
In Vitro and In Vivo Models to Assess the Immune-Related Effects of Nanomaterials



Abstract
:1. Introduction: Why Nanoimmunosafety Is Important
1.1. Nanotechnology and the Use of Engineered Nanoparticles
1.2. Immunological Safety: How the Immune System Works
1.3. Immunological Safety of ENPs: Need of Relevant Experimental Models
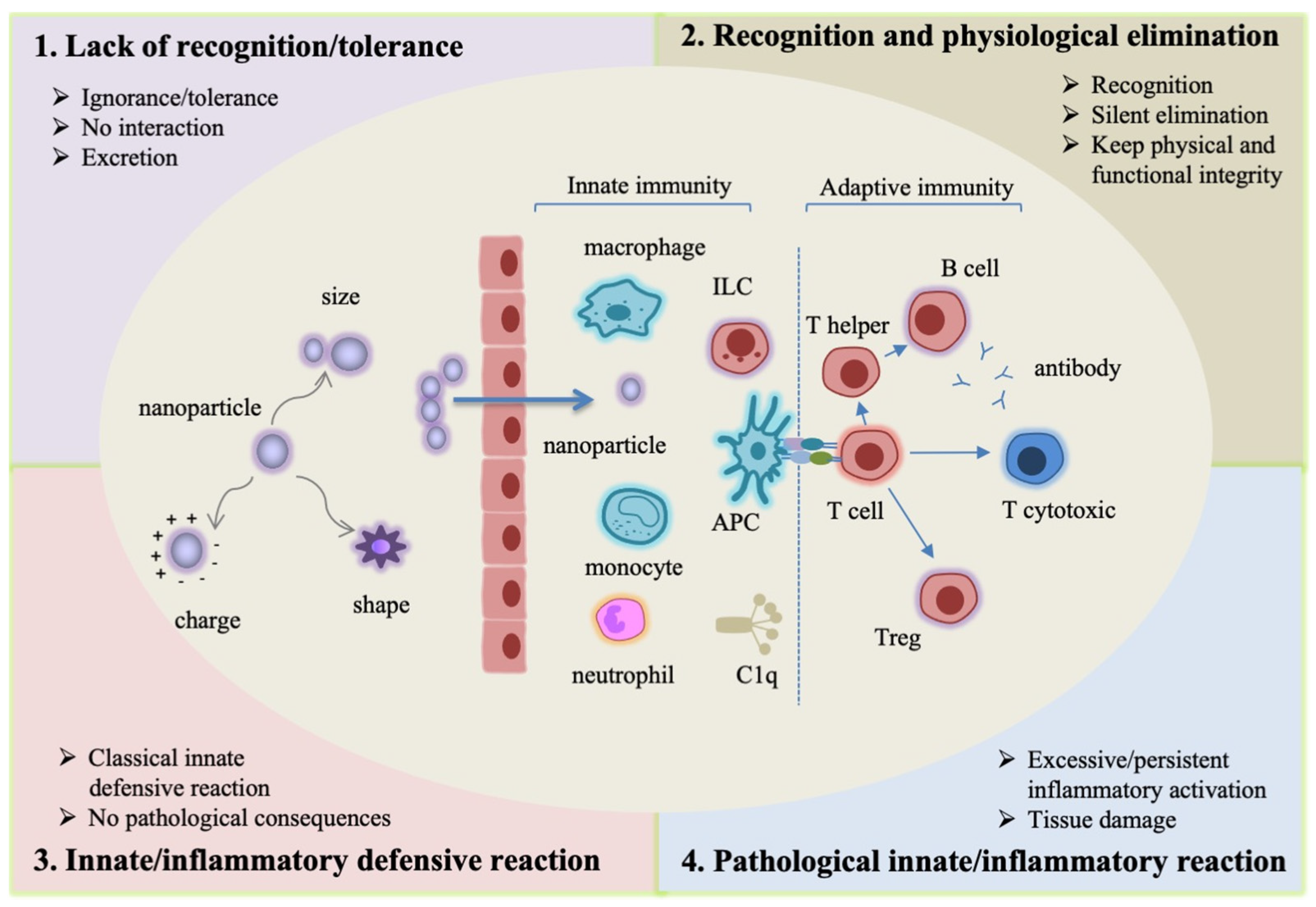
2. Immune Recognition and Response: Distinguishing between the “Normal” Defensive Response to Nanomaterials and Its Pathological/Damaging Dysregulation
- Lack of recognition/tolerance. This case encompasses two different events: ignorance (the immune system does not perceive the nanomaterial) and tolerance (the nanomaterial is detected by the immune system but not considered as a danger, and therefore does not trigger any reaction). Many ENMs fall into these two categories, and are eliminated as such through renal filtration and excretion with the urine and faeces [33,34,35,36]. Both particles <6 nm [33] and larger particles [34,36] can be excreted by renal filtration without causing a reaction. This kind of non-interaction results in rapid elimination without consequences for the organism.
- Recognition and physiological elimination. As for many particulate matters, the immune system can recognize an ENM as a potential danger and start an elimination process. Elimination mostly occurs by the action of the mononuclear phagocyte system (MPS), which encompasses phagocytes, such as macrophages, which have the specific role of engulfing particles and fragments of damaged tissue (dying cells, misfolded proteins) and degrading them into phagolysosomes, with the final goal of maintaining the tissue’s physical and functional integrity. This mechanism of silent/physiological elimination occurs constantly in all tissues. This process is physiological and causes no consequences to the body [33,37,38,39].
- Innate/inflammatory defensive reaction. This is a classical innate defensive reaction, when the innate immune system perceives an ENM as a potential danger that needs a powerful reaction to be eliminated. Most exogenous agents, such as microorganisms, trigger this kind of reaction, which involves several types of innate cells and soluble mediators. The ordered structure of nanomaterial surfaces resembles that of microorganisms, thereby facilitating their recognition by innate molecules (such as the complement component C1q) and receptors (such as the Toll-like receptors, TLR, and the scavenger receptors) and the consequent activation of an inflammatory reaction. When the triggering agent is successfully eliminated, the reaction ends with a mechanism of resolution and subsequent repair. In fact, as already mentioned, an innate/inflammatory reaction is a powerful non-specific response that not only targets the dangerous agent, but that can also damage the surrounding tissue. Thus, once the triggering agent is eliminated, the same innate cells involved in the defensive reaction (such as macrophages) are functionally redirected into anti-inflammation and tissue repair [32]. An innate/inflammatory reaction is not a pathological reaction, because its scope is the elimination of the dangerous agent and the re-establishment of tissue integrity, although it can cause transient damage and the death of a number of cells (both immune and bystander cells) [25,40,41,42,43].
- 4.
- Pathological innate/inflammatory reaction. In rare cases, an innate/inflammatory reaction may reach excessive levels or fail to resolve, thereby causing severe or permanent damage to the organism. This is the case of some microorganisms that can survive within macrophages [44] and, in the case of ENMs, it can happen with indigestible or toxic materials or with high aspect ratio particles that cannot be engulfed by phagocytes. In these cases, a significant cell death takes place, and the reaction becomes persistent, with the formation of new non-functional “scarring” tissue (as in the case of fibromas and granulomas) and the consequent impairment of tissue functionality [45,46,47,48]. Another circumstance that can lead to pathological inflammation is a chronic exposure, which may result in a persistent immune challenge and inflammatory activation, with a risk of chronic inflammation and consequent persistent tissue functional damage [49,50,51].
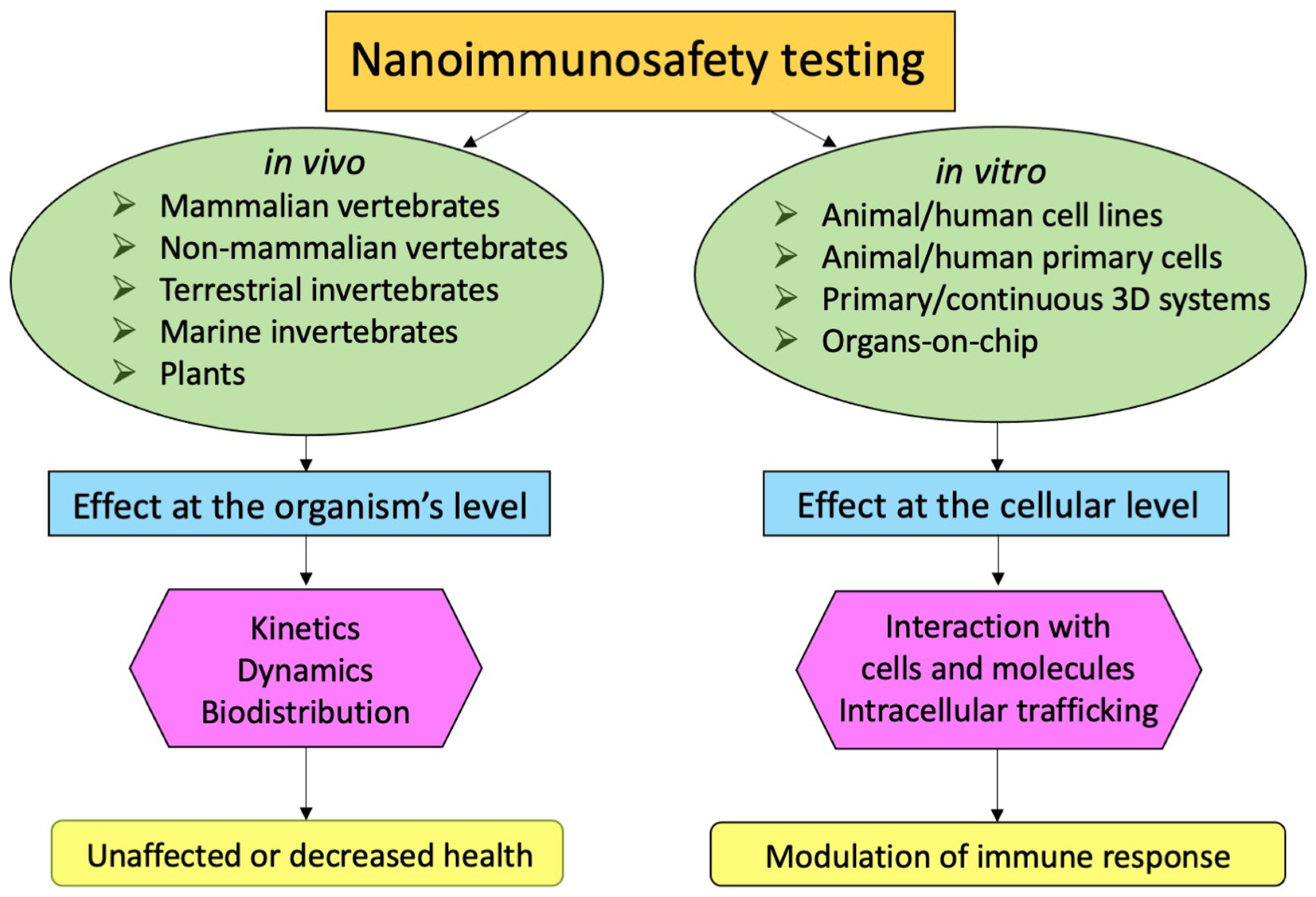
3. In Vivo Immunosafety Models, from Plants to Mammals
3.1. Selecting the In Vivo Nanoimmunosafety Model
3.2. Drawbacks and Limitations of In Vivo Models
3.3. Common Immunological Features across Living Species
4. In Vitro Immunosafety Models: Cell Lines and Primary Cells
4.1. Transformed Cell Lines
4.2. Primary Cells
4.3. Good In Vitro Method Practices and Immunoactive Contaminations
5. Conclusions and Future Perspectives: Addressing Biological Complexity for a More Realistic Nanoimmunosafety Assessment
Author Contributions
Funding
Conflicts of Interest
References
- Piccinno, F.; Gottschalk, F.; Seeger, S.; Nowack, B. Industrial production quantities and uses of ten engineered nanomaterials in Europe and the world. J. Nanoparticle Res. 2012, 14, 1109. [Google Scholar] [CrossRef] [Green Version]
- Mohajerani, A.; Burnett, L.; Smith, J.V.; Kurmus, H.; Milas, J.; Arulrajah, A.; Horpibulsuk, S.; Abdul Kadir, A. Nanoparticles in construction materials and other applications, and implications of nanoparticle use. Materials 2019, 12, 3052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Magaye, R.; Castranova, V.; Zhao, J. Titanium dioxide nanoparticles: A review of current toxicological data. Part. Fibre Toxicol. 2013, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolkova, B.; El Yamani, N.; Collins, A.R.; Gutleb, A.C.; Dusinska, M. Nanoparticles in food. Epigenetic changes induced by nanomaterials and possible impact on health. Food Chem. Toxicol. 2015, 77, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Xu, K.; Taratula, O.; Farsad, K. Applications of nanoparticles in biomedical imaging. Nanoscale 2019, 11, 799–819. [Google Scholar] [CrossRef] [PubMed]
- Nune, S.K.; Gunda, P.; Thallapally, P.K.; Lin, Y.-Y.; Forrest, M.L.; Berkland, C.J. Nanoparticles for biomedical imaging. Expert Opin. Drug Deliv. 2009, 6, 1175–1194. [Google Scholar] [CrossRef] [Green Version]
- Dadfar, S.M.; Roemhild, K.; Drude, N.I.; von Stillfried, S.; Knüchel, R.; Kiessling, F.; Lammers, T. Iron oxide nanoparticles: Diagnostic, therapeutic and theranostic applications. Adv. Drug Deliv. Rev. 2019, 138, 302–325. [Google Scholar] [CrossRef] [PubMed]
- Rosen, J.E.; Yoffe, S.; Meerasa, A.; Verma, M.; Gu, F.X. Nanotechnology and diagnostic imaging: New advances in contrast agent technology. J. Nanomed. Nanotechnol. 2011, 2, 1000115. [Google Scholar] [CrossRef]
- Lu, M.; Cohen, M.H.; Rieves, D.; Pazdur, R. FDA report: Ferumoxytol for intravenous iron therapy in adult patients with chronic kidney disease. Am. J. Hematol. 2010, 85, 315–319. [Google Scholar] [CrossRef] [PubMed]
- Thiesen, B.; Jordan, A. Clinical applications of magnetic nanoparticles for hyperthermia. Int. J. Hyperth. 2008, 24, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Dulinska-Litewka, J.; Lazarczyk, A.; Halubiec, P.; Szafranski, O.; Karnas, K.; Karewicz, A. Superparamagnetic iron oxide nanoparticles — Current and prospective medical applications. Materials 2019, 12, 617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soetaert, F.; Korangath, P.; Serantes, D.; Fiering, S.; Ivkov, R. Cancer therapy with iron oxide nanoparticles: Agents of thermal and immune therapies. Adv. Drug Deliv. Rev. 2020, 163, 65–83. [Google Scholar] [CrossRef]
- Rezvantalab, S.; Drude, N.I.; Moraveji, M.K.; Guvener, N.; Koons, E.K.; Shi, Y.; Lammers, T.; Kiessling, F. PLGA-based nanoparticles in cancer treatment. Front. Pharmacol. 2018, 9, 1260. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.; Jain, D.S. Poly Lactic-Co-Glycolic Acid (PLGA) copolymer and its pharmaceutical application. Handb. Polym. Pharm. Technol. 2015, 2, 151–172. [Google Scholar] [CrossRef]
- Danhier, F.; Ansorena, E.; Silva, J.M.; Coco, R.; Le Breton, A.; Preat, V. PLGA-based nanoparticles: An overview of biomedical applications. J. Control. Release 2012, 161, 505–522. [Google Scholar] [CrossRef] [PubMed]
- Tran, S.; DeGiovanni, P.J.; Piel, B.; Rai, P. Cancer nanomedicine: A review of recent success in drug delivery. Clin. Transl. Med. 2017, 6, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, X.; Zhou, M.; Yu, S.; Jin, Z.; Zhao, K. An overview of biodegradable nanomaterials and applications in vaccines. Vaccine 2020, 38, 1096–1104. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Italiani, P. From antigen delivery system to adjuvanticy: The board application of nanoparticles in vaccinology. Vaccines 2015, 3, 930–939. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.; Duschl, A.; Himly, M. Nanotechnology-based vaccines for allergen-specific immunotherapy: Potentials and challenges of conventional and novel adjuvants under research. Vaccines 2020, 8, 237. [Google Scholar] [CrossRef] [PubMed]
- Oberdörster, G.; Oberdörster, E.; Oberdörster, J. Nanotoxicology: An emerging discipline evolving from studies of ultrafine particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Madler, L.; Li, N. Toxic potential of materials at the nanolevel. Science 2006, 311, 622–627. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Leong, D.T.; Lynch, I. Rethinking nanosafety: Harnessing progress and driving innovation. Small 2020, 16, e2002503. [Google Scholar] [CrossRef] [PubMed]
- Dobrovolskaia, M.A.; McNeil, S.E. Immunological properties of engineered nanomaterials. Nat. Nanotechnol. 2007, 2, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Duschl, A. Nanoparticles and the Immune System: Safety and Effects; Elsevier—Academic Press: Oxford, UK, 2014; p. 138. [Google Scholar]
- Murphy, K.M.; Weaver, C.; Berg, L.J. Janeway’s Immunobiology, 10th ed.; W.W. Norton & Company, Inc.: New York, NY, USA, 2022. [Google Scholar]
- Boraschi, D.; Alijagic, A.; Auguste, M.; Barbero, F.; Ferrari, E.; Hernadi, S.; Mayall, C.; Michelini, S.; Navarro Pacheco, N.I.; Prinelli, A.; et al. Addressing nanomaterial immunosafety by evaluating innate immunity across living species. Small 2020, 16, 2000598. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Fadeel, B.; Duschl, A. Immune system. In Adverse Effects of Engineered Nanomaterials. Exposure, Toxicology and Impact on Human Health, 2nd ed.; Fadeel, B., Pietroiusti, A., Shvedova, A., Eds.; Elsevier: Amsterdam, The Netherlands; San Diego, CA, USA, 2017; pp. 313–337. [Google Scholar]
- Fadeel, B. Hide and seek: Nanomaterial interactions with the immune system. Front. Immunol. 2019, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Pinsino, A.; Bastús, N.G.; Busquets-Fité, M.; Canesi, L.; Cesaroni, P.; Drobne, D.; Duschl, A.; Ewart, M.-A.; Gispert, I.; Horejs-Höck, J.; et al. Probing the immunological responses to nanoparticles across environmental species: A perspective of the EU-funded PANDORA project. Environ. Sci. Nano 2020, 7, 3216–3232. [Google Scholar] [CrossRef]
- Cronin, J.G.; Jones, N.; Thornton, C.A.; Jenkins, G.J.S.; Doak, S.H.; Clift, M.J.D. Nanomaterials and innate immunity: A perspective of the current status in nanosafety. Chem. Res. Toxicol. 2020, 33, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Italiani, P.; Boraschi, D. From monocytes to M1/M2 macrophages: Phenotypical vs. functional differentiation. Front. Immunol. 2014, 5, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, H.-J. Excretion and Clearance. In Radionanomedicine. Combined Nuclear and Nanomedicine; Biological and Medical Physics, Biomedical Engineering; Lee, D.S., Ed.; Springer International Publishing AG: Cham, Switzerland, 2018; pp. 347–368. [Google Scholar] [CrossRef]
- He, X.; Nie, H.; Wang, K.; Tan, W.; Wu, X.; Zhang, P. In vivo study of biodistribution and urinary excretion of surface-modified silica nanoparticles. Anal. Chem. 2008, 80, 9597–9603. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, L.; Liu, T.; Hao, N.; Liu, H.; Chen, D.; Tang, F. The shape effect of mesoporous silica nanoparticles on biodistribution, clearance, and biocompatibility in vivo. ACS Nano 2011, 5, 5390–5399. [Google Scholar] [CrossRef]
- Ferretti, A.M.; Usseglio, S.; Mondini, S.; Drago, C.; La Mattina, R.; Chini, B.; Verderio, C.; Leonzino, M.; Cagnoli, C.; Joshi, P.; et al. Towards bio-compatible magnetic nanoparticles: Immune-related effects, in vitro internalization, and in vivo bio-distribution of zwitterionic ferrite nanoparticles with unexpected renal clearance. J. Colloid Interface Sci. 2021, 582, 678–700. [Google Scholar] [CrossRef]
- Kumar, R.; Roy, I.; Ohulchanskky, T.Y.; Vathy, L.A.; Bergey, E.J.; Sajjad, M.; Prasad, P.N. In vivo biodistribution and clearance studies using multimodal organically modified silica nanoparticles. ACS Nano 2010, 4, 699–708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Li, L.; Teng, X.; Huang, X.; Liu, H.; Chen, D.; Ren, J.; He, J.; Tang, F. Single and repeated dose toxicity of mesoporous hollow silica nanoparticles in intravenously exposed mice. Biomaterials 2011, 32, 1657–1668. [Google Scholar] [CrossRef] [PubMed]
- Shao, D.; Lu, M.-M.; Zhao, Y.-W.; Zhang, F.; Tan, Y.-F.; Zheng, X.; Pan, Y.; Xiao, X.-A.; Wang, Z.; Dong, W.-F.; et al. The shape effect of magnetic mesoporous silica nanoparticles on endocytosis, biocompatibility and biodistribution. Acta Biomater. 2017, 49, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Gurtner, G.C.; Werner, S.; Barrandon, Y.; Longaker, M.T. Wound repair and regeneration. Nature 2008, 453, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Cho, W.S.; Han, B.S.; Cho, M.; Kim, S.Y.; Yi, J.Y.; Ahn, B.; Kim, S.H.; Jeong, J. Transient pulmonary fibrogenic effect induced by intratracheal instillation of ultrafine amorphous silica in A/J mice. Toxicol. Lett. 2008, 182, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nature 2016, 529, 307–315. [Google Scholar] [CrossRef] [PubMed]
- May, S.; Hirsch, C.; Rippl, A.; Bohmer, N.; Kaiser, J.-P.; Diener, L.; Wichser, A.; Bürkle, A.; Wick, P. Transient DNA damage following exposure to gold nanoparticles. Nanoscale 2018, 10, 15723–15735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Co, D.O.; Hogan, L.H.; Kim, S.-I.; Sandor, M. Mycobacterial granulomas: Keys to a long-lasting host-pathogen relationship. Clin. Immunol. 2004, 113, 130–136. [Google Scholar] [CrossRef]
- Porter, D.W.; Wu, N.; Hubbs, A.F.; Mercer, R.R.; Funk, K.; Meng, F.; Li, J.; Wolfarth, M.G.; Battelli, L.; Friend, S.; et al. Differential mouse pulmonary dose and time course responses to titanium dioxide nanospheres and nanobelts. Toxicol. Sci. 2013, 131, 179–193. [Google Scholar] [CrossRef] [Green Version]
- van den Brule, S.; Beckers, E.; Chaurand, P.; Liu, W.; Ibouraadaten, S.; Palmai-Pallag, M.; Uwambayinema, F.; Yakoub, Y.; Avellan, A.; Levard, C.; et al. Nanometer-long Ge-imogolite nanotubes cause sustained lung inflammation and fibrosis in rats. Part. Fibre Toxicol. 2014, 11, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.T.; Wu, C.T.; Huang, K.H.; Lin, W.C.; Chen, C.M.; Guan, S.S.; Chiang, C.K.; Liu, S.H. Titanium nanoparticle inhalation induces renal fibrosis in mice via an oxidative stress upregulated transforming growth factor-β pathway. Chem. Res. Toxicol. 2015, 28, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Ma, Q. Integration of inflammation, fibrosis, and cancer induced by carbon nanotubes. Nanotoxicology 2019, 13, 1244–1274. [Google Scholar] [CrossRef] [PubMed]
- Riebling, C.; Luch, A.; Götz, M.E. Comparative modeling of exposure to airborne nanoparticles released by consumer spray products. Nanotoxicology 2016, 10, 343–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadpour, R.; Dobrovolskaia, M.A.; Cheney, D.L.; Greish, K.F.; Ghandehari, H. Subchronic and chronic toxicity evaluation of inorganic nanoparticles for delivery applications. Adv. Drug Deliv. Rev. 2019, 144, 112–132. [Google Scholar] [CrossRef] [PubMed]
- Mohammadpour, R.; Cheney, D.L.; Grunberger, J.W.; Yazdimamaghani, M.; Jedrzkiewicz, J.; Isaacson, K.J.; Dobrovolskaia, M.A.; Ghandehari, H. One-year chronic toxicity evaluation of single dose intravenously administered silica nanoparticles in mice and their ex vivo human hemocompatibility. J. Control. Release 2020, 324, 471–481. [Google Scholar] [CrossRef] [PubMed]
- Boraschi, D.; Costantino, L.; Italiani, P. Interaction of nanoparticles with immunocompetent cells: Nanosafety considerations. Nanomedicine (Future Med.) 2012, 7, 121–131. [Google Scholar] [CrossRef]
- Kusaka, T.; Nakayama, M.; Nakamura, K.; Ishimiya, M.; Furusawa, E.; Ogasawara, K. Effect of silica particle size on macrophage inflammatory responses. PLoS ONE 2014, 9, e92634. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.M.; Rosser, A.M. Humans: The ultimate animal models. J. Neurol. Neurosurg. Psychiatry 2020, 91, 1132–1136. [Google Scholar] [CrossRef]
- Pulendran, B.; Davis, M.M. The science and medicine of human immunology. Science 2020, 369, eaay4014. [Google Scholar] [CrossRef] [PubMed]
- Sydney Brenner Urges Cancer Researchers to Consider ‘Bedside to Bench’ Approach. 2008. Available online: www.genomeweb.com/archive/sydney-brenner-urges-cancer-researchers-consider-bedside-bench-approach#.XcnGQi2ZNlM (accessed on 24 August 2021).
- Szebeni, J.; Alving, C.R.; Rosivall, L.; Bünger, R.; Baranyi, L.; Bedöcs, P.; Tóth, M.; Barenholz, Y. Animal models of complement-mediated hypersensitivity reactions to liposomes and other lipid-based nanoparticles. J. Liposome Res. 2007, 17, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Carrion, R., Jr.; Patterson, J.L. An animal model that reflects human disease: The common marmoset (Callithrix jacchus). Curr. Opin. Virol. 2012, 2, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Barclay, W.; Barr, I.; Fouchier, R.A.M.; Matsuyama, R.; Nishiura, H.; Peiris, M.; Russell, C.J.; Subbarao, K.; Zhu, H.; et al. Ferrets as models for influenza virus transmission studies and pandemic risk assessments. Emerg. Infect. Dis. 2018, 24, 965–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wager, L.E.; DiFazio, R.M.; Davie, M.M. Advanced model systems and tools for basic and translational human immunology. Genome Med. 2018, 10, 73. [Google Scholar] [CrossRef] [PubMed]
- Graham, A.L. Naturalizing mouse models for immunology. Nat. Immunol. 2021, 22, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Trede, N.S.; Langenau, D.M.; Traver, D.; Look, A.T.; Zon, L. The use of zebrafish to understand immunity. Immunity 2004, 20, 367–379. [Google Scholar] [CrossRef] [Green Version]
- Novoa, B.; Figueras, A. Zebrafish: Model for the study of inflammation and the innate immune response to infectious diseases. In Current Topics in Innate Immunity II. Advances in Experimental Medicine and Biology; Lambris, J., Hajishengallis, G., Eds.; Springer: New York, NY, USA, 2012; Volume 946, pp. 253–275. [Google Scholar] [CrossRef] [Green Version]
- Ruyra, A.; Cano-Sarabia, M.; García-Valtanen, P.; Yero, D.; Gibert, I.; Mackenzie, S.A.; Estepa, A.; Maspoch, D.; Roher, N. Targeting and stimulation of the zebrafish (Danio rerio) innate immune system with LPS/dsRNA-loaded nanoliposomes. Vaccine 2014, 32, 3955–3962. [Google Scholar] [CrossRef] [PubMed]
- Limonta, G.; Mancia, A.; Benkhalqui, A.; Bertolucci, C.; Abelli, L.; Fossa, M.C.; Panti, C. Microplastics induce transcriptional changes, immune response and behavioural alterations in adult zebrafish. Sci. Rep. 2019, 9, 15775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pensado-López, A.; Fernández-Rey, J.; Reimunde, P.; Crecente-Campo, J.; Sánchez, L.; Torres Andón, F. Zebrafish models for the safety and therapeutic testing of nanoparticles with a focus on macrophages. Nanomaterials 2021, 11, 1784. [Google Scholar] [CrossRef]
- EFPIA-European Federation of Pharmaceutical Industries and Federations. Animal Welfare: 3Rs—Replace, Refine, Reduce. Available online: www.efpia.eu/documents/41/90/Animal-welfare-3R-39-s-Replace-Refine-Reduce (accessed on 20 August 2021).
- Mestas, J.; Hughes, C.C.W. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.M. A prescription for human immunology. Immunity 2008, 29, 835–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nold, M.F.; Nold-Petry, C.A.; Zepp, J.A.; Palmer, B.E.; Bufler, P.; Dinarello, C.A. IL-37 is a fundamental inhibitor of innate immunity. Nat. Immunol. 2010, 11, 1014–1022. [Google Scholar] [CrossRef] [Green Version]
- Boraschi, D.; Lucchesi, D.; Hainzl, S.; Leitner, M.; Maier, E.; Mangelberger, D.; Oostingh, G.J.; Pfaller, T.; Pixner, C.; Posselt, G.; et al. IL-37: An anti-inflammatory cytokine of the IL-1 family. Eur. Cytokine Netw. 2011, 22, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Patera, A.C.; Pong-Kennedy, A.; Deno, G.; Gonsiorek, W.; Manfra, D.J.; Vassileva, G.; Zeng, M.; Jackson, C.; Sullivan, L.; et al. Murine CXCR1 is a functional receptor for GCP-2/CXCL6 and interleukin-8/CXCL8. J. Biol. Chem. 2007, 282, 11658–11666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjornson, M.; Pimprikar, P.; Nürnberger, T.; Zipfel, C. The transcriptional landscape of Arabidopsis thaliana pattern-triggered immunity. Nat. Plants 2021, 7, 579–586. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, J.A. The immune response of Drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Rowley, A.F. The blood cells of the sea squirt, Ciona intestinalis: Morphology, differential counts, and in vitro phagocytic activity. J. Invertebr. Pathol. 1981, 37, 91–100. [Google Scholar] [CrossRef]
- Irmak, P.; Kurtz, J.; Zimmer, M. Immune response in Porcellio scaber (Isopoda: Oniscidea): Copper revisited. Eur. J. Soil Biol. 2005, 41, 77–83. [Google Scholar] [CrossRef]
- Dales, R.P.; Kalaç, Y. Phagocytic defence by the earthworm Eisenia foetida against certain pathogenic bacteria. Comp. Biochem. Physiol. A 1992, 101, 487–490. [Google Scholar] [CrossRef]
- Pinsino, A.; Matranga, V. Sea urchin immune cells as sentinels of environmental stress. Dev. Comp. Immunol. 2015, 49, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Alijagic, A.; Gaglio, D.; Napodano, E.; Russo, R.; Costa, C.; Benada, O.; Kofroňová, O.; Pinsino, A. Titanium dioxide nanoparticles temporarily influence the sea urchin immunological state suppressing inflammatory-relate gene transcription and boosting antioxidant metabolic activity. J. Hazard. Mater. 2020, 384, 121389. [Google Scholar] [CrossRef] [PubMed]
- Auguste, M.; Balbi, T.; Ciacci, C.; Canesi, L. Conservation od cell communication systems in invertebrates host-defence mechanisms: Possible role in immunity and diusease. Biology 2020, 9, 234. [Google Scholar] [CrossRef]
- Balbi, T.; Auguste, M.; Ciacci, C.; Canesi, L. Immunological responses of marine bivalves to contaminant exposure: Contribution of the -omics approach. Front. Immunol. 2021, 12, 618726. [Google Scholar] [CrossRef]
- Swartzwelter, B.J.; Mayall, C.; Alijagic, A.; Barbero, F.; Ferrari, E.; Hernadi, S.; Michelini, S.; Navarro Pacheco, N.I.; Prinelli, A.; Swart, E.; et al. Cross-species comparisons of nanoparticle interactions with innate immune systems: A methodological review. Nanomaterials 2021, 11, 1528. [Google Scholar] [CrossRef] [PubMed]
- Goodman, C.L.; Kang, D.S.; Stanley, D. Cell line platforms support research into arthropod immunity. Insects 2021, 12, 738. [Google Scholar] [CrossRef] [PubMed]
- Wolff, S.M. Biological effects of bacterial endotoxins in man. J. Infect. Dis. 1973, 128 (Suppl. S1), 259–264. [Google Scholar] [CrossRef]
- Copeland, S.; Warren, H.S.; Lowry, S.F.; Calvanom, S.E.; Remick, D. The Inflammation and the Host Response to Injury Investigators. Acute inflammatory response to endotoxin in mice and humans. Clin. Diagn. Lab. Immunol. 2005, 12, 60–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauwels, A.; Vandendriessche, B.; Brouckaert, P. Of mice, men, and inflammation. Proc. Natl. Acad. Sci. USA 2013, 110, E3150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ATCC Catalogue. Product ATCC® TIB-71TM. Available online: https://www.atcc.org/products/tib-71#detailed-product-information (accessed on 20 August 2021).
- Italiani, P.; Mazza, E.M.C.; Lucchesi, D.; Cifola, I.; Gemelli, C.; Grande, A.; Battaglia, C.; Bicciato, S.; Boraschi, D. Transcriptomic profiling of the development of the inflammatory response in human monocytes in vitro. PLoS ONE 2014, 9, e87680. [Google Scholar] [CrossRef]
- Della Camera, G.; Madej, M.; Ferretti, A.M.; La Spina, R.; Li, Y.; Corteggio, A.; Heinzl, T.; Swartzwelter, B.J.; Sipos, G.; Gioria, S.; et al. Personalised profiling of innate immune memory induced by nano-imaging particles in human monocytes. Front. Immunol. 2021, 12, 692165. [Google Scholar] [CrossRef] [PubMed]
- Sestini, P.; Tagliabue, A.; Bartalini, M.; Boraschi, D. Asbestos induced modulation of release of regulatory molecules from alveolar and peritoneal macrophages. Chest 1986, 89, 161S–162S. [Google Scholar] [CrossRef]
- Leijh, P.C.; van Zwet, T.L.; ter Kuile, M.N.; van Furth, R. Effect of thioglycolate on phagocytic and microbicidal activities of peritoneal macrophages. Infect. Immun. 1984, 46, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Guidance Document on Good In Vitro Method Practices (GIVIMP). OECD Series on Testing and Assessment. Available online: https://www.oecd.org/env/guidance-document-on-good-in-vitro-method-practices-givimp-9789264304796-en.htm (accessed on 22 September 2021).
- Drexler, H.G.; Uphoff, C.C. Mycoplasma contamination of cell cultures: Incidence, sources, effects, detection, elimination, prevention. Cytotechnology 2002, 39, 75–90. [Google Scholar] [CrossRef]
- Gorbet, M.B.; Sefton, M.V. Endotoxin: The uninvited guest. Biomaterials 2005, 26, 6811–6817. [Google Scholar] [CrossRef]
- Li, Y.; Fujita, M.; Boraschi, D. Endotoxin contamination in nanomaterials leads to the misinterpretation of immunosafety results. Front. Immunol. 2017, 8, 472. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shi, Z.; Radauer-Preiml, I.; Andosch, A.; Casals, E.; Luetz-Meindl, U.; Cobaleda, M.; Lin, Z.; Jaberi-Douraki, M.; Italiani, P.; et al. Bacterial endotoxin (lipopolysaccharide) binds to the surface of gold nanoparticles, interferes with biocorona formation and induces human monocyte inflammatory activation. Nanotoxicology 2017, 11, 1157–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Italiani, P.; Casals, E.; Valkenborg, D.; Mertens, I.; Baggerman, G.; Nelissen, I.; Puntes, V.; Boraschi, D. Assessing the immunosafety of engineered nanoparticles with a novel in vitro model based on human primary monocytes. ACS Appl. Mater. Interfaces 2016, 8, 28437–28447. [Google Scholar] [CrossRef] [PubMed]
- Swartzwelter, B.J.; Verde, A.; Rehak, L.; Madej, M.; Puntes, V.F.; De Luca, A.C.; Boraschi, D.; Italiani, P. Interaction between macrophages and nanoparticles: In vitro 3D cultures for the realistic assessment of inflammatory activation and modulation of innate memory. Nanomaterials 2021, 11, 207. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Moeller, J.; Vogel, V. Mechanobiology of macrophages: How physical factors coregulate macrophage plasticity and phagocytosis. Annu. Rev. Biomed. Eng. 2019, 21, 267–297. [Google Scholar] [CrossRef] [PubMed]
- Fizesan, I.; Cambier, S.; Moschini, E.; Chary, A.; Nelissen, I.; Ziebel, J.; Audinot, J.-N.; Wirtz, T.; Kruszewski, M.; Pop, A.; et al. In vitro exposure of a 3D-tetraculture representative for the alveolar barrier at the air-liquid interface to silver particles and nanowires. Part. Fibre Toxicol. 2019, 16, 14. [Google Scholar] [CrossRef]
- Kulthong, K.; Duivenvoorde, L.; Sun, H.; Confederat, S.; Wu, J.; Spenkelink, B.; de Haan, L.; Marin, V.; van der Zande, M.; Bouwmeester, H. Microfluidic chip for culturing intestinal epithelial cell layers: Characterization and comparison of drug transport between dynamic and static models. Toxicol. In Vitro 2020, 65, 104815. [Google Scholar] [CrossRef]
- Jalili-Firoozinezhad, S.; Gazzaniga, F.S.; Calamari, E.L.; Camacho, D.M.; Fadel, C.W.; Bein, A.; Swenor, B.; Nestor, B.; Cronce, M.J.; Tovaglieri, A.; et al. A complex human gut microbiome cultured in an anaerobic intestine-on-a-chip. Nat. Biomed. Eng. 2019, 3, 520–531. [Google Scholar] [CrossRef]
- Tang, M.; Li, S.; Wei, L.; Hou, Z.; Qu, J.; Li, L. Do engineered nanomaterials affect immune responses by interacting with gut microbiota? Front. Immunol. 2021, 12, 684605. [Google Scholar] [CrossRef]
- Tlaskalová-Hogenová, H.; Stěpánková, R.; Kozáková, H.; Hudcovic, T.; Vannucci, L.; Tučková, L.; Rossmann, P.; Hrnčíř, T.; Kverka, M.; Zákostelská, Z.; et al. The role of gut microbiota (commensal bacteria) and the mucosal barrier in the pathogenesis of inflammatory and autoimmune diseases and cancer: Contribution of germ-free and gnotobiotic animal models of human diseases. Cell. Mol. Immunol. 2011, 8, 110–120. [Google Scholar] [CrossRef]
- Webster, J.I.; Tonelli, L.; Sternberg, E.M. Neuroendocrine regulation of immunity. Annu. Rev. Immunol. 2002, 20, 125–163. [Google Scholar] [CrossRef] [Green Version]
- Di Cosmo, A.; Polese, G. Neuroendocrine–immune systems response to environmental stressors in the cephalopod Octopus vulgaris. Front. Physiol. 2016, 7, 434. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Innate Reaction | Cellular Level | Tissue/Organ Level | Organism Level |
|---|---|---|---|
| Ignorance/tolerance | no effect | no effect | no effect |
| Silent elimination | activation | no effect | no effect |
| autophagy, apoptosis | |||
| Resolving inflammation Strong reaction with eventual elimination | inflammatory death of immune and bystander cells | transient damage | no effect |
| Chronic inflammation Strong non-resolving reaction | continuous inflammatory death of immune and bystander cells | persistent damage, tissue destruction and neoformation | pathology |
| Models | Pros | Cons |
|---|---|---|
| in vivo |
|
|
| in vitro | Cell lines
Organs-on-chip
| Cell lines
Primary cells
Organs-on-chip
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boraschi, D.; Li, D.; Li, Y.; Italiani, P. In Vitro and In Vivo Models to Assess the Immune-Related Effects of Nanomaterials. Int. J. Environ. Res. Public Health 2021, 18, 11769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182211769
Boraschi D, Li D, Li Y, Italiani P. In Vitro and In Vivo Models to Assess the Immune-Related Effects of Nanomaterials. International Journal of Environmental Research and Public Health. 2021; 18(22):11769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182211769
Chicago/Turabian StyleBoraschi, Diana, Dongjie Li, Yang Li, and Paola Italiani. 2021. "In Vitro and In Vivo Models to Assess the Immune-Related Effects of Nanomaterials" International Journal of Environmental Research and Public Health 18, no. 22: 11769. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182211769