Chimeric Virus Made from crTMV RNA and the Coat Protein of Potato Leafroll Virus is Targeted to the Nucleolus and Can Infect Nicotiana benthamiana Mechanically

 ,
, {kind=link}
{kind=link}
{kind=link}
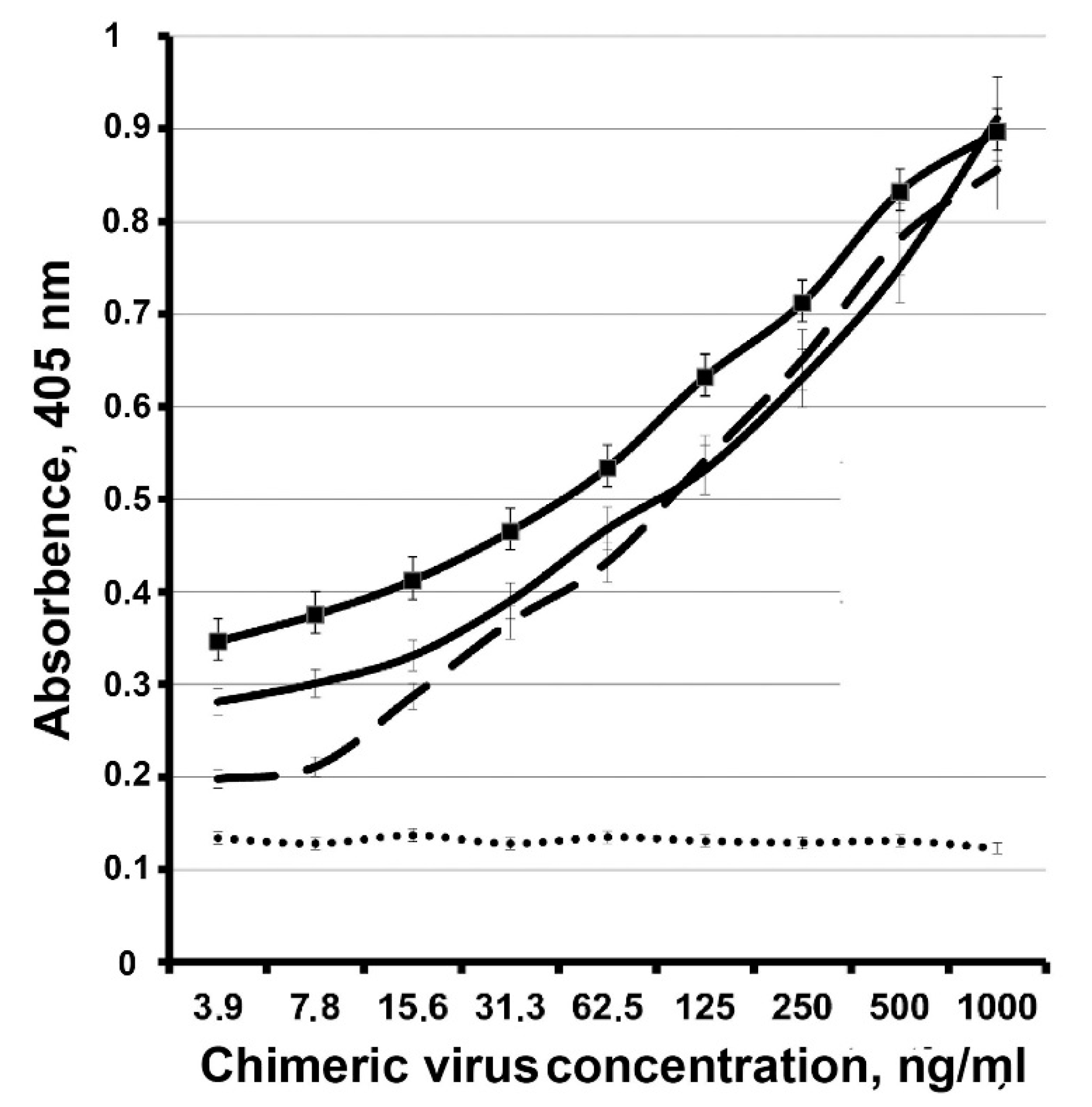{kind=link}
{kind=link}
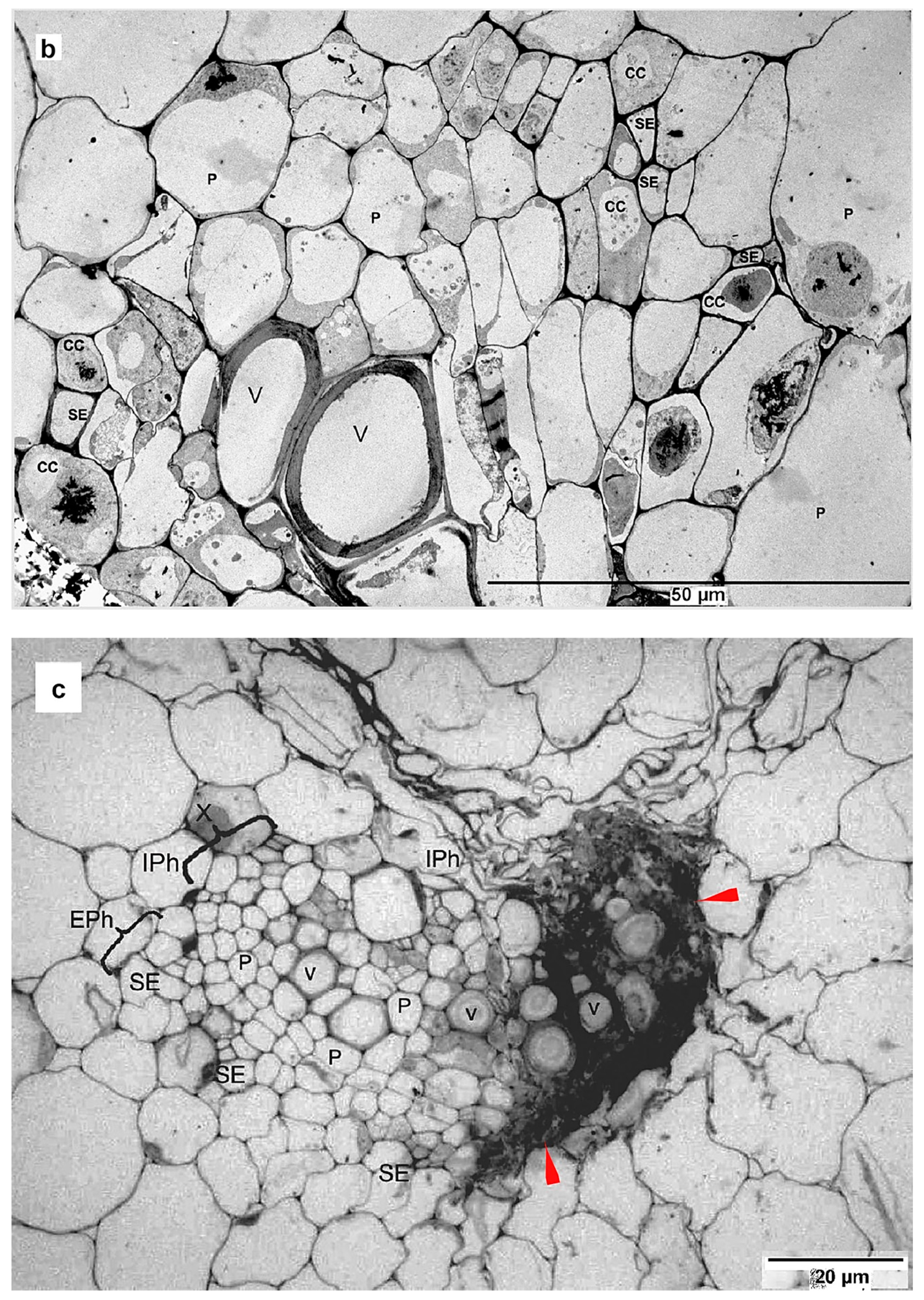{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
- 1)
- 5‘-ttctcgagatgagtacggtcgtggttaaaggaaacgtcaacgg-3‘ PLRV-CP (+)
- 2)
- 5‘-ttggtaccctatttggggttttgcaaagcca-3‘PLRV-CP (-)
- 3)
- 5‘-ggtggtgcacaccaaccacaatcccttcgcaggcgc-3‘ PLRV (+)
- 4)
- 5‘-gcgcctgcgaagggattgtggttggtgtgcaccacc-3‘ PLRV (-),
3. Results and Discussion
3.1. Previous TMV/PLRV Synthetic Vectors
3.2. Chimeric Virus Infectiousness
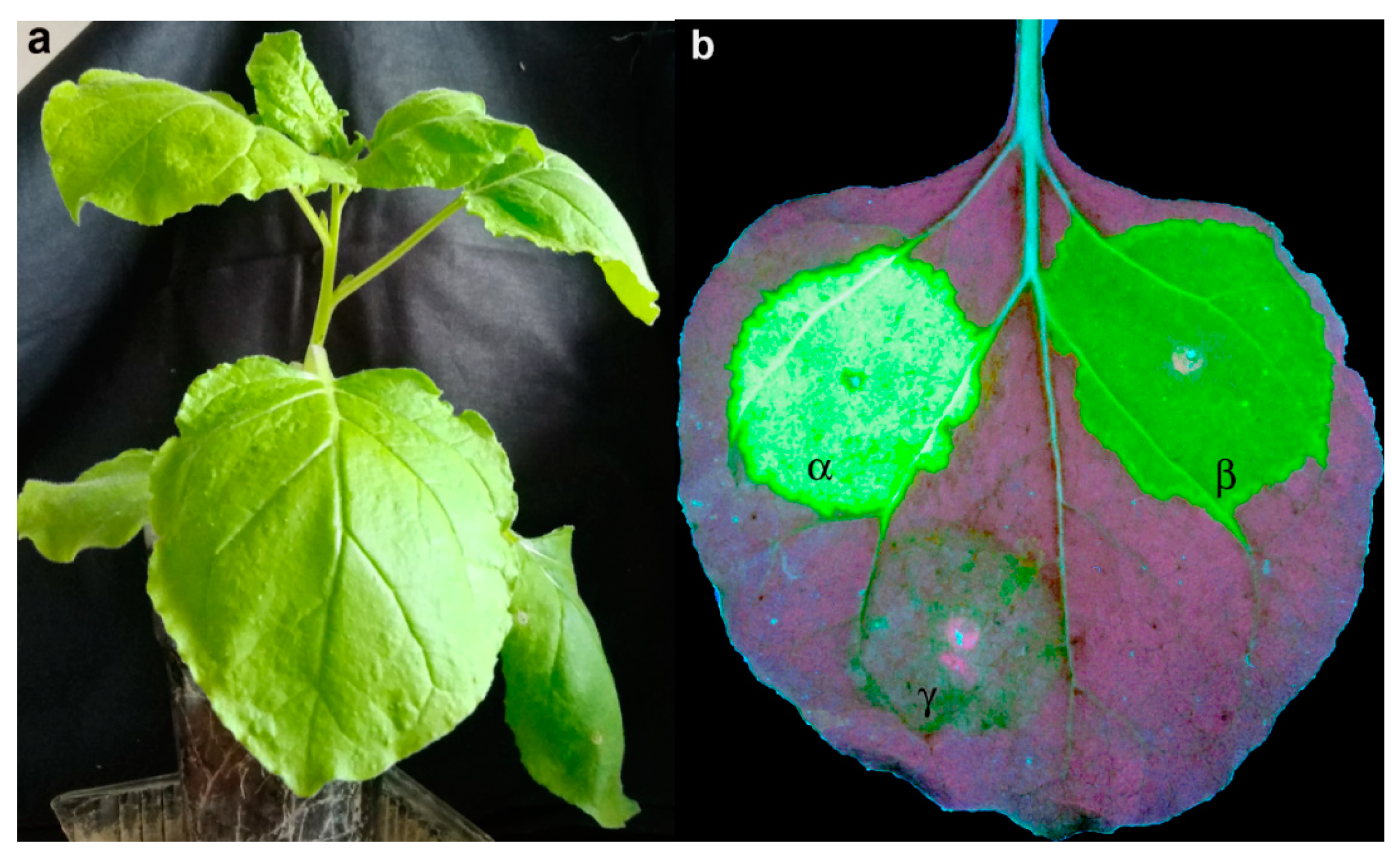
3.3. crTMV-CP-PLRV Exits the Phloem Into Nonvascular Tissues of N. benthamiana
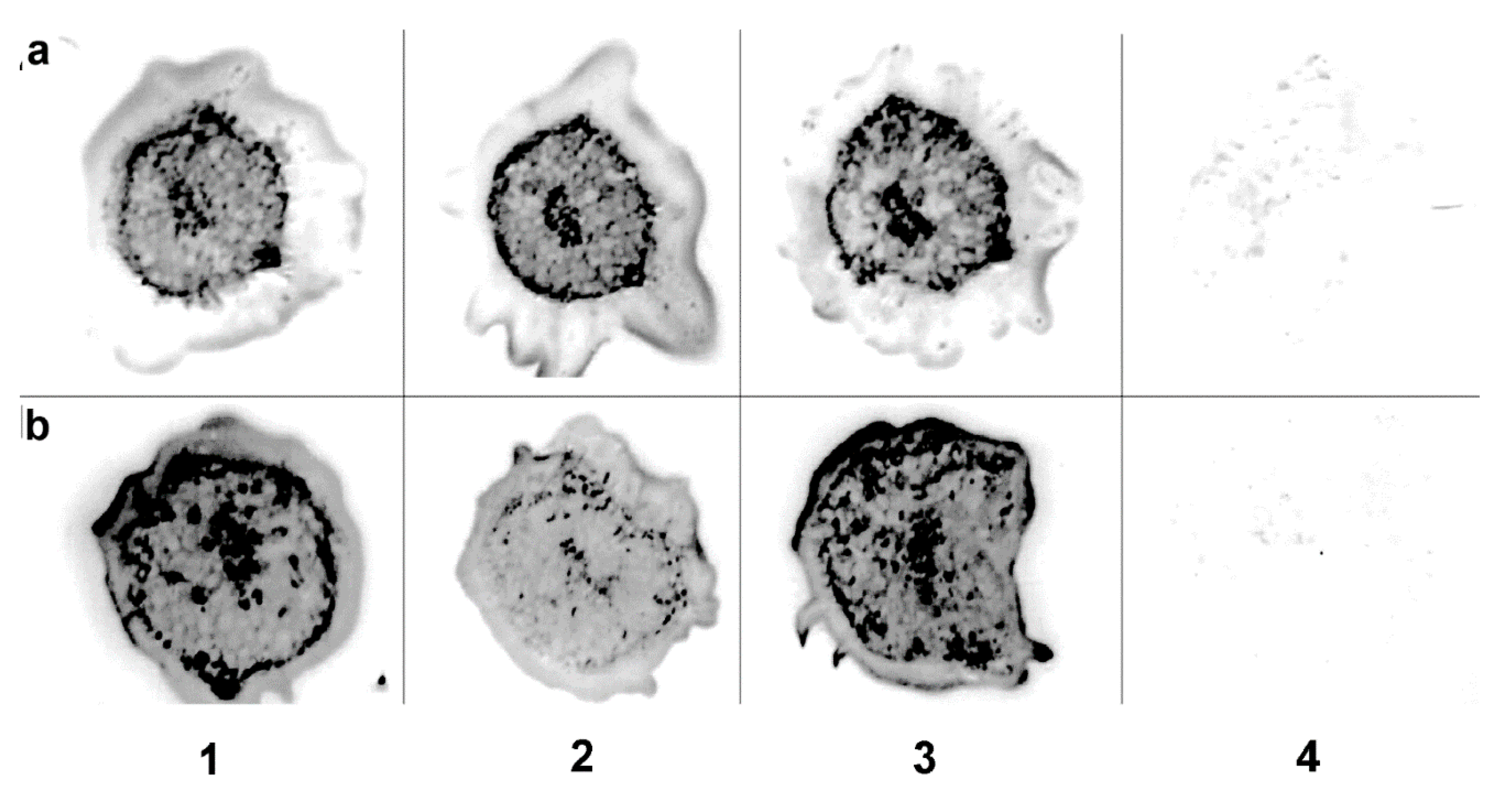
3.4. Fresh Virus Inoculum Can Infect Mechanically
3.5. Chimeric Virus Infection Induces Necrosis
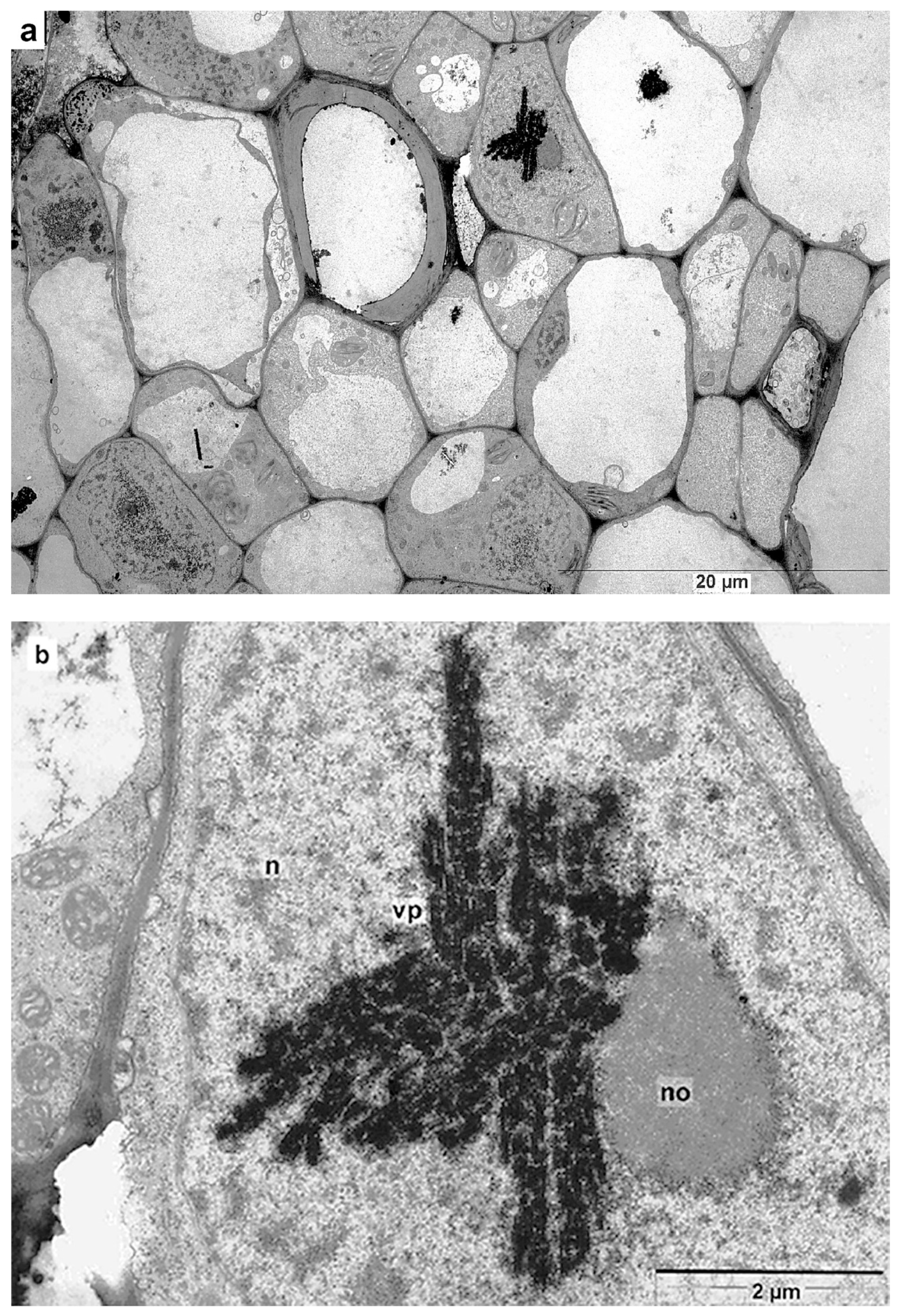
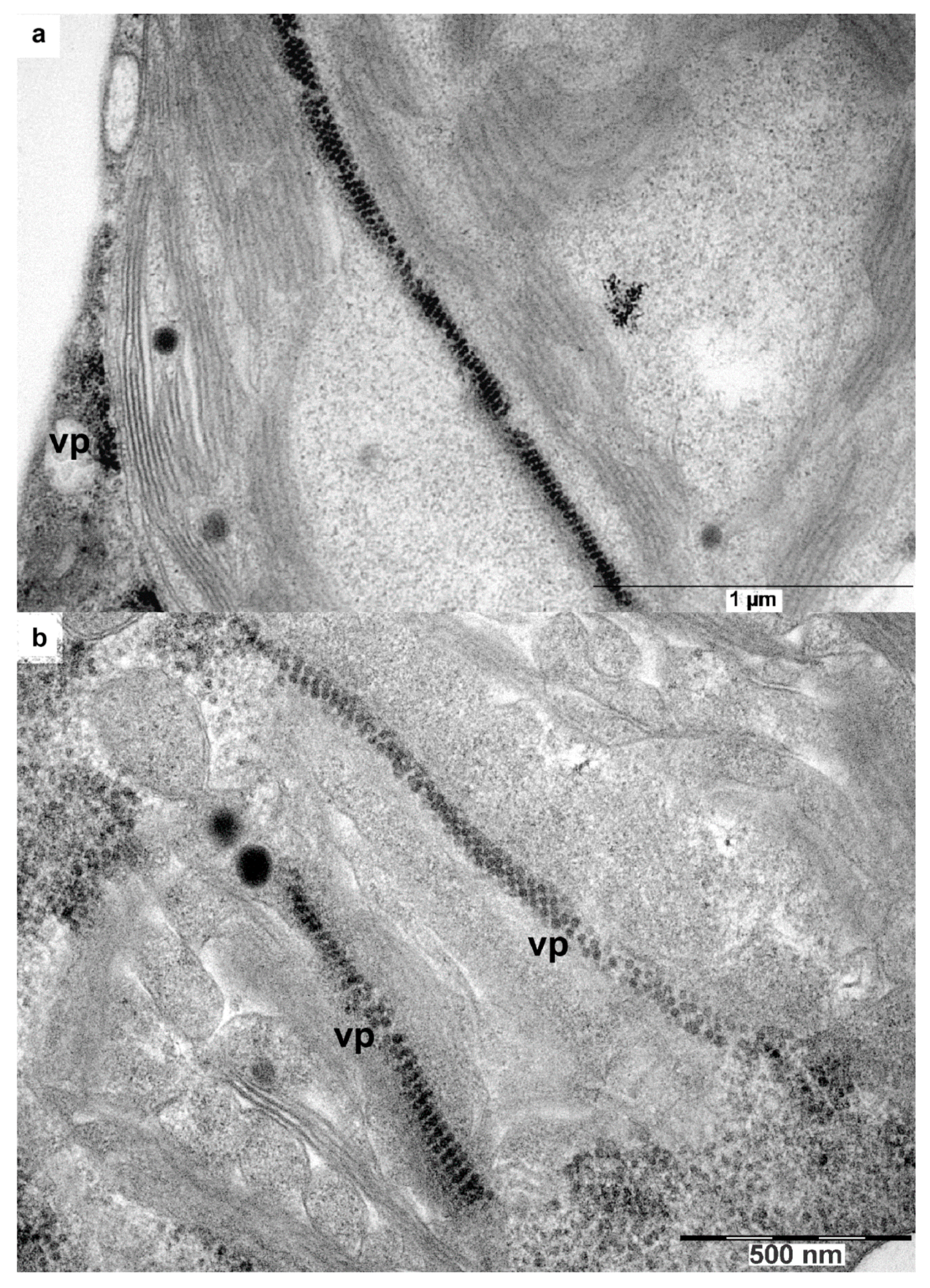
3.6. Intracellular Localization of crTMV-CP-PLRV
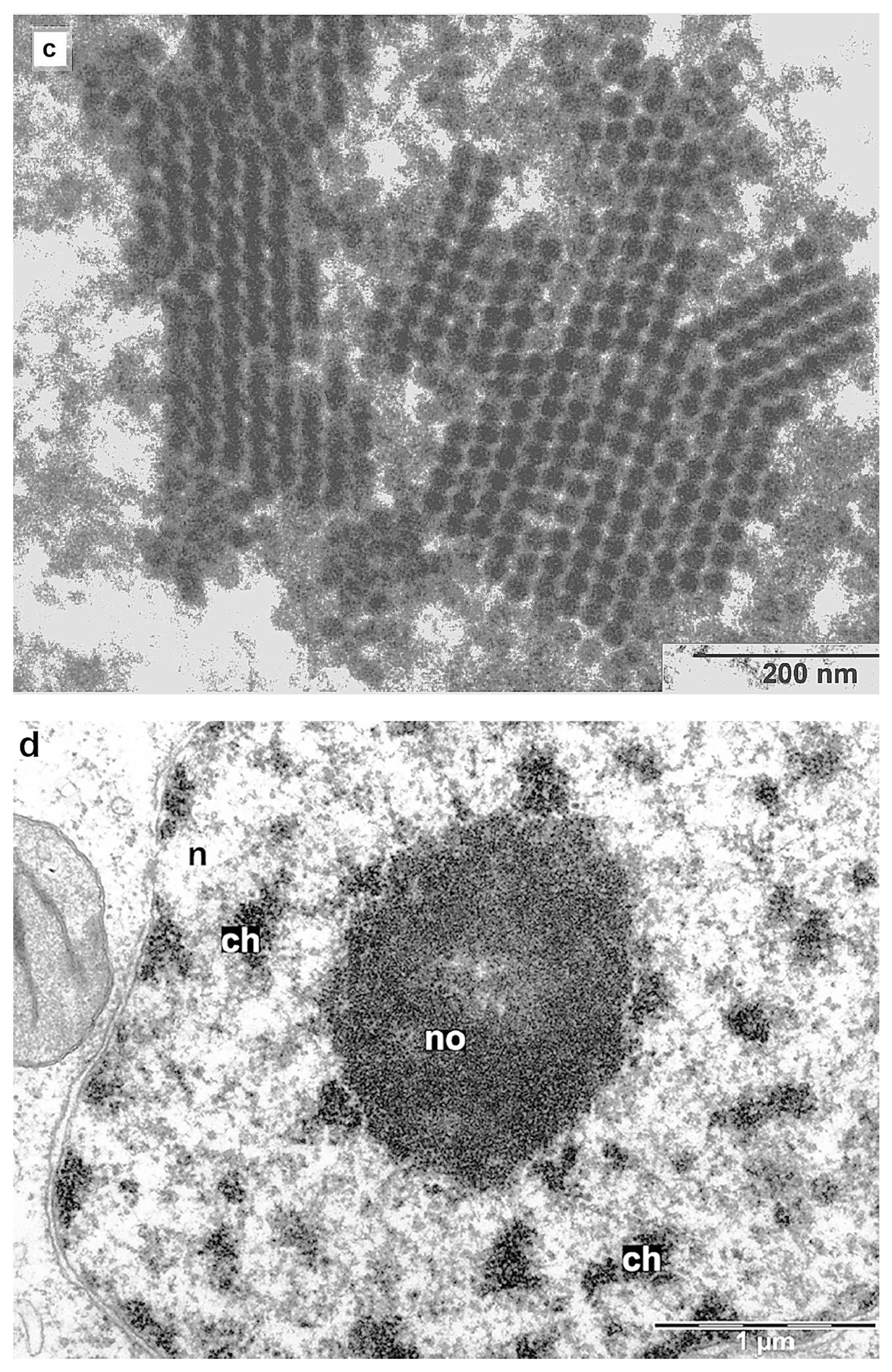
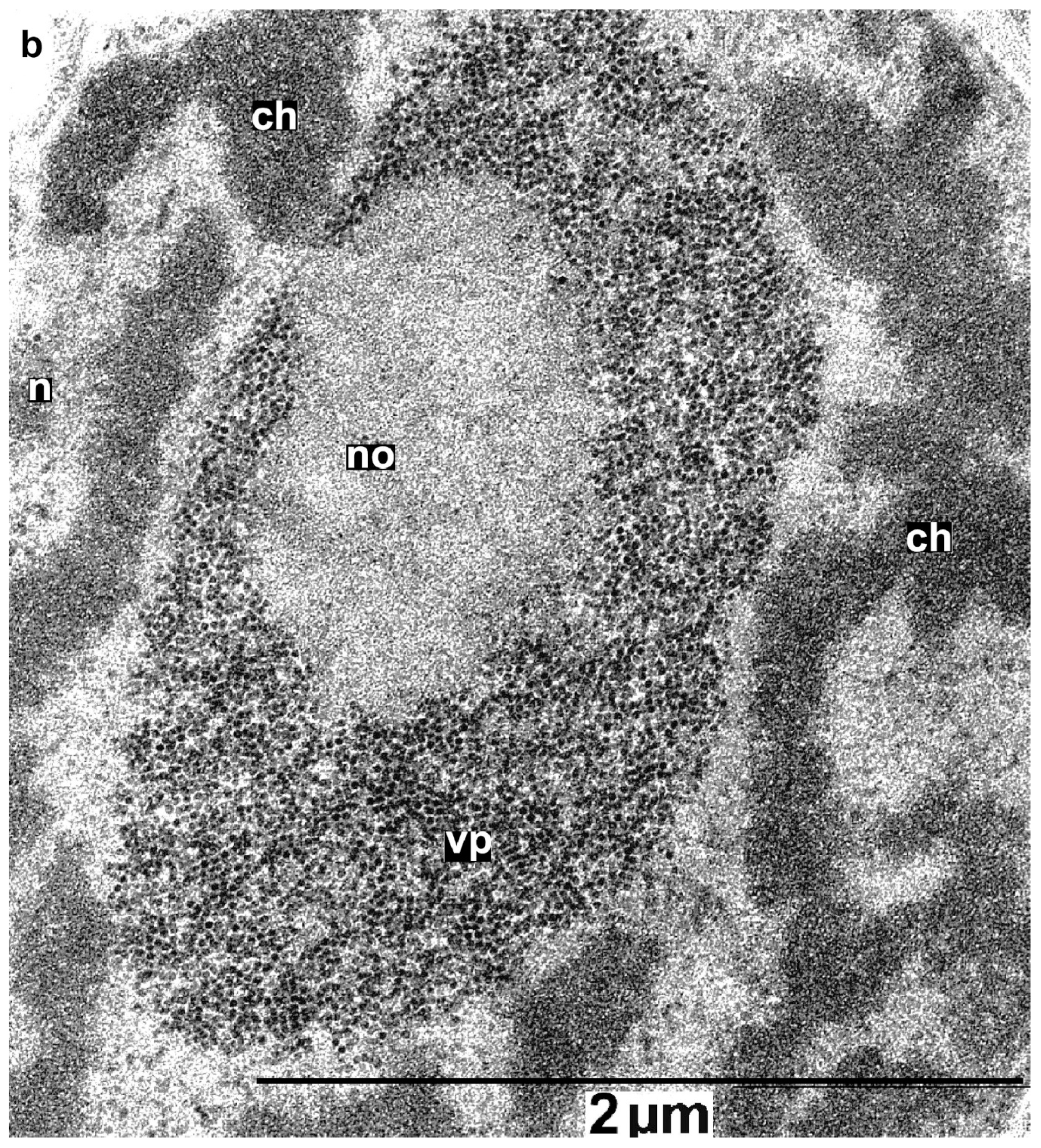
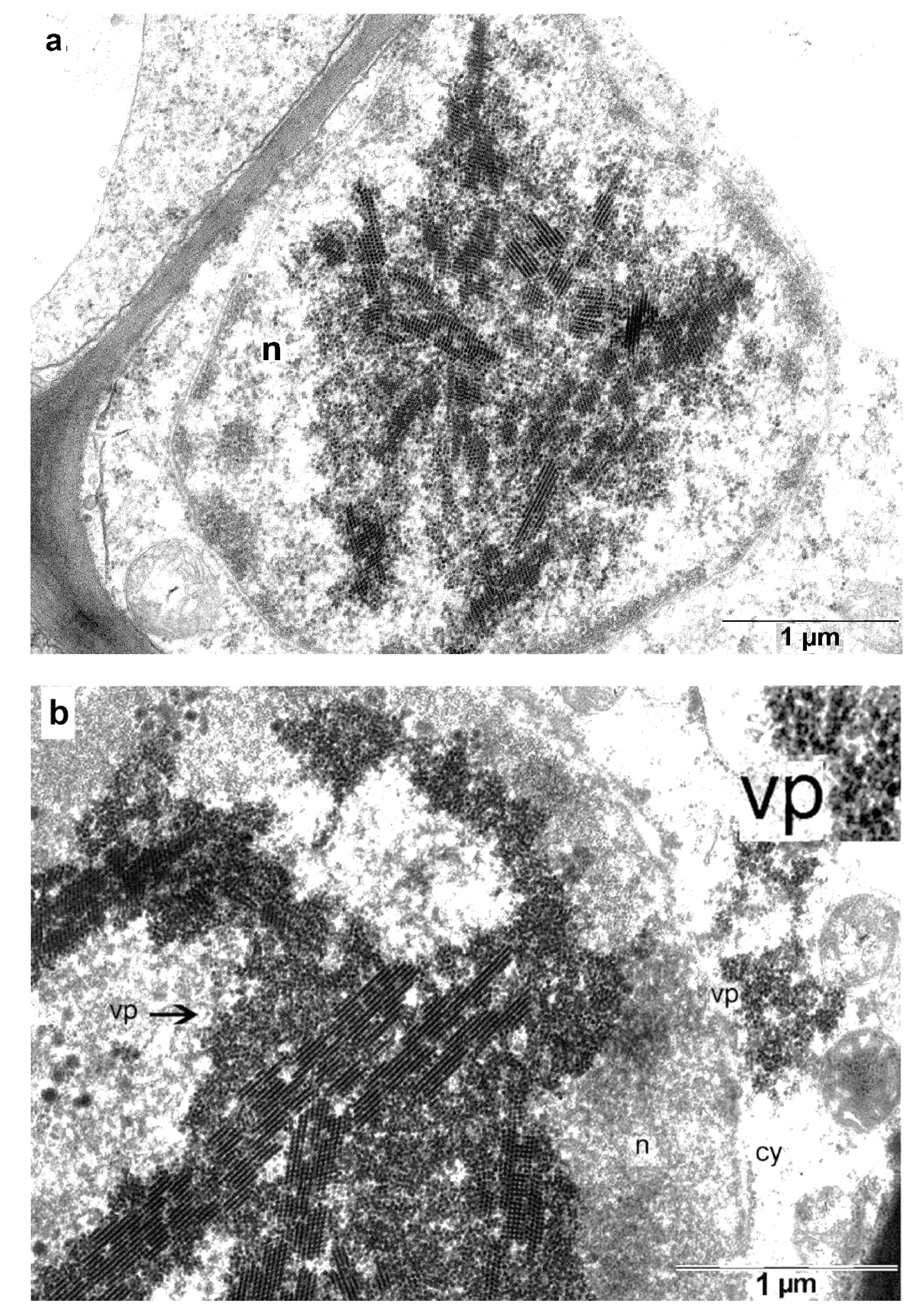
3.7. Major PLRV Coat Protein Targets the Chimeric Virus to Nucleolus
3.8. Transport of the Chimeric Virus Infection
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Wilk, F.; Huisman, M.J.; Cornelissen, B.J.; Huttinga, H.; Goldbach, R. Nucleotide sequence and organization of potato leafroll virus genomic RNA. FEBS Lett. 1989, 245, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Mayo, M.A.; Ziegler-Graff, V. Molecular biology of luteoviruses. Adv. Virus Res. 1996, 46, 413–460. [Google Scholar] [PubMed]
- Taliansky, M.; Mayo, M.A.; Barker, H. Potato leafroll virus: A classic pathogen shows some new tricks. Mol. Plant Path. 2003, 4, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.V.; Fraser, G.; Mayo, M.A.; Barker, H.; Taliansky, M. Umbravirus Gene Expression Helps Potato leafroll virus to Invade Mesophyll Tissues and to be Transmitted Mechanically between Plants. Virology 2001, 286, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Voinnet, O.O.; Pinto, Y.M.; Baulcombe, D.C. Suppression of gene silencing: A general strategy used by diverse DNA and RNA viruses of plants. Proc. Natl. Acad. Sci. USA 1999, 96, 14147–14152. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, P.M.; Wang, M.B.; Lough, T. Gene silencing as an adaptive defence against viruses. Nature 2001, 411, 834–842. [Google Scholar] [CrossRef]
- Barker, H.; McGeachy, K.D.; Ryabov, E.V.; Commandeur, U.; Mayo, M.A.; Taliansky, M. Evidence for RNA mediated defense effects on the accumulation of potato leafroll virus. J. Gen. Virol. 2001, 82, 3099–3106. [Google Scholar] [CrossRef] [Green Version]
- Atabekov, J.G.; Dorokhov, Y.L. Plant virus-specific transport function and resistance of plants to viruses. Adv. Virus Res. 1984, 29, 313–364. [Google Scholar] [CrossRef]
- Barker, H. Invasion of non-phloem tissue in Nicotiana clevelandii by potato leafroll virus is enhanced in plants also infected with potato Y potyvirus. J. Gen. Virol. 1987, 68, 1223–1227. [Google Scholar] [CrossRef]
- Barker, H. Specificity of the effect of sap-transmissible viruses in increasing the accumulation of luteoviruses in coinfected plants. Annu. Appl. Biol. 1989, 115, 71–78. [Google Scholar] [CrossRef]
- Rowhani, A.; Stace-Smith, R. Purification and characterization of potato leafroll virus. Virology 1979, 98, 45–54. [Google Scholar] [CrossRef]
- Takanami, Y.; Kubo, S. Enzyme-assisted purification of two phloem-limited plant viruses: Tobacco necrotic dwarf and potato leafroll. J. Gen. Virol. 1979, 44, 153–159. [Google Scholar] [CrossRef]
- Skurat, E.V.; Butenko, K.O.; Kondakova, O.A.; Nikitin, N.A.; Karpova, O.V.; Drygin, Y.F.; Atabekov, J.G. Chimeric Virus as a Source of the Potato Leafroll Virus Antigen. Mol. Biotechnol. 2017, 59, 469–481. [Google Scholar] [CrossRef] [PubMed]
- Skurat, E.V.; Drygin, Y.F.; Kondakova, O.A.; Butenko, K.O.; Atabekov, J.G. Method of Obtaining Preparative Quantities of Viral Particles of Phloem-Restricted Viruses. Patent RF RU2013123640 (A), 27 November 2014. [Google Scholar]
- Dorokhov, Y.L.; Ivanov, P.A.; Novikov, V.K.; Agranovsky, A.A.; Morozov, S.Y.; Efimov, V.A.; Casper, R.; Atabekov, J.G. Complete nucleotide sequence and genome organization of a tobamovirus infecting cruciferae plants. FEBS Lett. 1994, 350, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, G.R.; Helenius, A. Nuclear Import and Export of Viruses and Virus Genomes. Virology 1998, 246, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Solovyev, A.G.; Savenkov, E.I. Factors involved in the systemic transport of plant RNA viruses: The emerging role of the nucleus. J. Exp. Botany 2014, 65, 1689–1697. [Google Scholar] [CrossRef] [Green Version]
- Novoa, R.R.; Calderita, R.; Arranz, R.; Fontana, J.; Granzow, H.; Risco, C. Virus factories: Associations of cell organelles for viral replication and morphogenesis. Biol. Cell 2005, 97, 147–172. [Google Scholar] [CrossRef]
- Jin, X.; Cao, X.; Wang, X.; Jiang, J.; Wan, J.; Laliberté, J.-F.; Zhang, Y. Three-Dimensional Architecture and Biogenesis of Membrane Structures Associated with Plant Virus Replication. Front. Plant Sci. 2018, 9, 57. [Google Scholar] [CrossRef]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; CSHL Press: New York, NY, USA, 1989. [Google Scholar]
- Drygin, Y.F.; Blintsov, A.N.; Grigorenko, V.G.; Andreeva, I.P.; Osipov, A.P.; Varitzev, Y.A.; Uskov, A.I.; Kravchenko, D.V.; Atabekov, J.G. Highly sensitive field test lateral flow immunodiagnostics of PVX infection. Appl. Microbiol. Biotechnol. 2012, 93, 179–189. [Google Scholar] [CrossRef]
- Kuo, J. (Ed.) Electron Microscopy: Methods and Protocols, 2nd ed.; Humana Press: Totowa, NJ, USA, 2007. [Google Scholar]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Gill, C.C.; Chong, J. Development of the infection in oat leaves inoculated with barley yellow dwarf virus. Virology 1975, 66, 440–453. [Google Scholar] [CrossRef]
- Krenz, B.; Bronikowski, A.; Lu, X.; Ziebell, H.; Thompson, J.R.; Perry, K.L. Visual monitoring of cucumber mosaic virus infection in Nicotiana benthamiana following transmission by the aphid vector Myzus Persicae. J. Gen. Virol. 2015, 96, 2904–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peter, K.A.; Gildow, F.; Palukaitis, P.; Gray, S.M. The C Terminus of the Polerovirus P5 Readthrough Domain Limits Virus Infection to the Phloem. J. Virol. 2009, 83, 5419–5429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bald, J.G. Cytological Evidence for the Production of Plant Virus Ribonucleic Acid in the Nucleus. Virology 1964, 22, 377–387. [Google Scholar] [CrossRef]
- Milne, R.G. Electron microscopy of tobacco mosaic virus in leaves of Nicotiana Glutinosa. Virology 1966, 28, 527–532. [Google Scholar] [CrossRef]
- Gaur, R.K.; Khurana, S.M.P.; Dorokhov, Y.L. (Eds.) Plant Viruses. Diversity, Interaction and Management; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Wieczorek, P.; Wrzesinska, B.; Frackowiak, P.; Przybylska, A.; Obrępalska-Stęplowska, A. A Contribution of Tomato torrado virus Vp26 coat protein subunit to systemic necrosis induction and virus infectivity in Solanum lycopersicum. Virol. J. 2019, 16, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Shepardson, S.; Esau, K.; McCrum, R. Ultrastructure of potato leaf phloem infected with potato leafroll virus. Virology 1980, 105, 379–392. [Google Scholar] [CrossRef]
- Haupt, S.; Stroganova, T.; Ryabov, E.; Kim, S.H.; Fraser, G.; Dunkan, G.; Mayo, M.A.; Barker, H.; Taliansky, M. Nucleolar localization of potato leafroll virus capsid Proteins. J. Gen. Virol. 2005, 86, 2891–2896. [Google Scholar] [CrossRef] [PubMed]
- Shikata, E.; Maramorosch, K. Electron microscopy of pea enation mosaic virus in plant cell nuclei. Virology 1966, 30, 439–454. [Google Scholar] [CrossRef]
- Esau, K.; Hoefert, L.L. Development of Infection with Beet Western Yellows Virus in the Sugarbeet. Virology 1972, 48, 724–738. [Google Scholar] [CrossRef]
- Hiscox, J.A. RNA viruses: Hijacking the dynamic nucleolus. Nat. Rev. Microbiol. 2007, 5, 119–127. [Google Scholar] [CrossRef]
- Emmott, E.; Dove, B.K.; Howell, G. Viral nucleolar localisation signals determine dynamic trafficking within the nucleolus. Virology 2008, 380, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salvetti, A.; Greco, A. Viruses and the nucleolus: The fatal attraction. BBA Mol. Basis Dis. 2014, 1842, 840–847. [Google Scholar] [CrossRef]
- Taliansky, M.E.; Brown, J.W.; Rajamaki, M.L.; Valkonen, J.P.; Kalinina, N.O. Involvement of the plant nucleolus in virus and viroid infections: Parallels with animal pathosystems. Adv. Virus Res. 2010, 77, 119–158. [Google Scholar] [PubMed]
- Ashby, J.; Boutant, E.; Seemanpillai, M.; Sambade, A.; Ritzenthaler, C.; Heinlein, M. Tobacco Mosaic Virus Movement Protein Functions as a Structural Microtubule-Associated Protein. J. Virol. 2006, 80, 8329–8344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lartey, R.T.; Voss, T.C.; Melcher, U. Tobamovirus evolution: Gene overlaps, recombination, and taxonomic implications. Mol. Biol. Evol. 1996, 13, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Lartey, R.; Ghoshroy, S.; Ho, J.; Citovsky, V. Movement and subcellular localization of a tobamovirus in Arabidopsis. Plant. J. 1997, 12, 537–545. [Google Scholar] [CrossRef]
- Martin, R.M.; Ter-Avetisyan, G.; Herce, H.D.; Ludwig, A.K.; Lättig-Tünnemann, G.; Cardoso, M.C. Principles of protein targeting to the nucleolus. Nucleus 2015, 6, 314–325. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, I.B.; Lee, L.; Ripoll, D.R.; Palukaitis, P.; Gildow, F.; Gray, S.M. Point mutations in the potato leafroll virus major capsid protein alter virion stability and aphid transmission. J. Gen. Virol. 2007, 88, 1821–1830. [Google Scholar] [CrossRef]
- Hipper, C.; Brault, V.; Ziegler-Graff, V.; Revers, F. Viral and Cellular Factors Involved in Phloem Transport of Plant Viruses. Front. Plant Sci. 2013, 4, 154–247. [Google Scholar] [CrossRef] [Green Version]
- Sit, T.L.; Haikal, P.R.; Callaway, S.A.; Lommel, S.A. A Single Amino Acid Mutation in the Carnation Ringspot Virus Capsid Protein Allows Virion Formation but Prevents Systemic Infection. J. Virol. 2001, 75, 9538–9542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitsin, S.; Steplewski, K.; Fleysh, N.; Belanger, H.; Mikheeva, T.; Shivprasad, S.; Dawson, W.; Koprowski, H.; Yusibov, V. Expression of alfalfa mosaic virus coat protein in tobacco mosaic virus (TMV) deficient in the production of its native coat protein supports long-distance movement of a chimeric TMV. Proc. Natl. Acad. Sci. USA 1999, 96, 2549–2553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, P.H.; Bujarski, J.J. Multiple functions of capsid proteins in (+) stranded RNA viruses during plant-virus interactions. Virus Res. 2015, 196, 140–149. [Google Scholar] [CrossRef]
- Levy, A.; Zheng, J.Y.; Lazarowitz, S.G. The Tobamovirus Turnip Vein Clearing Virus 30-Kilodalton Movement Protein Localizes to Novel Nuclear Filaments to Enhance Virus Infection. J. Virol. 2013, 87, 6428–6440. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.J.; Steele, J.F.C.; Hesketh, E.L.; Walden, M.; Thompson, R.F.; Lomonossoff, G.P.; Ranson, N.A. Combining Transient Expression and Cryo-EM to Obtain High-Resolution Structures of Luteovirid Particles. Structure 2019, 27, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Plevka, P.; Tars, K.; Zeltins, A.; Balke, I.; Truve, E. The three-dimensional structure of ryegrass mottle virus at 2.9 Å resolution. Virology 2007, 369, 364–374. [Google Scholar] [CrossRef] [Green Version]













© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butenko, K.O.; Chaban, I.A.; Skurat, E.V.; Kondakova, O.A.; Drygin, Y.F. Chimeric Virus Made from crTMV RNA and the Coat Protein of Potato Leafroll Virus is Targeted to the Nucleolus and Can Infect Nicotiana benthamiana Mechanically. High-Throughput 2020, 9, 11. https://0-doi-org.brum.beds.ac.uk/10.3390/ht9020011
Butenko KO, Chaban IA, Skurat EV, Kondakova OA, Drygin YF. Chimeric Virus Made from crTMV RNA and the Coat Protein of Potato Leafroll Virus is Targeted to the Nucleolus and Can Infect Nicotiana benthamiana Mechanically. High-Throughput. 2020; 9(2):11. https://0-doi-org.brum.beds.ac.uk/10.3390/ht9020011
Chicago/Turabian StyleButenko, Konstantin O., Inna A. Chaban, Eugene V. Skurat, Olga A. Kondakova, and Yuri F. Drygin. 2020. "Chimeric Virus Made from crTMV RNA and the Coat Protein of Potato Leafroll Virus is Targeted to the Nucleolus and Can Infect Nicotiana benthamiana Mechanically" High-Throughput 9, no. 2: 11. https://0-doi-org.brum.beds.ac.uk/10.3390/ht9020011