Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas geniculata-Modulated Biochemical Responses Provide Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil

 , ,
, , 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characterization and Identification of Microbial Strains
2.2. Root Colonization Assay
2.3. Green Fluorescent Protein (GFP)-Tagging of the Pseudomonas Geniculata
2.4. In Planta Assay
2.4.1. Soil Preparation and Analysis
2.4.2. Experimental Set up
2.5. Sampling and Analysis
2.6. Statistical Analyses
3. Results
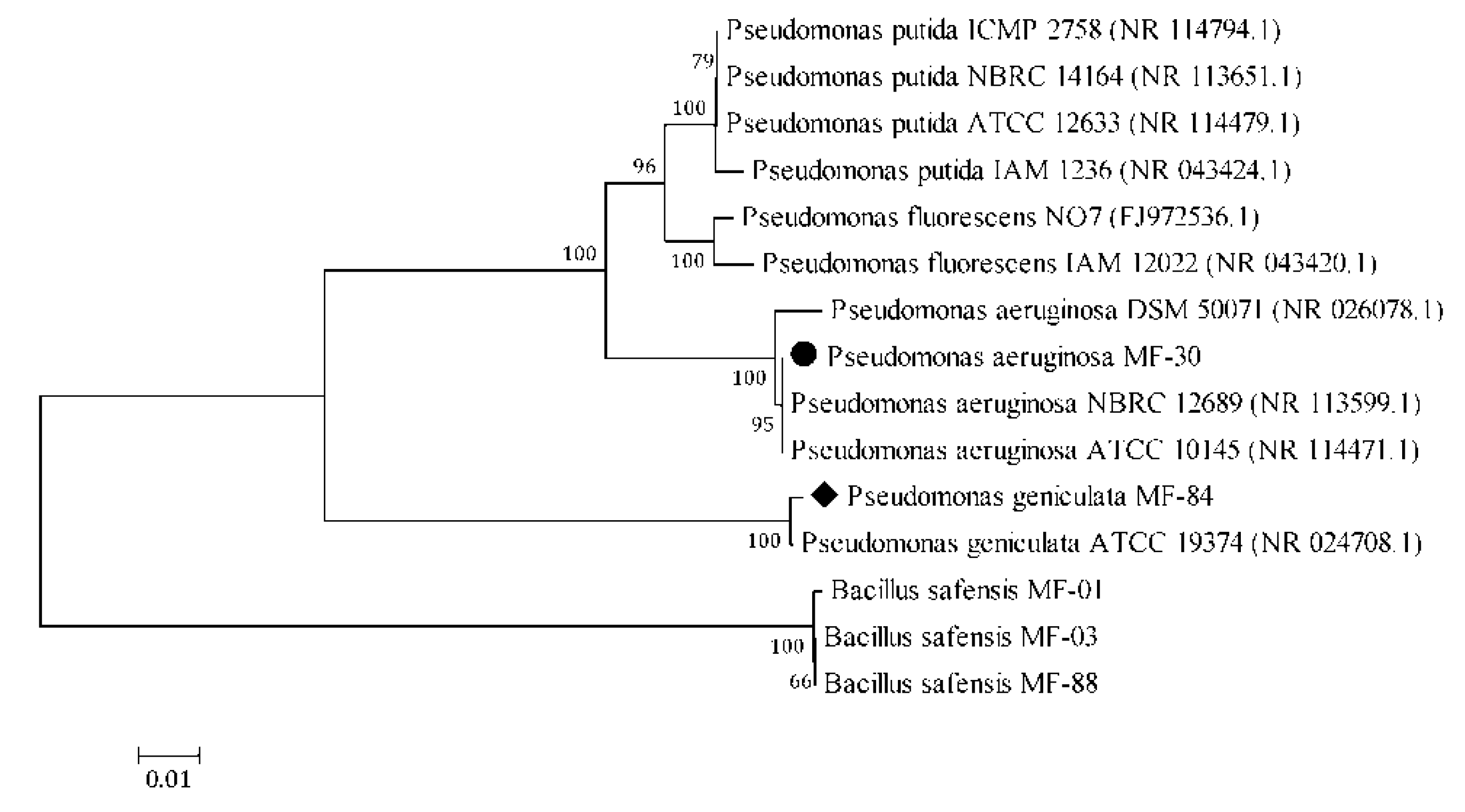
3.1. Characterization and Identification of Microbial Strains
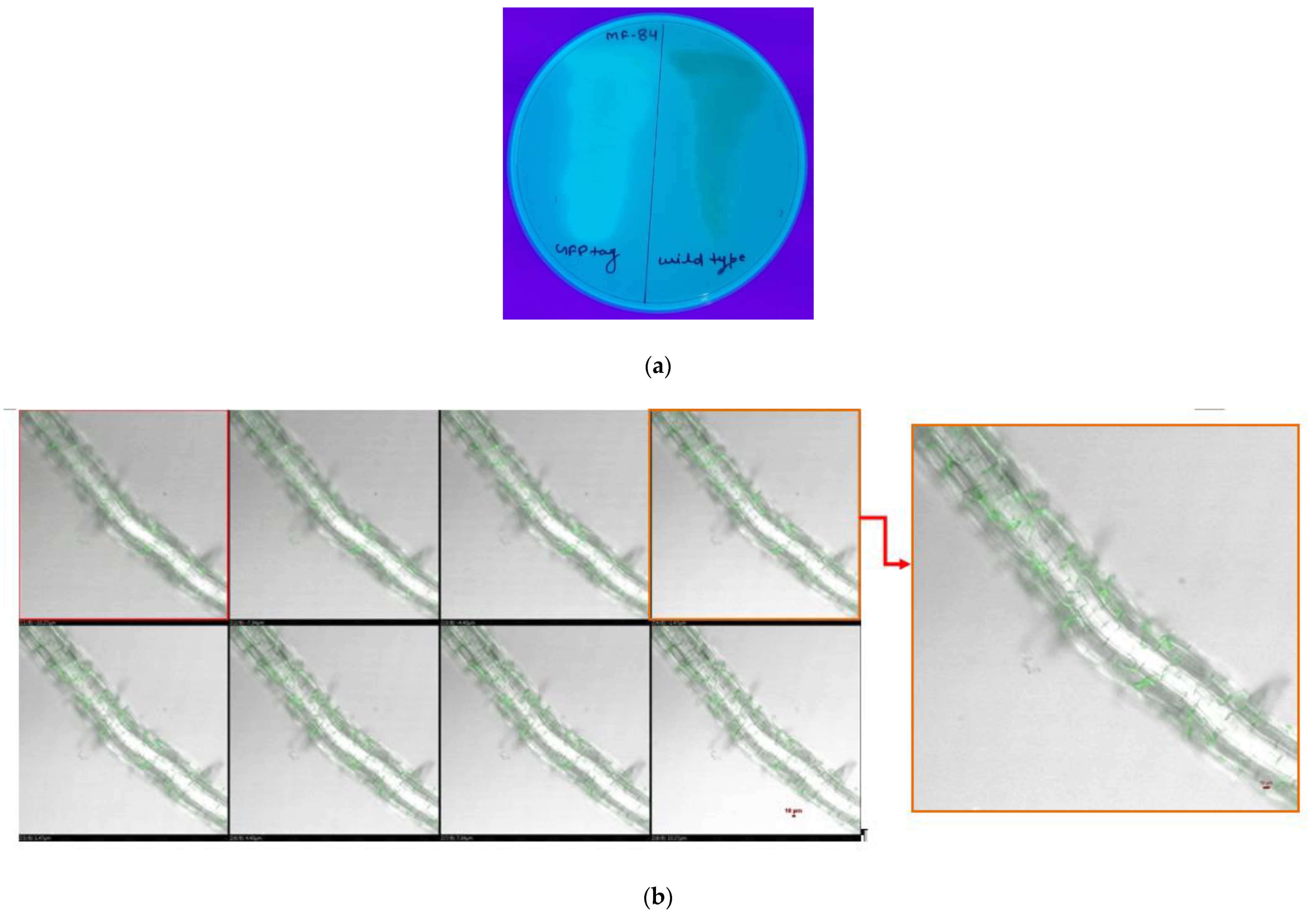
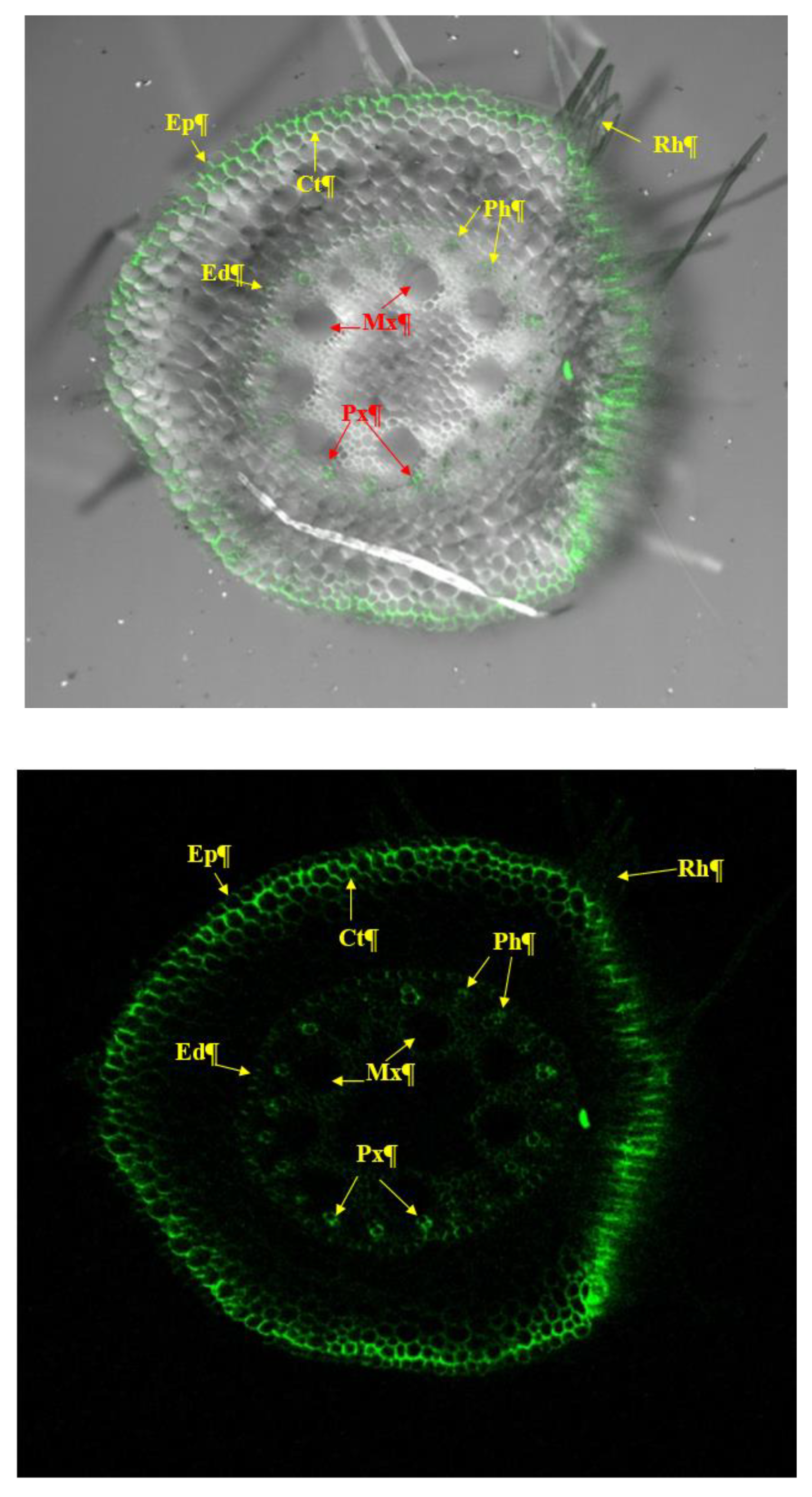
3.2. Root Colonization Assay
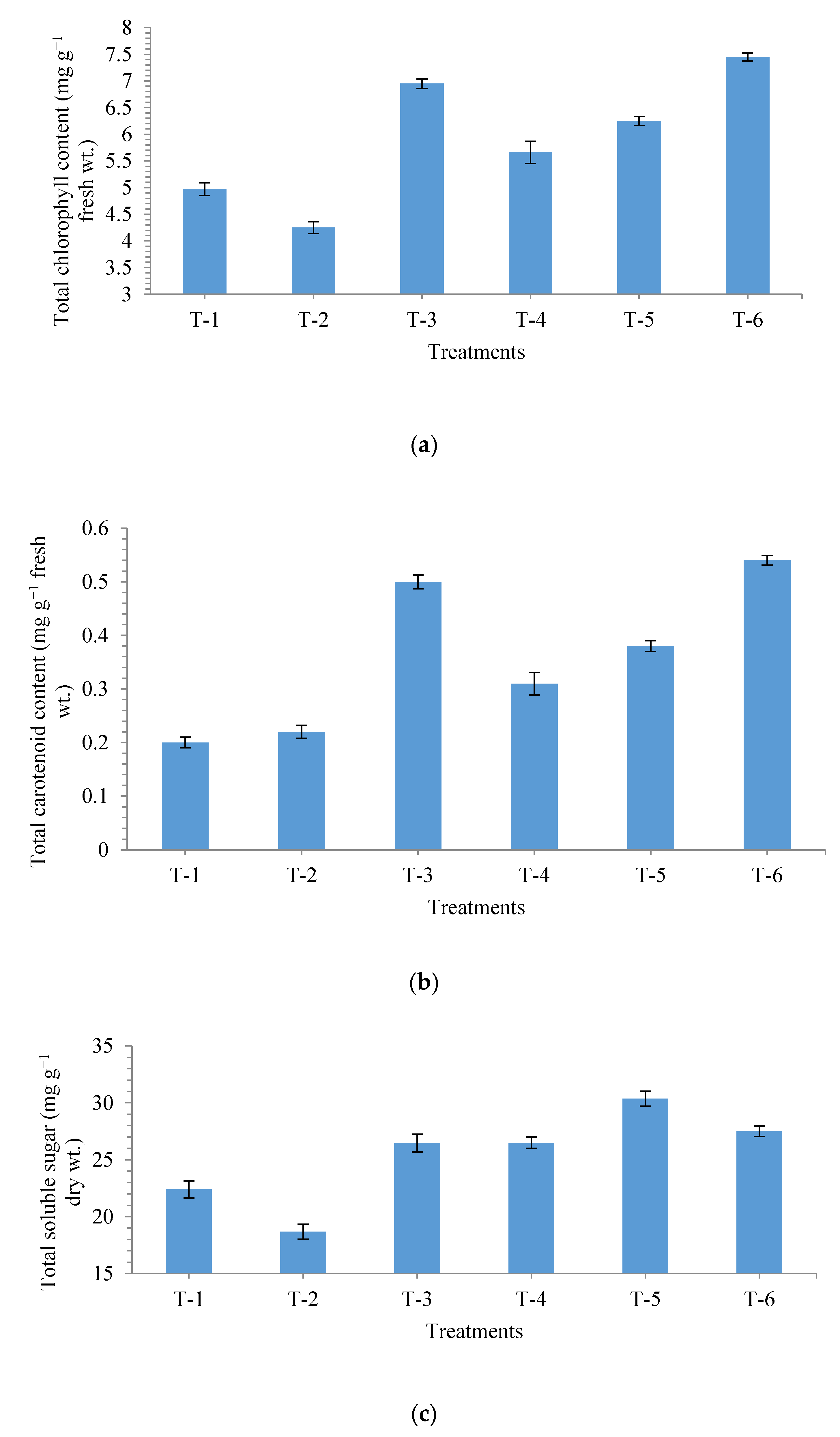
3.3. Effects of Seed-Biopriming on Biochemical Properties of Maize
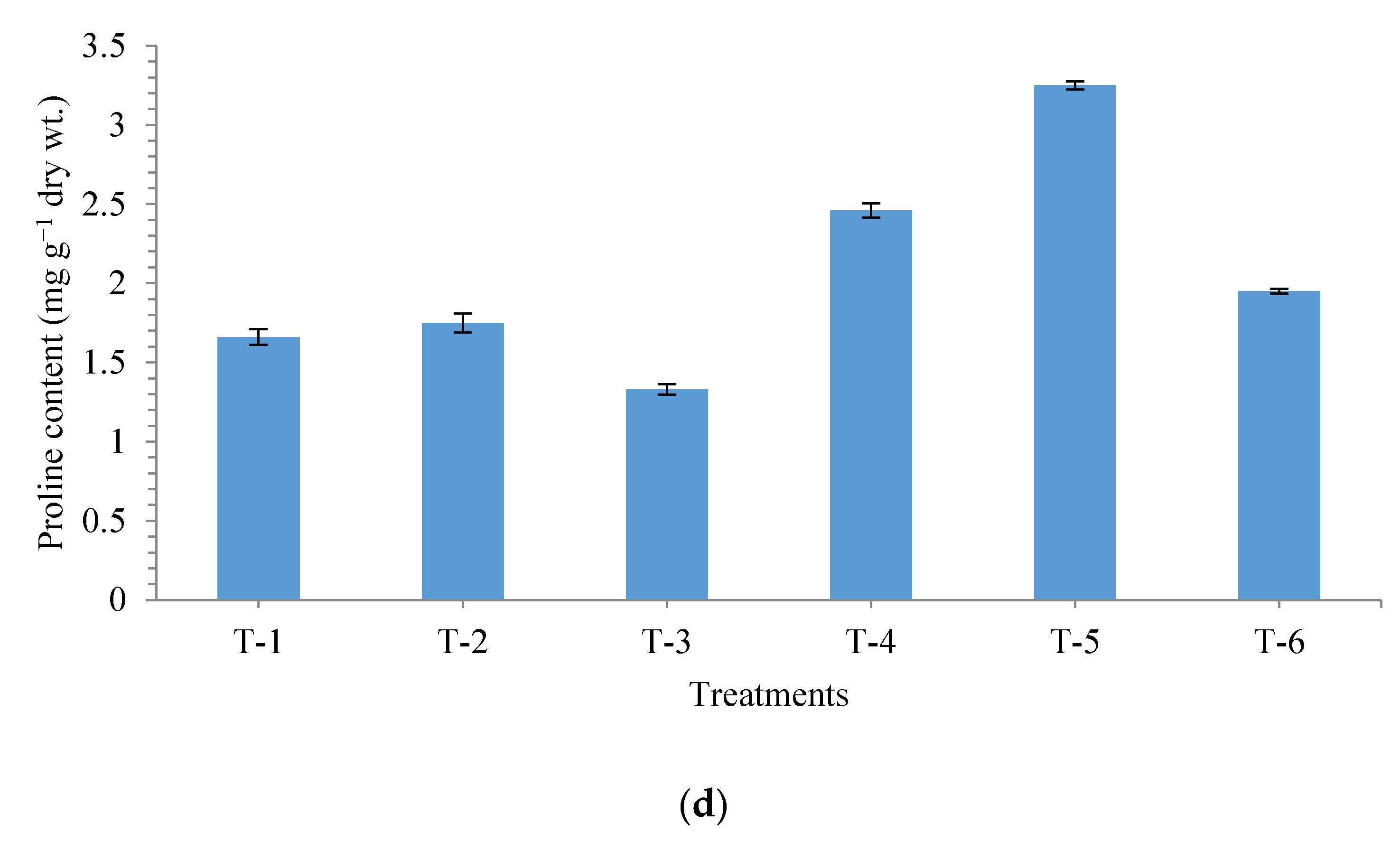
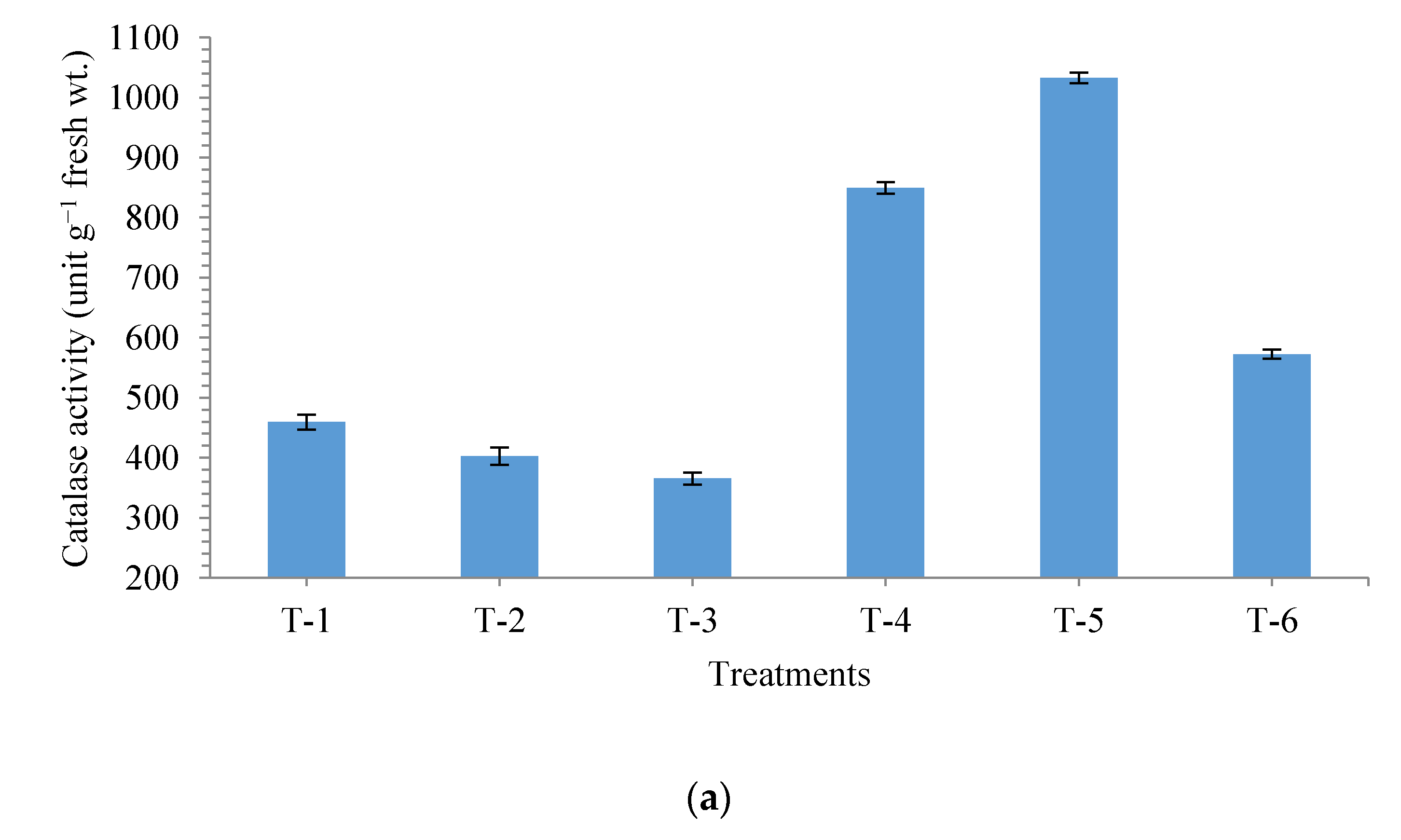
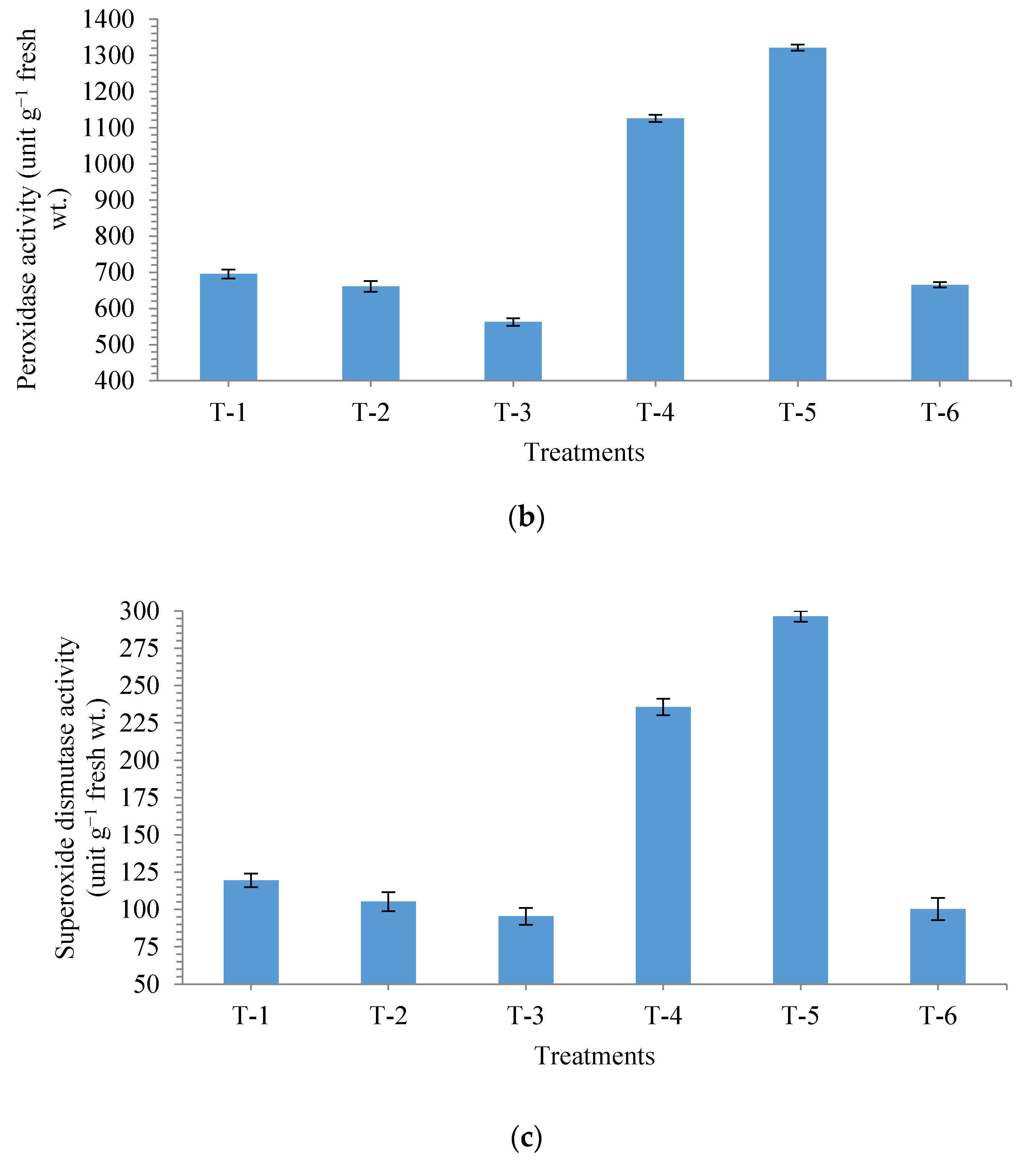
3.4. Effects of Seed Bio-Priming on Antioxidant Enzymes in Maize
3.5. Effects of Seed Bio-Priming on Plant Growth
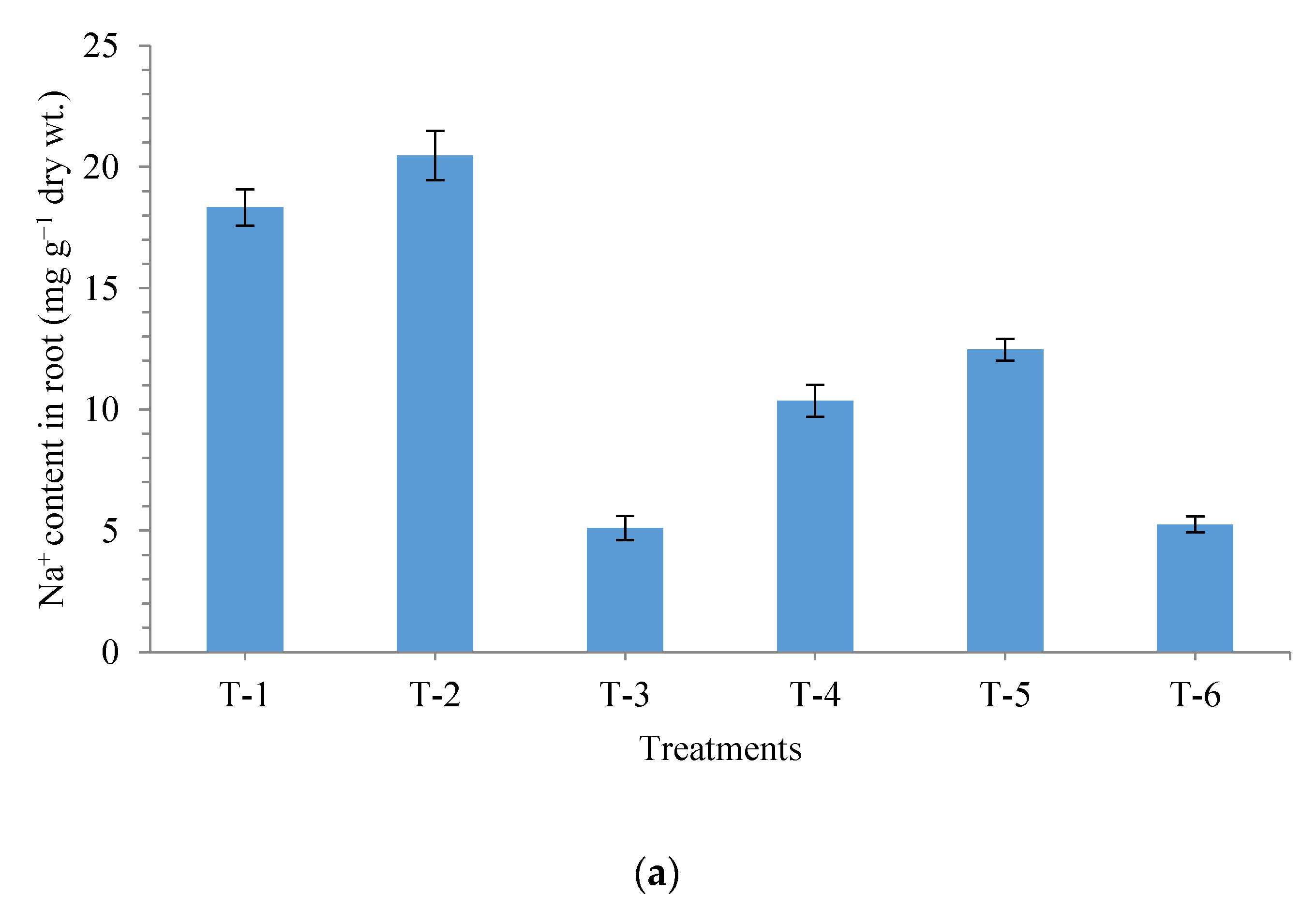
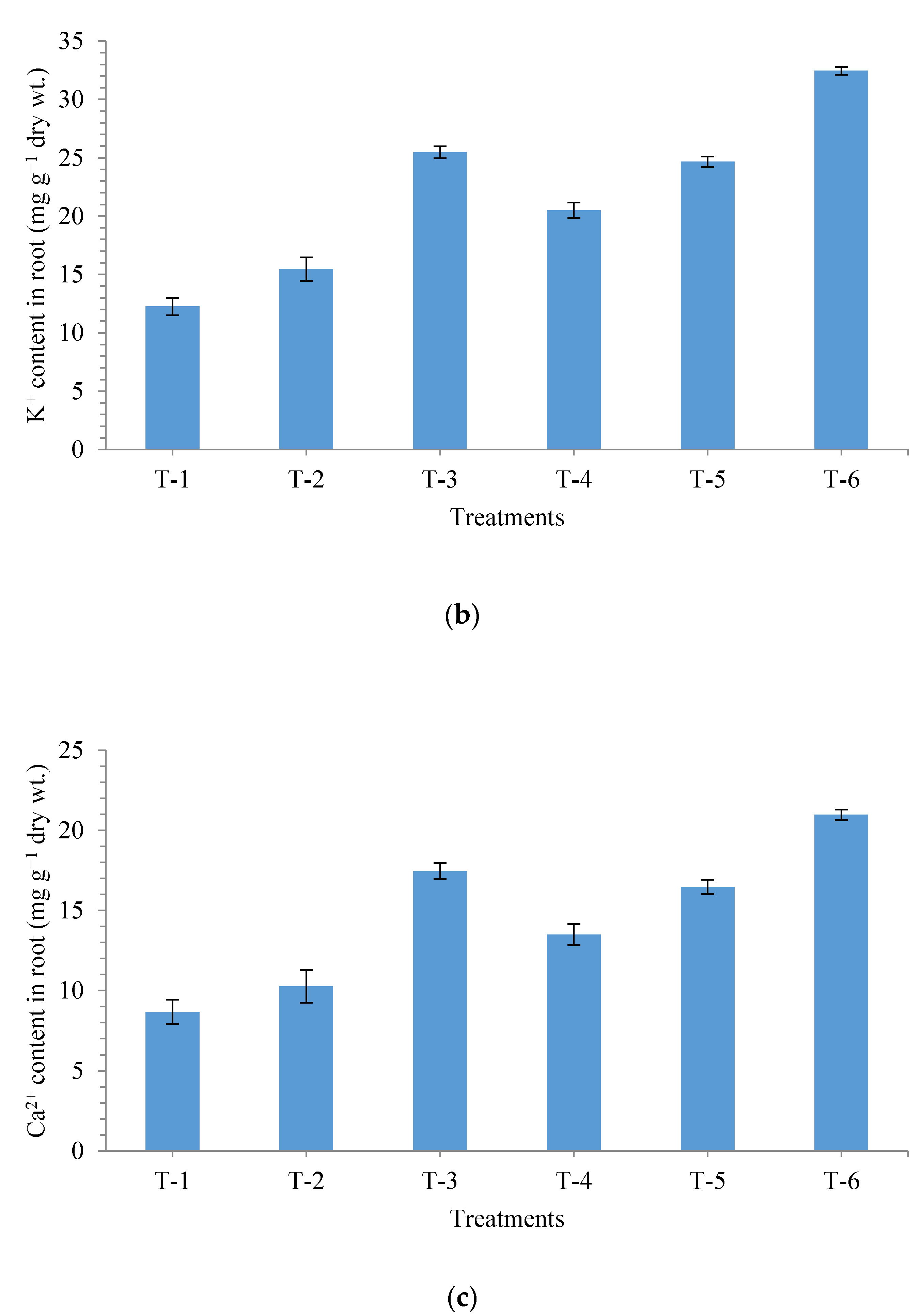
3.6. Effects of Seed Bio-Priming on Uptake of Na+, K+ and Ca2+
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anonymous. Annual Report (AICRP on Maize): Kharif Maize; Indian Institute of Maize Research: Ludhiyana, India, 2016; p. 318.
- Anonymous. Annual Report (AICRP on Maize): Rabi Maize; Indian Institute of Maize Research: Ludhiyana, India, 2017–2018; p. 992.
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issueand plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [Green Version]
- Shahbaz, M.; Ashraf, M. Improving salinity tolerance in cereals. Crit. Rev. Plant Sci. 2013, 32, 237–249. [Google Scholar] [CrossRef]
- Das, D.K. Introductory Soil Science; Kalyani Publishers: New Delhi, India, 2009; pp. 194–214. [Google Scholar]
- Blaylock, A.D. Soil Salinity, Salt Tolerance and Growth Potentialof Horticultural and Landscape Plants; Co-operative Extension Service, University of Wyoming, Department ofPlant, Soil and Insect Sciences, College of Agriculture: Laramie, WY, USA, 1994. [Google Scholar]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Imran, M.; Pathak, N.; Alam, M.; Rai, J.P.; Singh, R.K.; Sarma, B.K.; Sharma, P.K.; et al. Compatible salt-tolerant rhizosphere microbe-mediated induction of phenylpropanoid cascade and induced systemic responses against Bipolaris sorokiniana (Sacc.) Shoemaker causing spot blotch disease in wheat (Triticum aestivum L.). Appl. Soil Ecol. 2016, 108, 300–306. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Blumwald, E. Developing salt-tolerant cropplants: Challenges and Opportunities. Trends Plant Sci. 2005, 10, 615–620. [Google Scholar] [CrossRef] [PubMed]
- Flowers, T.J.; Garcia, A.; Koyama, M.; Yeo, A.R. Breeding of salt tolerance in crop plants-the role of molecular biology. Acta Physiol. Plant. 1997, 19, 427–433. [Google Scholar] [CrossRef]
- Dimkpa, C.; Weinand, T.; Ash, F. Plant-rhizobacteria interactions alleviate abiotic stress conditions. Plant Cell Environ. 2009, 32, 1682–1694. [Google Scholar] [CrossRef] [PubMed]
- Sarma, B.K.; Yadav, S.K.; Singh, S.; Singh, H.B. Microbial consortium mediated plant defense against phytopathogens: Readdressing for enhancing efficacy. Soil Biol. Biochem. 2015, 87, 25–33. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, W.; Meng, Y.; Xie, T.; Li, L.; Li, J.; Wei, S. γ-Aminobutyric acid imparts partial protection from salt stress injury to maize seedlings by improving photosynthesis and upregulating osmoprotectants and antioxidants. Sci. Rep. 2017, 7, 43609. [Google Scholar] [CrossRef]
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of exogenous selenium supply on photosynthesis, Na+ accumulation and antioxidative capacity of maize (Zea mays L.) under salinity stress. Sci. Rep. 2017, 7, 42039. [Google Scholar] [CrossRef] [Green Version]
- Nia, S.H.; Zarea, M.J.; Rejali, F.; Varma, A. Yield and yield components of wheat as affected by salinity and inoculation with Azospirillum strains from saline or non-saline soil. J. Saudi Soc. Agric. Sci. 2012, 11, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Ramadoss, D.; Lakkineni, V.K.; Bose, P.; Ali, S.; Annapurna, K. Mitigation of salt stress in wheat seedlings by halo tolerant bacteria isolated from saline habitats. Springer Plus 2013, 2, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yao, L.; Wu, Z.; Zheng, Y.; Kaleem, I.; Li, C. Growth promotion and protection against salt stress by Pseudomonas putida Rs-198 on cotton. Eur. J. Soil Biol. 2010, 46, 49–54. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.K.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by exo-polysaccharides producing Pseudomonas putida strain P45. Biol. Fertil. Soil 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Bona, E.; Scarafoni, A.; Marsano, F.; Boatti, L.; Copetta, A.; Massa, N.; Gamalero, E.; D’Agostino, G.; Cesaro, P.; Cavaletto, M.; et al. Arbuscular mycorrhizal symbiosis affects the grain proteome of Zea mays: A field study. Sci. Rep. 2016, 6, 26439. [Google Scholar] [CrossRef] [Green Version]
- Arora, N.K.; Khare, E.; Mishra, J. Multifacete interactins between endophytes and plants: Devlopment and prospects. Front. Microbiol. 2018, 6, 26439. [Google Scholar]
- Liu, S.; Hao, H.; Lu, X.; Zhao, X.; Wang, Y.; Zhang, Y.; Xie, Z.; Wang, R. Transcriptome profiling of genes involved in induced systemic salt tolerance conferred by Bacillus amyloliquefaciens FZB42 in Arabidopsis thaliana. Sci. Rep. 2017, 7, 10795. [Google Scholar] [CrossRef] [Green Version]
- Bano, A.; Fatima, M. Salt tolerance in Zea mays (L.) following inoculation with Rhizobium and Pseudomonas. Biol. Fertil. Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Paul, D.; Lade, H. Plant-growth-promoting rhizobacteria to improve crop growth in saline soils: A review. Agron. Sustain. Dev. 2014, 34, 737–752. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kim, M.S.; Sun, Y.; Dowd, S.E.; Shi, H.; Pare, P.W. Soil bacteria confer plant salt tolerance by tissue-specific regulation of the sodium transporter HKT1. Mol. Plant Microbe Interact. 2008, 21, 737–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhinandan, K.; Skori, L.; Stanic, M.; Hickerson, N.M.N.; Jamshed, M.; Samuel, M.A. Abiotic stress signaling in wheat—An inclusive overview of hormonal interactions during abiotic stress responses in wheat. Front. Plant Sci. 2018, 9, 734. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Sugawara, M.; Okunou, K.; Nakayama, H.; Schroeder, J.I.; Shinmyo, A.; Yoshida, K. Functions of HKT transporters in sodium transport in roots and in protecting leaves from salinity stress. Plant Biotech. 2009, 25, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Haynes, R.J.; Swift, R.S. Stability of soil aggregates in relation to organic constituents and soil water content. J. Soil Sci. 1990, 41, 73–83. [Google Scholar] [CrossRef]
- Singh, U.B.; Malviya, D.; Singh, S.; Pradhan, J.K.; Singh, B.P.; Roy, M.; Imram, M.; Pathak, N.; Baisyal, B.M.; Rai, J.P.; et al. Bio-protective microbial agents from rhizosphere eco-systems trigger plant defense responses provide protection against sheath blight disease inrice (Oryza sativa L.). Microbiol. Res. 2016, 192, 300–312. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall Till Blockerl, H.; Emde, M.; Bottger, E.C. Isolation anddirect complete nucleotide determination of entire genes: Characterization of agene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [Green Version]
- Singh, U.B.; Malviya, D.; Singh, S.; Kumar, M.; Sahu, P.K.; Singh, H.V.; Kumar, S.; Roy, M.; Imran, M.; Rai, J.P.; et al. Trichoderma harzianum- and methyl jasmonate-induced resistance to Bipolaris sorokiniana through enhanced phenylpropanoid activities in bread wheat (Triticum aestivum L.). Front. Microbiol. 2019, 10, 1697. [Google Scholar] [CrossRef] [Green Version]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International (P) Ltd.: New Delhi, India, 1996; p. 256. [Google Scholar]
- Thimmaiah, S.R. Standard Methods of Biochemical Analysis; Kalyani Publishers: New Delhi, India, 2012; pp. 421–426. [Google Scholar]
- Lee, E.H.; Bennet, J.H. Superoxide Dismutase. Plant Physiol. 1982, 69, 1444–1449. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, S.; Srinivas, V.; Prakash, B.; Sathya, A.; Vijayabharathi, R. Plant growth-promoting traits of Pseudomonas geniculata isolated from chickpea nodules. 3 Biotech 2015, 5, 653–661. [Google Scholar] [CrossRef] [Green Version]
- Danhorn, T.; Fuqua, C. Biofilm formation by plant-associated bacteria. Annu. Rev. Microbiol. 2007, 61, 401–422. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeier, S.; Mancuso, M.; Tecon, R.; Harms, H.; van der Meer, J.R.; Wells, M. Bioreporters: Gfpversus lux revisited and single-cell response. Biosens. Bioelectron. 2007, 22, 1578–1585. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Kloepper, J.W.; Ryu, C.M. Rhizosphere bacteria helpplants tolerate abiotic stress. Trends Plant Sci. 2009, 14, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, A.; Naeem, A.H.; Asghar, M. Inducing salt tolerance in mung bean through coinoculation with rhizobia and plant-growth-promoting rhizobacteria containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011, 57, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Arshad, M. The combined application of rhizobial strains and plant growth promoting rhizobacteria improves growth and productivity of mung bean (Vigna radiata L.) under salt-stressed conditions. Ann. Microbiol. 2012, 62, 1321–1330. [Google Scholar] [CrossRef]
- Ahmad, M.; Zahir, Z.A.; Nazli, F.; Akram, F.; Arshad, M.; Khalid, M. Effectiveness of halotolerant, auxin producing Pseudomonas and Rhizobium strains to improve osmotic stress tolerance in mung bean (Vigna radiata L.). Braz. J. Microbiol. 2013, 44, 1341–1348. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Wei, H.; Cao, J.; Liu, R.; Wang, Y.; Zheng, C. Expression of Bacillus subtilis proAB genes and reduction of feedback inhibition of proline synthesis increases proline production and confers osmo-tolerance in transgenic Arabdopsis. J. Biochem. Mol. Biol. 2007, 40, 396–403. [Google Scholar]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant growth promoting rhizobacteria Dietzia natronolimnaea modulates the expression of stress responsive genes providing protection of wheat from salinity stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef] [Green Version]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 2nd ed.; Springer: Berlin, Germany, 1980. [Google Scholar]
- Dodd, I.C.; Perez-Alfocea, F. Microbial amelioration of crop salinity stress. J. Exp. Bot. 2012, 63, 3415–3428. [Google Scholar] [CrossRef] [Green Version]
- Boiero, L.; Perrig, D.; Masciarelli, O.; Penna, C.; Cassan, F.; Luna, V. Phytohormone production by three strains of Bradyrhizobium japonicum and possible physiological and technological implications. Appl. Microbiol. Biotechnol. 2007, 74, 874–880. [Google Scholar] [CrossRef]
- Xu, Z.H.; Saffigna, P.G.; Farquhar, G.D.; Simpson, J.A.; Haines, R.J.; Walker, S. Carbon isotope discrimination and oxygen isotope composition in clones of the F(1) hybrid between slash pine and Caribbean pine in relation to tree growth, water-use efficiency and foliar nutrient concentration. Tree Physiol. 2000, 20, 1209–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, Y.P.; Mishra, V.K.; Singh, S.; Sharma, D.K.; Singh, D.; Singh, U.S.; Singh, R.K.; Haefele, S.M.; Ismail, A.M. Productivity of sodic soils can be enhanced through the use of salt-tolerant rice varieties and proper agronomic practices. Field Crops Res. 2016, 190, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, R.P.; Jha, P.; Jha, P.N. The plant-growth-promoting bacterium Klebsiella sp. SBP-8 confers induced systemic tolerance in wheat (Triticum aestivum) under salt stress. J. Plant Physiol. 2015, 184, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Hu, X.; Wang, S.; Tian, L.; Pang, Y.; Han, Z.; Wu, L.; Chen, Y. Quantitative analysis of changes in the phosphoproteome of maize induced by the plant hormone salicylic acid. Sci. Rep. 2015, 5, 18155. [Google Scholar] [CrossRef]
- Hu, X.; Li, N.; Wu, L.; Li, C.; Li, C.; Zhang, L.; Liu, T.; Wang, W. Quantitative iTRAQ-based proteomic analysis of phosphoproteins and ABA-regulated phosphoproteins in maize leaves under osmotic stress. Sci. Rep. 2015, 5, 15626. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.S.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Cholineand osmotic-stress tolerance induced in Arabidopsis by the soil microbe Bacillus subtilis (GB03). Mol. Plant Microbe Interact. 2010, 23, 1097–1104. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Gan, Y.; Xu, B. Application of Plant-Growth- Promoting Fungi Trichoderma longibrachiatum T6 enhances tolerance of wheat to salt stress through improvement of antioxidative defense system and gene expression. Front. Plant Sci. 2016, 7, 1405. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [Green Version]
- Hichem, H.; Naceur, E.A.; Mounir, D. Effects of salt stress on photosynthesis, PS II photochemistry and thermal energy dissipation in leaves of two corn (Zea mays L.) varieties. Photosynthetica 2009, 47, 517–526. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Davranov, K.; Wirth, S.; Hashem, A.; Allah, E.F.A. Impact of soil salinity on the plant-growth-promoting and biological control abilities of root associated bacteria. Saudi J. Biol. Sci. 2017, 24, 1601–1608. [Google Scholar] [CrossRef]
- Ebel, C.; BenFeki, A.; Hanin, M.; Solano, R.; Chini, A. Characterization of wheat (Triticum aestivum) TIFY family and role of Triticum Durum TdTIFY11a in salt stress tolerance. PLoS ONE 2018, 13, e0200566. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, P.J.; Broadley, M.R. Chloride in soils and its uptake and movement within the plant: A review. Ann. Bot. 2001, 88, 967–988. [Google Scholar] [CrossRef] [Green Version]
- Tester, M.; Davenport, R. Na+ tolerance and Na+ transport in higher plants. Ann. Bot. 2003, 91, 503–527. [Google Scholar] [CrossRef] [PubMed]
- Santoyoa, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Parida, A.K.; Das, A.B. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf. 2005, 60, 324–349. [Google Scholar] [CrossRef]
- Liu, S.H.; Kang, Y.H. Changes of soil microbial characteristics in saline-sodic soils under drip irrigation. J. Soil Sci. Plant Nutr. 2014, 14, 139–150. [Google Scholar] [CrossRef]
- Kumar, K.; Manigundan, K.; Natarajan, A. Influence of salt-tolerant Trichoderma spp. on growth of maize (Zea mays) under different salinity conditions. J. Basic Microbiol. 2016, 56, 1–10. [Google Scholar]
- Kasim, W.A.; Gaafar, R.M.; Abou-Ali, R.M.; Omar, M.N.; Hewait, H.M. Effect of biofilm forming plant growth promoting rhizobacteria on salinity tolerance in barley. Ann. Agric. Sci. 2016, 61, 217–227. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Smith, D.L. Plant growth promoting rhizobacteria in amelioration of salinity stress: A systems biology perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef]
- Wong, V.N.L.; Greene, R.S.B.; Dalal, R.C.; Murphy, B.W. Soil carbon dynamics in saline and sodic soils: A review. Soil Use Manag. 2010, 26, 2–11. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Soil Properties | Saline Sodic Soil | Non-Saline Sodic Soil |
|---|---|---|---|
| 1. | Soil type | Silty clay loam | Silty clay loam |
| 2. | pH | 8.5 | 7.2 |
| 3. | EC (dS m−1) | 2.58 | 0.45 |
| 4. | OC (g kg−1) | 3.50 | 5.50 |
| 5. | OM (g kg−1) | 7.25 | 9.46 |
| 6. | Available macronutrients (kg ha−1) | ||
| I. | Nitrogen | 175.33 | 205.66 |
| II. | Phosphorous | 32.45 | 42.36 |
| III. | Potassium | 142.50 | 162.75 |
| IV. | Sulphur | 6.45 | 8.26 |
| V. | |||
| 7. | Available micronutrients (mg kg−1) | ||
| I. | Fe | 4.26 | 11.50 |
| II. | Cu | 0.52 | 0.91 |
| III. | Zn | 0.75 | 0.82 |
| IV. | B | 0.09 | 0.12 |
| Treatments Code | Treatment Details |
|---|---|
| T1 | Saline sodic soil (Negative control) |
| T2 | Non-saline sodic soil + Salt (NaCl 150 mM) |
| T3 | Non-saline sodic soil (Positive control) |
| T4 | Saline sodic soil + Pseudomonas geniculata MF-84 |
| T5 | Non-saline sodic soil + Salt (NaCl 150 mM) + Pseudomonas geniculata MF-84 |
| T6 | Non-saline sodic soil + Pseudomonas geniculata MF-84 |
| Treatments | Shoot Length (cm) | Root Length (cm) | Fresh wt. of Shoot (g) | Fresh wt. of Root (g) | Dry wt. of Shoot (g) | Dry wt. of Root (g) |
|---|---|---|---|---|---|---|
| T1 | 15.25 ± 0.75 d | 12.50 ± 0.66 c | 4.97 ± 0.22 d | 2.15 ± 0.16 e | 1.05 ± 0.05 d | 0.66 ± 0.01 c |
| T2 | 14.37 ± 1.05 d | 10.47 ± 0.54 d | 3.42 ± 0.35 e | 2.05 ± 0.25 e | 0.97 ± 0.12 d | 0.75 ± 0.03 c |
| T3 | 22.75 ± 1.33 c | 18.36 ± 0.85 b | 6.50 ± 0.25 b | 3.97 ± 0.31 b | 1.66 ± 0.09 b | 1.02 ± 0.04 b |
| T4 | 21.92 ± 0.95 c | 17.89 ± 1.25 b | 5.90 ± 0.15 c | 3.15 ± 0.17 d | 1.35 ± 0.04 c | 1.15 ± 0.04 b |
| T5 | 23.46 ± 1.15 b | 20.50 ± 1.01 b | 4.87 ± 0.25 d | 3.36 ± 0.21 c | 1.42 ± 0.05 c | 1.25 ± 0.05 b |
| T6 | 28.35 ± 1.26 a | 24.78 ± 0.96 a | 8.05 ± 0.33 a | 5.25 ± 0.28 a | 2.25 ± 0.11 a | 1.76 ± 0.02 a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, S.; Singh, U.B.; Trivedi, M.; Sahu, P.K.; Paul, S.; Paul, D.; Saxena, A.K. Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas geniculata-Modulated Biochemical Responses Provide Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil. Int. J. Environ. Res. Public Health 2020, 17, 253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010253
Singh S, Singh UB, Trivedi M, Sahu PK, Paul S, Paul D, Saxena AK. Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas geniculata-Modulated Biochemical Responses Provide Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil. International Journal of Environmental Research and Public Health. 2020; 17(1):253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010253
Chicago/Turabian StyleSingh, Shailendra, Udai B. Singh, Mala Trivedi, Pramod Kumar Sahu, Surinder Paul, Diby Paul, and Anil Kumar Saxena. 2020. "Seed Biopriming with Salt-Tolerant Endophytic Pseudomonas geniculata-Modulated Biochemical Responses Provide Ecological Fitness in Maize (Zea mays L.) Grown in Saline Sodic Soil" International Journal of Environmental Research and Public Health 17, no. 1: 253. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17010253