
Propofol and Sevoflurane Anesthesia in Early Childhood Do Not Influence Seizure Threshold in Adult Rats

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Exposition to Sevoflurane
2.3. Exposure to Propofol
2.4. Experimental Seizure Models
2.5. Statistical Analysis
3. Results
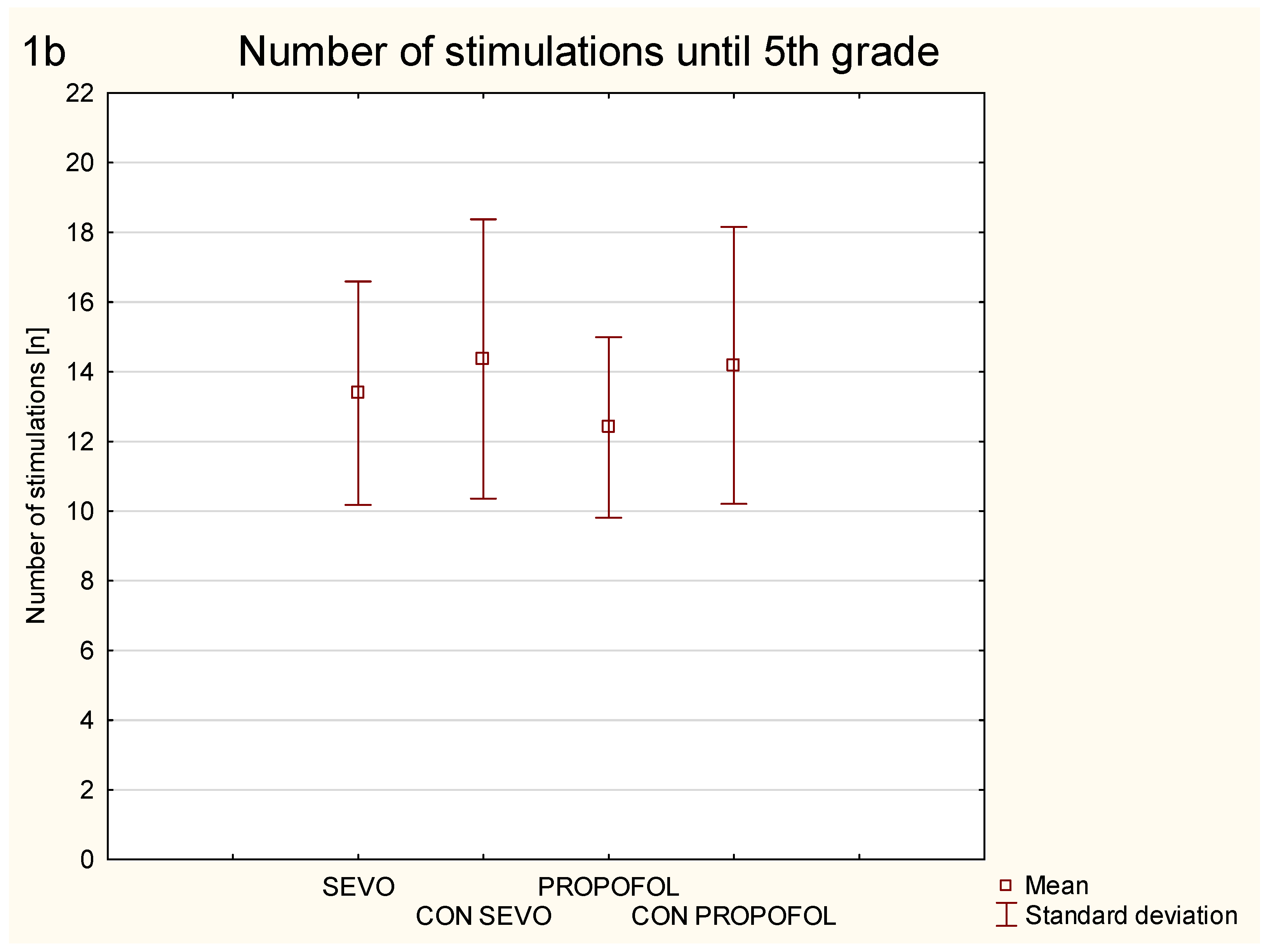
3.1. Corneal Kindling
3.2. Drug Induced Seizures
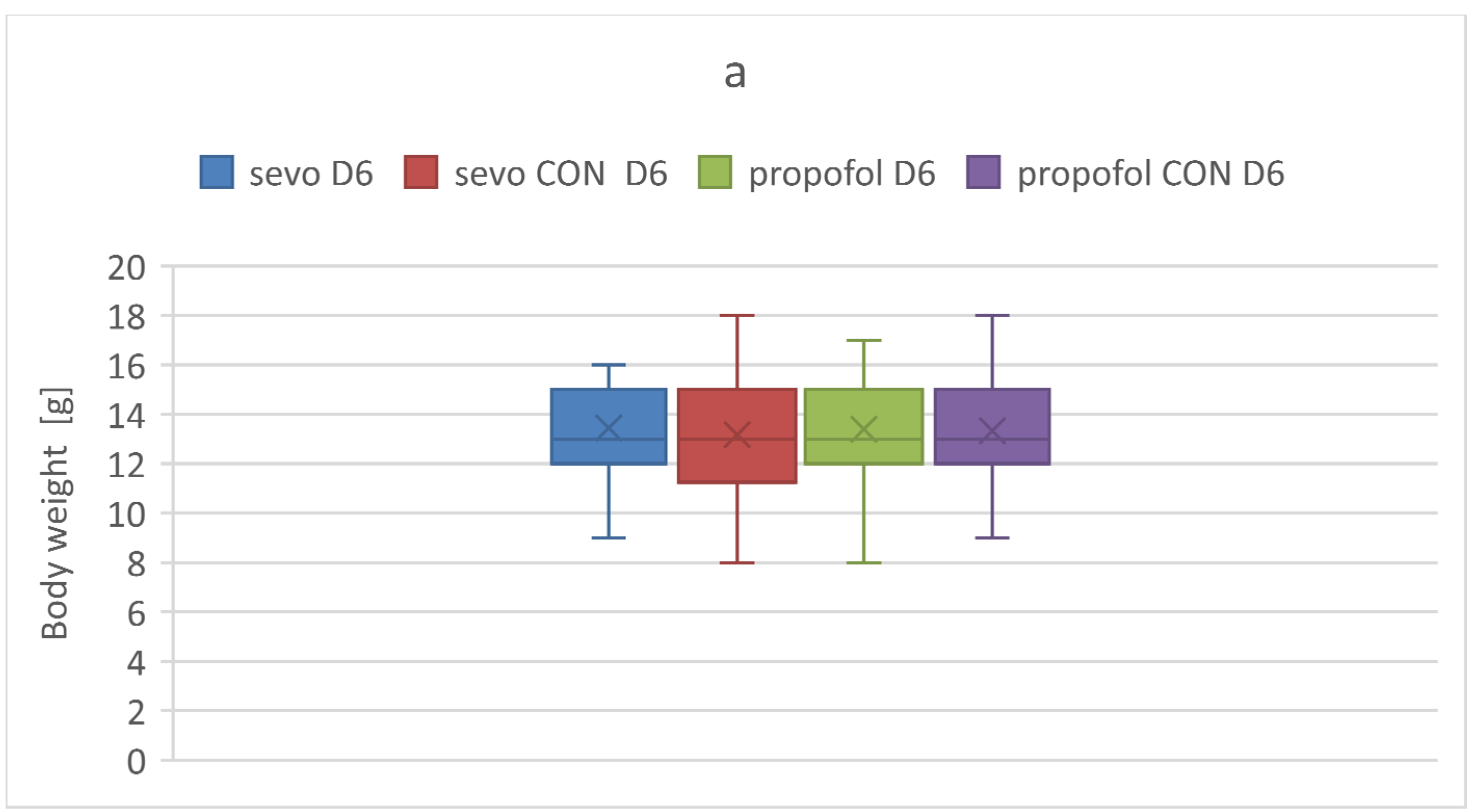
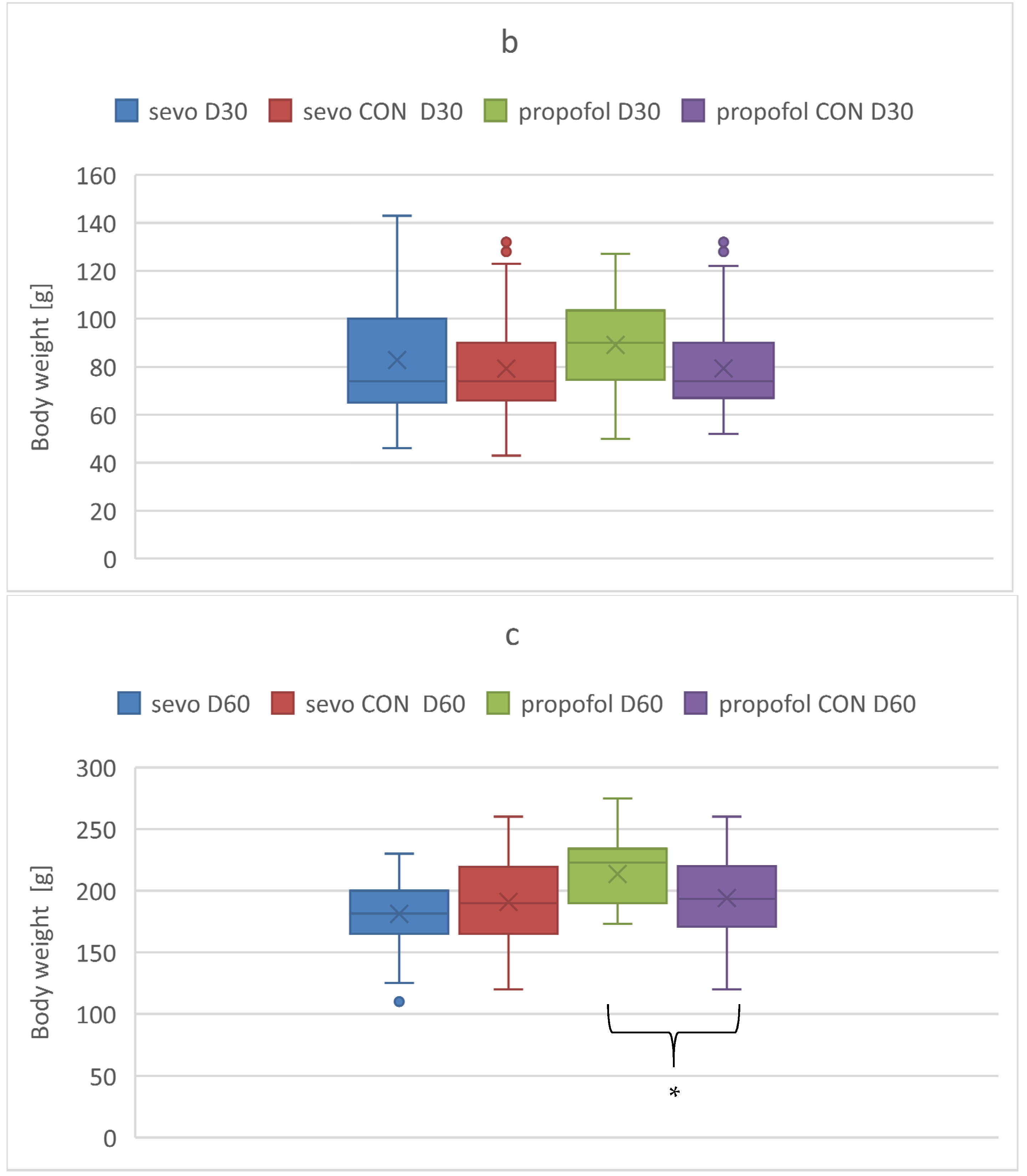
3.3. Body Weight
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Rabbitts, J.A.; Groenewald, C.B. Epidemiology of pediatric surgery in the United States. Pediatr. Anesth. 2020, 30, 1083–1090. [Google Scholar] [CrossRef]
- Ikonomidou, C.; Bosch, F.; Miksa, M.; Bittigau, P.; Vöckler, J.; Dikranian, K.; Tenkova, T.I.; Stefovska, V.; Turski, L.; Olney, J.W. Blockade of NMDA receptors and apoptotic neurodegeneration in the developing brain. Science 1999, 283, 70–74. [Google Scholar] [CrossRef]
- Jevtovic-Todorovic, V.; Hartman, R.E.; Izumi, Y.; Benshoff, N.D.; Dikranian, K.; Zorumski, C.F.; Olney, J.W.; Wozniak, D.F. Early exposure to common anesthetic agents causes widespread neurodegeneration in the developing rat brain and persistent learning deficits. J. Neurosci. 2003, 23, 876–882. [Google Scholar] [CrossRef] [Green Version]
- Bercker, S.; Bert, B.; Bittigau, P.; Felderhoff-Müser, U.; Bührer, C.; Ikonomidou, C.; Weise, M.; Kaisers, U.X.; Kerner, T. Neurodegeneration in newborn rats following propofol and sevoflurane anesthesia. Neurotox. Res. 2009, 16, 140–147. [Google Scholar] [CrossRef]
- Zhou, Z.W.; Shu, Y.; Li, M.; Guo, X.; Pac-Soo, C.; Maze, M.; Ma, D. The glutaminergic, GABAergic, dopaminergic but not cholinergic neurons are susceptible to anaesthesia-induced cell death in the rat developing brain. Neuroscience 2011, 174, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Brambrink, A.M.; Back, S.A.; Riddle, A.; Gong, X.; Moravec, M.D.; Dissen, G.A.; Creeley, C.E.; Dikranian, K.T.; Olney, J.W. Isoflurane-induced apoptosis of oligodendrocytes in the neonatal primate brain. Ann. Neurol. 2012, 72, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Raper, J.; Alvarado, M.C.; Murphy, K.L.; Baxter, M.G. Multiple anesthetic exposure in infant monkeys alters emotional reactivity to an acute stressor. Anesthesiology 2015, 123, 1084–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krasowski, M.D.; Harrison, N.L. General anaesthetic actions on ligand-gated ion channels. Cell. Mol. Life Sci. 1999, 55, 1278–1303. [Google Scholar] [CrossRef]
- Kotani, Y.; Shimazawa, M.; Yoshimura, S.; Iwama, T.; Hara, H. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci. Ther. 2008, 14, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Kuizenga, M.H.; Colin, P.J.; Reyntjens, K.M.E.M.; Touw, D.J.; Nalbat, H.; Knotnerus, F.H.; Vereecke, H.E.M.; Struys, M.M.R.F. Population Pharmacodynamics of Propofol and Sevoflurane in Healthy Volunteers Using a Clinical Score and the Patient State Index: A Crossover Study. Anesthesiology 2019, 131, 1223–1238. [Google Scholar] [CrossRef]
- Nikizad, H.; Yon, J.H.; Carter, L.B.; Jevtovic-Todorovic, V. Early exposure to general anesthesia causes significant neuronal deletion in the developing rat brain. Ann. N. Y. Acad. Sci. 2007, 1122, 69–82. [Google Scholar] [CrossRef]
- Dikranian, K.; Ishimaru, M.J.; Tenkova, T.; Labruyere, J.; Qin, Y.; Ikonomidou, C.; Olney, J.W. Apoptosis in the in vivo mammalian forebrain. Neurobiol. Dis. 2001, 8, 359–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tong, D.; Ma, Z.; Su, P.; Wang, S.; Xu, Y.; Zhang, L.M.; Wu, Z.; Liu, K.; Zhao, P. Sevoflurane-induced neuroapoptosis in rat dentate gyrus is activated by autophagy through NF-κB signaling on the late-stage progenitor granule cells. Front. Cell. Neurosci. 2020, 14, 590577. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.J.A.; Parnell, S.E.; West, J.R. Effects of alcohol and nicotine on developing olfactory bulb: Loss of mitral cells and alterations in neurotransmitter levels. Alcohol. Clin. Exp. Res. 1999, 23, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Bailey, C.D.; Brien, J.F.; Reynolds, J.N. Altered GABA A benzodiazepine receptor number and pharmacology in the adult guinea pig cerebral cortex after chronic prenatal ethanol exposure. Alcohol. Clin. Exp. Res. 1999, 23, 1816–1824. [Google Scholar] [CrossRef]
- Bouilleret, V.; Ridoux, V.; Depaulis, A.; Marescaux, C.; Nehlig, A.; Salle, G.L.G.L. Recurrent seizures and hippocampal sclerosis following intrahippocampal kainate injection in adult mice: Electroencephalography, histopathology and synaptic reorganization similar to mesial temporal lobe epilepsy. Neuroscience 1999, 89, 717–729. [Google Scholar] [CrossRef]
- Cavazos, J.E.; Sutula, T.P. Progressive neuronal loss induced by kindling: A possible mechanism for mossy fiber synaptic reorganization and hippocampal sclerosis. Brain Res. 1990, 527, 1–6. [Google Scholar] [CrossRef]
- Germano, I.M.; Sperber, E.F.; Ahuja, S.; Moshé, S.L. Evidence of enhanced kindling and hippocampal neuronal injury in immature rats with neuronal migration disorders. Epilepsia 1998, 39, 1253–1260. [Google Scholar] [CrossRef]
- Stringer, J.L. Pentylenetetrazol elicits epileptiform activity in the dentate gyrus of the urethane anesthetized rat by activation of the entorhinal cortex. Brain Res. 1994, 636, 221–226. [Google Scholar] [CrossRef]
- Moshe, S. Epileptogenesis and the immature brain. Epilepsia 1987, 28, 3–15. [Google Scholar] [CrossRef]
- Lothman, E.W. Pathophysiology of Seizures and Epilepsy in the Mature and Immature Brain: Cells, Synapses and Circuits in Pediatric Epilepsy: Diagnosis and Therapy; Demos Publications: New York, NY, USA, 1993; pp. 1–15. [Google Scholar]
- Nadler, J.V. Axon sprouting in epilepsy. In Encyclopedia of Basic Epilepsy Research; Academic Press: Oxford, UK, 2009; pp. 1143–1148. [Google Scholar]
- Hauser, W.; Kurland, L.T. The epidemiology of epilepsy in Rochester, Minnesota, 1935–1967. Epilepsia 1975, 16, 1–66. [Google Scholar] [CrossRef]
- Holmes, G.L. Effect of seizures on brain development: Lessons from the laboratory. Pediatr. Neurol. 2005, 33, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pitkänen, A.; Lukasiuk, K. Molecular and cellular basis of epileptogenesis in symptomatic epilepsy. Epilepsy Behav. 2009, 14, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Fisher, R.; Acevedo, C.; Arzimanoglou, A. A practical clinical definition of epilepsy. Epilepsia 2014, 55, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Epilepsy: Aetiology, Epidemiology and Prognosis; WHO: Geneva, Switzerland, 2001; Volume Fact Sheet N 165. [Google Scholar]
- Ngugi, A.K.; Bottomley, C.; Kleinschmidt, I.; Sander, J.; Newton, C. Estimation of the burden of active and life-time epilepsy: A meta-analytic approach. Epilepsia 2010, 51, 883–890. [Google Scholar] [CrossRef]
- Sillanpää, M.; Gissler, M.; Schmidt, D. Efforts in epilepsy prevention in the last 40 years: Lessons from a large nationwide study. JAMA Neurol. 2016, 73, 390–395. [Google Scholar] [CrossRef] [Green Version]
- Turski, W.A.; Cavalheiro, E.A.; Schwarz, M.; Czuczwar, S.J.; Kleinrok, Z.; Turski, L. Limbic seizures produced by pilocarpine in rats: Behavioural, electroencephalographic and neuropathological study. Behav. Brain Res. 1983, 9, 315–335. [Google Scholar] [CrossRef]
- Pitkänen, A.; Engel, J., Jr. Past and present definitions of epileptogenesis and its biomarkers. Neurotherapeutics 2014, 11, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Schubert, M.; Siegmund, H.; Pape, H.C.; Albrecht, D. Kindling-induced changes in plasticity of the rat amygdala and hippocampus. Learn. Mem. 2005, 12, 520–526. [Google Scholar] [CrossRef] [Green Version]
- Pitkänen, A.; Kharatishvili, I.; Karhunen, H.; Lukasiuk, K.; Immonen, R.; Nairismägi, J.; Gröhn, O.; Nissinen, J. Epileptogenesis in experimental models. Epilepsia 2007, 48, 13–20. [Google Scholar] [CrossRef]
- Löscher, W.; Brandt, C. Prevention or modification of epileptogenesis after brain insults: Experimental approaches and translational research. Pharmacol. Rev. 2010, 62, 668–700. [Google Scholar] [CrossRef] [Green Version]
- Forcelli, P.A.; Kozlowski, R.; Snyder, C.; Kondratyev, A.; Gale, K. Effects of neonatal antiepileptic drug exposure on cognitive, emotional, and motor function in adult rats. Pharmacol. Exp. Ther. 2012, 340, 558–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonthius, D.J.; Woodhouse, J.; Bonthius, N.E.; Taggard, D.A.; Lothman, E.W. Reduced seizure threshold and hippocampal cell loss in rats exposed to alcohol during the brain growth spurt. Alcohol. Clin. Exp. Res. 2001, 25, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.E.; Cain, D.P. A single pentylenetetrazol or hyperthermia convulsion increases kindling susceptibility in the adult rat. Brain Res. 1985, 22, 169–180. [Google Scholar] [CrossRef]
- Wlaź, P.; Potschka, H.; Loscher, W. Frontal versus transcorneal stimulation to induce maximal electroshock seizures or kindling in mice and rats. Epilepsy Res. 1998, 30, 219–229. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation. II. Motor seizure. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 281–294. [Google Scholar] [CrossRef]
- Litchfield, J.T.; Wilcoxon, F. A simplified method of evaluating dose-effect experiments. J. Pharm. Exp. Ther. 1949, 96, 99–113. [Google Scholar]
- Lévesque, M.; Avoli, M. The subiculum and its role in focal epileptic disorders. Rev. Neurosci. 2020, 32, 249–273. [Google Scholar] [CrossRef]
- Dengler, C.G.; Coulter, D.A. Normal and epilepsy-associated pathologic function of the dentate gyrus. Prog. Brain Res. 2016, 226, 155–178. [Google Scholar]
- Fang, F.; Xue, Z.; Cang, J. Sevoflurane exposure in 7-day-old rats affects neurogenesis, neurodegeneration and neurocognitive function. Neurosci. Bull. 2012, 28, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Chen, K.; Shen, X. Environmental Enrichment Attenuated Sevoflurane-Induced Neurotoxicity through the PPAR-γ Signaling Pathway. BioMed Res. Int. 2015, 2015, 107149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwan, P.; Sander, J.W. The natural history of epilepsy: An epidemiological view. J. Neurol. Neurosurg. 2004, 75, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
- Tagashira, E.; Nakao, K.; Urano, T.; Hiramori, T.; Yanaura, S. Alteration of convulsive threshold and sensitivity to CNS acting drugs in sedativehypnotics-experienced rat offspring. Jpn. J. Pharmacol. 1982, 32, 263–271. [Google Scholar] [CrossRef]
- Marcus, J.C. Neurological findings in the fetal alcohol syndrome. Neuropediatrics 1987, 18, 158–160. [Google Scholar] [CrossRef]
- Ioffe, S.; Chernick, V. Prediction of subsequent motor and mental retardation in newborn infants exposed to alcohol in utero by computerized EEG analysis. Neuropediatrics 1990, 21, 11–17. [Google Scholar]
- Kang, M.-H.; Spigelman, I.; Olsen, R.W. Alteration in the sensitivity of GABA A receptors to allosteric modulatory drugs in rat hippocampus after chronic intermittent ethanol treatment. Alcohol. Clin. Exp. Res. 1998, 22, 2165–2173. [Google Scholar] [CrossRef] [PubMed]
- Makaryus, R.; Lee, H.; Feng, T.; Park, J.H.; Nedergaard, M.; Jacob, Z.; Enikolopov, G.; Benveniste, H. Brain maturation in neonatal rodents is impeded by sevoflurane anesthesia. Anesthesiology 2015, 123, 557–568. [Google Scholar] [CrossRef] [Green Version]
- Peraino, C.; Ehret, C.F.; Groh, K.R.; Meinert, J.C.; D’Arcy-Gomez, G. Phenobarbital effects on weight gain and circadian cycling of food intake and body temperature. Proc. Soc. Exp. Biol. Med. 1980, 165, 473–479. [Google Scholar] [CrossRef]
- Satomoto, M.; Satoh, Y.; Terui, K.; Miyao, H.; Takishima, K.; Ito, M.; Imaki, J. Neonatal exposure to sevoflurane induces abnormal social behaviors and deficits in fear conditioning in mice. Anesthesiology 2009, 110, 628–637. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substance | CD50 Pilocarpine (mg/kg) | n | p |
|---|---|---|---|
| Propofol control group | 344.1 (337.7–350.7) | 7 | 0.63 |
| Propofol | 339.4 (330–349.1) | 8 | |
| Sevoflurane control group | 341.9 (317–368.7) | 8 | 0.42 |
| Sevoflurane | 323.3 (305–362.1) | 8 |
| Substance | CD50 Pentylenetetrazole (mg/kg) | n | p |
|---|---|---|---|
| Propofol control group | 108.5 (92.8–126.8) | 8 | 0.21 |
| Propofol | 95.6 (77.9–117.3) | 8 | |
| Sevoflurane control group | 104.9 (86.8–126.9) | 8 | 0.19 |
| Sevoflurane | 94.3 (78–114) | 8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piwowarczyk, P.; Rypulak, E.; Sysiak-Sławecka, J.; Nieoczym, D.; Socała, K.; Wlaź, A.; Wlaź, P.; Turski, W.; Czuczwar, M.; Borys, M. Propofol and Sevoflurane Anesthesia in Early Childhood Do Not Influence Seizure Threshold in Adult Rats. Int. J. Environ. Res. Public Health 2021, 18, 12367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182312367
Piwowarczyk P, Rypulak E, Sysiak-Sławecka J, Nieoczym D, Socała K, Wlaź A, Wlaź P, Turski W, Czuczwar M, Borys M. Propofol and Sevoflurane Anesthesia in Early Childhood Do Not Influence Seizure Threshold in Adult Rats. International Journal of Environmental Research and Public Health. 2021; 18(23):12367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182312367
Chicago/Turabian StylePiwowarczyk, Paweł, Elżbieta Rypulak, Justyna Sysiak-Sławecka, Dorota Nieoczym, Katarzyna Socała, Aleksandra Wlaź, Piotr Wlaź, Waldemar Turski, Mirosław Czuczwar, and Michał Borys. 2021. "Propofol and Sevoflurane Anesthesia in Early Childhood Do Not Influence Seizure Threshold in Adult Rats" International Journal of Environmental Research and Public Health 18, no. 23: 12367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph182312367