
n-Butyl Benzyl Phthalate Exposure Promotes Lesion Survival in a Murine Endometriosis Model




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice and the Surgically Induced Endometriosis Model
2.2. Flow Cytometry for Analysis of Immune Cell Subsets
2.3. Immunofluorescence Staining
2.4. Vaginal Cytology
2.5. Statistical Analyses
3. Results
3.1. Chronic Exposure to Low-Dose BBP Promotes Endometriotic Lesion Survival in a Surgically Induced Murine Model
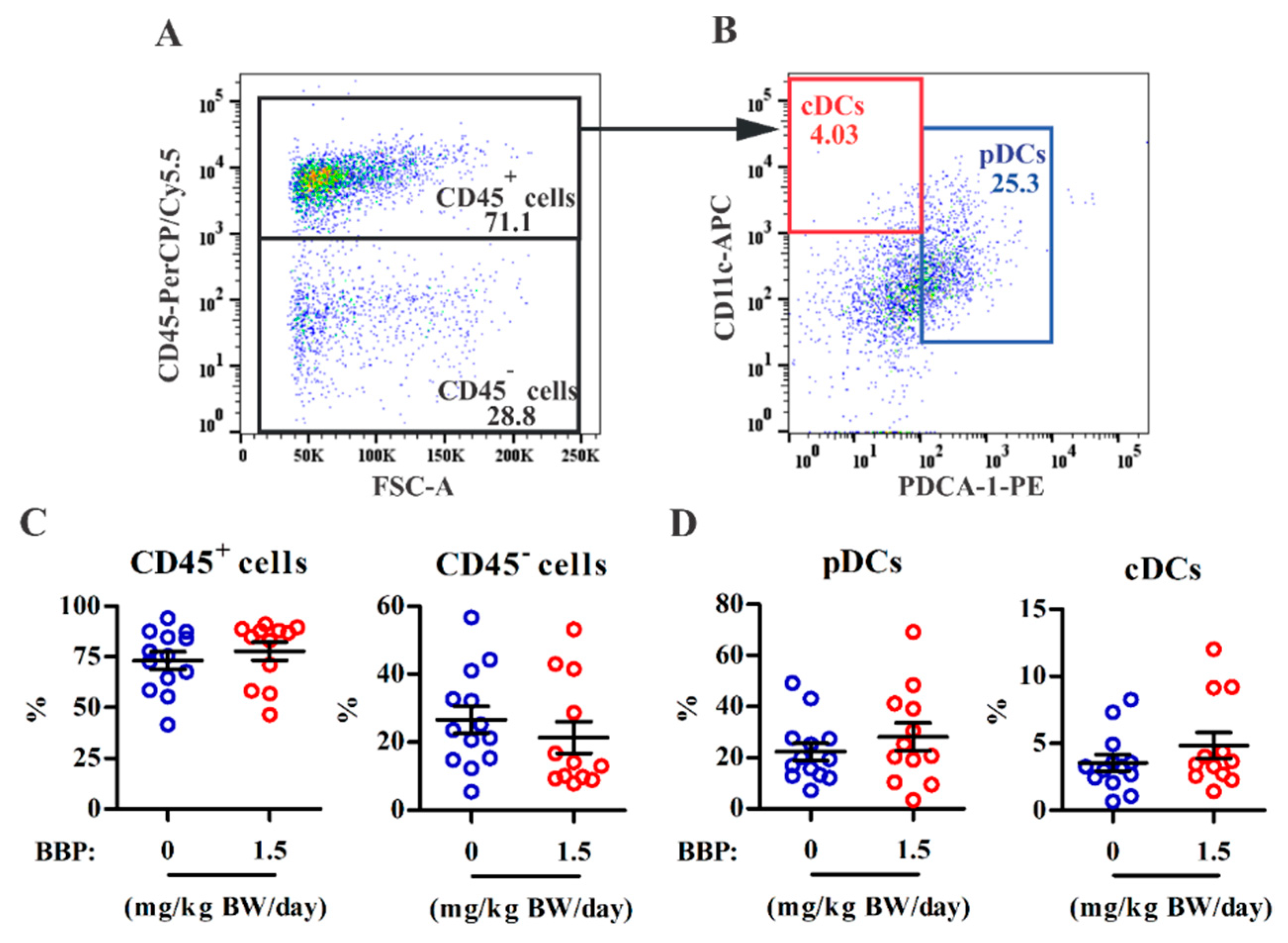
3.2. BBP-Mediated Lesion Survival Is Not Associated with Infiltration of Innate Immune Subsets in Lesions
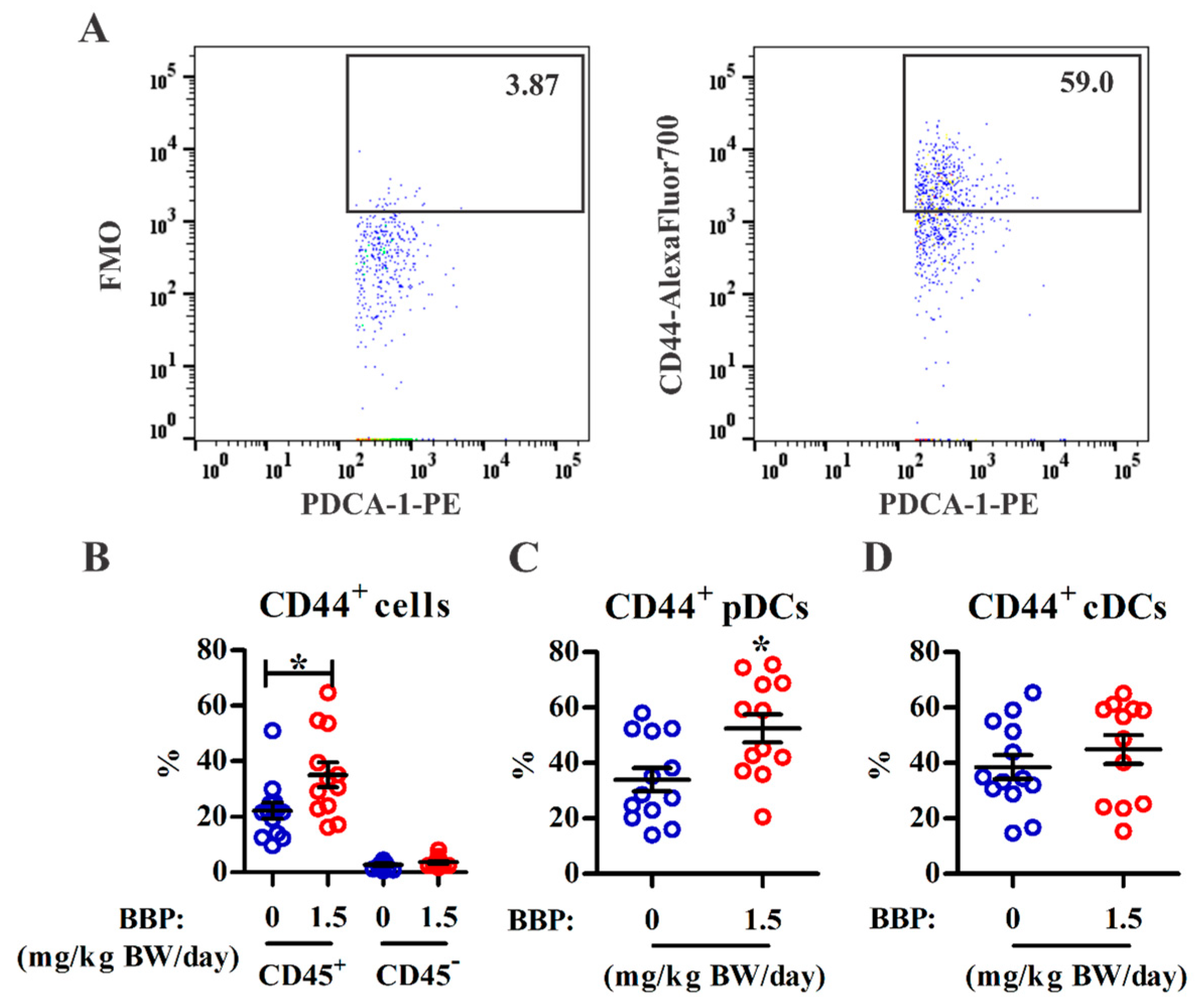
3.3. BBP Exposure Enhances Adhesion Marker Expression on Infiltrating pDCs
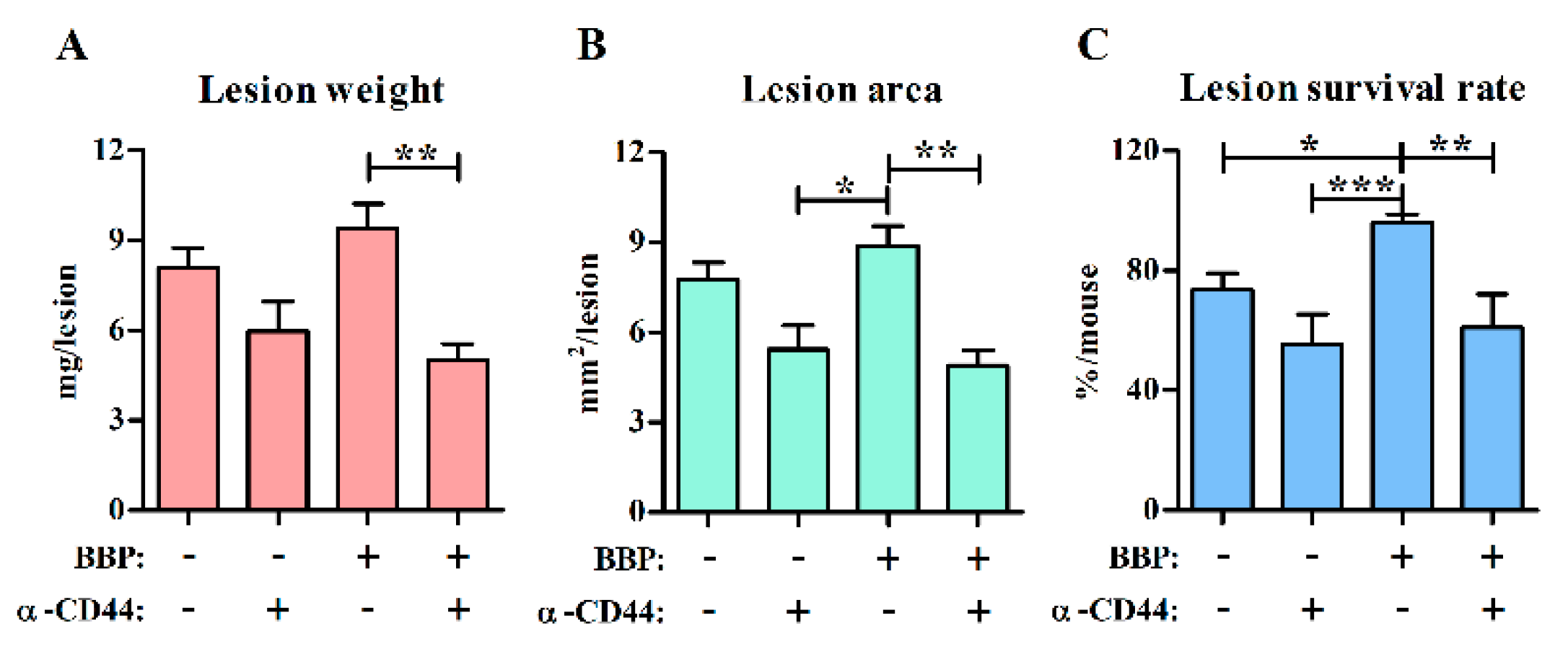
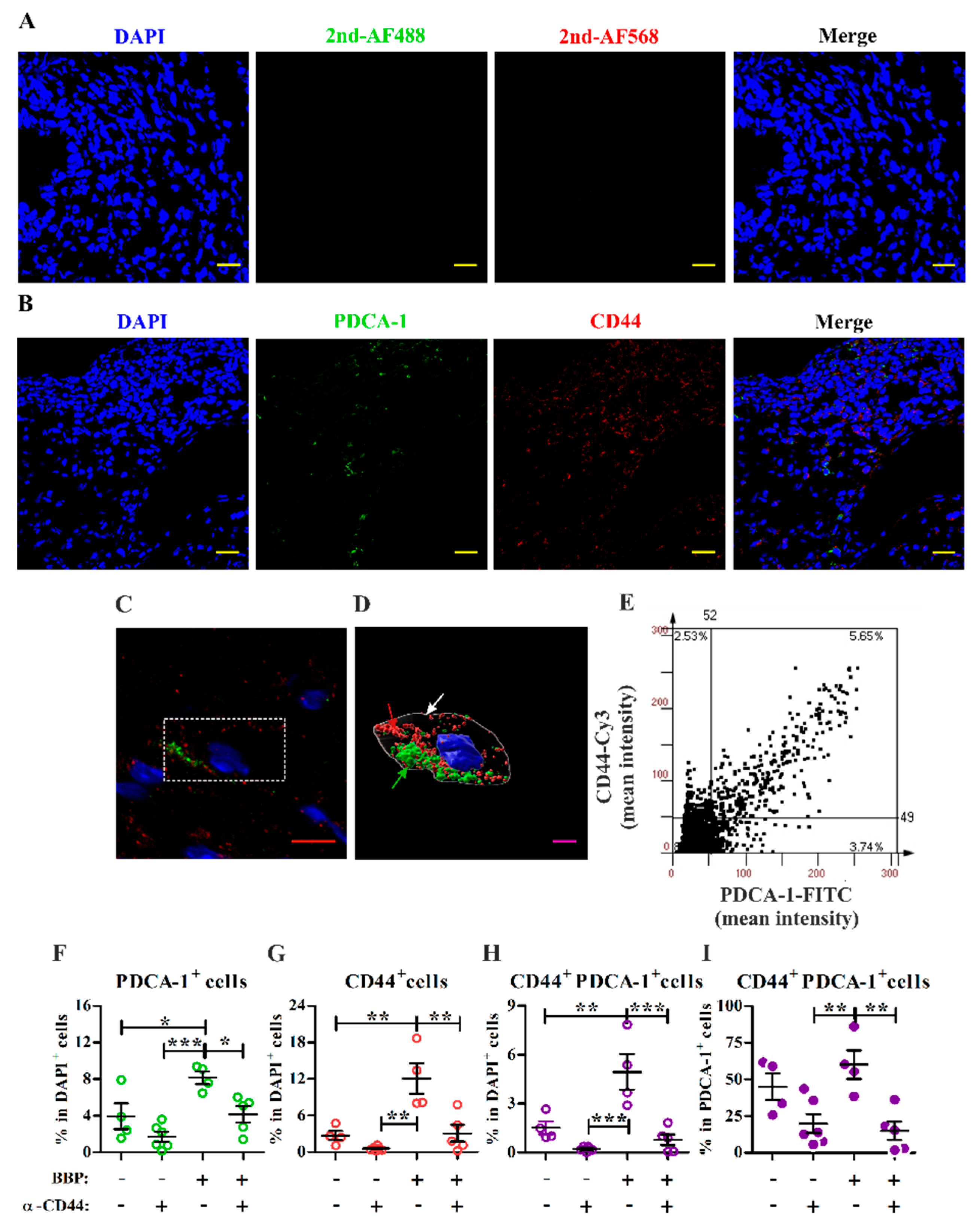
3.4. Blocking Interactions with CD44 Inhibits Endometriotic Lesion Development and CD44+ pDC Infiltration into Endometriotic Lesions
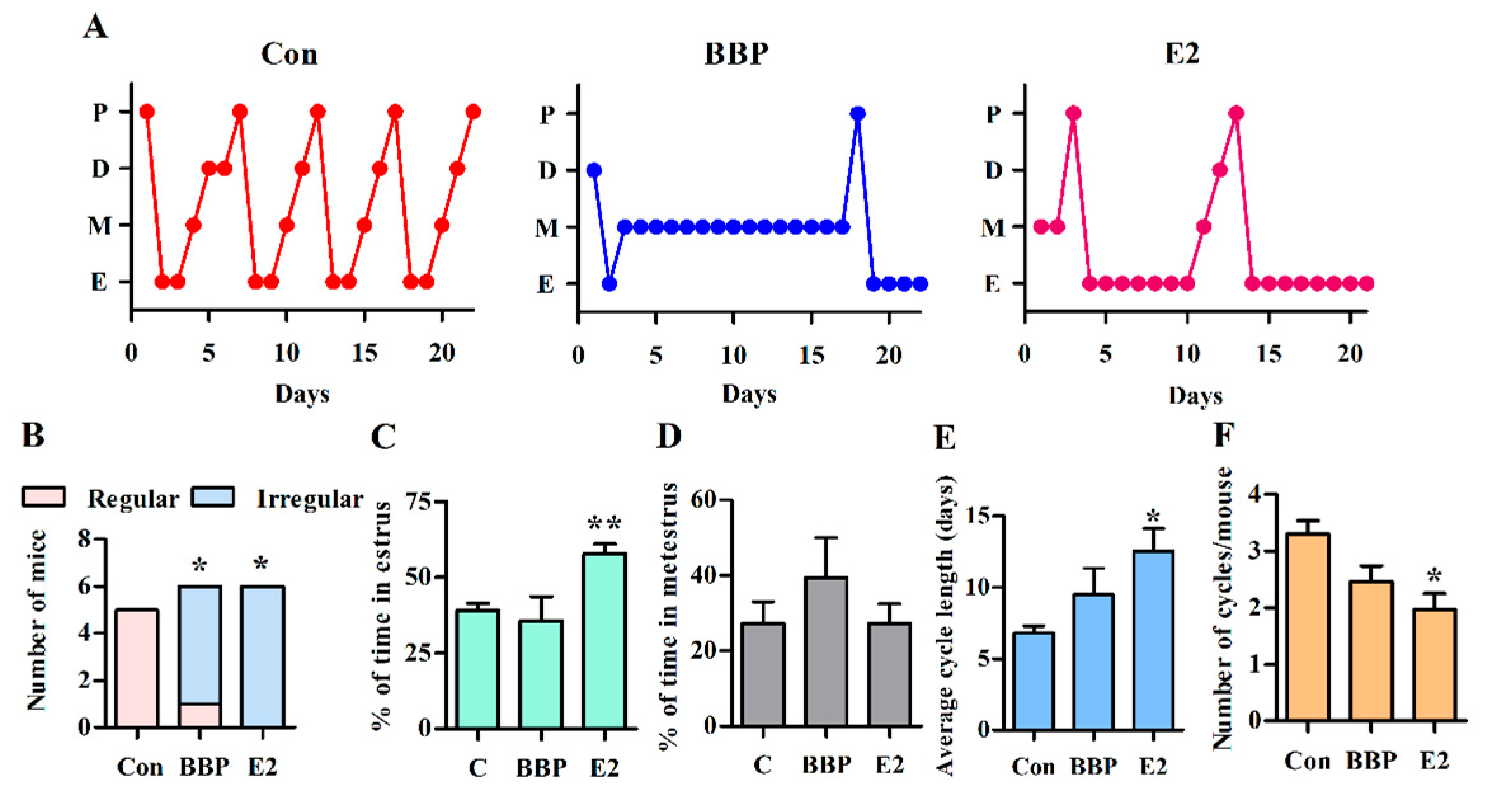
3.5. Chronic Exposure to Low-Dose BBP Alters the Estrous Cycle in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zondervan, K.T.; Becker, C.M.; Koga, K.; Missmer, S.A.; Taylor, R.N.; Vigano, P. Endometriosis. Nat. Rev. Dis. Primers 2018, 4, 9. [Google Scholar] [CrossRef] [PubMed]
- Vercellini, P.; Vigano, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Halme, J.; Hammond, M.G.; Hulka, J.F.; Raj, S.G.; Talbert, L.M. Retrograde menstruation in healthy women and in patients with endometriosis. Obstet. Gynecol. 1984, 64, 151–154. [Google Scholar] [PubMed]
- Burney, R.O.; Giudice, L.C. Pathogenesis and pathophysiology of endometriosis. Fertil Steril 2012, 98, 511–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symons, L.K.; Miller, J.E.; Kay, V.R.; Marks, R.M.; Liblik, K.; Koti, M.; Tayade, C. The Immunopathophysiology of Endometriosis. Trends Mol. Med. 2018, 24, 748–762. [Google Scholar] [CrossRef] [PubMed]
- Suen, J.L.; Chang, Y.; Shiu, Y.S.; Hsu, C.Y.; Sharma, P.; Chiu, C.C.; Chen, Y.J.; Hour, T.C.; Tsai, E.M. IL-10 from plasmacytoid dendritic cells promotes angiogenesis in the early stage of endometriosis. J. Pathol. 2019, 249, 485–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suen, J.L.; Chang, Y.; Chiu, P.R.; Hsieh, T.H.; Hsi, E.; Chen, Y.C.; Chen, Y.F.; Tsai, E.M. Serum level of IL-10 is increased in patients with endometriosis, and IL-10 promotes the growth of lesions in a murine model. Am. J. Pathol. 2014, 184, 464–471. [Google Scholar] [CrossRef]
- Chairakaki, A.D.; Saridaki, M.I.; Pyrillou, K.; Mouratis, M.A.; Koltsida, O.; Walton, R.P.; Bartlett, N.W.; Stavropoulos, A.; Boon, L.; Rovina, N.; et al. Plasmacytoid dendritic cells drive acute asthma exacerbations. J. Allergy Clin. Immunol. 2018, 142, 542–556.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.K.; Liu, L.B.; Jin, L.P.; Zhang, B.; Mei, J.; Li, H.; Wei, C.Y.; Zhou, W.J.; Zhu, X.Y.; Shao, J.; et al. IL-27 triggers IL-10 production in Th17 cells via a c-Maf/RORgammat/Blimp-1 signal to promote the progression of endometriosis. Cell Death Dis. 2017, 8, e2666. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.; Peiris-John, R.; Wickremasinghe, R.; Senanayake, H.; Sathiakumar, N. Cadmium a metalloestrogen: Are we convinced? J. Appl. Toxicol. 2012, 32, 318–332. [Google Scholar] [CrossRef]
- Schiattarella, A.; Colacurci, N.; Morlando, M.; Ammaturo, F.P.; Genovese, G.; Miraglia, N.; Franciscis, P.D. Plasma and urinary levels of lead and cadmium in patients with endometriosis. Ital. J. Gynaecol. Obstet. 2018, 30, 47–52. [Google Scholar]
- De Franciscis, P.; Guadagno, M.; Miraglia, N.; D’Eufemia, D.; Schiattarella, A.; Labriola, D.; Cucinella, G.; Colacurci, N. Follicular PB levels in women attending in vitro fertilization: Role of endometriosis on the outcome. Ital. J. Gynaecol. Obstet. 2018, 30, 21–27. [Google Scholar]
- Upson, K.; Sathyanarayana, S.; De Roos, A.J.; Thompson, M.L.; Scholes, D.; Dills, R.; Holt, V.L. Phthalates and risk of endometriosis. Environ. Res. 2013, 126, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heudorf, U.; Mersch-Sundermann, V.; Angerer, J. Phthalates: Toxicology and exposure. Int. J. Hyg. Environ. Health 2007, 210, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Shi, J.; Bo, T.; Li, H.; Crittenden, J.C. Occurrence and risk assessment of selected phthalates in drinking water from waterworks in China. Environ. Sci. Pollut. Res. Int. 2015, 22, 10690–10698. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Morueco, N.; Gonzalez-Alonso, S.; Valcarcel, Y. Phthalate occurrence in rivers and tap water from central Spain. Sci. Total Environ. 2014, 500–501, 139–146. [Google Scholar] [CrossRef]
- Bolger, R.; Wiese, T.E.; Ervin, K.; Nestich, S.; Checovich, W. Rapid screening of environmental chemicals for estrogen receptor binding capacity. Environ. Health Perspect. 1998, 106, 551–557. [Google Scholar] [CrossRef]
- Moore, N.P. The oestrogenic potential of the phthalate esters. Reprod. Toxicol. 2000, 14, 183–192. [Google Scholar] [CrossRef]
- Fujita, T.; Kobayashi, Y.; Wada, O.; Tateishi, Y.; Kitada, L.; Yamamoto, Y.; Takashima, H.; Murayama, A.; Yano, T.; Baba, T.; et al. Full activation of estrogen receptor alpha activation function-1 induces proliferation of breast cancer cells. J. Biol. Chem. 2003, 278, 26704–26714. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.S.; Hsu, C.Y.; Chang, Y.C.; Chuang, H.Y.; Long, C.Y.; Hsieh, T.H.; Tsai, E.M. Benzyl butyl phthalate decreases myogenic differentiation of endometrial mesenchymal stem/stromal cells through miR-137-mediated regulation of PITX2. Sci. Rep. 2017, 7, 186. [Google Scholar] [CrossRef] [Green Version]
- Reddy, B.S.; Rozati, R.; Reddy, B.V.R.; Raman, N.V.V.S.S. General gynaecology: Association of phthalate esters with endometriosis in Indian women. BJOG Int. J. Obstet. Gynaecol. 2006, 113, 515–520. [Google Scholar] [CrossRef]
- EFSA Panel on Food Contact Materials, Enzymes and Processing Aids (CEP); Silano, V.; Barat Baviera, J.M.; Bolognesi, C.; Chesson, A.; Cocconcelli, P.S.; Crebelli, R.; Gott, D.M.; Grob, K.; Lampi, E.; et al. Update of the risk assessment of ‘wood flour and fibres, untreated’(FCM No 96) for use in food contact materials, and criteria for future applications of materials from plant origin as additives for plastic food contact materials. EFSA J. 2019, 17, e05902. [Google Scholar] [CrossRef]
- McLean, A.C.; Valenzuela, N.; Fai, S.; Bennett, S.A. Performing vaginal lavage, crystal violet staining, and vaginal cytological evaluation for mouse estrous cycle staging identification. J. Vis. Exp. 2012, 67, e4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Jiang, D.G.; Sui, H.X.; Wu, P.G.; Liu, A.D.; Yang da, J.; Liu, Z.P.; Song, Y.; Li, N. Dietary Exposure to Benzyl Butyl Phthalate in China. Biomed. Environ. Sci. 2016, 29, 365–373. [Google Scholar]
- Wittassek, M.; Wiesmuller, G.A.; Koch, H.M.; Eckard, R.; Dobler, L.; Muller, J.; Angerer, J.; Schluter, C. Internal phthalate exposure over the last two decades--a retrospective human biomonitoring study. Int. J. Hyg. Environ. Health 2007, 210, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Kay, V.R.; Chambers, C.; Foster, W.G. Reproductive and developmental effects of phthalate diesters in females. Crit. Rev. Toxicol. 2013, 43, 200–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyl, R.W.; Myers, C.B.; Marr, M.C.; Fail, P.A.; Seely, J.C.; Brine, D.R.; Barter, R.A.; Butala, J.H. Reproductive toxicity evaluation of dietary butyl benzyl phthalate (BBP) in rats. Reprod. Toxicol. 2004, 18, 241–264. [Google Scholar] [CrossRef]
- Sharma, P.; Tseng, H.H.; Lee, J.L.; Tsai, E.M.; Suen, J.L. A prominent environmental endocrine disruptor, 4-nonylphenol, promotes endometriosis development via plasmacytoid dendritic cells. Mol. Hum. Reprod. 2020, 26, 601–614. [Google Scholar] [CrossRef]
- Fainaru, O.; Adini, A.; Benny, O.; Adini, I.; Short, S.; Bazinet, L.; Nakai, K.; Pravda, E.; Hornstein, M.D.; D’Amato, R.J.; et al. Dendritic cells support angiogenesis and promote lesion growth in a murine model of endometriosis. FASEB J. 2008, 22, 522–529. [Google Scholar] [CrossRef] [PubMed]
- Capobianco, A.; Rovere-Querini, P. Endometriosis, a disease of the macrophage. Front. Immunol. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.W.; Huang, M.T.; Wang, P.H.; Huang, B.S.; Chen, Y.J.; Hsieh, S.L. Decoy receptor 3 promotes cell adhesion and enhances endometriosis development. J. Pathol. 2018, 244, 189–202. [Google Scholar] [CrossRef] [PubMed]
- Marafioti, T.; Paterson, J.C.; Ballabio, E.; Reichard, K.K.; Tedoldi, S.; Hollowood, K.; Dictor, M.; Hansmann, M.L.; Pileri, S.A.; Dyer, M.J.; et al. Novel markers of normal and neoplastic human plasmacytoid dendritic cells. Blood 2008, 111, 3778–3792. [Google Scholar] [CrossRef] [Green Version]
- Jordan, A.R.; Racine, R.R.; Hennig, M.J.; Lokeshwar, V.B. The Role of CD44 in Disease Pathophysiology and Targeted Treatment. Front. Immunol. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed]
- Cannistra, S.A.; Kansas, G.S.; Niloff, J.; Defranzo, B.; Kim, Y.; Ottensmeier, C. Binding of Ovarian-Cancer Cells to Peritoneal Mesothelium in-Vitro Is Partly Mediated by Cd44h. Cancer Res. 1993, 53, 3830–3838. [Google Scholar] [PubMed]
- Lessan, K.; Aguiar, D.J.; Oegema, T.; Siebenson, L.; Skubitz, A.P. CD44 and beta1 integrin mediate ovarian carcinoma cell adhesion to peritoneal mesothelial cells. Am. J. Pathol. 1999, 154, 1525–1537. [Google Scholar] [CrossRef]
- Videira, P.A.; Silva, M.; Martin, K.C.; Sackstein, R. Ligation of the CD44 Glycoform HCELL on Culture-Expanded Human Monocyte-Derived Dendritic Cells Programs Transendothelial Migration. J. Immunol. 2018, 201, 1030–1043. [Google Scholar] [CrossRef]
- Termeer, C.; Averbeck, M.; Hara, H.; Eibel, H.; Herrlich, P.; Sleeman, J.; Simon, J.C. Targeting dendritic cells with CD44 monoclonal antibodies selectively inhibits the proliferation of naive CD4+ T-helper cells by induction of FAS-independent T-cell apoptosis. Immunology 2003, 109, 32–40. [Google Scholar] [CrossRef]
- Schulke, L.; Berbic, M.; Manconi, F.; Tokushige, N.; Markham, R.; Fraser, I.S. Dendritic cell populations in the eutopic and ectopic endometrium of women with endometriosis. Hum. Reprod. 2009, 24, 1695–1703. [Google Scholar] [CrossRef] [Green Version]
- Pencovich, N.; Luk, J.; Hantisteanu, S.; Hornstein, M.D.; Fainaru, O. The development of endometriosis in a murine model is dependent on the presence of dendritic cells. Reprod. Biomed. Online 2014, 28, 515–521. [Google Scholar] [CrossRef] [Green Version]
- Rubin, B.S.; Murray, M.K.; Damassa, D.A.; King, J.C.; Soto, A.M. Perinatal exposure to low doses of bisphenol A affects body weight, patterns of estrous cyclicity, and plasma LH levels. Environ. Health Perspect. 2001, 109, 675–680. [Google Scholar] [CrossRef]
- Greaves, E.; Cousins, F.L.; Murray, A.; Esnal-Zufiaurre, A.; Fassbender, A.; Horne, A.W.; Saunders, P.T. A novel mouse model of endometriosis mimics human phenotype and reveals insights into the inflammatory contribution of shed endometrium. Am. J. Pathol. 2014, 184, 1930–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Li, D.; An, M.; Li, Q.; Zhang, L.; Wang, G. Rediscovering peritoneal macrophages in a murine endometriosis model. Hum. Reprod. 2017, 32, 94–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, P.; Lee, J.-Y.L.; Tsai, E.-M.; Chang, Y.; Suen, J.-L. n-Butyl Benzyl Phthalate Exposure Promotes Lesion Survival in a Murine Endometriosis Model. Int. J. Environ. Res. Public Health 2021, 18, 3640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073640
Sharma P, Lee J-YL, Tsai E-M, Chang Y, Suen J-L. n-Butyl Benzyl Phthalate Exposure Promotes Lesion Survival in a Murine Endometriosis Model. International Journal of Environmental Research and Public Health. 2021; 18(7):3640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073640
Chicago/Turabian StyleSharma, Pooja, Jo-Yu Lynn Lee, Eing-Mei Tsai, Yu Chang, and Jau-Ling Suen. 2021. "n-Butyl Benzyl Phthalate Exposure Promotes Lesion Survival in a Murine Endometriosis Model" International Journal of Environmental Research and Public Health 18, no. 7: 3640. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18073640