucosinolate-derived isothiocyanates/indoles: A focus on metabolic syndrome. Front. Nutr. 2020, 7, 111. [Google Scholar] [CrossRef]
- Bell, L.; Oloyede, O.O.; Lignou, S.; Wagstaff, C.; Methven, L. Taste and flavor perceptions of glucosinolates, isothiocyanates, and related compounds. Mol. Nutr. Food Res. 2018, 62, 1700990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, C.H.; Yeo, H.J.; Kim, Y.J.; Nguyen, B.V.; Park, Y.E.; Sathasivam, R.; Kim, J.K.; Park, S.U. Profiles of secondary metabolites (phenolic acids, carotenoids, anthocyanins, and galantamine) and primary metabolites (carbohydrates, amino acids, and organic acids) during flower development in Lycoris radiata. Biomolecules 2021, 11, 248. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The role of polyphenols in human health and food systems: A mini-review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Wang, J.; Qiu, Y.; Wang, H.; Wang, P.; Zhang, X.; Li, C.; Song, J.; Gui, W.; Shen, D. SSR-sequencing reveals the inter-and intraspecific genetic variation and phylogenetic relationships among an extensive collection of Radish (Raphanus) germplasm resources. Biology 2021, 10, 1250. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.-J.; Moon, S.-K.; Kim, C.-H. Raphanus sativus and its isothiocyanates inhibit vascular smooth muscle cells proliferation and induce G1 cell cycle arrest. Int. Immunopharmacol. 2006, 6, 854–861. [Google Scholar] [CrossRef]
- Gamba, M.; Asllanaj, E.; Raguindin, P.F.; Glisic, M.; Franco, O.H.; Minder, B.; Bussler, W.; Metzger, B.; Kern, H.; Muka, T. Nutritional and phytochemical characterization of radish (Raphanus sativus): A systematic review. Trends Food Sci. Technol. 2021, 113, 205–218. [Google Scholar] [CrossRef]
- Park, C.H.; Baskar, T.B.; Park, S.-Y.; Kim, S.-J.; Arasu, M.V.; Al-Dhabi, N.A.; Kim, J.K.; Park, S.U. Metabolic profiling and antioxidant assay of metabolites from three radish cultivars (Raphanus sativus). Molecules 2016, 21, 157. [Google Scholar] [CrossRef] [Green Version]
- Park, C.H.; Park, S.-Y.; Park, Y.J.; Kim, J.K.; Park, S.U. Metabolite profiling and comparative analysis of secondary metabolites in Chinese cabbage, radish, and hybrid xBrassicoraphanus. J. Agric. Food Chem. 2020, 68, 13711–13719. [Google Scholar] [CrossRef]
- Zhang, J.; Qiu, X.; Tan, Q.; Xiao, Q.; Mei, S. A comparative metabolomics study of flavonoids in radish with different skin and flesh colors (Raphanus sativus L.). J. Agric. Food Chem. 2020, 68, 14463–14470. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4. 3.1–F4. 3.8. [Google Scholar] [CrossRef]
- Lopez, M.J.; Mohiuddin, S.S. Biochemistry, Essential Amino Acids; StatPearls Publishing: Tampa, FL, USA, 2020. [Google Scholar]
- Hepsomali, P.; Groeger, J.A.; Nishihira, J.; Scholey, A. Effects of oral gamma-aminobutyric acid (GABA) administration on stress and sleep in humans: A systematic review. Front. Neurosci. 2020, 14, 923. [Google Scholar] [CrossRef] [PubMed]
- Ponrasu, T.; Jamuna, S.; Mathew, A.; Madhukumar, K.N.; Ganeshkumar, M.; Iyappan, K.; Suguna, L. Efficacy of L-proline administration on the early responses during cutaneous wound healing in rats. Amino Acids 2013, 45, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, Q.; Sun, H.; Zhang, Z.; Qian, H.; Zhao, X.; He, H.; Zhang, L. Glucosinolate profiles in different organs of 111 radish accessions and candidate genes involved in converting glucobrassicin to 4-Hydroxyglucobrassicin. J. Agric. Food Chem. 2022, 70, 2. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-J.; Uddin, M.R.; Park, S.U. Glucosinolate accumulation in three important radish (Raphanus sativus) cultivars. Aust. J. Crop Sci. 2013, 7, 1843–1847. [Google Scholar]
- Ko, H.-C.; Sung, J.-S.; Hur, O.-S.; Baek, H.-J.; Jeon, Y.-A.; Luitel, B.P.; Ryu, K.-Y.; Kim, J.-B.; Rhee, J.-H. Comparison of glucosinolate contents in leaves and roots of radish (Raphanus spp.). Korean J. Plant Res. 2017, 30, 579–589. [Google Scholar]
- Bajkacz, S.; Ligor, M.; Baranowska, I.; Buszewski, B. Separation and determination of chemopreventive phytochemicals of flavonoids from Brassicaceae plants. Molecules 2021, 26, 4734. [Google Scholar] [CrossRef] [PubMed]
- Goyeneche, R.; Roura, S.; Ponce, A.; Vega-Gálvez, A.; Quispe-Fuentes, I.; Uribe, E.; Scala, K.D. Chemical characterization and antioxidant capacity of red radish (Raphanus sativus L.) leaves and roots. J. Funct. Foods 2015, 16, 256–264. [Google Scholar] [CrossRef]
- Beevi, S.S.; Mangamoori, L.N.; Gowda, B.B. Polyphenolics profile and antioxidant properties of Raphanus sativus L. Nat. Prod. Res. 2012, 26, 557–563. [Google Scholar] [CrossRef]
- Park, C.H.; Park, Y.E.; Yeo, H.J.; Yoon, J.S.; Park, S.-Y.; Kim, J.K.; Park, S.U. Comparative analysis of secondary metabolites and metabolic profiling between diploid and tetraploid Morus alba L. J. Agric. Food Chem. 2021, 69, 1300–1307. [Google Scholar] [CrossRef]
- Park, C.H.; Bong, S.J.; Lim, C.J.; Kim, J.K.; Park, S.U. Transcriptome analysis and metabolic profiling of green and red mizuna (Brassica rapa L. var. japonica). Foods 2020, 9, 1079. [Google Scholar] [CrossRef]
- Sakuta, M.; Hirano, H.; Kakegawa, K.; Suda, J.; Hirose, M.; Joy, R.W.; Sugiyama, M.; Komamine, A. Regulatory mechanisms of biosynthesis of betacyanin and anthocyanin in relation to cell division activity in suspension cultures. Plant Cell Tissue Organ Cult. 1994, 38, 167–169. [Google Scholar] [CrossRef]
- Singh, D.; Bahadur, A.; Sarma, B.; Maurya, S.; Singh, H.; Singh, U. Exogenous application of L-phenylalanine and ferulic acid enhance phenylalanine ammonia lyase activity and accumulation of phenolic acids in pea (Pisum sativum) to offer protection against Erysiphe pisi. Arch. Phytopathol. Plant Prot. 2010, 43, 1454–1462. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, Y.E.; Kim, Y.J.; Park, C.; Kim, J.K.; Park, S.U. Integrated analysis of transcriptome and metabolome and evaluation of antioxidant activities in Lavandula pubescens. Antioxidants 2021, 10, 1027. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Choi, M.-G.; Lee, H.K.; Cho, M.; Choi, S.-B.; Choi, G.; Park, Y.-I. Calcium dependent sucrose uptake links sugar signaling to anthocyanin biosynthesis in Arabidopsis. Biochem. Biophys. Res. Commun. 2013, 430, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Zakhleniuk, O.V.; Raines, C.A.; Lloyd, J.C. Pho3: A phosphorus-deficient mutant of Arabidopsis thaliana (L.) Heynh. Planta 2001, 212, 529–534. [Google Scholar] [CrossRef]
- Park, C.H.; Xu, H.; Yeo, H.J.; Park, Y.E.; Hwang, G.-S.; Park, N.I.; Park, S.U. Enhancement of the flavone contents of Scutellaria baicalensis hairy roots via metabolic engineering using maize Lc and Arabidopsis PAP1 transcription factors. Metab. Eng. 2021, 64, 64–73. [Google Scholar] [CrossRef]
- Hara, M.; Oki, K.; Hoshino, K.; Kuboi, T. Enhancement of anthocyanin biosynthesis by sugar in radish (Raphanus sativus) hypocotyl. Plant Sci. 2003, 164, 259–265. [Google Scholar] [CrossRef]
- Tsukaya, H.; Ohshima, T.; Naito, S.; Chino, M.; Komeda, Y. Sugar-dependent expression of the CHS-A gene for chalcone synthase from petunia in transgenic Arabidopsis. Plant Physiol. 1991, 97, 1414–1421. [Google Scholar] [CrossRef] [Green Version]
- Morkunas, I.; Narożna, D.; Nowak, W.; Samardakiewicz, S.; Remlein-Starosta, D. Cross-talk interactions of sucrose and Fusarium oxysporum in the phenylpropanoid pathway and the accumulation and localization of flavonoids in embryo axes of yellow lupine. J. Plant Physiol. 2011, 168, 424–433. [Google Scholar] [CrossRef]
- Park, C.H.; Yeo, H.J.; Park, S.-Y.; Kim, J.K.; Park, S.U. Comparative phytochemical analyses and metabolic profiling of different phenotypes of Chinese cabbage (Brassica rapa ssp. pekinensis). Foods 2019, 8, 587. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Photograph of (A) green-colored radishes; and (B) white-colored radishes.

Figure 2.
Scores and loading plots of PCA model for metabolites present in green and white radish roots.
Figure 2.
Scores and loading plots of PCA model for metabolites present in green and white radish roots.

Figure 3.
Correlation matrix of metabolites present in green and white radish roots.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Glucosinolate content in white and green radish cultivars (μg g−1).
| Compounds | White Radish Root | Green Radish Root |
|---|---|---|
| Glucoraphanin | 0.67 ± 0.03 | 0.71 ± 0.03 |
| Glucoalyssin | 0.08 ± 0.01 | 0.13 ± 0.02 *1 |
| Gluconapin | 0.08 ± 0.01 ** | 0.04 ± 0.01 |
| 4-Hydroxyglucobrassicin | 0.27 ± 0.03 * | 0.12 ± 0.00 |
| Glucoerucin | 0.28 ± 0.01 *** | 0.15 ± 0.03 |
| Glucoraphasatin | 11.34 ± 0.57 * | 9.93 ± 0.33 |
| 4-Methoxyglucobrassicin | 0.35 ± 0.00 | 0.49 ± 0.02 ** |
| Total | 13.06 ± 0.55 * | 11.56 ± 0.37 |
1 Student’s t-test, * p < 0.05, ** p < 0.01, *** p < 0.005.
Table 2.
Phenolic content in white and green radish cultivars (μg g−1).
| Compounds | White Radish Root | Green Radish Root |
|---|---|---|
| Gallic acid | 8.17 ± 0.01 | 11.64 ± 0.20 ***1 |
| Catechin | 84.68 ± 0.64 | 90.96 ± 0.69 *** |
| 4-hydroxybenzoic acid | 1.58 ± 0.34 | 3.22 ± 0.37 *** |
| Kaempferol | 40.33 ± 0.16 | 46.13 ± 2.11 * |
| Total | 134.75 ± 0.80 | 151.96 ± 2.48 * |
1 Student’s t-test, * p < 0.05, *** p < 0.005.
Table 3.
Chlorophyll content in white and green radish cultivars (μg g−1).
| Compounds | White Radish Root | Green Radish Root |
|---|---|---|
| Chlorophyll a | 5.64 ± 0.30 | 57.85 ± 6.45 ***1 |
| Chlorophyll b | 7.02 ± 0.14 | 14.26 ± 0.42 *** |
| Total | 12.66 ± 0.17 | 72.11 ± 0.37 *** |
1 Student’s t-test, *** p < 0.005.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Park, C.H.; Ki, W.; Kim, N.S.; Park, S.-Y.; Kim, J.K.; Park, S.U. Metabolic Profiling of White and Green Radish Cultivars (Raphanus sativus). Horticulturae 2022, 8, 310. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8040310
AMA Style
Park CH, Ki W, Kim NS, Park S-Y, Kim JK, Park SU. Metabolic Profiling of White and Green Radish Cultivars (Raphanus sativus). Horticulturae. 2022; 8(4):310. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8040310
Chicago/Turabian StylePark, Chang Ha, Wooyeal Ki, Nam Su Kim, Soo-Yun Park, Jae Kwang Kim, and Sang Un Park. 2022. "Metabolic Profiling of White and Green Radish Cultivars (Raphanus sativus)" Horticulturae 8, no. 4: 310. https://0-doi-org.brum.beds.ac.uk/10.3390/horticulturae8040310
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.