
Influence of Amino Acid Feeding on Production of Calcimycin and Analogs in Streptomyces chartreusis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Cultivation Conditions
2.2. Compound Extraction
2.3. LC-MS/MS Measurements
2.4. Molecular Networking
2.5. Heatmap Modeling
3. Results
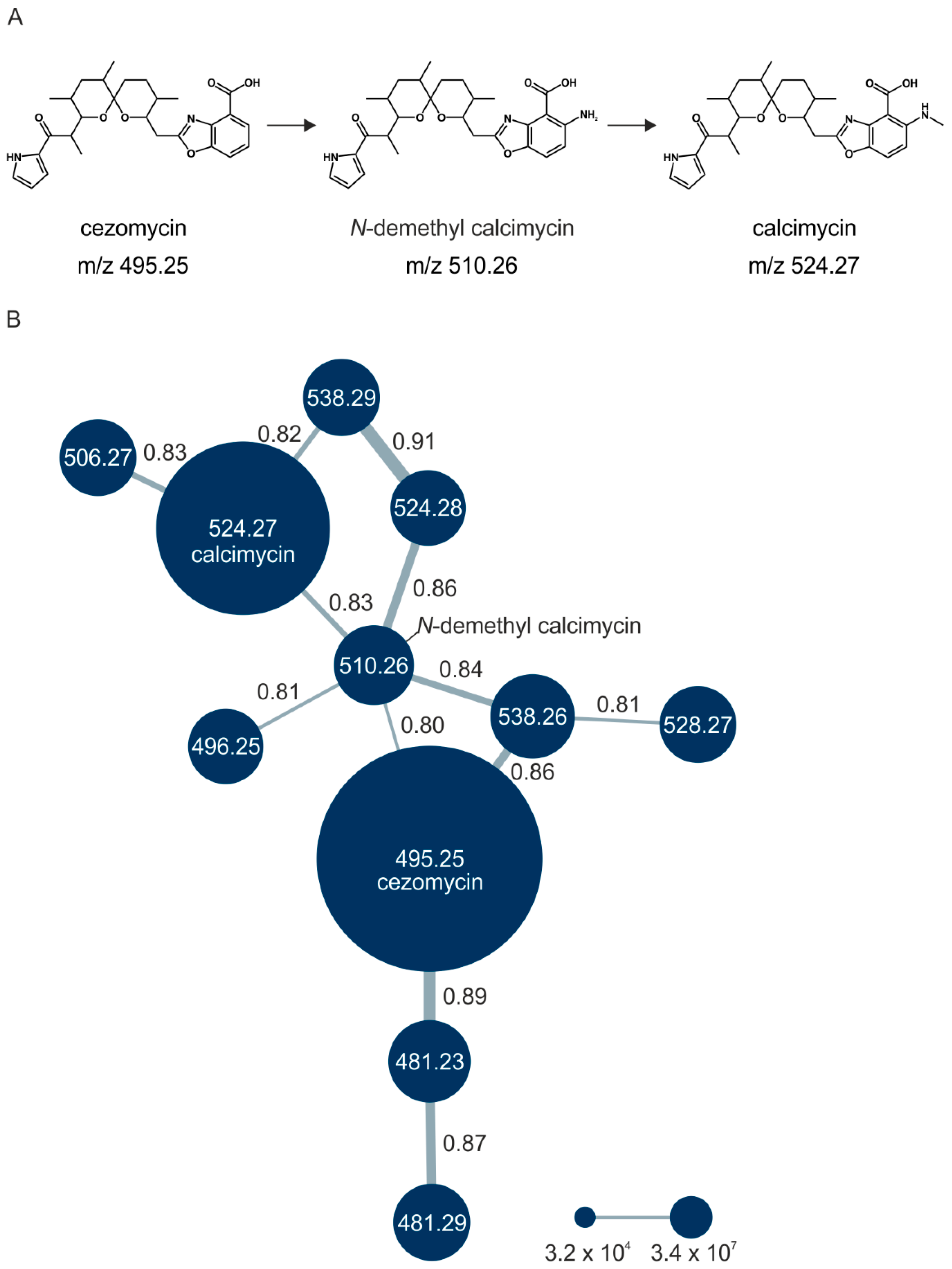
3.1. Amino Acid Supplementation Leads to the Detection of New Calcimycin Analogs
3.2. Cezomycin Is the Most Abundant Analog in Chemically Defined Medium
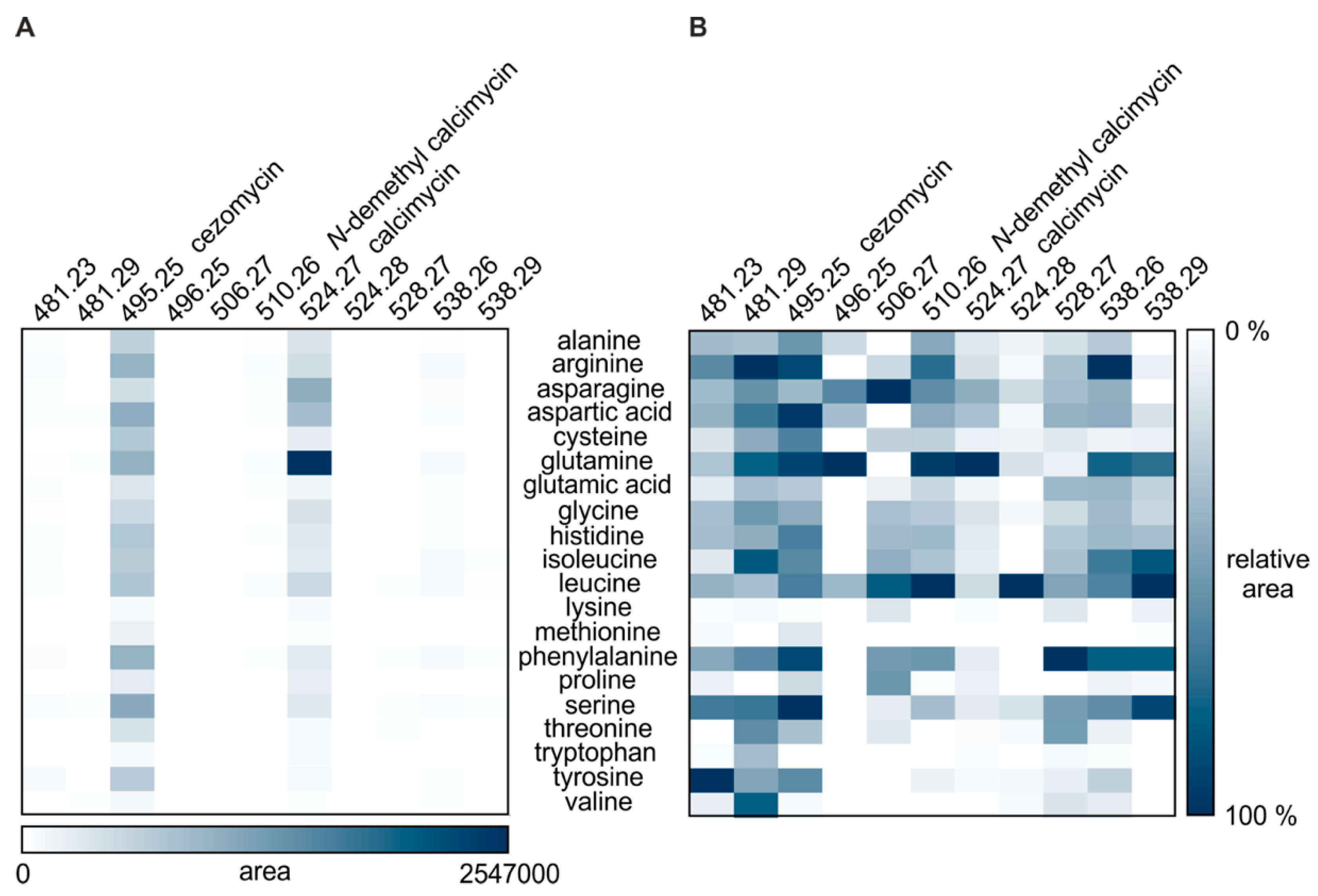
3.3. The Presence of Canonical Amino Acids Results in the Production of New Analogs
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boot, J.H.; van Hilten, J.A. The use of the divalent calcium-ionophore A23187 as a biochemical tool in pharmacological and in vitro toxicological studies. Cell Struct. Funct. 1996, 21, 97–99. [Google Scholar]
- Wang, X.; Yamamoto, T.; Kadowaki, M.; Yang, Y. Identification of key pathways and gene expression in the activation of mast cells via calcium flux using bioinformatics analysis. Biocell 2021, 45, 395–415. [Google Scholar] [CrossRef]
- Hara, H.; Kanazawa, T. Selective inhibition by ionophore A23187 of the enzyme isomerization in the catalytic cycle of sarcoplasmic reticulum Ca2+—ATPase. J. Biol. Chem. 1986, 261, 16584–16590. [Google Scholar] [CrossRef]
- Andreo, C.S.; Vallejos, R.H. Uncoupling of photophosphorylation in spinach chloroplasts by the ionophorous antibiotic A23187. FEBS Lett. 1974, 46, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Kajitani, N.; Kobuchi, H.; Fujita, H.; Yano, H.; Fujiwara, T.; Yasuda, T.; Utsumi, K. Mechanism of A23187-induced apoptosis in HL-60 cells: Dependency on mitochondrial permeability transition but not on NADPH oxidase. Biosci. Biotechnol. Biochem. 2007, 71, 2701–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raatschen, N.; Wenzel, M.; Leichert, L.I.O.; Düchting, P.; Krämer, U.; Bandow, J.E. Extracting iron and manganese from bacteria with ionophores—A mechanism against competitors characterized by increased potency in environments low in micronutrients. Proteomics 2013, 13, 1358–1370. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liang, J.; Lin, S.; Zhou, X.; Bai, L.; Deng, Z.; Wang, Z. Characterization of the biosynthesis gene cluster for the pyrrole polyether antibiotic calcimycin (A23187) in Streptomyces chartreusis NRRL 3882. Antimicrob. Agents Chemother. 2011, 55, 974–982. [Google Scholar] [CrossRef] [Green Version]
- Zmijewski, M.J. Biosynthesis of antibiotic A23187. Incorporation of precursors into A23187. J. Antibiot. 1980, 33, 447–450. [Google Scholar] [CrossRef] [Green Version]
- David, L.; Emadzadeh, S. Biosynthesis of the ionophorus antibiotic A23187. J. Antibiot. 1982, 35, 1616–1617. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.; Gou, L.; Lin, S.; Liang, J.; Yin, J.; Zhou, X.; Bai, L.; An, D.; Deng, Z.; Wang, Z. Characterization of the N-methyltransferase CalM involved in calcimycin biosynthesis by Streptomyces chartreusis NRRL 3882. Biochimie 2013, 95, 1487–1493. [Google Scholar] [CrossRef]
- Senges, C.H.R.; Al-Dilaimi, A.; Marchbank, D.H.; Wibberg, D.; Winkler, A.; Haltli, B.; Nowrousian, M.; Kalinowski, J.; Kerr, R.G.; Bandow, J.E. The secreted metabolome of Streptomyces chartreusis and implications for bacterial chemistry. Proc. Natl. Acad. Sci. USA 2018, 115, 2490–2495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, L.; Wu, Q.; Lin, S.; Li, X.; Liang, J.; Zhou, X.; An, D.; Deng, Z.; Wang, Z. Mutasynthesis of pyrrole spiroketal compound using calcimycin 3-hydroxy anthranilic acid biosynthetic mutant. Appl. Microbiol. Biotechnol. 2013, 97, 8183–8191. [Google Scholar] [CrossRef]
- David, L.; Kergomard, A. Production by controlled biosynthesis of a novel ionophore antibiotic, cezomycin (demethylamino A23187). J. Antibiot. 1982, 35, 1409–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, G.D.; Duax, W.L. Crystal and molecular structure of the calcium ion complex of A23187. J. Am. Chem. Soc. 1976, 98, 1578–1580. [Google Scholar] [CrossRef]
- Albrecht-Gary, A.M.; Blanc, S.; David, L.; Jeminet, G. Closely related ionophores cezomycin and calcimycin (A 23187): Cooperative formation of the transporting species. Inorg. Chem. 1994, 33, 518–524. [Google Scholar] [CrossRef]
- Erdahl, W.L.; Chapman, C.J.; Wang, E.; Taylor, R.W.; Pfeiffer, D.R. Ionophore 4-BrA23187 Transports Zn2+ and Mn2+ with High Selectivity Over Ca2+. Biochemistry 1996, 35, 13817–13825. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big Effects from Small Changes: Possible Ways to Explore Nature’s Chemical Diversity. ChemBioChem 2002, 3, 619. [Google Scholar] [CrossRef]
- Tangerina, M.M.P.; Furtado, L.C.; Leite, V.M.B.; Bauermeister, A.; Velasco-Alzate, K.; Jimenez, P.C.; Garrido, L.M.; Padilla, G.; Lopes, N.P.; Costa-Lotufo, L.V.; et al. Metabolomic study of marine Streptomyces sp.: Secondary metabolites and the production of potential anticancer compounds. PLoS ONE 2020, 15, e0244385. [Google Scholar] [CrossRef] [PubMed]
- Bauermeister, A.; Pereira, F.; Grilo, I.R.; Godinho, C.C.; Paulino, M.; Almeida, V.; Gobbo-Neto, L.; Prieto-Davó, A.; Sobral, R.G.; Lopes, N.P.; et al. Intra-clade metabolomic profiling of MAR4 Streptomyces from the Macaronesia Atlantic region reveals a source of anti-biofilm metabolites. Environ. Microbiol. 2019, 21, 1099–1112. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Fang, Q.; Maglangit, F.; Wu, L.; Ebel, R.; Kyeremeh, K.; Andersen, J.H.; Annang, F.; Pérez-Moreno, G.; Reyes, F.; Deng, H. Signalling and Bioactive Metabolites from Streptomyces sp. RK44. Molecules 2020, 25, 460. [Google Scholar] [CrossRef] [Green Version]
- Machushynets, N.V.; Wu, C.; Elsayed, S.S.; Hankemeier, T.; van Wezel, G.P. Discovery of novel glycerolated quinazolinones from Streptomyces sp. MBT27. J. Ind. Microbiol. Biotechnol. 2019, 46, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Zhang, Y.X.; Hutchinson, C.R. Amino acid catabolism and antibiotic synthesis: Valine is a source of precursors for macrolide biosynthesis in Streptomyces ambofaciens and Streptomyces fradiae. J. Appl. Bacteriol. 1994, 176, 6107–6119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, J.; Liras, P.; Martin, J.F. Utilization of ornithine and arginine as specific precursors of clavulanic acid. Appl. Environ. Microbiol. 1986, 52, 892–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luengo, J.M.; Revilla, G.; Villanueva, J.R.; Martin, J.F. Lysine regulation of penicillin biosynthesis in low-producing and industrial strains of Penicillium chrysogenum. Microbiology 1979, 115, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Masurekar, P.S.; Demain, A.L. Lysine control of penicillin biosynthesis. Can. J. Microbiol. 1972, 18, 1045–1048. [Google Scholar] [CrossRef]
- Pavlikova, M.; Kamenik, Z.; Janata, J.; Kadlcik, S.; Kuzma, M.; Najmanova, L. Novel pathway of 3-hydroxyanthranilic acid formation in limazepine biosynthesis reveals evolutionary relation between phenazines and pyrrolobenzodiazepines. Sci. Rep. 2018, 8, 7810. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R. The shikimate pathway—A metabolic tree with many branches. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 307–384. [Google Scholar] [CrossRef]
- Dewick, P.M. The biosynthesis of shikimate metabolites. Nat. Prod. Rep. 1989, 6, 263–290. [Google Scholar] [CrossRef]
- Walker, G.E.; Dubar, B.; Hunter, I.S.; Nimmo, H.G.; Cogginy, J.R. Evidence for a novel class of microbial 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase in Streptomyces coelicolor A3(2), Streptomyces rimosus and Neurospora crassa. Microbiology 1996, 142, 1973–1982. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.D.; Doy, C.H. End-product regulation of the general aromatic pathway in Escherichia coli W. Biochim. Biophys. Acta 1963, 3, 170–172. [Google Scholar] [CrossRef]
- Smith, L.C.; Ravel, J.M.; Lax, S.R.; Shiver, W. The control of 3-deoxy-D-arabino-heptulosonic acid 7-phosphate synthesis by phenylalanine and tyrosine. J. Biol. Chem. 1962, 237, 3566–3570. [Google Scholar] [CrossRef]
- Losada, A.A.; Cano-Prieto, C.; García-Salcedo, R.; Braña, A.F.; Méndez, C.; Salas, J.A.; Olano, C. Caboxamycin biosynthesis pathway and identification of novel benzoxazoles produced by cross-talk in Streptomyces sp. NTK 937. Microb. Biotechnol. 2017, 10, 873–885. [Google Scholar] [CrossRef]
- Wu, H.; Liang, J.; Gou, L.; Wu, Q.; Liang, W.J.; Zhou, X.; Bruce, I.J.; Deng, Z.; Wang, Z. Recycling of Overactivated Acyls by a Type II Thioesterase during Calcimycin Biosynthesis in Streptomyces chartreusis NRRL 3882. Appl. Environ. Microbiol. 2018, 84, e00587–e18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, M.C.; van der Donk, W.A. The many roles of glutamate in metabolism. J. Ind. Microbiol. Biotechnol. 2016, 43, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, C.; Lu, F.; He, Y.; Han, Z.; Du, L. Regulation of avilamycin biosynthesis in Streptomyces viridochromogenes: Effects of glucose, ammonium ion, and inorganic phosphate. Appl. Microbiol. Biotechnol. 2007, 73, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- Lebrihi, A.; Lamsaif, D.; Lefebvre, G.; Germain, P. Effect of ammonium ions on spiramycin biosynthesis in Streptomyces ambofaciens. Appl. Microbiol. Biotechnol. 1992, 37, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Khaoua, S.; Lebrihi, A.; Germain, P.; Lefebvre, G. Cephamycin C biosynthesis in Streptomyces cattleya: Nitrogen source regulation. Appl. Mircobiol. Biotechnol. 1991, 35, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Huh, J.H.; Yang, Y.Y.; Kang, C.M.; Lee, I.H.; Hyun, C.G.; Hong, S.K.; Suh, J.W. Accumulation of S-adenosyl-L-methionine enhances production of actinorhodin but inhibits sporulation in Streptomyces lividans TK23. J. Bacteriol. 2003, 185, 592–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huh, J.H.; Kim, D.J.; Zhao, X.Q.; Li, M.; Jo, Y.Y.; Yoon, T.M.; Shin, S.K.; Yong, J.H.; Ryu, Y.W.; Yang, Y.Y.; et al. Widespread activation of antibiotic biosynthesis by S-adenosylmethionine in streptomycetes. FEMS Microbiol. Lett. 2004, 238, 439–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontecave, M.; Atta, M.; Mulliez, E. S-adenosylmethionine: Nothing goes to waste. Trends Biochem. Sci. 2004, 29, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Li, S.M.; Westrich, L.; Schmidt, J.; Kuhnt, C.; Heide, L. Methyltransferase genes in Streptomyces rishiriensis: New coumermycin derivatives from gene-inactivation experiments. Microbiology 2002, 148, 317–3326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inahashi, Y.; Zhou, S.; Bibb, M.J.; Song, L.; Al-Bassam, M.M.; Bibb, M.J.; Challis, G.L. Watasemycin biosynthesis in Streptomyces venezuelae: Thiazoline C-methylation by a type B radical-SAM methylase homologue. Chem. Sci. 2017, 8, 2823–2831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Time [Min] | % H2O with 0.1% FA | % ACN with 0.1% FA |
|---|---|---|
| 0 | 95 | 5 |
| 2 | 95 | 5 |
| 21 | 0.5 | 99.5 |
| 23 | 0.5 | 99.5 |
| 28 | 95 | 5 |
| 30 | 95 | 5 |
| Measured Mass [M + H]+/ Δppm 1 | RT [min]/SD 2 | Fragment Spectrum 3 |
|---|---|---|
| 481.2369/ - | 21.39/ 0.19 |  |
| 481.2950/ - | 19.19/ 3.10 |  |
| Cezomycin 495.2509/ 2.827 | 22.83/ 0.18 |  |
| 496.2529/ - | 22.52/ 0.16 |  |
| 506.2727/ - | 24.57/ 0.05 |  |
| N-demethyl- calcimycin 510.2662/ 11.367 | 23.43/ 0.19 |  |
| calcimycin 524.2776/ 2.861 | 24.32/ 0.50 |  |
| 524.2822/ - | 23.51/ 0.03 |  |
| 528.2720/ - | 19.46/ 0.08 |  |
| 538.2606/ - | 23.13/ 0.04 |  |
| 538.2975/ - | 24.30/ 0.03 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arend, K.I.; Bandow, J.E. Influence of Amino Acid Feeding on Production of Calcimycin and Analogs in Streptomyces chartreusis. Int. J. Environ. Res. Public Health 2021, 18, 8740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168740
Arend KI, Bandow JE. Influence of Amino Acid Feeding on Production of Calcimycin and Analogs in Streptomyces chartreusis. International Journal of Environmental Research and Public Health. 2021; 18(16):8740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168740
Chicago/Turabian StyleArend, Kirstin I., and Julia E. Bandow. 2021. "Influence of Amino Acid Feeding on Production of Calcimycin and Analogs in Streptomyces chartreusis" International Journal of Environmental Research and Public Health 18, no. 16: 8740. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph18168740