
Vegetation Dynamic Assessment by NDVI and Field Observations for Sustainability of China’s Wulagai River Basin



 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
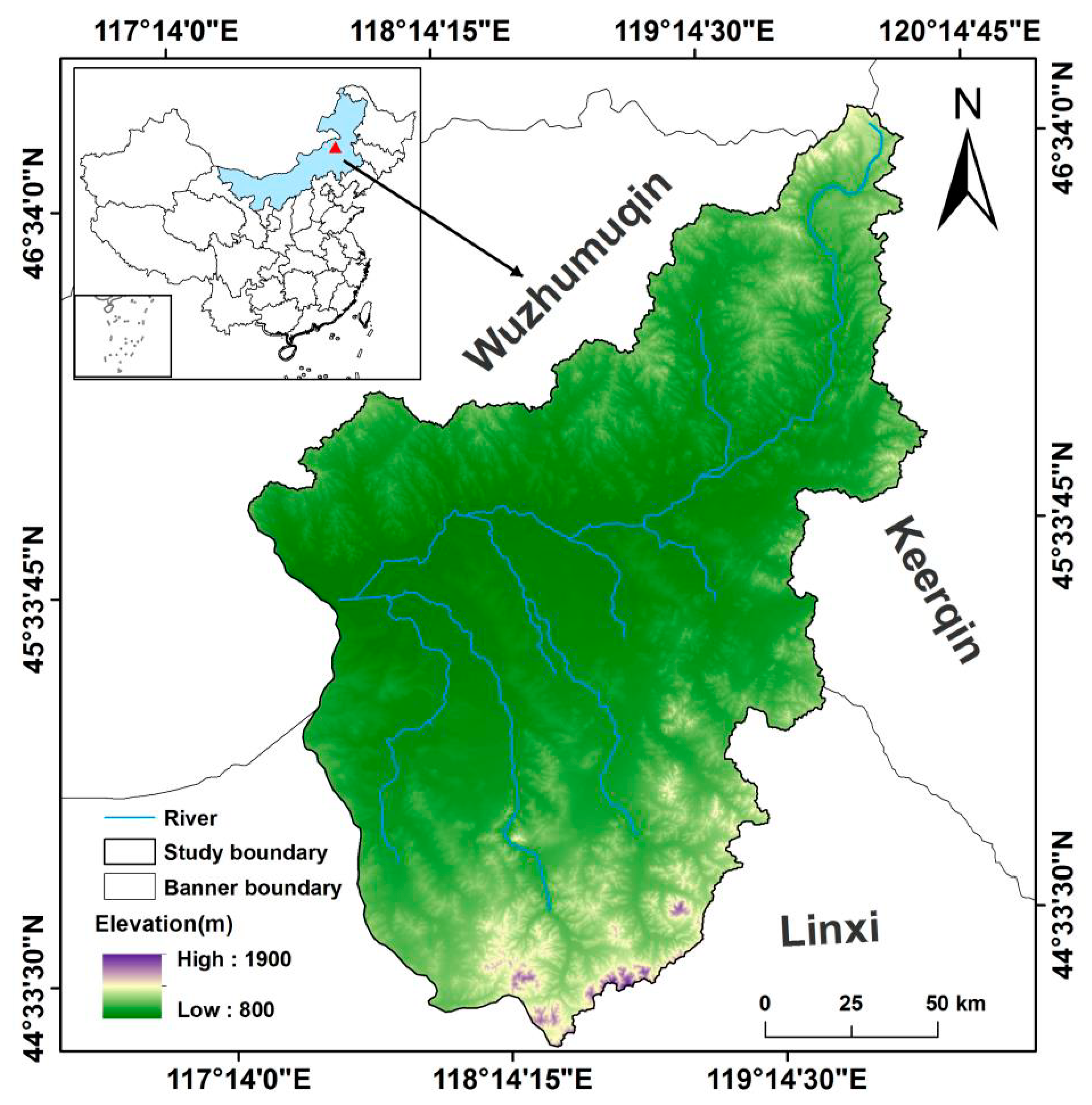
2.1. Study Area
2.2. Data
2.2.1. Normalized Difference Vegetation Index (NDVI) Data Sets
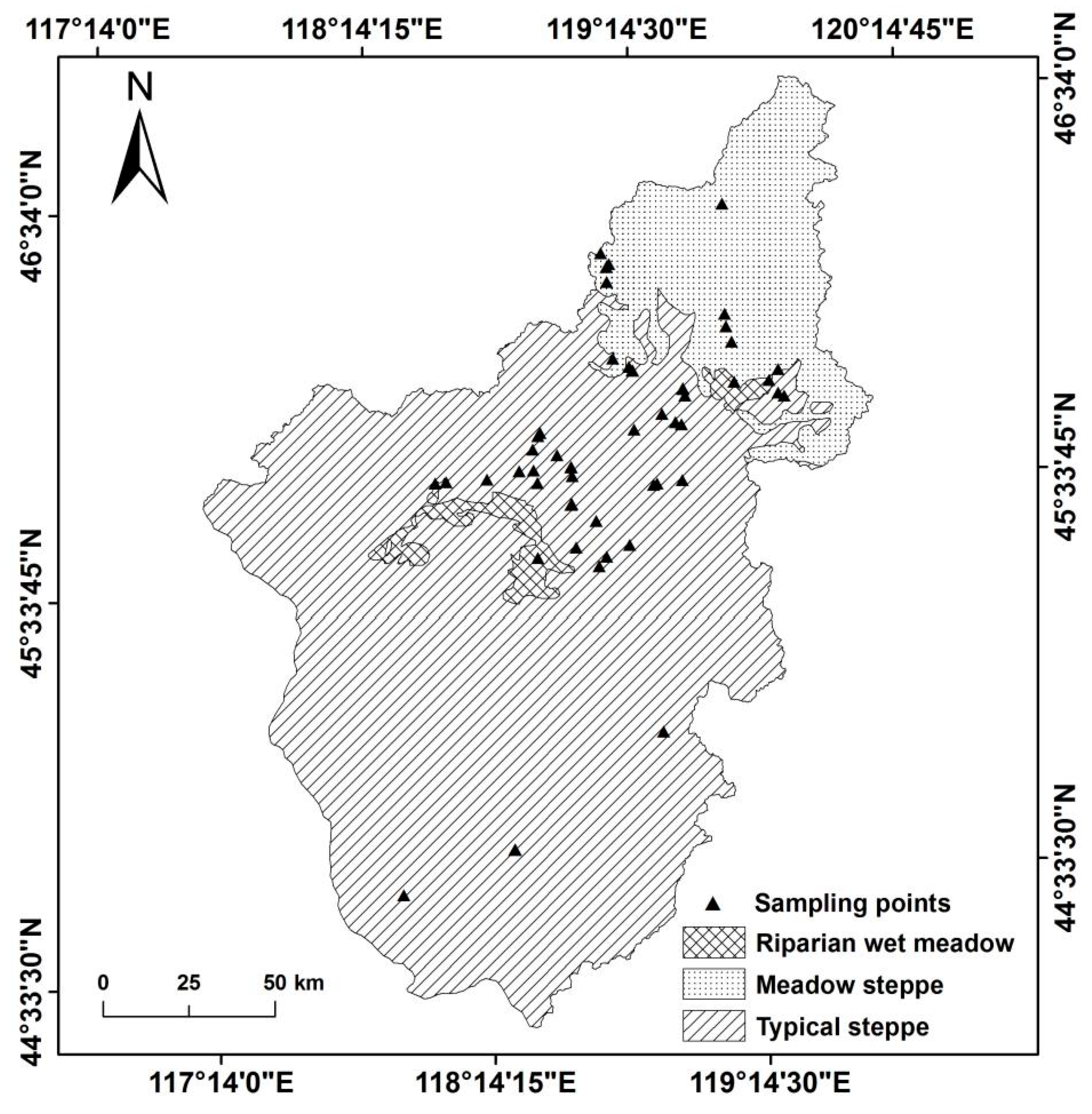
2.2.2. Plant Observations
2.2.3. Meteorological Data
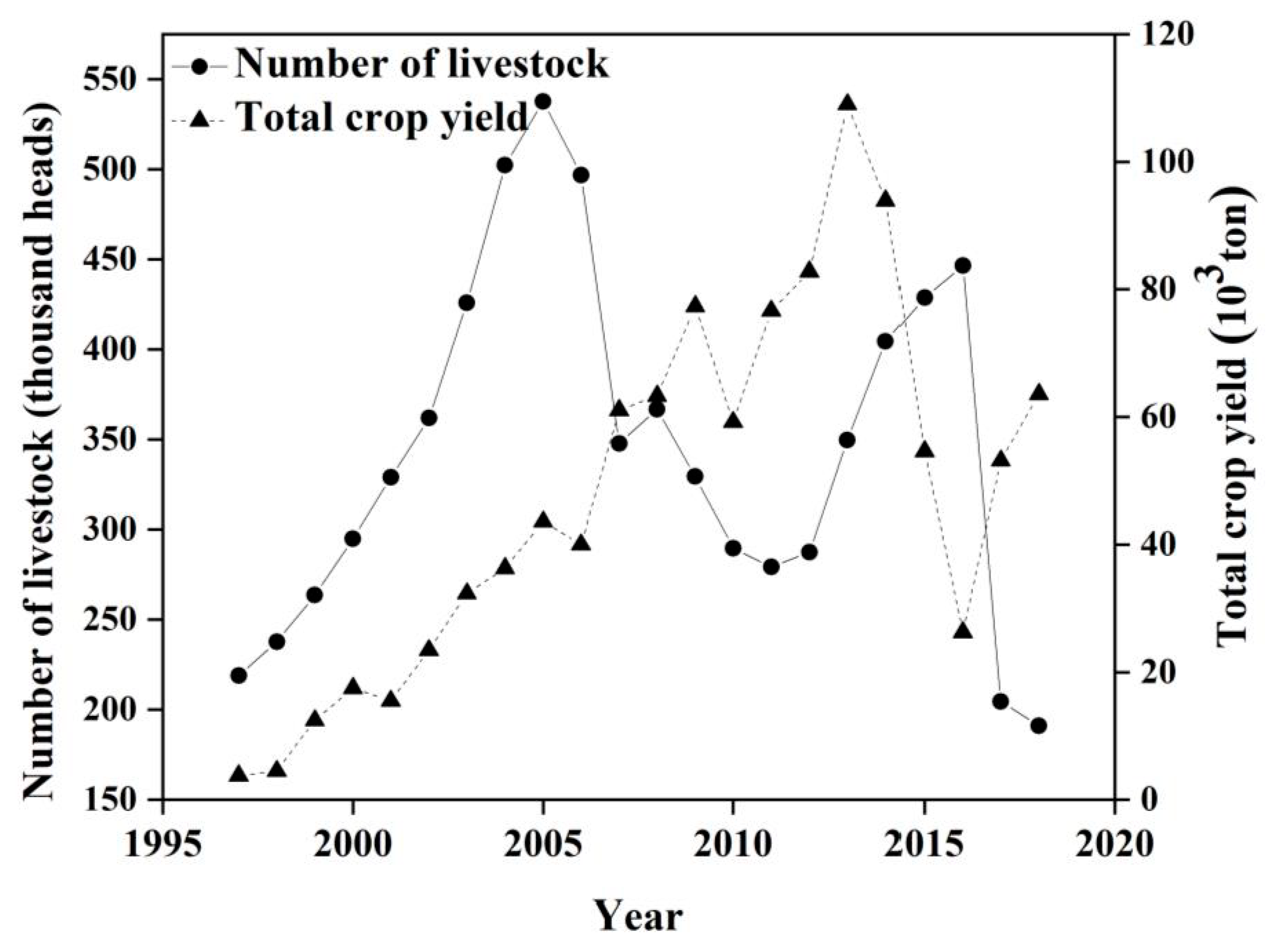
2.2.4. Livestock and Crop Yield Statistics
2.3. Methods
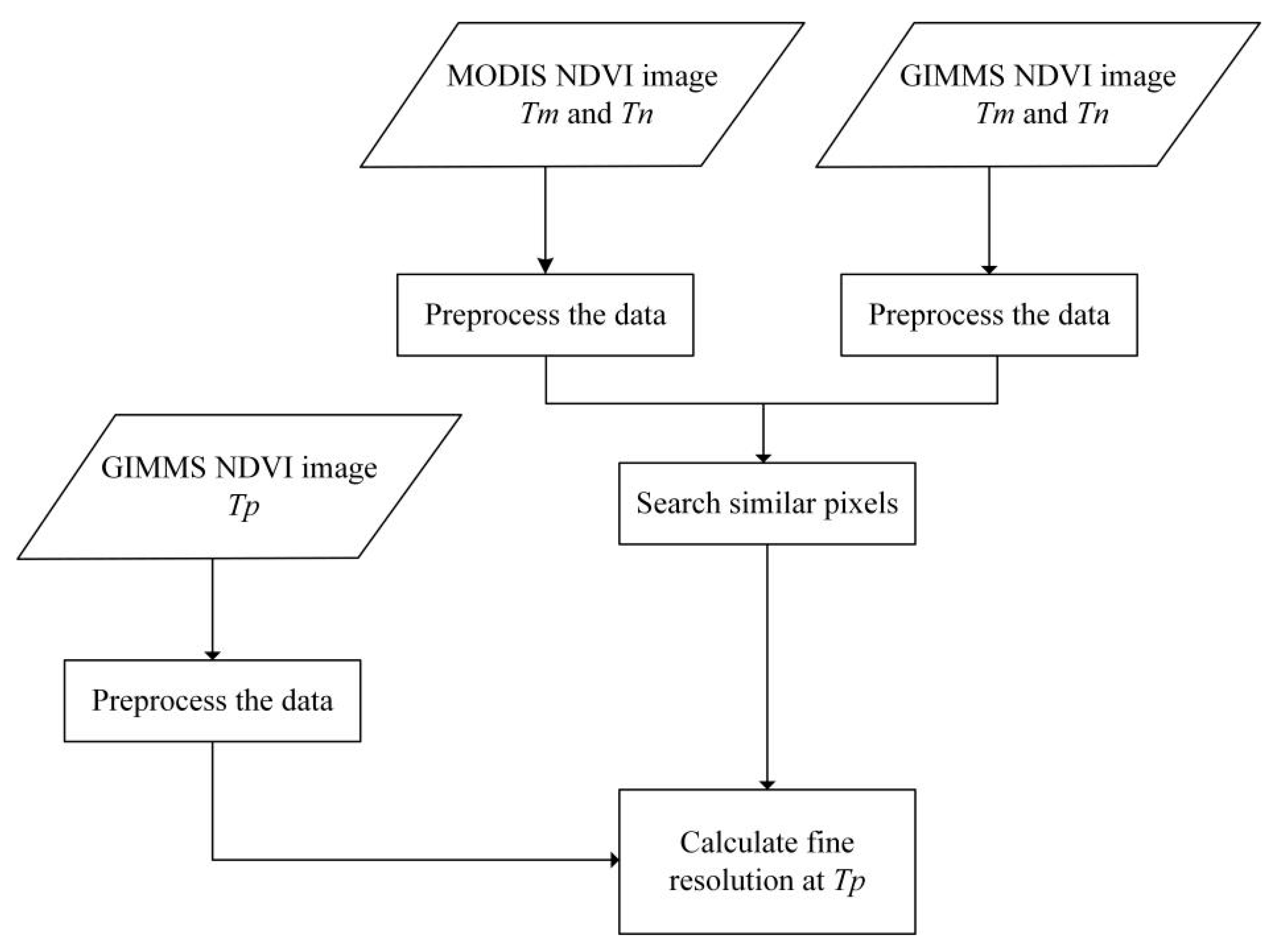
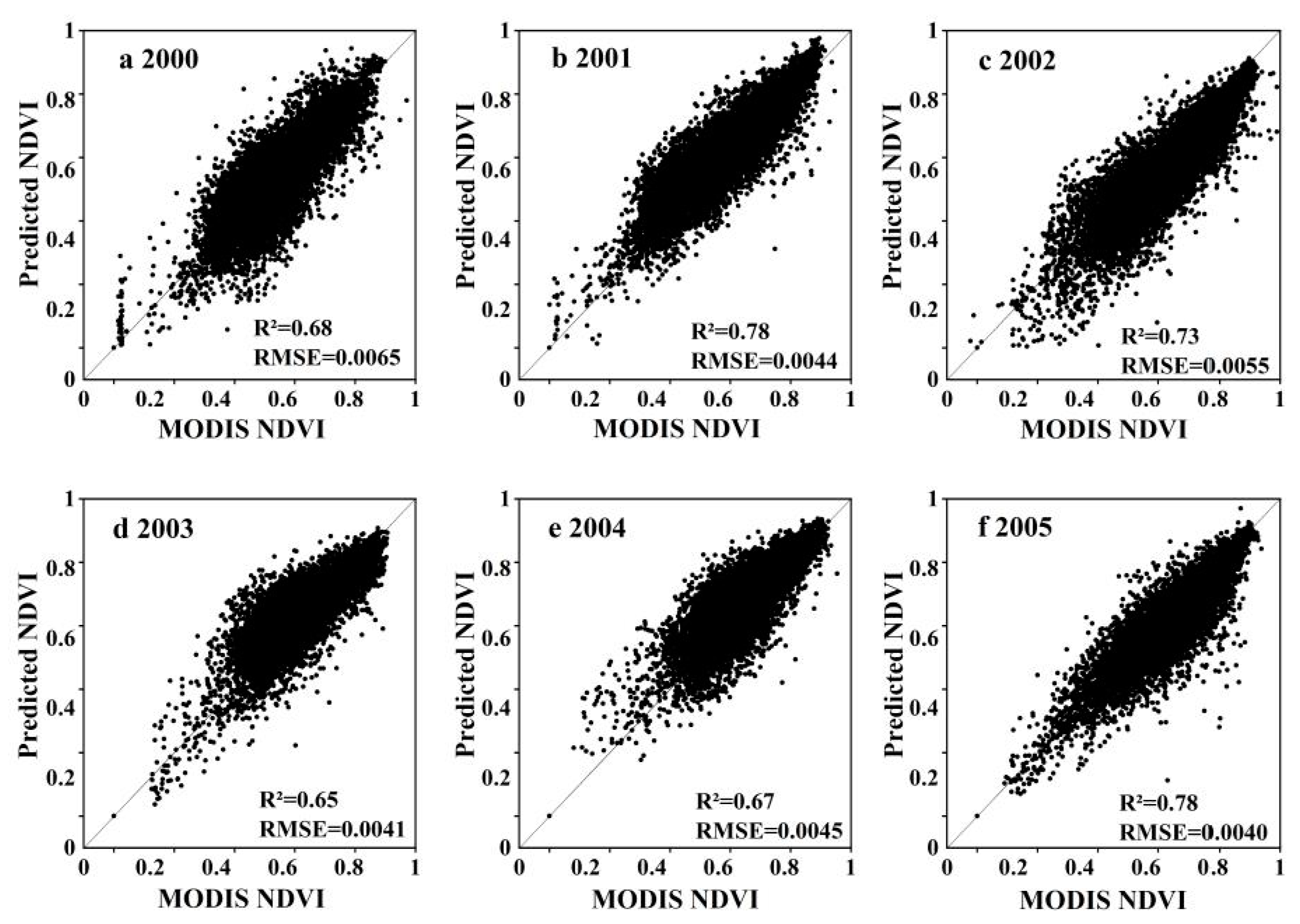
2.3.1. Building NDVI Data Sets
2.3.2. The RESTREND Method
3. Results
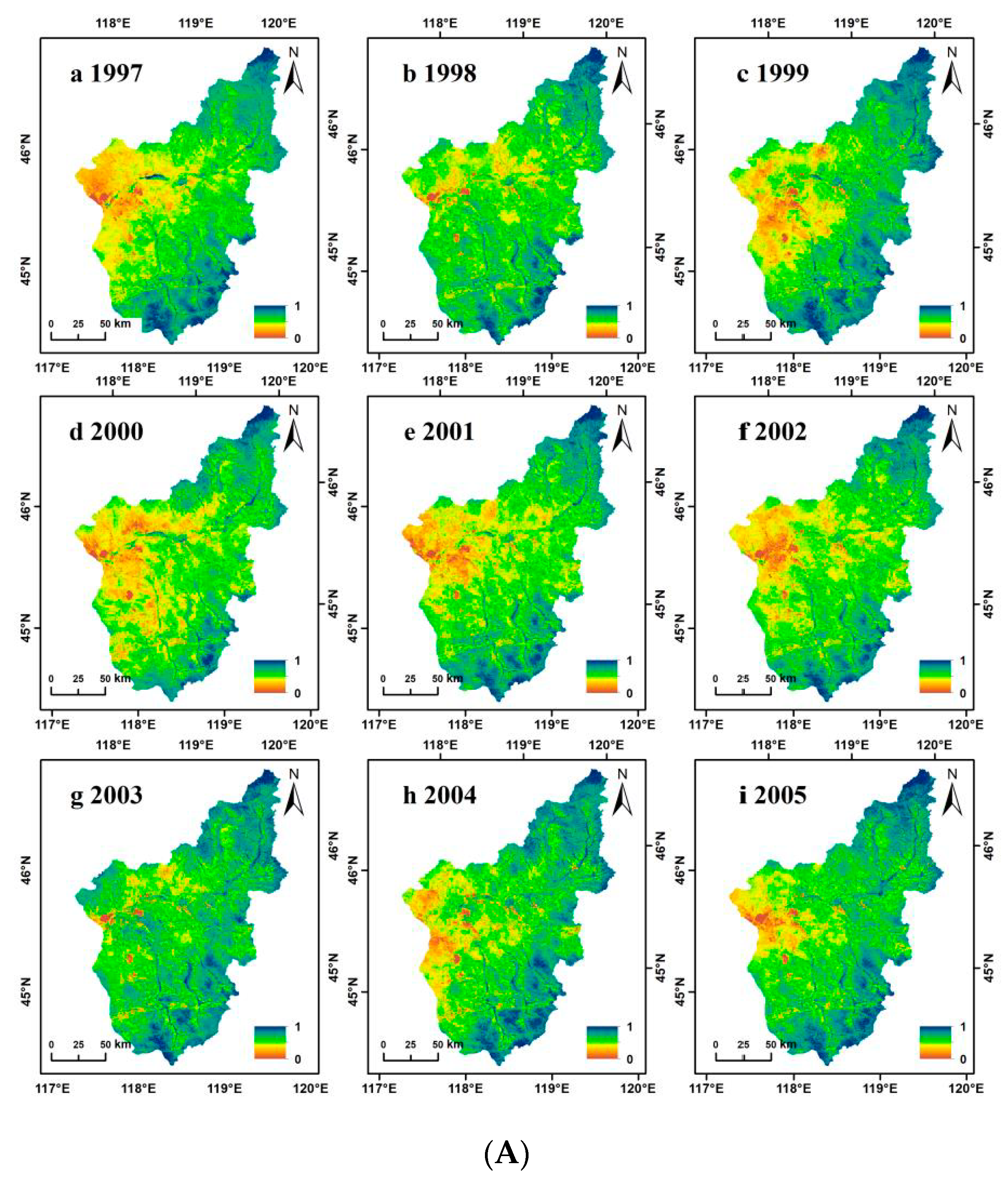
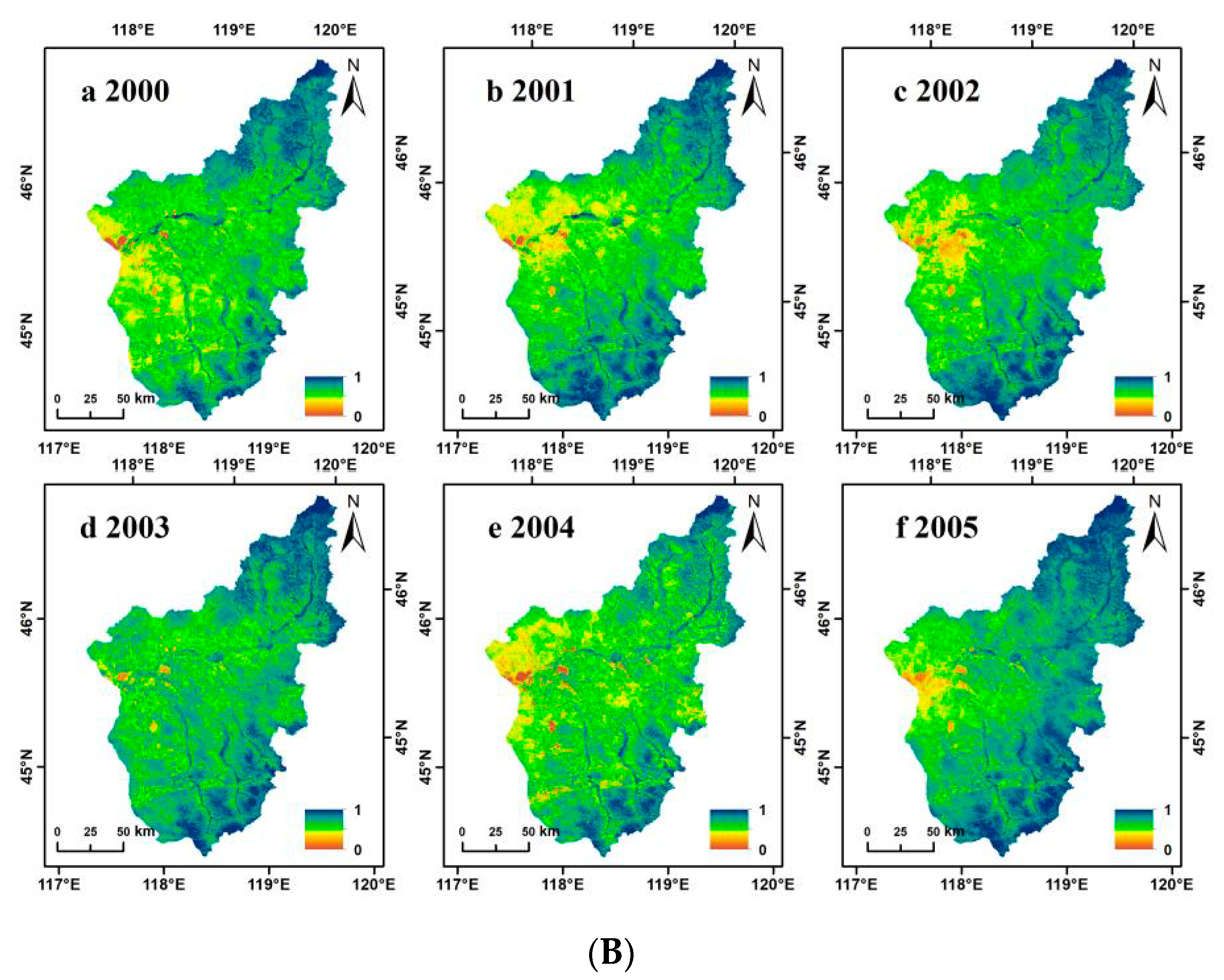
3.1. NDVI Dynamic Change Characteristics
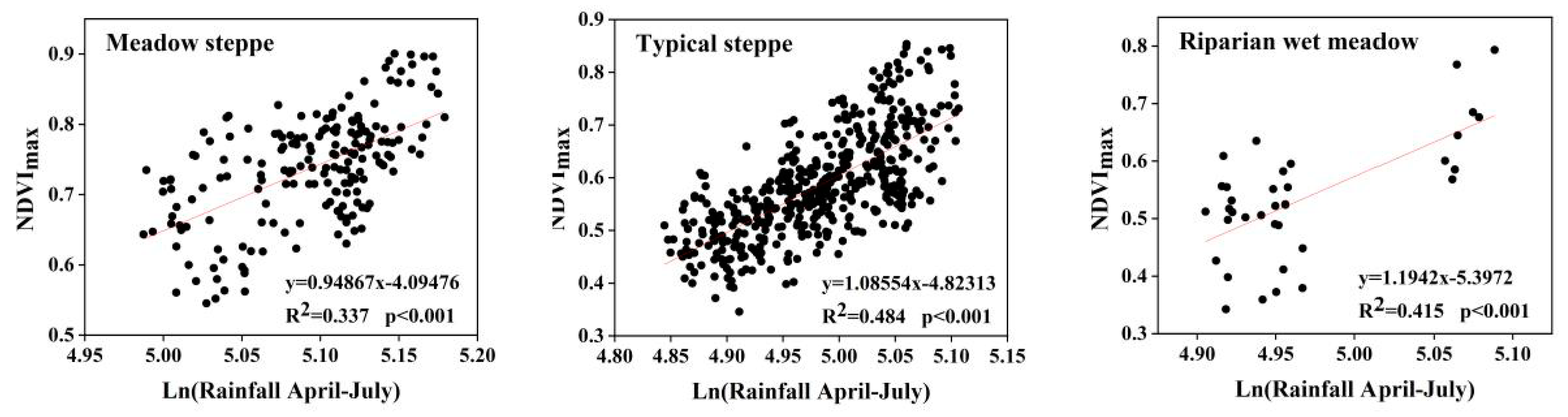
3.2. Correlation between NDVImax Spatial Distribution and Climatic Factors
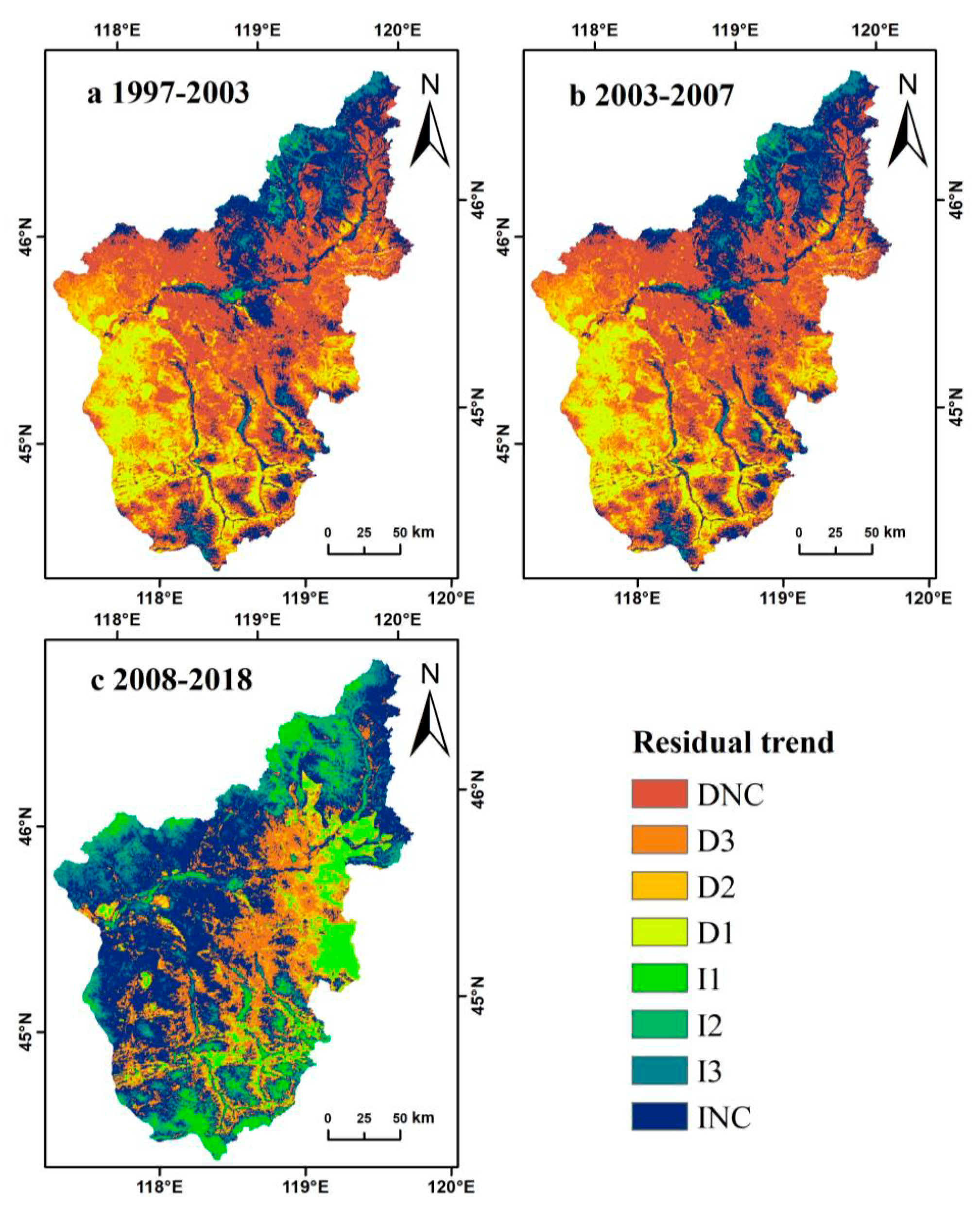
3.3. Residual Analysis
3.4. Community Structure Characteristics of Ecosystem Types in the Wulagai River Basin
4. Discussion
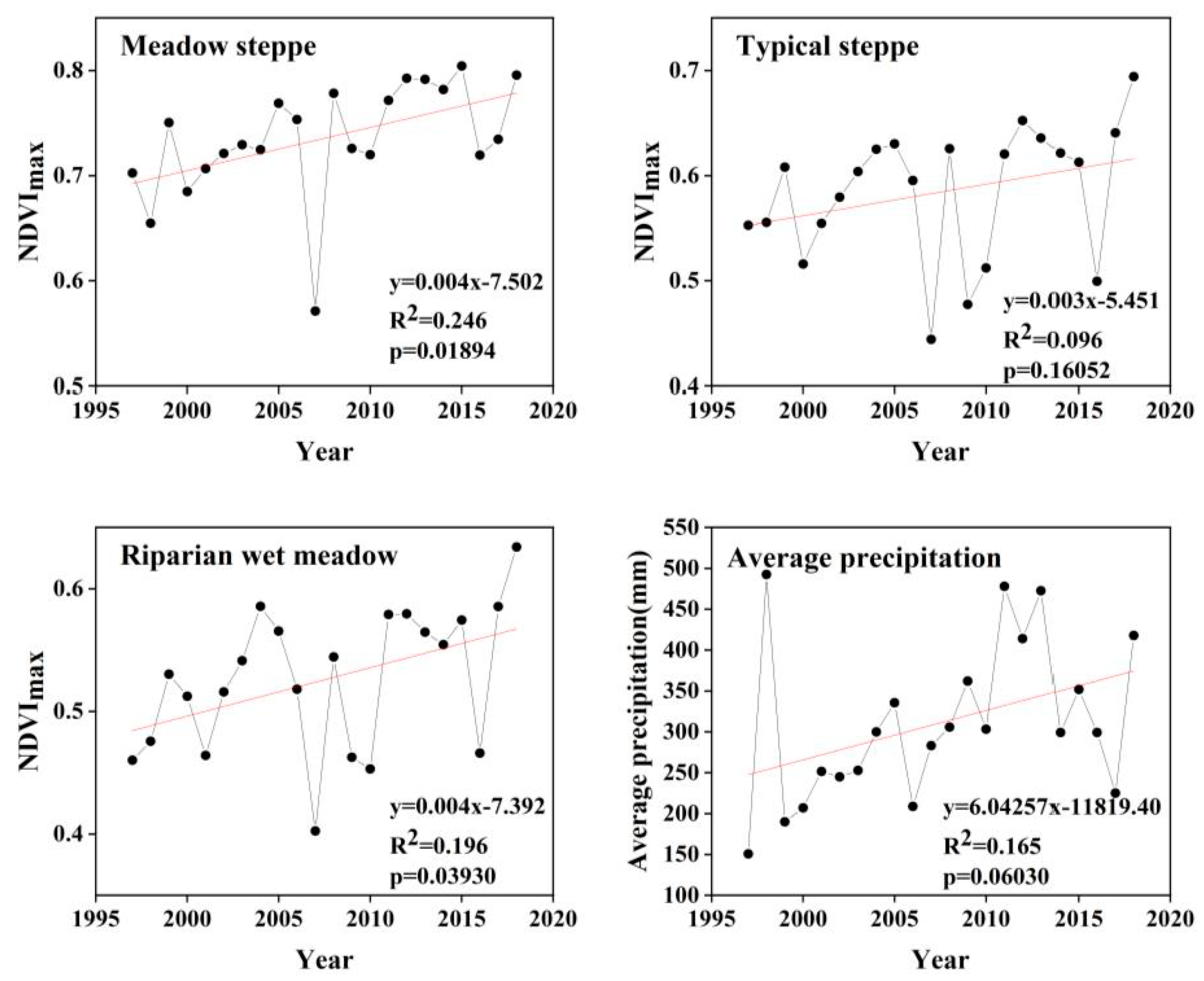
4.1. The Dynamic Changes and Influencing Factors of NDVI in Different Ecosystems
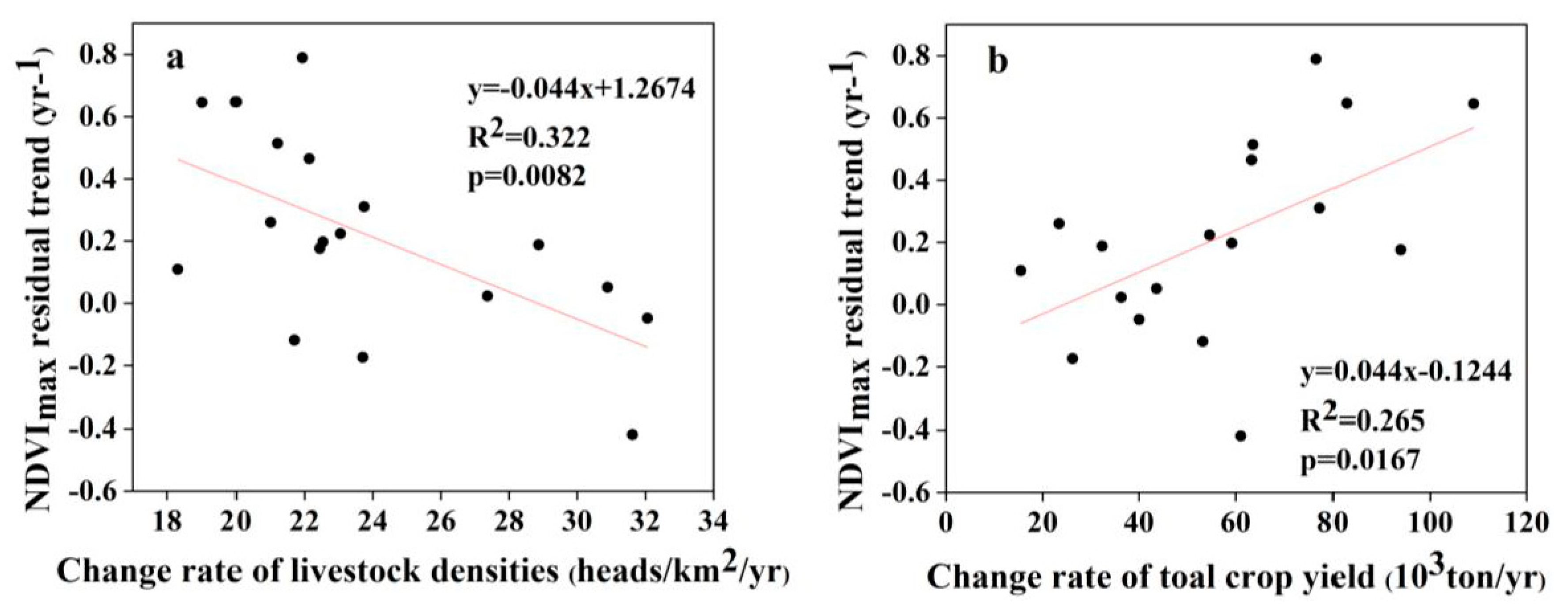
4.2. The Influence of Anthropogenic Factors on the Residual Trend
4.3. Comparison of NDVI and Community Structure Changes in the Same Ecosystem
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Abberton, M.; Conant, R.; Batello, C. Grassland Carbon Sequestration: Management, Policy and Economics. In Proceedings of the Workshop on the Role of Grassland Carbon Sequestration in the Mitigation of Climate Change; FAO: Rome, Italy, 2010; ISBN 978-92-5-106695-9. [Google Scholar]
- Liu, J.; Li, S.; Ouyang, Z.; Tam, C.; Chen, X. Ecological and socioeconomic effects of China’s policies for ecosystem services. Proc. Natl. Acad. Sci. USA 2008, 105, 9477–9482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Long, R.; Shang, Z. Evaluation method of ecological services function and their value for grassland ecosystems. Acta Pratacult. Sin. 2011, 20, 167–174. (In Chinese) [Google Scholar]
- Lu, H.; Raupach, M.R.; McVicar, T.R.; Barrett, D.J. Decomposition of vegetation cover into woody and herbaceous components using AVHRR NDVI time series. Remote Sens. Environ. 2003, 86, 1–18. [Google Scholar] [CrossRef]
- Song, Y.; Ma, M. Study on vegetation cover change in Northwest China based on SPOT VEGETATION data. J. Desert Res. 2007, 27, 89–93. [Google Scholar]
- Justice, C.O.; Hiernaux, P.H. Monitoring the grasslands of the Sahel using NOAA AVHRR data: Niger 1983. Int. J. Remote Sens. 1986, 7, 1475–1497. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R.; Vermote, E.F.; El Saleous, N. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Gallo, K.; Ji, L.; Reed, B.; Dwyer, J.; Eidenshink, J. Comparison of MODIS and AVHRR 16-day normalized difference vegetation index composite data. Geophys. Res. Lett. 2004, 31, L07502. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.Q.; Huang, W.J.; Niu, Z.; Wang, C.Y. Combining HJ CCD, GF-1 WFV and MODIS Data to Generate Daily High Spatial Resolution Synthetic Data for Environmental Process Monitoring. Int. J. Environ. Res. Public Health 2015, 12, 9920–9937. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Zhang, H.; Wu, Z.; Zhao, J.; Zhang, Z. Comparison and evaluation of annual NDVI time series in China derived from the NOAA AVHRR LTDR and Terra MODIS MOD13C1 products. Sensors 2017, 17, 1298. [Google Scholar]
- Du, J.; Zhao, C.; Shu, J.; Jiaerheng, A.; Yuan, X.; Yin, J.; Fang, S.; He, P. Spatiotemporal changes of vegetation on the Tibetan Plateau and relationship to climatic variables during multiyear periods from 1982–2012. Environ. Earth Sci. 2016, 75, 77. [Google Scholar] [CrossRef]
- Gao, F.; Masek, J.; Schwaller, M.; Hall, F. On the blending of the Landsat and MODIS surface reflectance: Predicting daily Landsat surface reflectance. IEEE Trans. Geosci. Remote Sens. 2006, 44, 2207–2218. [Google Scholar]
- Meng, J.; Wang, B.; Du, X.; Niu, L.; Zhang, F. Method to construct high spatial and temporal resolution NDVI DataSet-STAVFM. J. Remote Sens. 2011, 15, 44–59. [Google Scholar]
- Zhu, X.; Chen, J.; Gao, F.; Chen, X.; Masek, J.G. An enhanced spatial and temporal adaptive reflectance fusion model for complex heterogeneous regions. Remote Sens. Environ. 2010, 114, 2610–2623. [Google Scholar] [CrossRef]
- Wang, L.; Liu, H.; Yang, J.; Liang, C.; Wang, W.; Zhang, J. Climatic change of Mu Us Sandy Land and its influence on vegetation coverage. J. Nat. Resour. 2010, 4, 2030–2039. (In Chinese) [Google Scholar]
- Chen, L.; Ma, Z.; Zhao, T. Modeling and analysis of the potential impacts on regional climate due to vegetation degradation over arid and semi-arid regions of China. Clim. Chang. 2017, 144, 461–473. [Google Scholar] [CrossRef]
- Hilker, T.; Natsagdorj, E.; Waring, R.H.; Lyapustin, A.; Wang, Y. Satellite observed widespread decline in Mongolian grasslands largely due to overgrazing. Glob. Chang. Biol. 2014, 20, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Zhao, X.; Wang, S.; Zhao, L.; Duan, J.; Zhang, Z.; Ge, S.; Zhu, X. Grazing intensifies degradation of a Tibetan Plateau alpine meadow through plant–pest interaction. Ecol. Evol. 2015, 5, 2478–2486. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Liu, H.; Fu, Q.; Guan, H.; Ye, Q.; Zhang, X.; Kong, F. Mapping the fallowed area of paddy fields on Sanjiang Plain of Northeast China to assist water security assessments. J. Int. Agric. 2020, 19, 1885–1896. [Google Scholar] [CrossRef]
- Zhou, X.; Yamaguchi, Y.; Arjasakusuma, S. Distinguishing the vegetation dynamics induced by anthropogenic factors using vegetation optical depth and AVHRR NDVI: A cross-border study on the Mongolian Plateau. Sci. Total Environ. 2018, 616, 730–743. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sun, X.; Wang, Z.; Zhang, Y.; Liu, Q.; Zhang, B.; Paudel, B.; Xie, F. Vegetation Changes and Their Response to Global Change Based on NDVI in the Koshi River Basin of Central Himalayas Since 2000. Sustainability 2020, 12, 6644. [Google Scholar] [CrossRef]
- Du, M.; Kawashima, S.; Yonemura, S.; Zhang, X.; Chen, S. Mutual influence between human activities and climate change in the Tibetan Plateau during recent years. Glob. Planet. Chang. 2004, 41, 241–249. [Google Scholar] [CrossRef]
- Huang, K.; Zhang, Y.; Zhu, J.; Liu, Y.; Zu, J.; Zhang, J. The influences of climate change and human activities on vegetation dynamics in the Qinghai-Tibet Plateau. Remote Sens. 2016, 8, 876. [Google Scholar] [CrossRef] [Green Version]
- Willis, C.G.; Ruhfel, B.; Primack, R.B.; Miller-Rushing, A.J.; Davis, C.C. Phylogenetic patterns of species loss in Thoreau’s woods are driven by climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 17029–17033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavergne, S.; Mouquet, N.; Thuiller, W.; Ronce, O. Biodiversity and climate change: Integrating evolutionary and ecological responses of species and communities. Ann. Rev. Ecol. Evol. Syst. 2010, 41, 321–350. [Google Scholar] [CrossRef] [Green Version]
- Cislaghi, A.; Giupponi, L.; Tamburini, A.; Giorgi, A.; Bischetti, G.B. The effects of mountain grazing abandonment on plant community, forage value and soil properties: Observations and field measurements in an alpine area. Catena 2019, 181, 104086. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Dinnage, R.; Tilman, D. Phylogenetic diversity promotes ecosystem stability. Ecology 2012, 93, S223–S233. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Yang, X.; Xu, X. Human-induced grassland degradation/restoration in the central Tibetan Plateau: The effects of ecological protection and restoration projects. Ecol. Eng. 2015, 83, 112–119. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, C.; Wang, Z.; Chen, Y.; Gang, C.; An, R.; Li, J. Vegetation dynamics and its driving forces from climate change and human activities in the Three-River Source Region, China from 1982 to 2012. Sci. Total Environ. 2016, 563–564, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.; Geerken, R. Discrimination between climate and human-induced dryland degradation. J. Arid Environ. 2004, 57, 535–554. [Google Scholar] [CrossRef]
- Wessels, K.J.; Prince, S.; Malherbe, J.; Small, J.; Frost, P.; VanZyl, D. Can human-induced land degradation be distinguished from the effects of rainfall variability? A case study in South Africa. J. Arid Environ. 2007, 68, 271–297. [Google Scholar] [CrossRef]
- Li, A.; Wu, J.; Huang, J. Distinguishing between human-induced and climate-driven vegetation changes: A critical application of RESTREND in inner Mongolia. Landsc. Ecol. 2012, 27, 969–982. [Google Scholar] [CrossRef]
- Pang, Z. Discussion on the Management of District of WULAGAI Grassland Ecological Protection and Construction. Beijing Agric. 2011, 4, 145. (In Chinese) [Google Scholar]
- Fang, J.; Wang, X.; Shen, Z.; Tang, Z.; He, J.; Yu, D.; Jiang, Y.; Wang, Z.; Zheng, C.; Zhu, J. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533–548. [Google Scholar]
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens. 1986, 7, 1417–1434. [Google Scholar] [CrossRef]
- Wang, Z.; Bovik, A.C. A universal image quality index. IEEE Signal Process. Lett. 2002, 9, 81–84. [Google Scholar] [CrossRef]
- Nkonya, E.; Anderson, W.; Kato, E.; Koo, J.; Mirzabaev, A.; von Braun, J.; Meyer, S. Global cost of land degradation. In Economics of Land Degradation and Improvement–A Global Assessment for Sustainable Development; Springer: Berlin, Germany, 2016; pp. 117–165. [Google Scholar]
- Davis, J.; Pavlova, A.; Thompson, R.; Sunnucks, P. Evolutionary refugia and ecological refuges: Key concepts for conserving Australian arid zone freshwater biodiversity under climate change. Glob. Chang. Biol. 2013, 19, 1970–1984. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Viña, A.; Yang, W.; Chen, X.; Zhang, J.; Ouyang, Z.; Liang, Z.; Liu, J. Effects of conservation policies on forest cover change in giant panda habitat regions, China. Land Use Policy 2013, 33, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, S.E.; Davenport, M.L.; Malo, A.R. A comparison of the vegetation response to rainfall in the Sahel and East Africa, using normalized difference vegetation index from NOAA AVHRR. Clim. Chang. 1990, 17, 209–241. [Google Scholar] [CrossRef]
- Li, B.; Tao, S.; Dawson, R. Relations between AVHRR NDVI and ecoclimatic parameters in China. Int. J. Remote Sens. 2002, 23, 989–999. [Google Scholar] [CrossRef]
- Mao, D.; Wang, Z.; Luo, L.; Ren, C. Integrating AVHRR and MODIS data to monitor NDVI changes and their relationships with climatic parameters in Northeast China. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 528–536. [Google Scholar] [CrossRef]
- Fang, J.; Piao, S.; He, J.; Ma, W. Increasing terrestrial vegetation activity in China, 1982–1999. Sci. China Ser. C Life Sci. 2004, 47, 229–240. [Google Scholar]
- Piao, S.; Mohammat, A.; Fang, J.; Cai, Q.; Feng, J. NDVI-based increase in growth of temperate grasslands and its responses to climate changes in China. Glob. Environ. Chang. 2006, 16, 340–348. [Google Scholar] [CrossRef]
- Jiang, G.; Han, X.; Wu, J. Restoration and management of the Inner Mongolia grassland require a sustainable strategy. AMBIO J. Human Environ. 2006, 35, 269–270. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Gu, Z.; Chen, J.; Liu, J.; Shi, P. Analysis of human-induced steppe degradation based on remote sensing in Xilin Gole, Inner Mongolia, China. Acta Pratacult. Sin. 2006, 30, 268–277. (In Chinese) [Google Scholar]
- Kohyani, P.T.; Bossuyt, B.; Bonte, D.; Hoffmann, M. Differential herbivory tolerance of dominant and subordinate plant species along gradients of nutrient availability and competition. In Herbaceous Plant Ecology; Springer: Berlin, Germany, 2009; pp. 247–255. [Google Scholar]
- Zhuo, L.; Cao, X.; Chen, J.; Chen, Z.; Shi, P. Assessment of grassland ecological restoration project in Xilin Gol grassland. Acta Geogr. Sin. 2007, 62, 471–480. [Google Scholar]
- Wessels, K.J.; Van Den Bergh, F.; Scholes, R. Limits to detectability of land degradation by trend analysis of vegetation index data. Remote Sens. Environ. 2012, 125, 10–22. [Google Scholar] [CrossRef]
- Hu, Y.; Dao, R.; Hu, Y. Vegetation change and driving factors: Contribution analysis in the loess plateau of China during 2000–2015. Sustainability 2019, 11, 1320. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Scottá, F.C.; Da Fonseca, E.L. Multiscale trend analysis for pampa grasslands using ground data and vegetation sensor imagery. Sensors 2015, 15, 17666–17692. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cumulative Rainfall Period | Rainfall April–July | Rainfall April–August | Rainfall June–August | Ln (Rainfall April–July) | Ln (Rainfall April–August) | Ln (Rainfall June–August) | Cumulative Temp January–August | Ln (Cumulative Temp January–August) | |
|---|---|---|---|---|---|---|---|---|---|
| Meadow steppe | Correlation coefficient | 0.32 | 0.22 | 0.22 | 0.39 | 0.18 | 0.20 | −0.25 | −0.04 |
| Percentage of p < 0.05 | 27.62 | 6.84 | 7.66 | 49.66 | 4.72 | 5.56 | 11.95 | 1.22 | |
| Typical steppe | Correlation coefficient | 0.33 | 0.27 | 0.30 | 0.41 | 0.28 | 0.33 | −0.21 | −0.09 |
| Percentage of p < 0.05 | 32.36 | 24.73 | 32.03 | 51.65 | 27.77 | 36.17 | 10.56 | 1.28 | |
| Riparian wet meadow | Correlation coefficient | 0.25 | 0.23 | 0.30 | 0.33 | 0.23 | 0.30 | −0.115 | −0.025 |
| Percentage of p < 0.05 | 22.21 | 16.81 | 35.41 | 42.60 | 16.61 | 35.23 | 3.96 | 1.65 |
| Residual | 1997–2003 (%) | 2003–2007 (%) | 2008–2018 (%) |
|---|---|---|---|
| D1, D2, D3 | 36.31 | 1.76 | 15.58 |
| DNC | 37.75 | 16.01 | 18.37 |
| INC | 21.08 | 58.96 | 39.08 |
| I1, I2, I3 | 4.86 | 23.27 | 26.97 |
| Total | 100.00 | 100.00 | 100.00 |
| Human Factors | Period | Coefficient of Determination (Direction) |
|---|---|---|
| Rate of change in livestock densities | 1997–2001 | 0.009 (−) |
| 2001–2018 | 0.044 ** (−) | |
| Rate of change in total crop yield | 1997–2001 | 3.38E-5 (−) |
| 2001–2018 | 6.225E-6 * (+) |
| Ecosystem Type | Years | Community Type | Main Species | Average Plant Height (cm) | Above-Ground Biomass (g/m2) |
|---|---|---|---|---|---|
| Meadow steppe | 1997 | Stipa baicalensis + Filifolium sibircum | Stipa baicalensis, Filifolium sibircum, Carex pediformis, Artemisia tanacetifolia, Leucopoa albida | 18.21 ± 0.5 | 148.31 ± 3 |
| 2018 | S. baicalensis + Carex korshinskyi | Stipa baicalensis, Carex korshinskyi, Sanguisorba officinalis, Filifolium sibircum, Serratula centauroides | 20.77 ± 0.2 | 159.02 ± 2 | |
| Typical steppe | 1997 | S. grandis + Leymus chinensis | Stipa grandis, Leymus chinensis, Artemisia frigida, Euphorbia fischeriana, Scutellaria baicalensis | 14.17 ± 0.2 | 83.96 ± 5 |
| 2018 | S. grandis + S. krylovii + L. chinensis | Stipa grandis, Stipa krylovii, Leymus chinensis, Euphorbia fischeriana, Artemisia frigida, Alium ramosm | 12.53 ± 0.3 | 60.89 ± 3 | |
| Riparian wet meadow | 1997 | Agrostis alba + Potentilla anserina | Agrostis alba, Potentilla anserina, Halerpestes ruthenica, Suaeda glauca, Carex korshinskyi | 14.66 ± 0.5 | 196.26 ± 4 |
| 2018 | C. korshinskyi + Hemerocallis minor | Carex korshinskyi, Hemerocallis minor, Agrostis alba, Potentilla anserina, Suaeda glauca | 18.78 ± 0.3 | 233.16 ± 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.; Liu, H.; Wang, Z.; Mao, D.; Liang, C.; Wen, L.; Li, Z.; Zhang, J.; Liu, D.; Zhuo, Y.; et al. Vegetation Dynamic Assessment by NDVI and Field Observations for Sustainability of China’s Wulagai River Basin. Int. J. Environ. Res. Public Health 2021, 18, 2528. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052528
Chen P, Liu H, Wang Z, Mao D, Liang C, Wen L, Li Z, Zhang J, Liu D, Zhuo Y, et al. Vegetation Dynamic Assessment by NDVI and Field Observations for Sustainability of China’s Wulagai River Basin. International Journal of Environmental Research and Public Health. 2021; 18(5):2528. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052528
Chicago/Turabian StyleChen, Panpan, Huamin Liu, Zongming Wang, Dehua Mao, Cunzhu Liang, Lu Wen, Zhiyong Li, Jinghui Zhang, Dongwei Liu, Yi Zhuo, and et al. 2021. "Vegetation Dynamic Assessment by NDVI and Field Observations for Sustainability of China’s Wulagai River Basin" International Journal of Environmental Research and Public Health 18, no. 5: 2528. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18052528