
The Modulation of Cholesterol Metabolism Is Involved in the Antiviral Effect of Nitazoxanide

 , , , and
, , , and 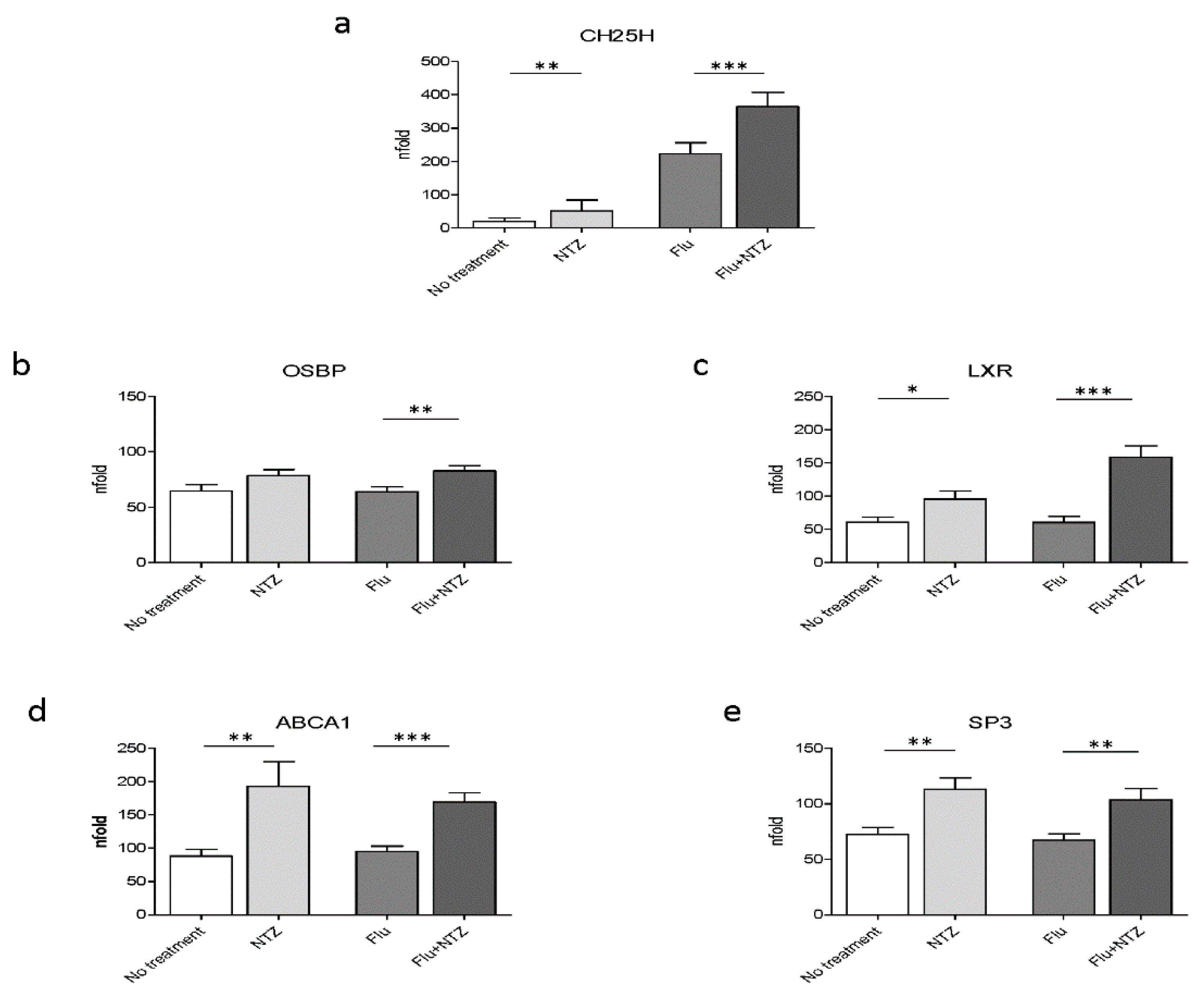{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. PBMC Isolation and Culture
2.3. RNA Extraction, Retro-Transcription (RT) and Real-Time PCR Analysis
2.4. Statistical Analysis
3. Results
3.1. Cholesterol Homeostasis Genes
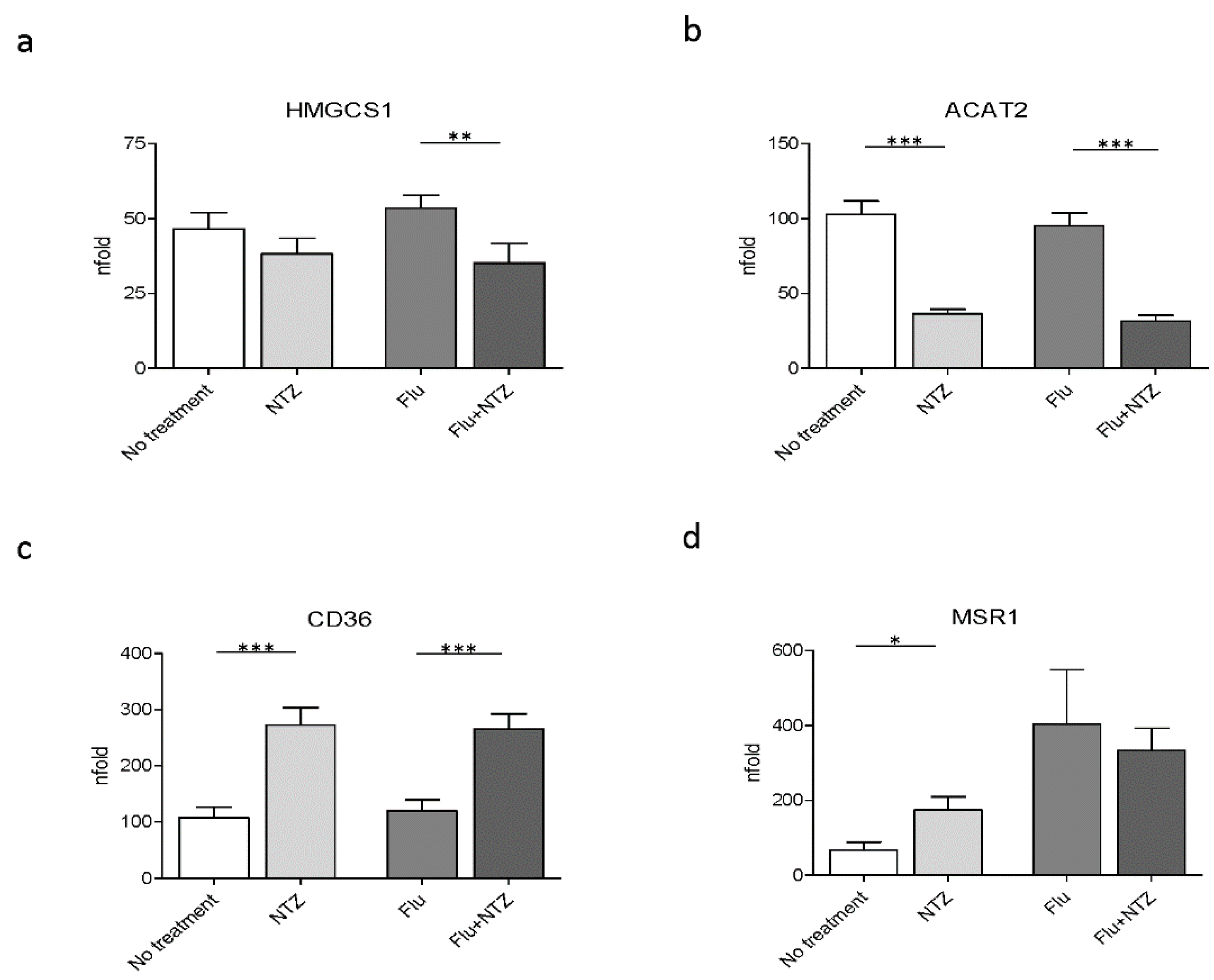
3.2. Cholesterol Biosynthesis Genes
3.3. Scavenger Receptor Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amorosa, V.K.; Luetkemeyer, A.; Kang, M.; Johnson, V.A.; Umbleja, T.; Haas, D.W.; Yesmin, S.; Bardin, M.C.; Chung, R.T.; Alston-Smith, B.; et al. Addition of Nitazoxanide to PEG-IFN and Ribavirin to Improve HCV Treatment Response in HIV-1 and HCV Genotype 1 Coinfected Persons Naïve to HCV Therapy: Results of the ACTG A5269 Trial. HIV Clin. Trials. 2013, 14, 274–283. [Google Scholar] [CrossRef]
- Haffizulla, J.; Hartman, A.; Hoppers, M.; Resnick, H.; Samudrala, S.; Ginocchio, C.; Bardin, M.; Rossignol, J.-F. Effect of nitazoxanide in adults and adolescents with acute uncomplicated influenza: A double-blind, randomised, placebo-controlled, phase 2b/3 trial. Lancet Infect. Dis. 2014, 14, 609–618. [Google Scholar] [CrossRef]
- Hui, D.S.; Lee, N.; Chan, P.K.; Beigel, J.H. The role of adjuvant immunomodulatory agents for treatment of severe influenza. Antivir. Res. 2018, 150, 202–216. [Google Scholar] [CrossRef]
- Kaufman, S.S.; Green, K.Y.; Korba, B.E. Treatment of norovirus infections: Moving antivirals from the bench to the bedside. Antivir. Res. 2014, 105, 80. [Google Scholar] [CrossRef] [Green Version]
- Korba, B.E.; Montero, A.B.; Farrar, K.; Gaye, K.; Mukerjee, S.; Ayers, M.S.; Rossignol, J.-F. Nitazoxanide, tizoxanide and other thiazolides are potent inhibitors of hepatitis B virus and hepatitis C virus replication. Antivir. Res. 2008, 77, 56–63. [Google Scholar] [CrossRef]
- Rossignol, J.-F.; Elfert, A.; El–Gohary, Y.; Keeffe, E.B. Improved Virologic Response in Chronic Hepatitis C Genotype 4 Treated With Nitazoxanide, Peginterferon, and Ribavirin. Gastroenterology 2009, 136, 856–862. [Google Scholar] [CrossRef]
- Rossignol, J.-F. Nitazoxanide, a new drug candidate for the treatment of Middle East respiratory syndrome coronavirus. J. Infect. Public Health 2016, 9, 227–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossignol, J.-F. Nitazoxanide: A first-in-class broad-spectrum antiviral agent. Antivir. Res. 2014, 110, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, J.F.; Kabil, S.M.; El-Gohary, Y.; Elfert, A.; Keeffe, E.B. Clinical trial: Randomized, double-blind, placebo-controlled study of nitazoxanide monotherapy for the treatment of patients with chronic hepatitis C genotype. Aliment. Pharmacol. Ther. 2008, 28, 574–580. [Google Scholar] [CrossRef]
- Rossignol, J.-F.; El-Gohary, Y.M. Nitazoxanide in the treatment of viral gastroenteritis: A randomized double-blind placebo-controlled clinical trial. Aliment. Pharmacol. Ther. 2006, 24, 1423–1430. [Google Scholar] [CrossRef] [PubMed]
- Trabattoni, D.; Gnudi, F.; Ibba, S.V.; Saulle, I.; Agostini, S.; Masetti, M.; Biasin, M.; Rossignol, J.-F.; Clerici, M. Thiazolides Elicit Anti-Viral Innate Immunity and Reduce HIV Replication. Sci. Rep. 2016, 6, 27148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbod-Giannone, M.-C.; Li, Y.; Holleboom, A.; Han, S.; Hsu, L.-C.; Tabas, I.; Tall, A.R. TNFα induces ABCA1 through NF-κB in macrophages and in phagocytes ingesting apoptotic cells. Proc. Natl. Acad. Sci. USA 2006, 103, 3112–3117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maitra, U.; Parks, J.S.; Li, L. An Innate Immunity Signaling Process Suppresses Macrophage ABCA1 Expression through IRAK-1-Mediated Downregulation of Retinoic Acid Receptor α and NFATc. Mol. Cell Biol. 2009, 29, 5989–5997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenizia, C.; Rossignol, J.-F.; Clerici, M.; Biasin, M. Genetic and immune determinants of immune activation in HIV-exposed seronegative individuals and their role in protection against HIV infection. Infect. Genet. Evol. 2018, 66, 325–334. Available online: https://0-linkinghub-elsevier-com.brum.beds.ac.uk/retrieve/pii/S1567134817304392 (accessed on 7 June 2021). [CrossRef]
- Liu, S.-Y.; Aliyari, R.; Chikere, K.; Li, G.; Marsden, M.D.; Smith, J.K.; Pernet, O.; Guo, H.; Nusbaum, R.; Zack, J.A.; et al. Interferon-Inducible Cholesterol-25-Hydroxylase Broadly Inhibits Viral Entry by Production of 25-Hydroxycholesterol. Immunity 2013, 38, 92–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanc, M.; Hsieh, W.Y.; Robertson, K.A.; Watterson, S.; Shui, G.; Lacaze, P.; Khondoker, M.; Dickinson, P.; Sing, G.; Rodríguez-Martín, S.; et al. Host Defense against Viral Infection Involves Interferon Mediated Down-Regulation of Sterol Biosynthesis. PLoS Biol. 2011, 9, e1000598. [Google Scholar] [CrossRef] [Green Version]
- Civra, A.; Cagno, V.; Donalisio, M.; Biasi, F.; Leonarduzzi, G.; Poli, G.; Lembo, D. Inhibition of pathogenic non-enveloped viruses by 25-hydroxycholesterol and 27-hydroxycholesterol. Sci. Rep. 2014, 4, 7487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, B.; Gonçalves, S.; Disalvo, A.; Hollmann, A.; Santos, N.C. Effect of 25-hydroxycholesterol in viral membrane fusion: Insights on HIV inhibition. Biochim. Biophys. Acta BBA Biomembr. 2018, 1860, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Deng, Y.-Q.; Wang, S.; Ma, F.; Aliyari, R.; Huang, X.-Y.; Zhang, N.-N.; Watanabe, M.; Dong, H.-L.; Liu, P.; et al. 25-Hydroxycholesterol Protects Host against Zika Virus Infection and Its Associated Microcephaly in a Mouse Model. Immunity 2017, 46, 446–456. [Google Scholar] [CrossRef] [Green Version]
- Cyster, J.G.; Dang, E.V.; Reboldi, A.; Yi, T. 25-Hydroxycholesterols in innate and adaptive immunity. Nat. Rev. Immunol. 2014, 14, 731–743. [Google Scholar] [CrossRef]
- Lembo, D.; Cagno, V.; Civra, A.; Poli, G. Oxysterols: An emerging class of broad spectrum antiviral effectors. Mol. Asp. Med. 2016, 49, 23–30. [Google Scholar] [CrossRef]
- Fritsch, S.D.; Weichhart, T. Effects of Interferons and Viruses on Metabolism. Front. Immunol. 2016, 7, 630. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC5174094/ (accessed on 12 December 2018). [CrossRef] [Green Version]
- Martín-Acebes, M.A.; Vázquez-Calvo, Á.; Caridi, F.; Saiz, J.-C.; Sobrino, F. Lipid Involvement in Viral Infections: Present and Future Perspectives for the Design of Antiviral Strategies. In Lipid Metabolism; InTech: London, UK, 2013. [Google Scholar]
- Lipid Involvement in Viral Infections: Present and Future Perspectives for the Design of Antiviral Strategies. Available online: https://www.intechopen.com/books/lipid-metabolism/lipid-involvement-in-viral-infections-present-and-future-perspectives-for-the-design-of-antiviral-st (accessed on 3 December 2018).
- Mazzon, M.; Mercer, J. Lipid interactions during virus entry and infection. Cell Microbiol. 2014, 16, 1493–1502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chazal, N.; Gerlier, D. Virus Entry, Assembly, Budding, and Membrane Rafts. Microbiol. Mol. Biol. Rev. 2003, 67, 226–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.; Krijnse-Locker, J. Modification of intracellular membrane structures for virus replication. Nat. Rev. Microbiol. 2008, 6, 363–374. [Google Scholar] [CrossRef]
- Sabahi, A. Hepatitis C Virus entry: The early steps in the viral replication cycle. Virol. J. 2009, 6, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Whittaker, G.R. Role for Influenza Virus Envelope Cholesterol in Virus Entry and Infection. J. Virol. 2003, 77, 12543–12551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaitseva, E.; Yang, S.-T.; Melikov, K.; Pourmal, S.; Chernomordik, L.V. Dengue Virus Ensures Its Fusion in Late Endosomes Using Compartment-Specific Lipids. PLoS Pathog. 2010, 6, e1001131. [Google Scholar] [CrossRef] [Green Version]
- Heaton, N.S.; Randall, G. Dengue virus induced autophagy regulates lipid metabolism. Cell Host Microbe. 2010, 8, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Reiss, S.; Rebhan, I.; Backes, P.; Romero-Brey, I.; Erfle, H.; Matula, P.; Kaderali, L.; Poenisch, M.; Blankenburg, H.; Hiet, M.-S.; et al. Recruitment and activation of a lipid kinase by hepatitis C virus NS5A is essential for integrity of the membranous replication compartment. Cell Host Microbe. 2011, 9, 32–45. [Google Scholar] [CrossRef] [Green Version]
- Boasso, A.; Royle, C.M.; Doumazos, S.; Aquino, V.N.; Biasin, M.; Piacentini, L.; Tavano, B.; Fuchs, D.; Mazzotta, F.; Lo Caputo, S.; et al. Overactivation of plasmacytoid dendritic cells inhibits antiviral T-cell responses: A model for HIV immunopathogenesis. Blood 2011, 118, 5152–5162. [Google Scholar] [CrossRef] [Green Version]
- Egger, D.; Wölk, B.; Gosert, R.; Bianchi, L.; Blum, H.E.; Moradpour, D.; Bienz, K. Expression of Hepatitis C Virus Proteins Induces Distinct Membrane Alterations Including a Candidate Viral Replication Complex. J. Virol. 2002, 76, 5974–5984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fenizia, C.; Fiocchi, M.; Jones, K.; Parks, R.W.; Ceribelli, M.; Chevalier, S.A.; Edwards, D.; Ruscetti, F.; Pise-Masison, C.A.; Franchini, G. Human T-Cell Leukemia/Lymphoma Virus Type 1 p30, but Not p12/p8, Counteracts Toll-Like Receptor 3 (TLR3) and TLR4 Signaling in Human Monocytes and Dendritic Cells. J. Virol. 2014, 88, 393–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaccari, M.; Fenizia, C.; Ma, Z.-M.; Hryniewicz, A.; Boasso, A.; Doster, M.N.; Miller, C.J.; Lindegardh, N.; Joel Tarning, J.; Landay, A.L.; et al. Transient Increase of Interferon-Stimulated Genes and No Clinical Benefit by Chloroquine Treatment During Acute Simian Immunodeficiency Virus Infection of Macaques. AIDS Res. Hum. Retrovir. 2014, 30, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyea, M.M.; Heslop, C.L.; Sawyez, C.G.; Edwards, J.Y.; Markle, J.G.; Hegele, R.A.; Huff, M.W. Selective Up-regulation of LXR-regulated Genes ABCA1, ABCG1, and APOE in Macrophages through Increased Endogenous Synthesis of 24(S),25-Epoxycholesterol. J. Biol. Chem. 2007, 282, 5207–5216. [Google Scholar] [CrossRef] [Green Version]
- Sapetschnig, A.; Koch, F.; Rischitor, G.; Mennenga, T.; Suske, G. Complexity of Translationally Controlled Transcription Factor Sp3 Isoform Expression. J. Biol. Chem. 2004, 279, 42095–42105. [Google Scholar] [CrossRef] [Green Version]
- Silverstein, R.L.; Febbraio, M. CD36, a Scavenger Receptor Involved in Immunity, Metabolism, Angiogenesis, and Behavior. Sci. Signal. 2009, 2, re3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tall, A.R.; Yvan-Charvet, L. Cholesterol, inflammation and innate immunity. Nat. Rev. Immunol. 2015, 15, 104. [Google Scholar] [CrossRef] [Green Version]
- Amako, Y.; Sarkeshik, A.; Hotta, H.; Yates, J.; Siddiqui, A. Role of Oxysterol Binding Protein in Hepatitis C Virus infection. J. Virol. 2009, 83, 9237–9246. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, G.; Langmann, T. Transcriptional regulatory networks in lipid metabolism control ABCA1 expression. Biochim. Biophys. Acta BBA Mol. Cell. Biol. Lipids. 2005, 1735, 1–19. [Google Scholar] [CrossRef]
- Biasin, M.; Trabattoni, D.; Rossignol, J.-F.; Clerici, M. Immune correlates of protection against HIV infection and how to elicit them. Mucosal Immunol. 2017, 10, 827–828. [Google Scholar] [CrossRef] [PubMed]
- Gekonge, B.; Bardin, M.C.; Montaner, L.J. Short Communication: Nitazoxanide Inhibits HIV Viral Replication in Monocyte-Derived Macrophages. AIDS Res. Hum. Retrovir. 2015, 31, 237–241. [Google Scholar] [CrossRef] [Green Version]
- Cao, R.-Y.; Xu, Y.; Zhang, T.-H.; Yang, J.-J.; Yuan, Y.; Hao, P.; Shi, Y.; Zhong, J.; Zhong, W. Pediatric Drug Nitazoxanide: A Potential Choice for Control of Zika. Open Forum. Infect. Dis. 2017, 4, ofx009. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5414027/ (accessed on 7 June 2021).
- An, L.; Liu, R.; Tang, W.; Wu, J.-G.; Chen, X. Screening and identification of inhibitors against influenza A virus from a US drug collection of 1280 drugs. Antivir. Res. 2014, 109, 54–63. [Google Scholar] [CrossRef] [PubMed]
- Belardo, G.; Cenciarelli, O.; Frazia, S.L.; Rossignol, J.F.; Santoro, M.G. Synergistic Effect of Nitazoxanide with Neuraminidase Inhibitors against Influenza A Viruses In Vitro. Antimicrob. Agents Chemother. 2015, 59, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Rossignol, J.F.; La Frazia, S.; Chiappa, L.; Ciucci, A.; Santoro, M.G. Thiazolides, a New Class of Anti-influenza Molecules Targeting Viral Hemagglutinin at the Post-translational Level. J. Biol. Chem. 2009, 284, 29798–29808. [Google Scholar] [CrossRef] [Green Version]
- Sleeman, K.; Mishin, V.P.; Guo, Z.; Garten, R.J.; Balish, A.; Fry, A.M.; Villanueva, J.; Stevens, J.; Gubareva, L.V. Antiviral Susceptibility of Variant Influenza A(H3N2)v Viruses Isolated in the United States from 2011 to 2013. Antimicrob. Agents Chemother. 2014, 58, 2045–2051. [Google Scholar] [CrossRef] [Green Version]
- Koszalka, P.; Tilmanis, D.; Hurt, A.C. Influenza antivirals currently in late-phase clinical trial. Influenza Other Respir Viruses. 2017, 11, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Charlton-Menys, V.; Durrington, P.N. Human cholesterol metabolism and therapeutic molecules: Cholesterol metabolism and therapeutic molecules. Exp. Physiol. 2008, 93, 27–42. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fenizia, C.; Ibba, S.V.; Vanetti, C.; Strizzi, S.; Rossignol, J.-F.; Biasin, M.; Trabattoni, D.; Clerici, M. The Modulation of Cholesterol Metabolism Is Involved in the Antiviral Effect of Nitazoxanide. Infect. Dis. Rep. 2021, 13, 636-644. https://0-doi-org.brum.beds.ac.uk/10.3390/idr13030060
Fenizia C, Ibba SV, Vanetti C, Strizzi S, Rossignol J-F, Biasin M, Trabattoni D, Clerici M. The Modulation of Cholesterol Metabolism Is Involved in the Antiviral Effect of Nitazoxanide. Infectious Disease Reports. 2021; 13(3):636-644. https://0-doi-org.brum.beds.ac.uk/10.3390/idr13030060
Chicago/Turabian StyleFenizia, Claudio, Salomè Valentina Ibba, Claudia Vanetti, Sergio Strizzi, Jean-François Rossignol, Mara Biasin, Daria Trabattoni, and Mario Clerici. 2021. "The Modulation of Cholesterol Metabolism Is Involved in the Antiviral Effect of Nitazoxanide" Infectious Disease Reports 13, no. 3: 636-644. https://0-doi-org.brum.beds.ac.uk/10.3390/idr13030060