Functional Trait-Based Screening of Zn-Pb Tolerant Wild Plant Species at an Abandoned Mine Site in Gard (France) for Rehabilitation of Mediterranean Metal-Contaminated Soils

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
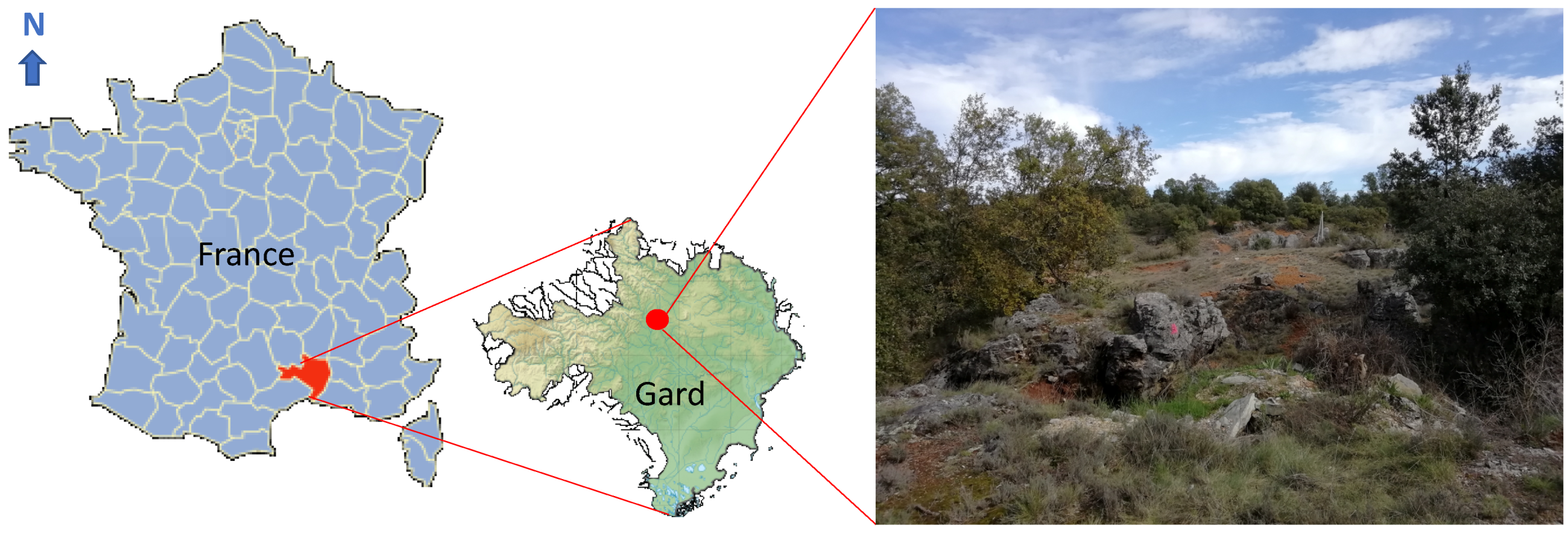
2.1. Site Location
2.2. Soil Sampling and Metal Analyses
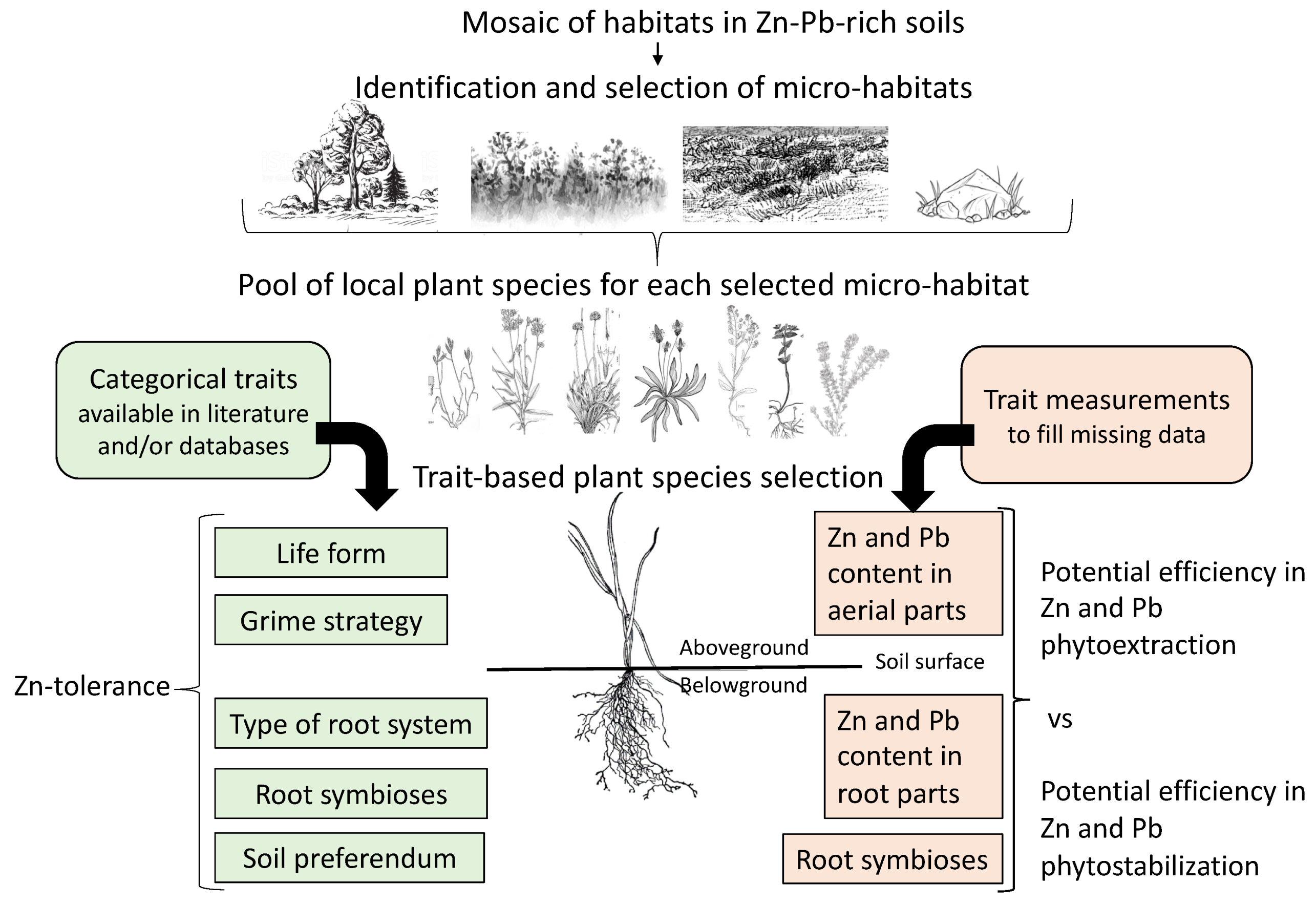
2.3. Experimental Approach for Plant Selection
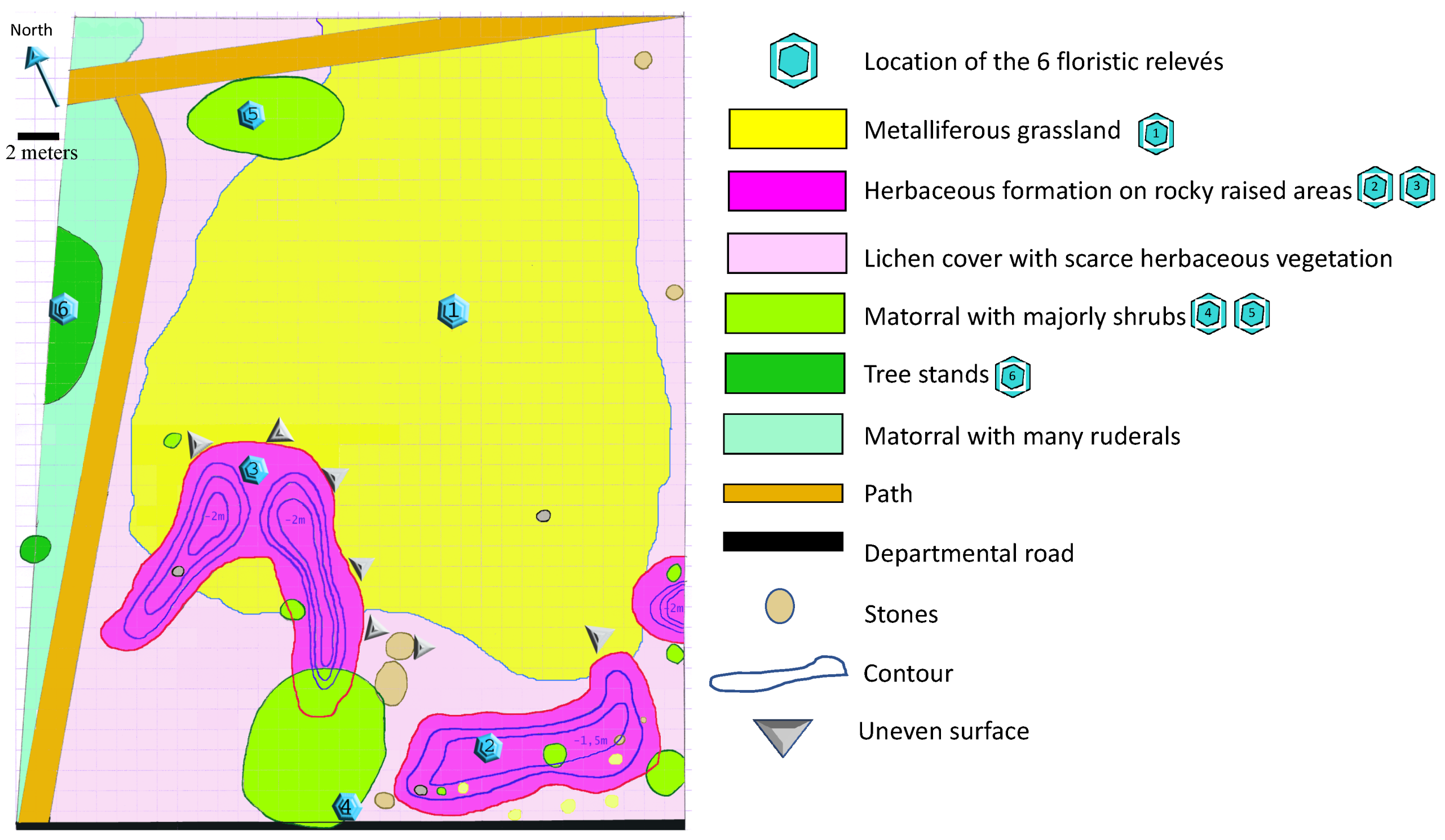
2.4. Floristic Analysis and Habitat Inventory
2.5. Bibliographical Functional Traits
2.6. Plant Analyses
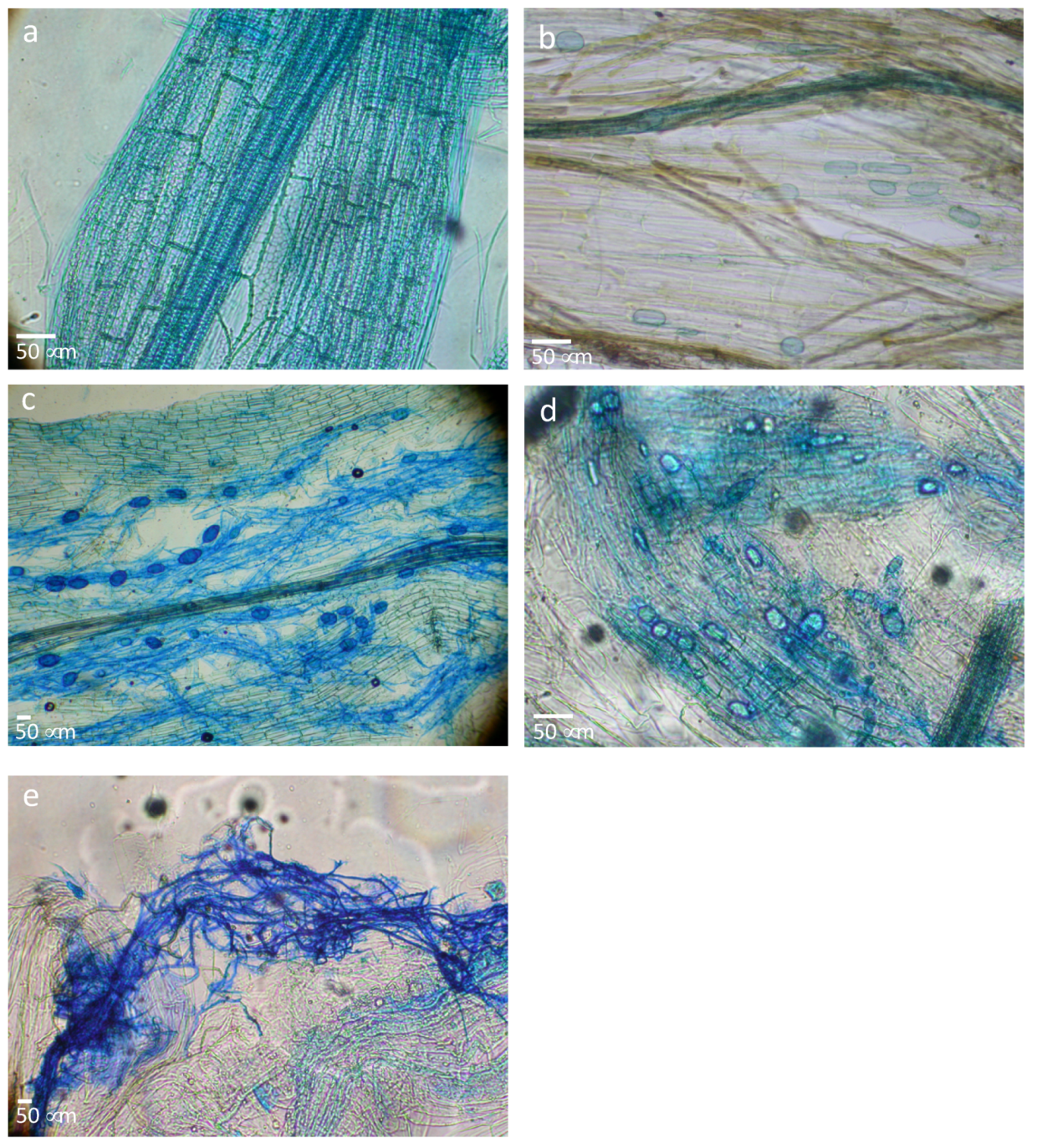
2.6.1. Root Symbiosis Assessment
2.6.2. Metal Content in Below-Ground and Above-Ground Plant Parts
2.7. Statistical Analysis
3. Results and Discussion
3.1. Soil Contamination
3.2. Diversity of Habitats and Plant Species
3.3. Analysis of Plant Traits Linked with TMM Tolerance
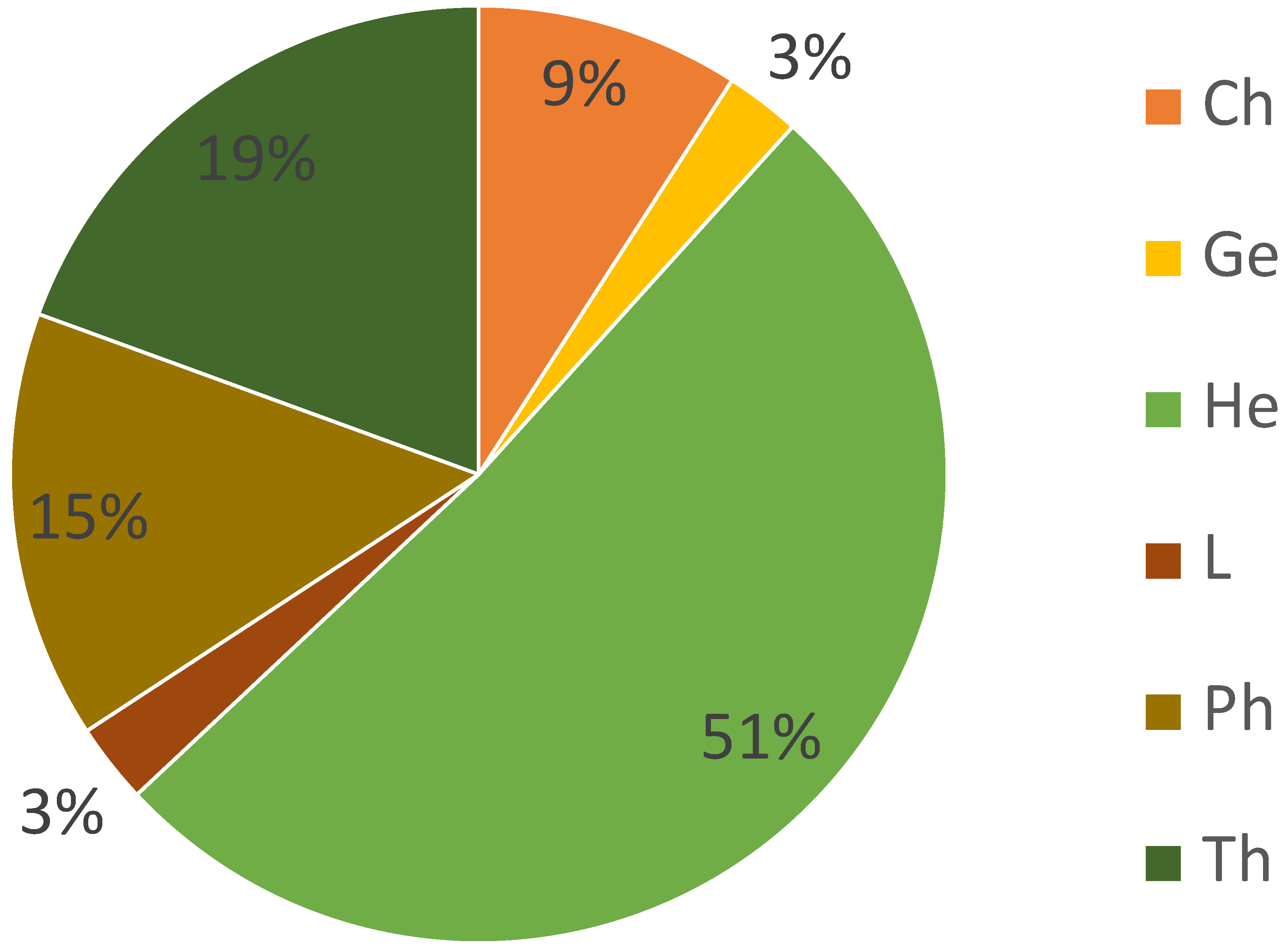
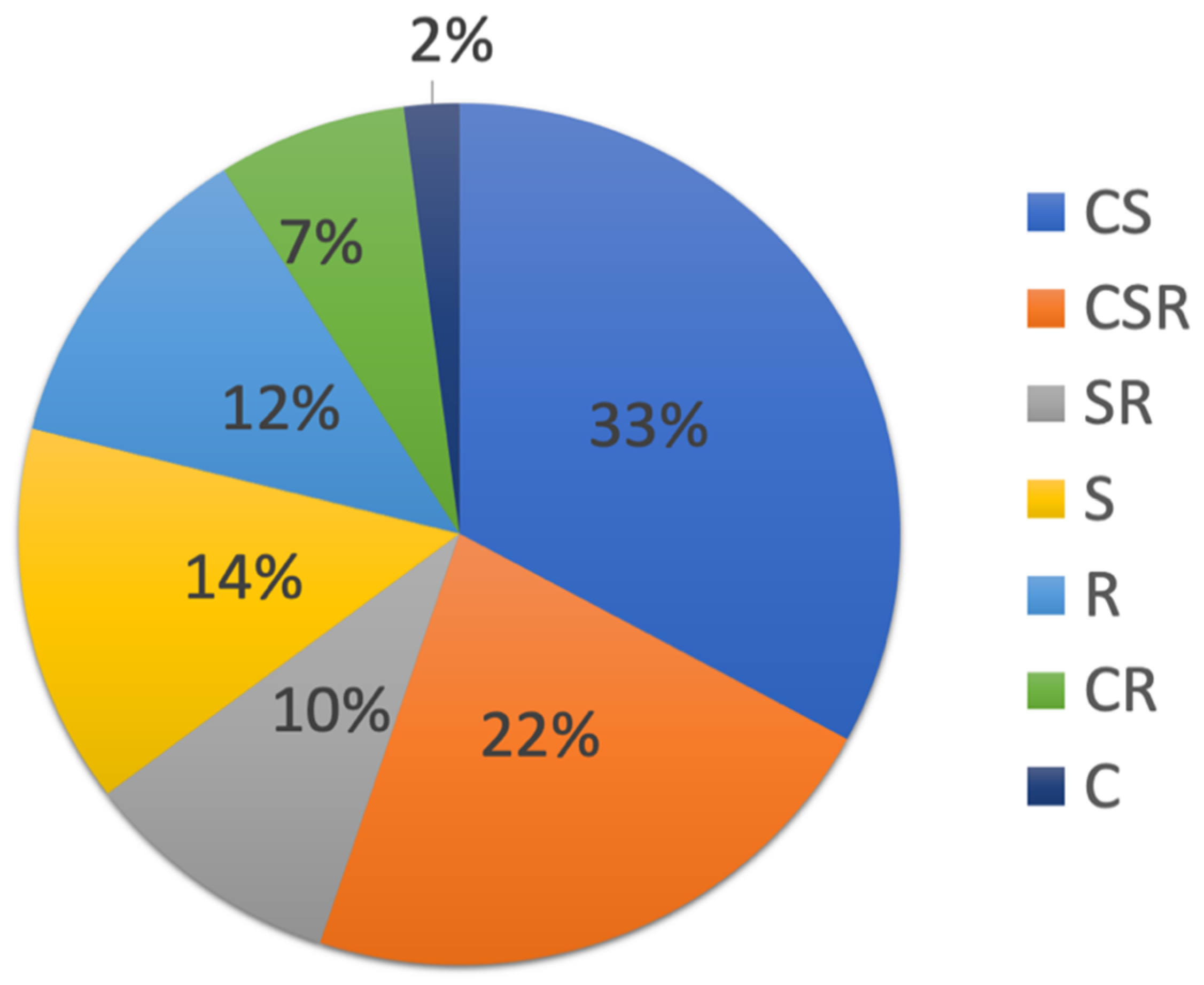
3.3.1. Categorical Trait Analysis of the 61 Identified Plant Species
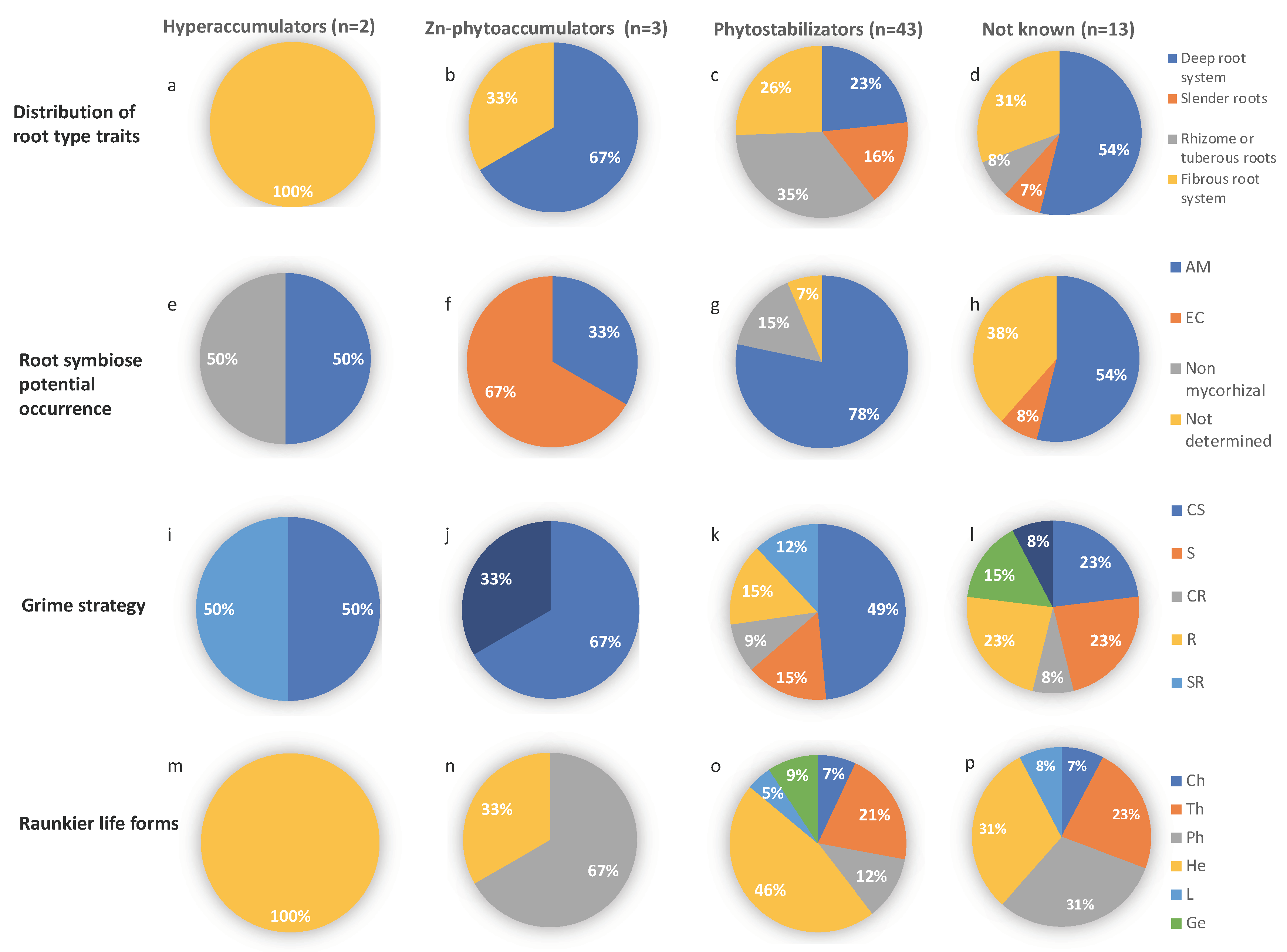
3.3.2. Analysis of Plant Traits by TMM Tolerance Strategy
3.3.3. Measured Trait Analysis on Selected Plant Species
3.3.4. Metal Content in 4 Plant Species for Phytoextraction Strategy
3.3.5. Spontaneous Plant Colonization at Abandoned Zn-Pb Mine Sites: A Matter of Geographical Situation or Plant Traits?
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Botanical Family | Root System | Root Symbioses | Soil Preferendum | Life Form | Grime Strategy | Zn Tolerance |
|---|---|---|---|---|---|---|---|
| Amelanchier ovalis Medik. | Rosaceae | Deep root system | AM [43] | Rocky dry soils | Ph | CS | Not known |
| Aphyllantes monspeliensis L. | Liliaceae | Fibrous root system | Not known | Dry soils | He | S | Not known |
| Arenaria serpyllifolia L. | Caryophyllaceae | Thin root system | NM [44,45] | Sandy and rocky soils, calcareous soils | Th | R | Population-specific Zn tolerance [46] |
| Argyrolobium zanonii (Turra) P.W.Ball | Fabaceae | Fibrous root system | AM [47] | Calcareous soils | Ch | S | Not known |
| Armeria arenaria subsp bupleuroides (Godr. & Gren.) Greuter & Burdet | Plumbaginaceae | Fibrous root system | Not known | Sandy and rocky soils | He | S | Population-specific Zn tolerance [48] |
| Asparagus acutifolius L. | Asparagaceae | Rhizome | AM [26] | Dry sand arid oils | He | CS | Population-specific Zn tolerance [20] |
| Asplenium ruta-muraria L. | Aspleniaceae | Rhizome | AM + NM [26] | Cliffs, rocks | He | CS | Population-specific Zn tolerance [49] |
| Biscutella laevigata L. | Brassicaceae | Fibrous root system | NM [26]/AM [27,28] | Rocky soils | He | SR | Species-wide Zn tolerance [8], Tl-, Cd-hyperaccumulator [50] |
| Brachypodium phoenicoides (L.) Roem. & Schult | Poaceae | Rhizome | AM [29] | Dry soils | He | CS | Population-specific Zn tolerance [19] |
| Brachypodium retusum (Pers.) P.Beauv. | Poaceae | Rhizome | AM [26] | Dry soils | He | CS | Population-specific Zn tolerance [8] |
| Bromus madritensis L. | Poaceae | Fibrous root system | AM [26] | Sandy soils, cultivated soils | Th | SR | Population-specific Zn tolerance [51] |
| Buxus sempervirens L. | Buxaceae | Deep root system | AM [26] | Dry calcareous soils | Ph | CS | Population-specific Zn tolerance [35] |
| Carex halleriana Asso | Cyperaceae | Rhizome | NM [52] | Dry calcareous soils | He | CSR | Population-specific Zn tolerance [37] |
| Centaurea pectinata L. | Asteraceae | Deep root system | Not known | Rocky soils | He | S | Population-specific Zn tolerance [20] |
| Cerastium pumilum Curtis | Caryophyllaceae | Slender root system | Not known | Sandy soils | Th | SR | Population-specific Zn tolerance [53] |
| Clematis vitalba L. | Ranunculaceae | fibrous root system | AM [26] | calcareous to acidic soils | L | CS | Population-specific Zn tolerance [54] |
| Clinopodium nepeta (L.) Kuntze | Lamiaceae | fibrous root system | AM [55] | Dry rocky soils | He | S | Population-specific Zn tolerance [56,57] |
| Dactylis glomerata L. | Poaceae | Fibrous root system | AM [26] | Cultivated soils | He | CSR | Population-specific Zn tolerance [20] |
| Dioscorea communis (L.) Caddick & Wilkin | Dioscoreaceae | Rhizome | AM [25] | Calcareous, well-drained soils | L | CS | Not known |
| Draba verna L. | Brassicaceae | Fibrous root system | Not known | Sandy and calcareous soils | Th | CR | Not known |
| Eryngium campestre L. | Apiaceae | Rhizome | AM [26] | Sandy soils | Ge | SR | Population-specific Zn tolerance [16] |
| Euphorbia cyparissias L. | Euphorbiacee | Fibrous root system | AM [26,58] | Calcareous soils | He | S | Population-specific Zn tolerance [58] |
| Festuca ovina sl L. | Poaceae | Fibrous root system | AM [26,27] | Dry soils | He | CSR | Population-specific Zn tolerance [59] |
| Galium aparine L. | Rubiaceae | Slender root system | AM + NM [26] | Cultivated soils | Th | CR | Population-specific Zn tolerance [60] |
| Galium corrudifolium Vill. | Rubiaceae | Deep and fibrous root system | Not known | Dry soils | He | CR | Not known |
| Helleborus foetidus L. | Ranunculaceae | Rhizome | AM [26] | Well- drained, calcareous soil | Ge | CSR | Population-specific Zn tolerance [17] |
| Hordeum murinum L. | Poaceae | Fibrous root system | AM [26] | Dry and sandy soils, disturbed soils | Th | R | Population-specific Zn tolerance [61] |
| Hornungia petraea (L.) ex Rchb. | Brassicaceae | Fibrous root system | Not known | Sandy and rocky soils | Th | R | Not known |
| Hypericum perforatum L. | Hypericaceae | Rhizome | AM [26] | Roadside soils, dry soils | He | SR | Population-specific Zn tolerance [62] |
| Juniperus oxycedrus L. | Cupressaceae | Deep root system | AM [26] | Arid soils | Ph | CS | Population-specific Zn tolerance [63] |
| Lactuca perennis L. | Asteraceae | Deep root system | Not known | Rocky and calcareous soils | He | S | Not known |
| Lepidium draba L. | Brassicaceae | Rhizome | NM [64] | Roadside soils | Ge | CR | Population-specific Zn tolerance [65] |
| Lysimachia arvensis (L.) U. Manns & Anderb. | Primulaceae | Fibrous root system | AM [25] | Cultivated and sandy soils | Th | R | Not known |
| Mibora minima (L.) Desv. | Poaceae | Slender root | Not known | Sandy soils | Th | R | Population-specific Zn tolerance [66] |
| Noccaea caerulescens (J.Presl & C.Presl) F.K.Mey | Brassicaceae | Fibrous root system | AM [26] | Well-drained soils, cultivated soils | He | CS | Species-wide Zn tolerance [20], Zn/Cd/Ni hyperaccumulator [3] |
| Pilosella officinarum Vaill. | Asteraceae | Rhizome | AM [67] | Dry soils | He | CS | Population-specific Zn tolerance [68] |
| Pinus sylvestris L. | Pinaceae | Deep root system | EC [26] | Dry soils, sandy and rocky soils | Ph | CS | Population-specific Zn tolerance [69], Zn accumulator [69] |
| Pistacia terebinthus L. | Anacardiaceae | Deep root system | AM [26] | Calcareous soils | Ph | CS | Population-specific Zn tolerance [70] |
| Plantago lanceolata L. | Plantaginaceae | Fibrous root system | AM [25] | Neutral to calcareous soils, rangelands | He | CSR | Population-specific Zn tolerance [20], Zn- accumulator [62] |
| Poa annua L. | Poaceae | Fibrous root system | AM + NM [26] | Well-drained soil | Th | R | Population-specific Zn tolerance [71] |
| Poterium sanguisorba L. | Rosaceae | deep-root system | AM [72] | Roadside soils, Rocky soils | He | CS | Population-specific Zn tolerance [20] |
| Pyrus spinosa Forssk. | Rosaceae | deep-root system | AM + NM [26] | Dry soils, Rocky soils | Ph | C | Not known |
| Quercus ilex L. | Fagaceae | deep-root system | EC [26] | Calcareous soils, well-drained soils | Ph | CS | Population-specific Zn tolerance [18], Zn- accumulator [18] |
| Quercus pubescens Willd. | Fagaceae | deep-root system | EC [26] | Well-drained soils, Calcareous soils | Ph | C | Not known |
| Ranunculus bulbosus L. | Ranunculaceae | Tuberous root system | AM [26,27] | Calcareous soils, Stony soil | Ge | CSR | Population-specific Zn tolerance [21] |
| Reseda lutea L. | Resedaceae | deep-root system | AM + NM [26,27] | Sandy soils, roadside soils, rangelands | He | CSR | Population-specific Zn tolerance [20] |
| Rosa canina L. | Rosaceae | deep-root system | AM [26] | Poor soils, sandy soils | Ph | CS | Population-specific Zn tolerance [73] |
| Rubia peregrina L. | Rubiaceae | Rhizome | AM [26] | Dry soils, well-drained soils | He | CS | Population-specific Zn tolerance [19] |
| Rubus ulmifolius Schott | Rosaceae | Rhizome | AM [26] | Calcareous soils | Ph | CS | Population-specific Zn tolerance [19] |
| Rumex intermedius D.C. | Polygonaceae | deep-root system | AM [26] | Rocky soils, dry soils | He | CSR | Not known |
| Ruscus aculeatus L. | Asparagaceae | Rhizome | AM [25] | Dry soils, rocky soils | Ch | CS | Population-specific Zn tolerance [19] |
| Scabiosa atropurpurea L. | Caprifoliaceae | Deep root system | Not known | Sandy soils, rangelands | He | CSR | Population-specific Zn tolerance [74] |
| Scrophularia lucida L. | Scrophulariaceae | Slender roots | Not known | Dry calcareous soils | He | CSR | Population-specific Zn tolerance [75] |
| Sedum acre L. | Crassulaceae | Slender roots | AM [26] | Rocky soils | Ch | S | Population-specific Zn tolerance [36] |
| Sedum annuum L. | Crassulaceae | Slender roots | Not known | Dry soils | Th | R | Not known |
| Senecio vulgaris L. | Asteraceae | Deep root system | AM [26,76] | Cultivated soils | Th | R | Population-specific Zn tolerance [36] |
| Silene vulgaris (Moench) Garcke | Caryophyllaceae | Slender roots | NM [27] | Well-drained soils, rangelands | He | CSR | Population-specific Zn tolerance [8] |
| Smilax aspera L. | Smilacaceae | Rhizome | AM [25] | Dry soils | L | CS | Population-specific Zn tolerance [77] |
| Thymus vulgaris L. | Lamiaceae | Fibrous root system | AM [26] | Dry soils | Ch | CS | Population-specific Zn tolerance [20] |
| Trifolium pratense L. | Fabaceae | Deep root system | AM + NM [26,27] | Meadows, woods | He | CSR | Population-specific Zn tolerance [27] |
| Ulex parviflorus Pourr. | Fabaceae | Deep root system | AM [26] | Dry, rocky soils, poor soils | Ph | CS | Not known |
References
- Bolan, N.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of Heavy Metal(Loid)s Contaminated Soils—To Mobilize or to Immobilize? J. Hazard. Mater. 2014, 266, 141–166. [Google Scholar] [CrossRef] [PubMed]
- Affholder, M.-C.; Laffont-Schwob, I.; Coulomb, B.; Rabier, J.; Borla, A.; Boudenne, J.-L.; Demelas, C.; Prudent, P. Implication of Phytometabolites on Metal Tolerance of the Pseudo-Metallophyte -Rosmarinus officinalis- in a Mediterranean Brownfield. Chemosphere 2020, 249, 126159. [Google Scholar] [CrossRef] [PubMed]
- Pollard, A.J.; Reeves, R.D.; Baker, A.J.M. Facultative Hyperaccumulation of Heavy Metals and Metalloids. Plant Sci. 2014, 217, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Mendez, M.O.; Maier, R.M. Phytostabilization of Mine Tailings in Arid and Semiarid Environments—An Emerging Remediation Technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, D.; Brereton, N.J.B.; Marchand, L.; Brisson, J.; Pitre, F.E.; Labrecque, M. Complementarity of Three Distinctive Phytoremediation Crops for Multiple-Trace Element Contaminated Soil. Sci. Total Environ. 2018, 610–611, 1428–1438. [Google Scholar] [CrossRef]
- Antoniadis, V.; Levizou, E.; Shaheen, S.M.; Ok, Y.S.; Sebastian, A.; Baum, C.; Prasad, M.N.V.; Wenzel, W.W.; Rinklebe, J. Trace Elements in the Soil-Plant Interface: Phytoavailability, Translocation, and Phytoremediation—A Review. Earth-Sci. Rev. 2017, 171, 621–645. [Google Scholar] [CrossRef]
- Wiszniewska, A.; Hanus-Fajerska, E.; Muszyńska, E.; Ciarkowska, K. Natural Organic Amendments for Improved Phytoremediation of Polluted Soils: A Review of Recent Progress. Pedosphere 2016, 26, 1–12. [Google Scholar] [CrossRef]
- Heckenroth, A.; Rabier, J.; Dutoit, T.; Torre, F.; Prudent, P.; Laffont-Schwob, I. Selection of Native Plants with Phytoremediation Potential for Highly Contaminated Mediterranean Soil Restoration: Tools for a Non-Destructive and Integrative Approach. J. Environ. Manag. 2016, 183, 850–863. [Google Scholar] [CrossRef]
- Navarro-Cano, J.A.; Goberna, M.; Verdú, M. Using Plant Functional Distances to Select Species for Restoration of Mining Sites. J. Appl. Ecol. 2019, 56, 2353–2362. [Google Scholar] [CrossRef]
- Ilunga wa Ilunga, E.; Mahy, G.; Piqueray, J.; Séleck, M.; Shutcha, M.N.; Meerts, P.; Faucon, M.-P. Plant Functional Traits as a Promising Tool for the Ecological Restoration of Degraded Tropical Metal-Rich Habitats and Revegetation of Metal-Rich Bare Soils: A Case Study in Copper Vegetation of Katanga, DRC. Ecol. Eng. 2015, 82, 214–221. [Google Scholar] [CrossRef]
- Salducci, M.-D.; Folzer, H.; Issartel, J.; Rabier, J.; Masotti, V.; Prudent, P.; Affre, L.; Hardion, L.; Tatoni, T.; Laffont-Schwob, I. How Can a Rare Protected Plant Cope with the Metal and Metalloid Soil Pollution Resulting from Past Industrial Activities? Phytometabolites, Antioxidant Activities and Root Symbiosis Involved in the Metal Tolerance of Astragalus tragacantha. Chemosphere 2019, 217, 887–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Affholder, M.-C.; Pricop, A.-D.; Laffont-Schwob, I.; Coulomb, B.; Rabier, J.; Borla, A.; Demelas, C.; Prudent, P. As, Pb, Sb, and Zn Transfer from Soil to Root of Wild Rosemary: Do Native Symbionts Matter? Plant Soil 2014, 382, 219–236. [Google Scholar] [CrossRef]
- Muszyńska, E.; Labudda, M.; Kral, A. Ecotype-Specific Pathways of Reactive Oxygen Species Deactivation in Facultative Metallophyte Silene Vulgaris (Moench) Garcke Treated with Heavy Metals. Antioxidants 2020, 9, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, J.H.; Anacker, B.L.; Strauss, S.Y.; Burke, D.J. Soil Microbial Community Variation Correlates Most Strongly with Plant Species Identity, Followed by Soil Chemistry, Spatial Location and Plant Genus. AoB Plants 2015, 7, plv030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Conesa, H.M.; García, G.; Faz, Á.; Arnaldos, R. Dynamics of Metal Tolerant Plant Communities’ Development in Mine Tailings from the Cartagena-La Unión Mining District (SE Spain) and Their Interest for Further Revegetation Purposes. Chemosphere 2007, 68, 1180–1185. [Google Scholar] [CrossRef] [PubMed]
- Fernández, S.; Poschenrieder, C.; Marcenò, C.; Gallego, J.R.; Jiménez-Gámez, D.; Bueno, A.; Afif, E. Phytoremediation Capability of Native Plant Species Living on Pb-Zn and Hg-As Mining Wastes in the Cantabrian Range, North of Spain. J. Geochem. Explor. 2017, 174, 10–20. [Google Scholar] [CrossRef]
- Higueras, P.; Esbrí, J.M.; García-Ordiales, E.; González-Corrochano, B.; López-Berdonces, M.A.; García-Noguero, E.M.; Alonso-Azcárate, J.; Martínez-Coronado, A. Potentially Harmful Elements in Soils and Holm-Oak Trees (Quercus Ilex L.) Growing in Mining Sites at the Valle de Alcudia Pb-Zn District (Spain)—Some Clues on Plant Metal Uptake. J. Geochem. Explor. 2017, 182, 166–179. [Google Scholar] [CrossRef]
- Pratas, J.; Favas, P.J.C.; D’Souza, R.; Varun, M.; Paul, M.S. Phytoremedial Assessment of Flora Tolerant to Heavy Metals in the Contaminated Soils of an Abandoned Pb Mine in Central Portugal. Chemosphere 2013, 90, 2216–2225. [Google Scholar] [CrossRef] [PubMed]
- Escarré, J.; Lefèbvre, C.; Raboyeau, S.; Dossantos, A.; Gruber, W.; Cleyet Marel, J.C.; Frérot, H.; Noret, N.; Mahieu, S.; Collin, C.; et al. Heavy Metal Concentration Survey in Soils and Plants of the Les Malines Mining District (Southern France): Implications for Soil Restoration. Water Air Soil Pollut. 2011, 216, 485–504. [Google Scholar] [CrossRef] [Green Version]
- Szarek-Łukaszewska, G. Vegetation of Reclaimed and Spontaneously Vegetated Zn-Pb Mine Wastes in Southern Poland. Pol. J. Environ. Stud. 2009, 18, 717–733. [Google Scholar]
- Pollock, L.J.; Morris, W.K.; Vesk, P.A. The Role of Functional Traits in Species Distributions Revealed through a Hierarchical Model. Ecography 2012, 35, 716–725. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Plant-Soil Interactions in Mediterranean Forest and Shrublands: Impacts of Climatic Change. Plant Soil 2013, 365, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierret, A.; Maeght, J.-L.; Clément, C.; Montoroi, J.-P.; Hartmann, C.; Gonkhamdee, S. Understanding Deep Roots and Their Functions in Ecosystems: An Advocacy for More Unconventional Research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azul, A.M.; Ramos, V.; Pato, A.; Azenha, M.; Freitas, H. Mycorrhizal Types in the Mediterranean Basin: Safety Teaching and Training. J. Biol. Educ. 2008, 42, 130–137. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.-L. Phylogenetic Distribution and Evolution of Mycorrhizas in Land Plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Pawlowska, T.E.; Błaszkowski, J.; Rühling, Å. The Mycorrhizal Status of Plants Colonizing a Calamine Spoil Mound in Southern Poland. Mycorrhiza 1997, 6, 499–505. [Google Scholar] [CrossRef]
- Regvar, M.; Vogel, K.; Irgel, N.; Wraber, T.; Hildebrandt, U.; Wilde, P.; Bothe, H. Colonization of Pennycresses (Thlaspi Spp.) of the Brassicaceae by Arbuscular Mycorrhizal Fungi. J. Plant Physiol. 2003, 160, 615–626. [Google Scholar] [CrossRef] [Green Version]
- Roumet, C.; Lafont, F.; Sari, M.; Warembourg, F.; Garnier, E. Root Traits and Taxonomic Affiliation of Nine Herbaceous Species Grown in Glasshouse Conditions. Plant Soil 2008, 312, 69–83. [Google Scholar] [CrossRef]
- Rajkumar, M.; Sandhya, S.; Prasad, M.N.V.; Freitas, H. Perspectives of Plant-Associated Microbes in Heavy Metal Phytoremediation. Biotechnol. Adv. 2012, 30, 1562–1574. [Google Scholar] [CrossRef]
- Dietterich, L.H.; Gonneau, C.; Casper, B.B. Arbuscular Mycorrhizal Colonization Has Little Consequence for Plant Heavy Metal Uptake in Contaminated Field Soils. Ecol. Appl. 2017, 27, 1862–1875. [Google Scholar] [CrossRef] [PubMed]
- Wang, F. Occurrence of Arbuscular Mycorrhizal Fungi in Mining-Impacted Sites and Their Contribution to Ecological Restoration: Mechanisms and Applications. Crit. Rev. Environ. Sci. Technol. 2017, 47, 1901–1957. [Google Scholar] [CrossRef]
- Nadgórska-Socha, A.; Kandziora-Ciupa, M.; Ciepał, R. Element Accumulation, Distribution, and Phytoremediation Potential in Selected Metallophytes Growing in a Contaminated Area. Environ. Monit. Assess. 2015, 187, 441. [Google Scholar] [CrossRef] [PubMed]
- Bothe, H. Plants in Heavy Metal Soils. In Detoxification of Heavy Metals; Sherameti, I., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; Volume 30, pp. 35–57. [Google Scholar]
- Marsili, S.; Roccotiello, E.; Carbone, C.; Marescotti, P.; Cornara, L.; Mariotti, M.G. Plant Colonization on a Contaminated Serpentine Site. Northeast. Nat. 2009, 16, 297–308. [Google Scholar] [CrossRef]
- Szarek-Łukaszewska, G.; Grodzińska, K. Grasslands of A Zn-Pb Post-Mining Area (Olkusz Ore-Bearing Region, S Poland). Pol. Bot. J. 2011, 56, 245–260. [Google Scholar]
- Bergmeier, E.; Konstantinou, M.; Tsiripidis, I.; Sýkora, K.V. Plant Communities on Metalliferous Soils in Northern Greece. Phytocoenologia 2009, 39, 411–438. [Google Scholar] [CrossRef]
- Woch, M.W. Species Trait-Environment Relationships in Semi-Dry Brachypodium pinnatum Grasslands on Old Waste Heaps Left by Zn-Pb Mining in the Western Małopolska Region (S Poland). Tuexenia 2017, 37, 247–270. [Google Scholar]
- Turnau, K.; Gawroński, S.; Ryszka, P.; Zook, D. Mycorrhizal-Based Phytostabilization of Zn–Pb Tailings: Lessons from the Trzebionka Mining Works (Southern Poland). In Bio-Geo Interactions in Metal-Contaminated Soils; Kothe, E., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 31, pp. 327–348. [Google Scholar]
- Hazard, C.; Johnson, D. Does Genotypic and Species Diversity of Mycorrhizal Plants and Fungi Affect Ecosystem Function? New Phytol. 2018, 220, 1122–1128. [Google Scholar] [CrossRef] [Green Version]
- Partida-Martínez, L.P.; Heil, M. The Microbe-Free Plant: Fact or Artifact? Front. Plant Sci. 2011, 2. [Google Scholar] [CrossRef] [Green Version]
- Raunkiær, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- Lombardo, E.; Bancheva, S.; Domina, G.; Venturella, G. Distribution, Ecological Role and Symbioses of Selected Shrubby Species in the Mediterranean Basin: A Review. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2020, 154, 1–31. [Google Scholar] [CrossRef]
- Bonis, A.; Grubb, P.J.; Coomes, A. Requirements of Gap-Demanding Species in Chalk Grassland: Reduction of Root Competition versus Nutrient Enrichment by Animals. J. Ecol. 1997, 85, 625–633. [Google Scholar] [CrossRef]
- Li, H.-Y.; Li, D.-W.; He, C.-M.; Zhou, Z.-P.; Mei, T.; Xu, H.-M. Diversity and Heavy Metal Tolerance of Endophytic Fungi from Six Dominant Plant Species in a Pb-Zn Mine Wasteland in China. Fungal Ecol. 2012, 5, 309–315. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, T.; Zhao, Z.-W. Colonization Characteristics and Composition of Dark Septate Endophytes (DSE) in a Lead and Zinc Slag Heap in Southwest China. Soil Sediment Contam. Int. J. 2013, 22, 532–545. [Google Scholar] [CrossRef]
- Oba, H.; Tawaray, K.; Wagatsuma, T. Arbuscular Mycorrhizal Colonization in Lupinus and Related Genera. Soil Sci. Plant Nutr. 2001, 47, 685–694. [Google Scholar] [CrossRef]
- Baumel, A.; Auda, P.; Torre, F.; Medail, F. Morphological Polymorphism and RDNA Internal Transcribed Spacer (ITS) Sequence Variation in Armeria (Plumbaginaceae) from South-Eastern France. Bot. J. Linn. Soc. 2009, 159, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Kachenko, A.G. General Introduction & Background. In Ecophysiology and Phytoremediation Potential of Heavy Metal (Loid) Accumulating Plants; The University of Sydney: New South Wales, Australia, 2008; pp. 1–52. [Google Scholar]
- Babst-Kostecka, A.A.; Waldmann, P.; Frérot, H.; Vollenweider, P. Plant Adaptation to Metal Polluted Environments—Physiological, Morphological, and Evolutionary Insights from Biscutella laevigata. Environ. Exp. Bot. 2016, 127, 1–13. [Google Scholar] [CrossRef]
- Pastor, J.; Hernández, A.J. Multi-Functional Role of Grassland Systems in the Ecological Restoration of Mines, Landfills, Roadside Slopes and Agroecosystems. Options Mediterr. 2008, 79, 6. [Google Scholar]
- Veselkin, D.V.; Konoplenko, M.A.; Betekhtina, A.A. Means for Soil Nutrient Uptake in Sedges with Different Ecological Strategies. Russ. J. Ecol. 2014, 45, 547–554. [Google Scholar] [CrossRef]
- Tsiripidis, I.; Papaioannou, A.; Sapounidis, V.; Bergmeier, E. Approaching the Serpentine Factor at a Local Scale—A Study in an Ultramafic Area in Northern Greece. Plant Soil 2010, 329, 35–50. [Google Scholar] [CrossRef] [Green Version]
- Meerts, P.; Grommesch, C. Soil Seed Banks in a Heavy-Metal Polluted Grassland at Prayon (Belgium). Plant Ecol. 2001, 155, 35–45. [Google Scholar] [CrossRef]
- Colombo, R.P.; Martínez, A.E.; Pardo, A.F.; Di Bidondo, L.F.; Van Baren, C.; Lira, P.d.L.; Godeas, A.M. Differential Effects of Two Strains of Rhizophagus intraradices on Dry Biomass and Essential Oil Yield and Composition in Calamintha nepeta. Rev. Argent. Microbiol. 2013, 45, 114–118. [Google Scholar] [CrossRef] [Green Version]
- Hüseyinova, R.; Kutbay, H.G.; Bilgin, A.; Kılıç, D.; Horuz, A.; Kırmanoğlu, C. Sulphur and Some Heavy Metal Contents in Foliage of Corylus avellana and Some Roadside Native Plants in Ordu Province, Turkey. Ekoloji 2009, 18, 10–16. [Google Scholar] [CrossRef]
- Shallari, S.; Schwartza, C.; Haskob, A.; MorelaT, J.L. Heavy Metals in Soils and Plants of Serpentine and Industrial Sites of Albania. Sci. Total Environ. 1998, 10. [Google Scholar] [CrossRef]
- Turnau, K. Heavy Metal Content and Localization in Mycorrhizal Euphorbia cyparissias from Zinc Wastes in Southern Poland. Acta Soc. Bot. Pol. 1998, 67, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.; Brinkmann, K. Heavy Metal Tolerance in Festuca ovina L. from Contaminated Sites in the Eifel Mountains, Germany. Plant Soil 1992, 143, 239–247. [Google Scholar] [CrossRef]
- Nworie, O.; Qin, J.; Lin, C. Trace Element Uptake by Herbaceous Plants from the Soils at a Multiple Trace Element-Contaminated Site. Toxics 2019, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boularbah, A.; Schwartz, C.; Bitton, G.; Aboudrar, W.; Ouhammou, A.; Morel, J.L. Heavy Metal Contamination from Mining Sites in South Morocco: 2. Assessment of Metal Accumulation and Toxicity in Plants. Chemosphere 2006, 63, 811–817. [Google Scholar] [CrossRef]
- Glavač, N.K.; Djogo, S.; Ražić, S.; Kreft, S.; Veber, M. Accumulation of Heavy Metals from Soil in Medicinal Plants. Arch. Ind. Hyg. Toxicol. 2017, 68, 236–244. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, D.; Kula, I.; Ay, G.; Baslar, S.; Dogan, Y. Determination of Trace Elements in the Plants of Mt. Bozdag, Izmir, Turkey. Arch. Biol. Sci. 2010, 62, 731–738. [Google Scholar] [CrossRef]
- Gucwa-Przepióra, E.; Chmura, D.; Sokołowska, K. AM and DSE Colonization of Invasive Plants in Urban Habitat: A Study of Upper Silesia (Southern Poland). J. Plant Res. 2016, 129, 603–614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suchkova, N.; Tsiripidis, I.; Alifragkis, D.; Ganoulis, J.; Darakas, E.; Sawidis, T. Assessment of Phytoremediation Potential of Native Plants during the Reclamation of an Area Affected by Sewage Sludge. Ecol. Eng. 2014, 69, 160–169. [Google Scholar] [CrossRef]
- Hernández, A.J.; Pastor, J. Incidencia De Metales Pesados En La Biodiversidad De Pastizales En El Emplazamiento De Una Explotación Abandonada De Baritina. In Los Sistemas Forrajeros: Entre La Producción Y El Paisaje; Reunión Científica XLVI, SEEP: Vitoria-Gasteiz, Spain, 2007; p. 8. [Google Scholar]
- Friede, M.; Unger, S.; Hellmann, C.; Beyschlag, W. Conditions Promoting Mycorrhizal Parasitism Are of Minor Importance for Competitive Interactions in Two Differentially Mycotrophic Species. Front. Plant Sci. 2016, 7, 1465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banásová, V.; Ďurišová, E.; Nadubinská, M.; Gurinová, E.; Čiamporová, M. Natural Vegetation, Metal Accumulation and Tolerance in Plants Growing on Heavy Metal Rich Soils. In Bio-Geo Interactions in Metal-Contaminated Soils; Kothe, E., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 31, pp. 233–250. [Google Scholar]
- Pająk, M.; Halecki, W.; Gąsiorek, M. Accumulative Response of Scots Pine (Pinus sylvestris L.) and Silver Birch (Betula pendula Roth) to Heavy Metals Enhanced by Pb-Zn Ore Mining and Processing Plants: Explicitly Spatial Considerations of Ordinary Kriging Based on a GIS Approach. Chemosphere 2017, 168, 851–859. [Google Scholar]
- Anawar, H.M.; Canha, N.; Santa-Regina, I.; Freitas, M.C. Adaptation, Tolerance, and Evolution of Plant Species in a Pyrite Mine in Response to Contamination Level and Properties of Mine Tailings: Sustainable Rehabilitation. J. Soils Sediments 2013, 13, 730–741. [Google Scholar] [CrossRef]
- Pedron, F.; Petruzzelli, G.; Barbafieri, M.; Tassi, E. Strategies to Use Phytoextraction in Very Acidic Soil Contaminated by Heavy Metals. Chemosphere 2009, 75, 808–814. [Google Scholar] [CrossRef]
- Newman, E.I.; Eason, W.R.; Eissenstat, D.M.; Ramos, M.I.R.F. Interactions between Plants: The Role of Mycorrhizae. Mycorrhiza 1992, 1, 47–53. [Google Scholar] [CrossRef]
- Kalinovic, J.V.; Serbula, S.M.; Radojevic, A.A.; Milosavljevic, J.S.; Kalinovic, T.S.; Steharnik, M.M. Assessment of As, Cd, Cu, Fe, Pb, and Zn Concentrations in Soil and Parts of Rosa spp. Sampled in Extremely Polluted Environment. Environ. Monit. Assess. 2019, 191, 15. [Google Scholar] [CrossRef]
- Zekri, J.; Rached, O.; Sahli, L.; Yilmaz, A.; Temel, H.; Tahar, A. Assessment of Soil Contamination and Plant Stress Tolerance in an Antimony Mining Area: Case Study for Scabiosa atropurpurea L. and Santolina chamaecyparissus. Environ. Ecol. 2019, 37, 747–757. [Google Scholar]
- Reeves, R.D.; Adigüzel, N.; Baker, A.J.M. Nickel Hyperaccumulation in Bornmuellera kiyakii Aytaç & Aksoy and Associated Plants of the Brassicaceae from Kızıldağ (Derebucak, Konya-Turkey). Turk. J. Bot. 2009, 33, 33–40. [Google Scholar]
- Barni, E.; Siniscalco, C. Vegetation Dynamics and Arbuscular Mycorrhiza in Old-Field Successions of the Western Italian Alps. Mycorrhiza 2000, 10, 63–72. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Llugany, M.; Lombini, A.; Dinelli, E.; Bech, J.; Barceló, J. Smilax aspera L. an Evergreen Mediterranean Climber for Phytoremediation. J. Geochem. Explor. 2012, 123, 41–44. [Google Scholar] [CrossRef]







| Element (mg/kg) | ||||||
|---|---|---|---|---|---|---|
| Cr | Cu | Fe | Mn | Ni | Pb | Zn |
| 103 ± 38 | 39 ± 17 | 173,377 ± 58,177 | 3000 ± 936 | 89 ± 30 | 17,230 ± 6804 | 117,321 ± 33,770 |
| Latin Name | Botanical Family | Occurrence in Each Relevé * | Frequency | |||||
|---|---|---|---|---|---|---|---|---|
| (1) | (2) | (3) | (4) | (5) | (6) | - | ||
| Amelanchier ovalis Medik. | Rosaceae | - | - | - | - | - | X | 1/6 |
| Aphyllantes monspeliensis L. | Liliaceae | - | X | - | - | X | - | 2/6 |
| Arenaria serpyllifolia L. | Caryophyllaceae | - | - | X | X | X | X | 4/6 |
| Argyrolobium zanonii (Turra) P.W.Ball | Fabaceae | - | X | - | X | - | X | 3/6 |
| Armeria arenaria subsp bupleuroides (Godr. & Gren.) Greuter & Burdet | Plumbaginaceae | X | X | X | X | - | - | 4/6 |
| Asparagus acutifolius L. | Asparagaceae | - | - | - | X | - | - | 1/6 |
| Asplenium ruta-muraria L. | Aspleniaceae | - | X | - | - | - | - | 1/6 |
| Biscutella laevigata L. | Brassicaceae | X | X | X | X | X | X | 6/6 |
| Brachypodium phoenicoides (L.) Roem. & Schult | Poaceae | - | - | - | - | - | X | 1/6 |
| Brachypodium retusum (Pers.) P.Beauv. | Poaceae | - | - | - | - | - | X | 1/6 |
| Bromus madritensis L. | Poaceae | - | - | - | X | - | - | 1/6 |
| Buxus sempervirens L. | Buxaceae | - | X | X | X | - | X | 4/6 |
| Carex halleriana Asso | Cyperaceae | - | - | - | - | X | - | 1/6 |
| Centaurea pectinata L. | Asteraceae | X | X | X | X | - | - | 4/6 |
| Cerastium pumilum Curtis | Caryophyllaceae | - | X | X | - | - | X | 3/6 |
| Clematis vitalba L. | Ranunculaceae | - | - | - | - | - | X | 1/6 |
| Clinopodium nepeta (L.) Kuntze | Lamiaceae | - | - | - | - | X | X | 1/6 |
| Dactylis glomerata L. | Poaceae | - | - | - | - | X | X | 2/6 |
| Dioscorea communis (L.) Caddick & Wilkin | Dioscoreaceae | - | - | - | - | - | X | 1/6 |
| Draba verna L. | Brassicaceae | - | X | X | - | - | - | 2/6 |
| Eryngium campestre L. | Apiaceae | - | - | - | X | - | - | 1/6 |
| Euphorbia cyparissias L. | Euphorbiacee | - | X | X | X | - | - | 3/6 |
| Festuca ovina sl L. | Poaceae | X | X | X | - | X | X | 5/6 |
| Galium aparine L. | Rubiaceae | - | X | X | X | X | - | 4/6 |
| Galium corrudifolium Vill. | Rubiaceae | - | - | - | - | X | X | 2/6 |
| Helleborus foetidus L. | Ranunculaceae | - | - | - | - | - | X | 1/6 |
| Hordeum murinum L. | Poaceae | - | - | - | X | - | - | 1/6 |
| Hornungia petraea (L.) ex Rchb. | Brassicaceae | - | X | X | - | - | - | 2/6 |
| Hypericum perforatum L. | Hypericaceae | - | X | - | - | - | - | 1/6 |
| Juniperus oxycedrus L. | Cupressaceae | - | - | - | X | - | X | 2/6 |
| Lactuca perennis L. | Asteraceae | - | - | - | - | - | X | 1/6 |
| Lepidium draba L. | Brassicaceae | - | - | - | X | - | - | 1/6 |
| Lysimachia arvensis (L.) U. Manns & Anderb. | Primulaceae | - | - | - | X | X | - | 2/6 |
| Mibora minima (L.) Desv. | Poaceae | X | X | X | X | - | - | 4/6 |
| Noccaea caerulescens (J.Presl & C.Presl) F.K.Mey | Brassicaceae | X | X | X | X | X | - | 5/6 |
| Pilosella officinarum Vaill. | Asteraceae | - | X | X | X | - | - | 3/6 |
| Pinus sylvestris L. | Pinaceae | - | X | X | - | - | X | 3/6 |
| Pistacia terebinthus L. | Anacardiaceae | - | - | - | - | - | X | 1/6 |
| Plantago lanceolata L. | Plantaginaceae | - | - | - | - | - | X | 1/6 |
| Poa annua L. | Poaceae | - | - | - | X | - | - | 1/6 |
| Poterium sanguisorba L. | Rosaceae | - | - | - | - | - | X | 1/6 |
| Pyrus spinosa Forssk. | Rosaceae | - | - | - | - | - | X | 1/6 |
| Quercus ilex L. | Fagaceae | - | X | X | X | X | X | 5/6 |
| Quercus pubescens Willd. | Fagaceae | - | - | - | - | X | X | 2/6 |
| Ranunculus bulbosus L. | Ranunculaceae | - | - | - | - | X | X | 2/6 |
| Reseda lutea L. | Resedaceae | X | X | X | X | - | - | 4/6 |
| Rosa canina L. | Rosaceae | - | - | - | - | - | X | 1/6 |
| Rubia peregrina L. | Rubiaceae | - | - | - | X | - | X | 2/6 |
| Rubus ulmifolius Schott | Rosaceae | - | - | - | X | - | X | 2/6 |
| Rumex intermedius D.C. | Polygonaceae | - | X | X | - | - | - | 2/6 |
| Ruscus aculeatus L. | Asparagaceae | - | - | - | - | X | X | 2/6 |
| Scabiosa atropurpurea L. | Caprifoliaceae | X | X | X | - | - | - | 3/6 |
| Scrophularia lucida L. | Scrophulariaceae | - | X | X | - | - | - | 2/6 |
| Sedum acre L. | Crassulaceae | - | - | - | X | - | - | 1/6 |
| Sedum annuum L. | Crassulaceae | - | X | X | - | - | - | 2/6 |
| Senecio vulgaris L. | Asteraceae | - | - | - | X | - | - | 1/6 |
| Silene vulgaris (Moench) Garcke | Caryophyllaceae | - | - | - | X | - | - | 1/6 |
| Smilax aspera L. | Smilacaceae | - | X | - | X | - | X | 3/6 |
| Thymus vulgaris L. | Lamiaceae | X | X | X | X | X | X | 6/6 |
| Trifolium pratense L. | Fabaceae | - | - | - | - | X | X | 2/6 |
| Ulex parviflorus Pourr. | Fabaceae | - | X | - | X | - | - | 2/6 |
| Element (mg/kg) | Plant Species | |||
|---|---|---|---|---|
| A. arenaria | B. laevigata | N. caerulescens | P. lanceolata | |
| In aerial parts | ||||
| Cr | 9.6 ± 4.3 | 10.7 ± 5.3 | 11.4 ± 4.2 | 11.4 ± 4.2 |
| Cu | 4.6 ± 0.3 | 3.9 ± 0.5 | 5.0 ± 1.3 | 7.3 ± 0.4 |
| Fe | 966.3 ± 375.4 | 325.6 ± 186.2 | 5780 ± 1878 | 728.5 ± 167.1 |
| Mn | 160.3 ± 99.1 | 204.9 ± 80.2 | 105.3 ± 25.0 | 36.6 ± 4.0 |
| Ni | 1.8 ± 0.2 ab | 1.8 ± 0.3 ab | 3.6 ± 0.8 a | 0.5 ± 0.2 b |
| Pb | 74.9 ± 6.8 ab | 48.2 ± 6.3 b | 496.2 ± 104.6 a | 69.2 ± 15.1 ab |
| Zn | 1,916 ± 201 ab | 1,775 ± 125 ab | 10,664 ± 563 a | 946 ± 88 b |
| In root parts | ||||
| Cr | 4.3 ± 1.9 | 2.5 ± 0.8 | 29.9 ± 9.1 | 10.1 ± 0.5 |
| Cu | 8.6 ± 1.7 | 6.9 ± 0.1 | 27.2 ± 6.7 | 21.3 ± 1.2 |
| Fe | 7,719 ± 3,565 | 5,472 ± 1,009 | 56,779 ± 24,995 | 15,210 ± 1,035 |
| Mn | 303.0 ± 89.7 | 223.2 ± 46.3 | 1,143.2 ± 427.7 | 405.7 ± 37.9 |
| Ni | 5.2 ± 2.5 | 5.9 ± 2.5 | 30.6 ± 13.7 | 7.8 ± 0.6 |
| Pb | 1,769 ± 49 | 1,393 ± 129 | 5,123 ± 1,879 | 1,630 ± 124 |
| Zn | 10,659 ± 2,763 | 6,902 ± 547 | 66,381 ± 24,010 | 12,217 ± 996 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laffont-Schwob, I.; Rabier, J.; Masotti, V.; Folzer, H.; Tosini, L.; Vassalo, L.; Salducci, M.-D.; Prudent, P. Functional Trait-Based Screening of Zn-Pb Tolerant Wild Plant Species at an Abandoned Mine Site in Gard (France) for Rehabilitation of Mediterranean Metal-Contaminated Soils. Int. J. Environ. Res. Public Health 2020, 17, 5506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17155506
Laffont-Schwob I, Rabier J, Masotti V, Folzer H, Tosini L, Vassalo L, Salducci M-D, Prudent P. Functional Trait-Based Screening of Zn-Pb Tolerant Wild Plant Species at an Abandoned Mine Site in Gard (France) for Rehabilitation of Mediterranean Metal-Contaminated Soils. International Journal of Environmental Research and Public Health. 2020; 17(15):5506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17155506
Chicago/Turabian StyleLaffont-Schwob, Isabelle, Jacques Rabier, Véronique Masotti, Hélène Folzer, Lorène Tosini, Laurent Vassalo, Marie-Dominique Salducci, and Pascale Prudent. 2020. "Functional Trait-Based Screening of Zn-Pb Tolerant Wild Plant Species at an Abandoned Mine Site in Gard (France) for Rehabilitation of Mediterranean Metal-Contaminated Soils" International Journal of Environmental Research and Public Health 17, no. 15: 5506. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph17155506