
Isolation and Characterization of a Biosurfactant Producing Strain Planococcus sp. XW-1 from the Cold Marine Environment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Culture Medium
2.2. Isolation of the Bacteria
2.2.1. Isolation of the Petroleum Degrading Bacteria
2.2.2. Surface Tension Measurement
2.2.3. Diesel Oil Degradation Measurement
2.3. Identification of the Bacteria
2.3.1. Morphological Identification
2.3.2. Molecular Identification
2.4. Extraction of Biosurfactant
2.5. Identification of Biosurfactant
2.6. Critical Micelle Concentration (CMC) Measurement
2.7. Influence of Environmental Factors on Biosurfactant Stability
2.8. Influence of Carbon Source on Biosurfactant Production
2.9. Biodegradation of Crude Oil
3. Results and Discussion
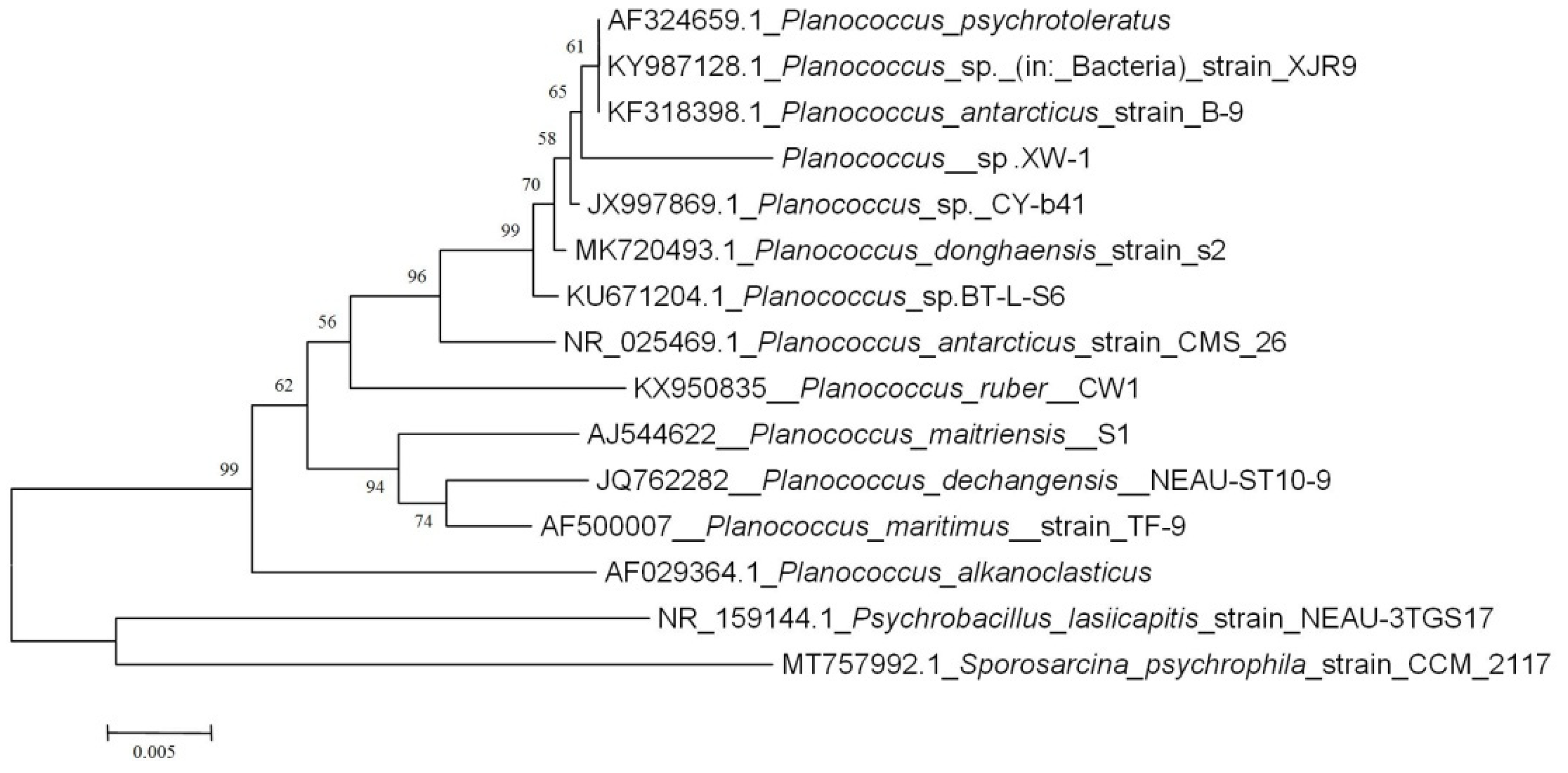
3.1. Isolation and Identification of the Petroleum Degrading and Surfactant Producing Bacteria
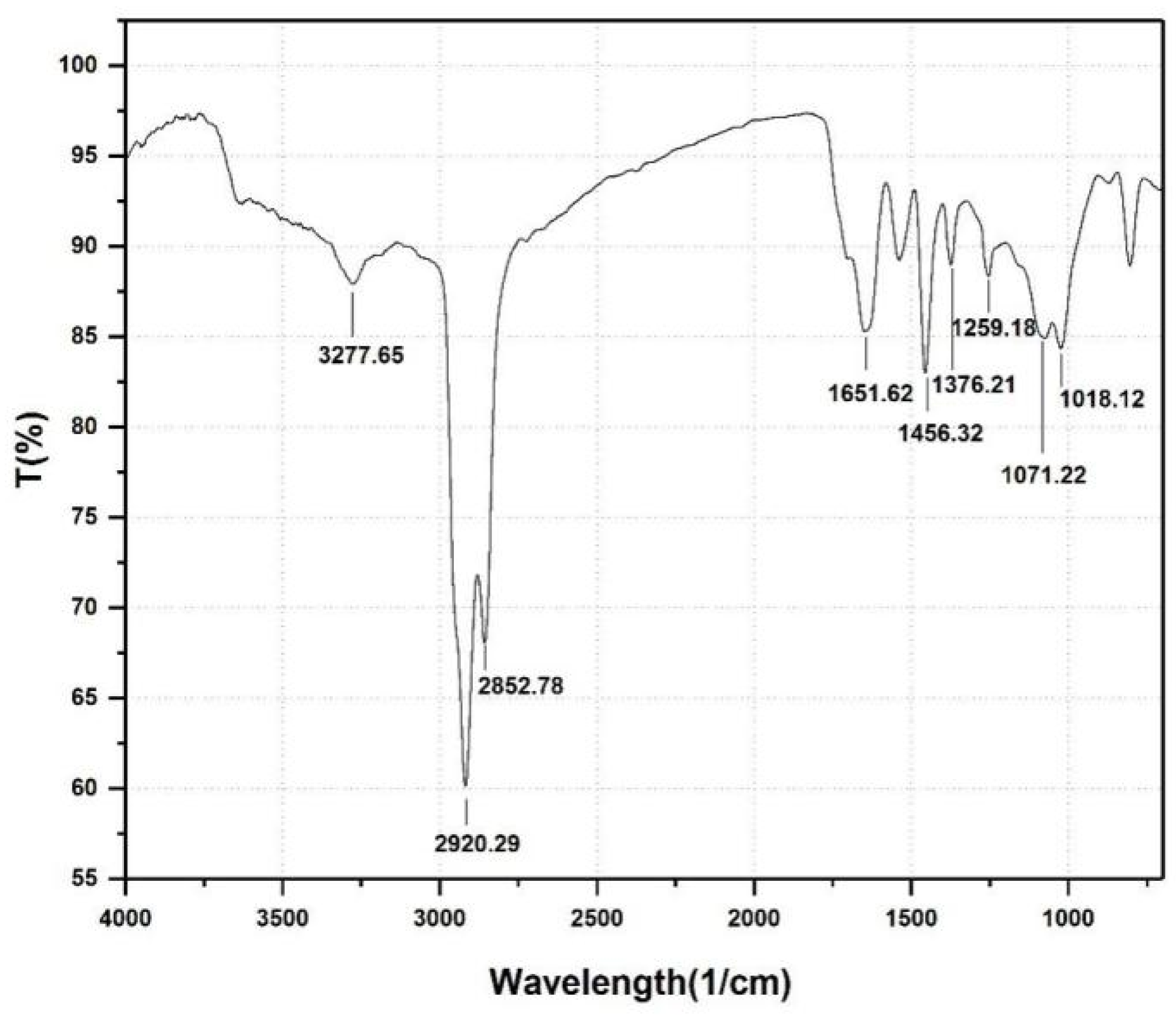
3.2. Identification of the Biosurcatant
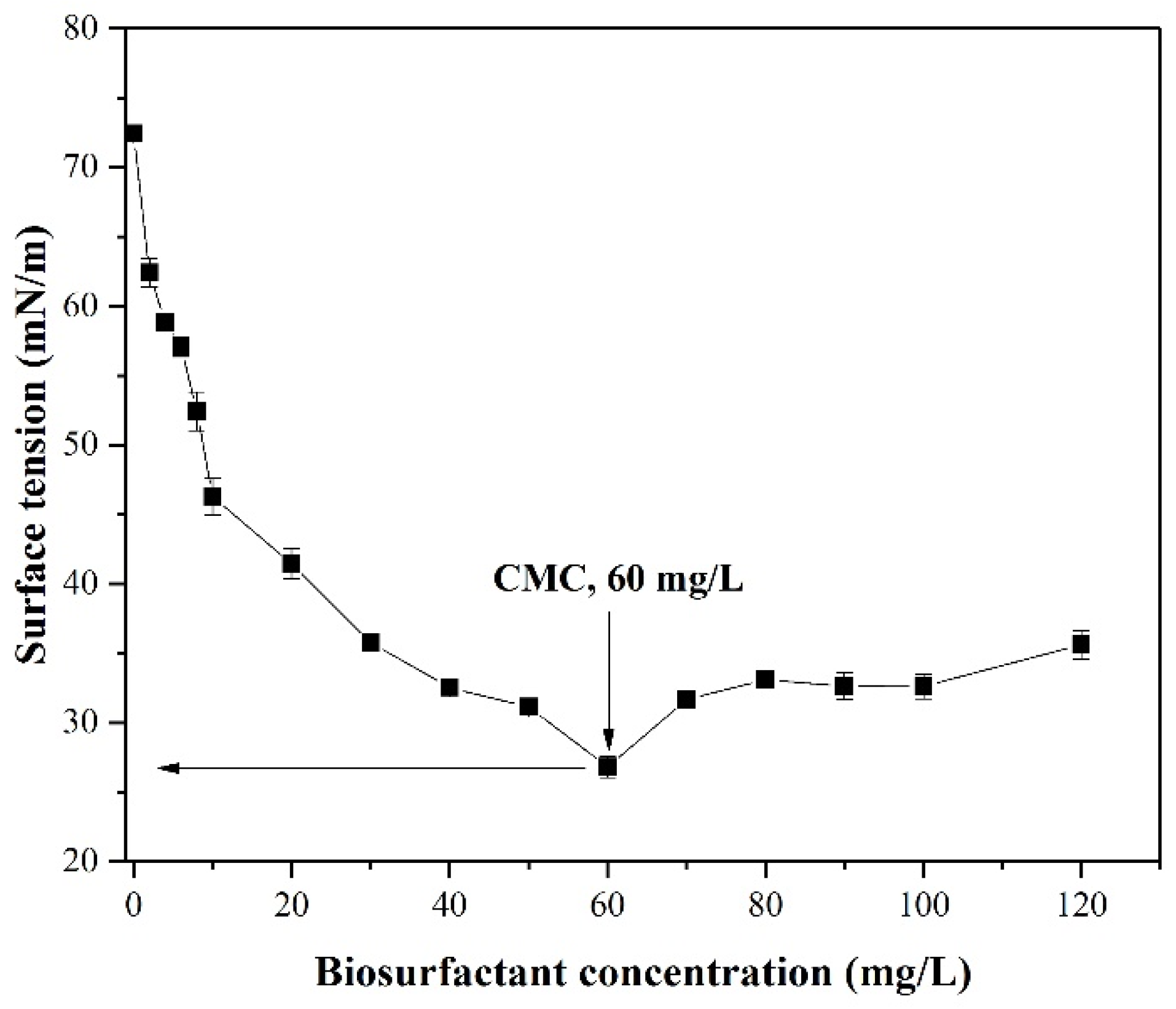
3.3. Determination of CMC Value
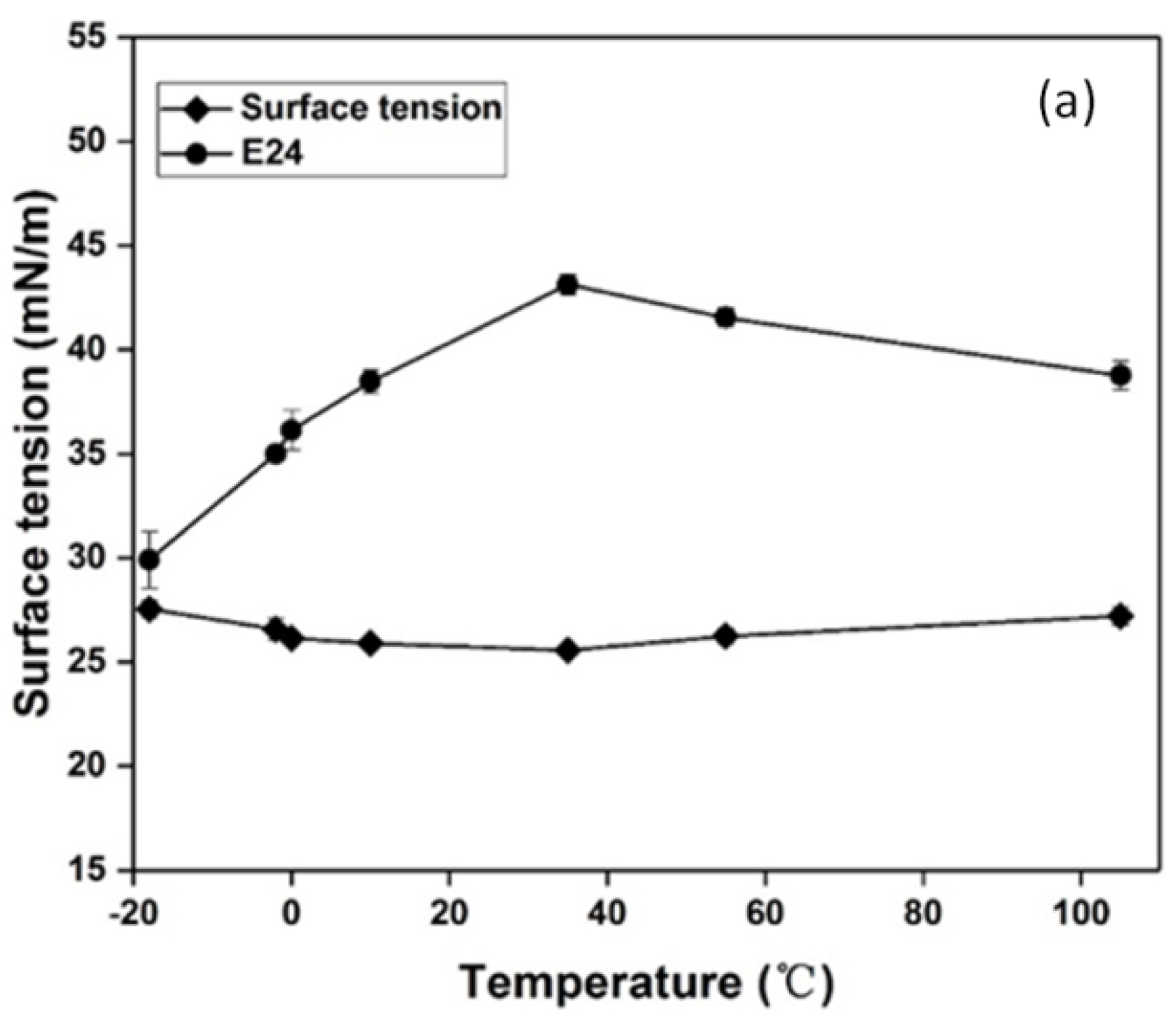
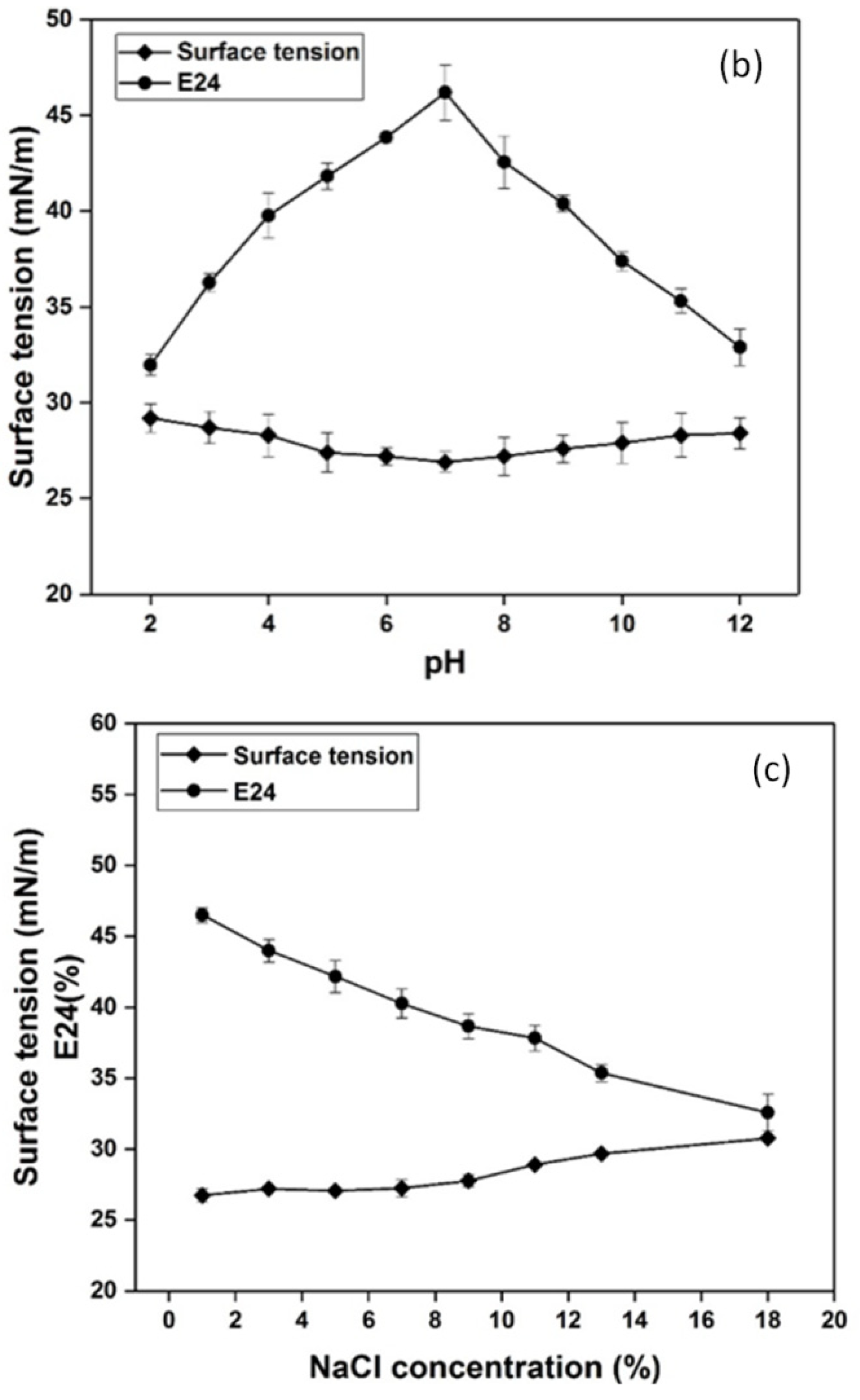
3.4. Effect of Temperature, pH and Salinity on the Stability of Biosurfactant
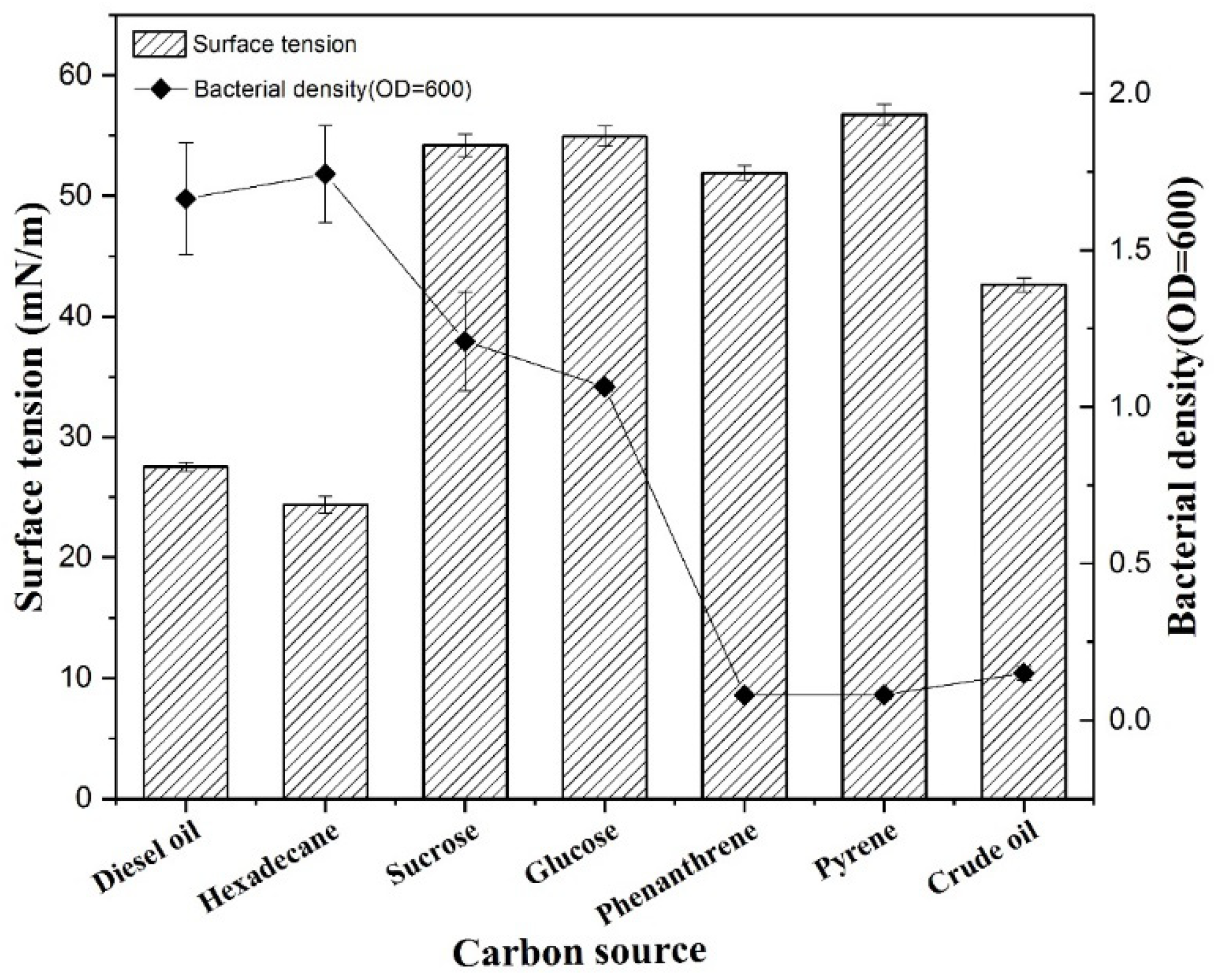
3.5. Effect of Seven Carbon Sources on Biosurfactant Production
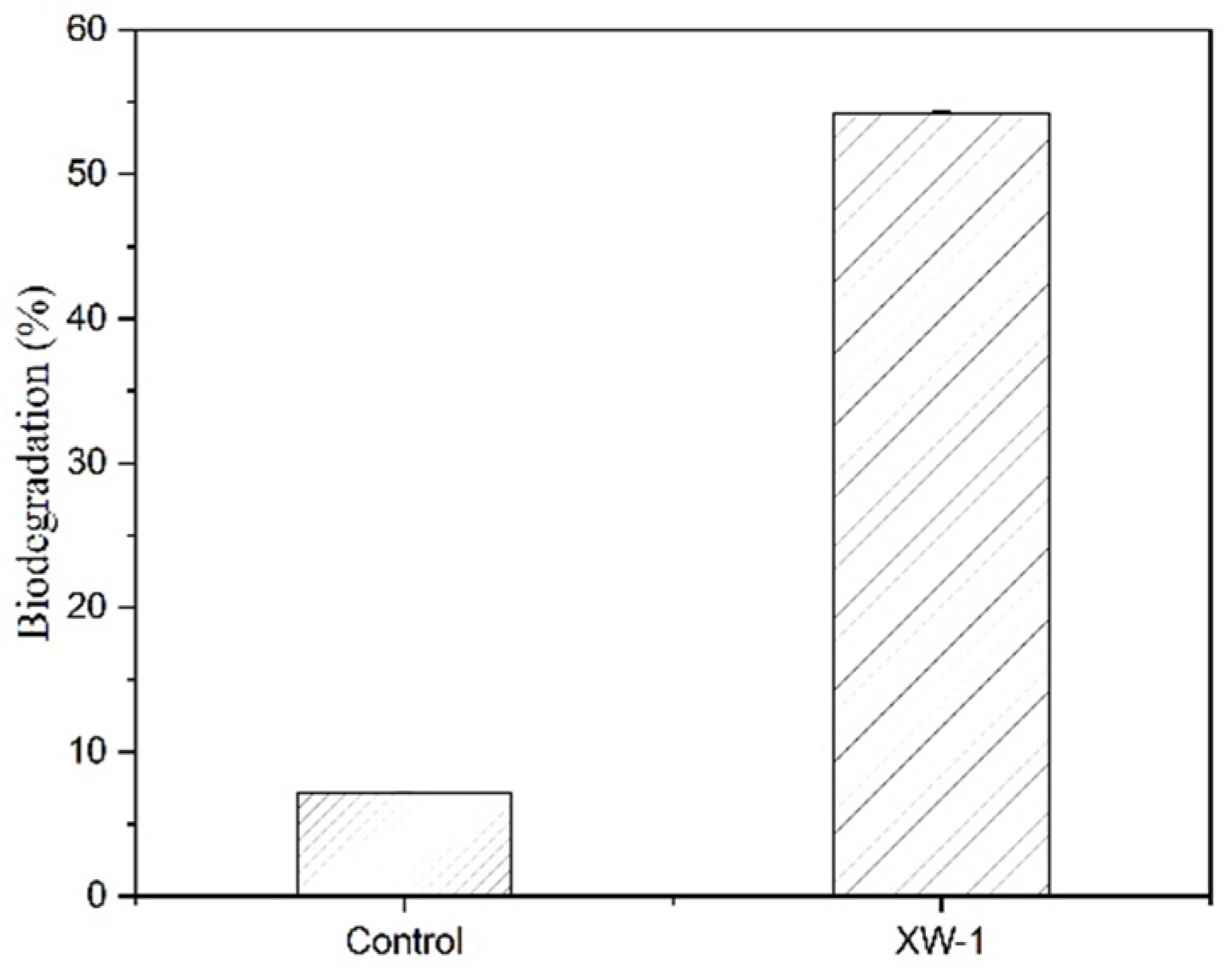
3.6. Biodegradation of Crude Oil
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gargouri, B.; Karray, F.; Mhiri, N.; Aloui, F.; Sayadi, S. Bioremediation of petroleum hydrocarbons-contaminated soil by bacterial consortium isolated from an industrial wastewater treatment plant. J. Chem. Technol. Biotechnol. 2014, 89, 978–987. [Google Scholar] [CrossRef]
- Xia, W.; Du, Z.; Cui, Q.; Dong, H.; Wang, F.; He, P.; Tang, Y. Biosurfactant produced by novel Pseudomonas sp. WJ6 with biodegradation of n-alkanes and polycyclic aromatic hydrocarbons. J. Hazard. Mater. 2014, 276, 489–498. [Google Scholar] [CrossRef]
- Chandankere, R.; Yao, J.; Cai, M.; Masakoralaabe, K.; Jain, A.K.; Martin, M.F.C. Properties and characterization of biosurfactant in crude oil biodegradation by bacterium Bacillus methylotrophicus USTBa. Fuel 2014, 122, 140–148. [Google Scholar] [CrossRef]
- Mohn, W.W.; Stewart, G.R. Limiting factors for hydrocarbon biodegradation at low temperature in Arctic soils. Soil Biol. Biochem. 2000, 32, 1161–1172. [Google Scholar] [CrossRef]
- Borah, D.; Yadav, R.N.S. Bioremediation of petroleum based contaminants with biosurfactant produced by a newly isolated petroleum oil degrading bacterial strain. Egypt. J. Pet. 2016, 26, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Giudice, A.L.; Fani, R. Cold-adapted bacteria from a coastal area of the Ross Sea (Terra Nova Bay, Antarctica): Linking microbial ecology to biotechnology. Hydrobiologia 2015, 761, 417–441. [Google Scholar] [CrossRef]
- Fingas, M.F. Modeling evaporation using models that are not boundary-layer regulated. J. Hazard. Mater. 2004, 107, 27–36. [Google Scholar] [CrossRef]
- Michaud, L.; Giudice, A.L.; Saitta, M.; De Domenico, M.; Bruni, V. The biodegradation efficiency on diesel oil by two psychotropic Antarctic marine bacteria during a two-month-long experiment. Mar. Pollut. Bull. 2004, 49, 405–409. [Google Scholar] [CrossRef] [PubMed]
- Bezza, F.A.; Beukes, M.; Chirwa, E.M.N. Application of biosurfactant produced by Ochrobactrum intermedium CN3 for enhancing petroleum sludge bioremediation. Process Biochem. 2015, 51, 343. [Google Scholar] [CrossRef]
- Petrikov, K.; Delegan, Y.; Surin, A.; Ponamoreva, O.; Puntus, I.; Filonov, A.; Boronin, A. Glycolipids of Pseudomonas and Rhodococcus oil-degrading bacteria used in bioremediation preparations: Formation and structure. Process Biochem. 2013, 48, 931–935. [Google Scholar] [CrossRef]
- Jung, J.H.; Joe, M.H.; Kim, D.H.; Park, H.; Choi, J.I.; Lim, S.Y. Complete genome sequence of Planococcus sp. PAMC21323 isolated from Antarctica and its metabolic potential to detoxify pollutants. Stand. Genomic. Sci. 2018, 13, 31. [Google Scholar] [CrossRef]
- Waghmode, S.; Suryavanshi, M.; Dama, L.; Kansara, S.; Ghattargi, V.; Das, P.; Banpurkar, A.; Satpute, S.K. Genomic Insights of Halophilic Planococcus maritimus SAMP MCC 3013 and Detail Investigation of Its Biosurfactant Production. Front. Microb. 2019, 10, 00235. [Google Scholar] [CrossRef]
- Waghmode, S.; Suryavanshi, M.; Sharma, D.; Satpute, S.K. Planococcus Species—An Imminent Resource to Explore Biosurfactant and Bioactive Metabolites for Industrial Applications. Front. Bioeng. Biotechnol. 2020, 8, 996. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.S.; Mody, K.; Jha, B. Evaluation of biosurfactant/bioemulsifier production by a marine bacterium. Bull. Environ. Contam. Toxicol. 2007, 79, 617–621. [Google Scholar] [CrossRef]
- Ebrahimipour, G.; Gilavand, F.; Karkhane, M.; Kavyanifard, A.; Teymouri, M.; Marzban, A. Bioemulsification activity assessment of an indigenous strain of halotolerant Planococcus and partial characterization of produced biosurfactants. Int. J. Environ. Sci. Technol. 2014, 11, 1379–1386. [Google Scholar] [CrossRef] [Green Version]
- Marchant, R.; Banat, I.M. Biosurfactants: A sustainable replacement for chemical surfactants. Biotechnol. Lett. 2012, 34, 1597–1605. [Google Scholar] [CrossRef] [PubMed]
- Varjani, S.J.; Upasani, V.N. Critical review on biosurfactant analysis, purification and characterization using rhamnolipid as a model biosurfactant. Bioresour. Technol. 2017, 232, 389–397. [Google Scholar] [CrossRef]
- Ohadi, M.; Dehghannoudeh, G.; Forootanfar, H.; Shakibaie, M.; Rajaee, M. Investigation of the structural, physicochemical properties, and aggregation behavior of lipopeptide biosurfactant produced by Acinetobacter junii B6. Int. J. Biol. Macromol. 2018, 112, 712–719. [Google Scholar] [CrossRef]
- Mulligan, C.N. Recent advances in the environmental applications of biosurfactants. Curr. Opin. Colloid Interface Sci. 2009, 14, 372–378. [Google Scholar] [CrossRef]
- Pepi, M.; Cesaro, A.; Liut, G.; Baldi, F. An antractic psychrotrophic bacterium Halomonas sp. ANT-3b, growing on n-hexadecane, produces a new emulsifying glycolipid. FEMS Microbiol. Ecol. 2005, 53, 157–166. [Google Scholar] [CrossRef]
- Bej, A.K.; Saul, D.; Aislabie, J. Cold-tolerant alkane-degrading Rhodococcus species from Antarctica. Polar Biol. 2000, 23, 100–105. [Google Scholar] [CrossRef]
- Ruberto, L.A.M.; Vazquez, S.; Lobalbo, A. Psychrotolerant hydrocarbon-degrading Rhodococcus strains isolated from polluted Antarctic soils. Antarct. Sci. 2005, 17, 47–56. [Google Scholar] [CrossRef]
- Giudice, A.L.; Casella, P.; Caruso, C.; Mangano, S.; Bruni, V.; Domenico, M.D.; Michaud, L. Occurrence and characterization of psychrotolerant hydrocarbon-oxidizing bacteria from surface seawater along the Victoria Land coast (Antarctica). Polar Biol. 2010, 33, 929–943. [Google Scholar] [CrossRef]
- Aislabie, J.; Foght, J.; Saul, D. Aromatic hydrocarbon-degrading bacteria from soil near Scott Base, Antarctica. Polar Biol. 2000, 23, 183–188. [Google Scholar] [CrossRef]
- Farrell, R.L.; Rhodes, P.L.; Aislabie, J. Toluene-degrading Antarctic Pseudomonas strains from fuel-contaminated soil. Biochem. Biophys. Res. Commun. 2003, 312, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Saul, D.J.; Aislabie, J.M.; Brown, C.E.; Harris, L.; Foght, J.M. Hydrocarbon contamination changes the bacterial diversity of soil from around Scott Base, Antarctica. FEMS Microbiol. Ecol. 2005, 53, 141–155. [Google Scholar] [CrossRef] [Green Version]
- Bao, X.W.; Wang, X.Q.; Gao, G.P.; Wu, D.X. The characteristics of the seasonal variability of the sea surface temperature field in the Bohai Sea, the Huanghai Sea and the East China Sea from AVHRR data. Acta Oceanol. Sin. 2002, 24, 125–133. [Google Scholar]
- Wang, C.; He, S.; Li, Y. Research on the status of marine oil spill pollution in China and its ecological impact. Mar. Sci. 2009, 33, 57–60. [Google Scholar]
- Li, L.; Shen, X.; Zhao, C.; Liu, Q.; Liu, X.; Wu, Y. Biodegradation of dibenzothiophene by efficient Pseudomonas sp. LKY-5 with the production of a biosurfactant. Ecotoxicol. Environ. Saf. 2019, 176, 50–57. [Google Scholar] [CrossRef]
- Cheng, X.Y.; Liu, W.W.; Xu, Y.; Zhou, N.Y. Screening and characterization of culturable hydrocarbon-degrading strains from the South and East China Seas. J. Tianjin Univ. Technol. 2019, 46, 975–985. [Google Scholar]
- Hassanshahian, M.; Emtiazi, G.; Cappello, S. Isolation and characterization of crude-oil-degrading bacteria from the Persian Gulf and the Caspian Sea. Mar. Pollut. Bull. 2012, 64, 7–12. [Google Scholar] [CrossRef]
- Pornsunthorntawee, O.; Wongpanit, P.; Chavadej, S. Structural and physicochemical characterization of crude biosurfactant produced by Pseudomonas aeruginosa SP4 isolated from petroleum-contaminated soil. Bioresour. Technol. 2008, 99, 1589–1595. [Google Scholar] [CrossRef]
- Durval, I.J.B.; Mendonça, A.H.R.; Rocha, I.V.; Luna, J.M.; Rufino, R.D.; Converti, A.; Sarubbo, L.A. Production, characterization, evaluation and toxicity assessment of a Bacillus cereus UCP 1615 biosurfactant for marine oil spills bioremediation. Mar. Pollut. Bull. 2020, 157, 111357. [Google Scholar] [CrossRef]
- Xue, J.L.; Wu, Y.N.; Liu, Z.X.; Li, M.L.; Sun, X.Y.; Wang, H.J.; Liu, B. Characteristic Assessment of Diesel-degrading Bacteria Immobilized on Natural Organic Carriers in Marine Environment: The Degradation Activity and Nutrient. Sci. Rep. 2017, 7, 8635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siamak, P.Y.; Henning, S.; Hans, J.S.L.; Geir, G. Use of magnetic beads for Gram staining of bacteria in aqueous suspension. J. Microbiol. Methods 2001, 47, 369–371. [Google Scholar]
- Atakpa, E.O.; Zhou, H.H.; Jiang, L.J.; Ma, Y.H.; Liang, Y.P.; Li, Y.H.; Zhang, D.D.; Zhang, C.F. Improved degradation of petroleum hydrocarbons by co-culture of fungi and biosurfactant-producing bacteria. Chemosphere 2022, 290, 133337. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Meyers, E.W.; Lipman, D.J. Basic local alighnment search tool. J. Mol. Biol. 1990, 215, 403. [Google Scholar] [CrossRef]
- Gaur, V.; Tripathi, V.; Gupta, P.; Dhiman, N.; Regar, R.K.; Gautam, K.; Srivastava, J.K.; Patnaik, S.; Patel, D.K.; Manickam, N. Rhamnolipids from planococcus spp. and their mechanism of action against pathogenic bacteria. Bioresour. Technol. 2020, 307, 123206. [Google Scholar] [CrossRef]
- Cooper, D.G.; Goldenberg, B.G. Surface-active agents from two Bacillus species. Appl. Environ. Microbiol. 1987, 53, 224–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, S.K.S.; Gudiña, E.J.; Schwan, R.F.; Rodrigues, L.R.; Dias, D.R.; Teixeira, J.A. Improvement of biosurfactant production by Wickerhamomyces anomalus CCMA 0358 and its potential application in bioremediation. J. Hazard. Mater. 2018, 346, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, M.D.; Noller, H.F. The draft genome of Planococcus donghaensis MPA1U2 reveals nonsporulation pathways controlled by a conserved Spo0A regulon. J. Bacteriol. 2011, 193, 6106. [Google Scholar] [CrossRef] [Green Version]
- Abelardo, M.; Miguel, G.; Borja, S. Genome sequence of the Antarctic psychrophile bacterium Planococcus antarcticus DSM 14505. J. Bacteriol. 2012, 194, 4465. [Google Scholar]
- Lotfabad, T.B.; Abassi, H.; Ahmadkhaniha, R.; Roostaazad, R.; Masoomi, F.; Zahiri, H.S.; Ahmadian, G.; Vali, H.; Noghabi, K.A. Structural characterization of a rhamnolipid-type biosurfactant produced by Pseudomonas aeruginosa MR01: Enhancement of di-rhamnolipid proportion using gamma irradiation. Colloids Surf. B Biointerfaces 2010, 81, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Raza, Z.A.; Khalid, Z.M.; Banat, I.M. Characterization of rhamnolipids produced by a Pseudomonas aeruginosa mutant strain grown on waste oils. J. Environ. Sci. Health Part A 2009, 44, 1367–1373. [Google Scholar] [CrossRef]
- Varjani, S.J.; Upasani, V.N. Core Flood study for Enhanced Oil Recovery through ex-situ bioaugmentation with thermo-and halo-tolerant rhamnolipid produced by Pseudomonas aeruginosa NCIM 5514. Bioresour. Technol. 2016, 220, 175–182. [Google Scholar] [CrossRef]
- Abouseoud, M.; Maachi, R.; Amrane, A.; Boudergua, S. Evaluation of different carbon and nitrogen sources in production of biosurfactant by Pseudomonas fluorescens. Desalination 2008, 223, 143–151. [Google Scholar] [CrossRef]
- Saikia, R.R.; Deka, S.; Deka, M.; Banat, I.M. Isolation of biosurfactant-producing Pseudomonas aeruginosa RS29 from oil-contaminated soil and evaluation of different nitrogen sources in biosurfactant production. Ann. Microbiol. 2012, 62, 753–763. [Google Scholar] [CrossRef]
- Li, S.; Pi, Y.; Bao, M.; Zhang, C.; Zhao, D.; Li, Y.; Sun, P.; Lu, J. Effect of rhamnolipid biosurfactant on solubilization of polycyclic aromatic hydrocarbons. Mar. Pollut. Bull. 2015, 101, 219–225. [Google Scholar] [CrossRef]
- Khadenolhosseini, R.; Jafari, A.; Mousavi, S.M.; Hajfarajollah, H.; Noghabi, K.A.; Manteghian, M. Physicochemical characterization and optimization of glycolipid biosurfactant production by a native strain of Pseudomonas aeruginosa HAK01 and its performance evaluation for the MEOR process. RSC Adv. 2019, 14, 7932. [Google Scholar] [CrossRef] [Green Version]
- Bezza, F.A.; Chirwa, E.M.N. The Role of Lipopeptide Biosurfactant on Microbial Remediation of Aged Polycyclic Aromatic Hydrocarbon (PAHs)-contaminated Soil. Chem. Eng. J. 2017, 309, 563–576. [Google Scholar] [CrossRef]
- Parkinson, M. Bio-surfactants. Biotechnol. Adv. 1985, 3, 65–83. [Google Scholar] [CrossRef]
- Kaustuvmani, P.; Rupshikha, P.; Kalita, M.C.; Deka, S. Characterization of Biosurfactant Produced during Degradation of Hydrocarbons Using Crude Oil As Sole Source of Carbon. Front. Microbiol. 2017, 8, 279. [Google Scholar]
- Heryani, H.; Putra, M.D. Kinetic study and modeling of biosurfactant production using, Bacillus sp. Electron. J. Biotechnol. 2017, 27, 49–54. [Google Scholar] [CrossRef]
- Almansoory, A.F.; Hasan, H.A.; Abdullah, S.R.H.; Idris, M.; Anuar, N.; AL-Adiwish, W.M. Biosurfactant produced by the hydrocarbon-degrading bacteria: Characterization, activity and applications in removing TPH from contaminated soil. Environ. Technol. Innov. 2019, 14, 100347. [Google Scholar] [CrossRef]
- Aparna, A.; Srinikethan, G.; Smitha, H. Production and characterization of biosurfactant produced by a novel Pseudomonas sp. 2B. Colloids Surf. B Biointerfaces 2012, 95, 23–29. [Google Scholar] [CrossRef]
- Shi, Q. Temporal and spatial modes and mechanisms of seasonal cycles of pH fields in the Yellow Sea. J. Appl. Ocean 2017, 36, 56–72. [Google Scholar]
- De Franca, I.W.L.; Lima, A.P.; Lemos, J.A.M.; Lemos, C.G.F.; Melo, V.M.M.; De Santana, H.B.; Goncalves, L.R.B. Production of a biosurfactant by Bacillus subtilis ICA56 aiming bioremediation of impacted soils. Catal. Today 2015, 255, 10–15. [Google Scholar] [CrossRef]
- Helvaci, S.S.; Peker, S.; Ozdemir, G. Effect of electrolytes on the surface behavior of rhamnolipids R1 and R2. Colloids Surf. B Biointerfaces 2004, 35, 225–233. [Google Scholar] [CrossRef]
- Dubey, K.V.; Charde, P.N.; Meshram, S.U.; Shendre, L.P.; Dubey, V.S.; Juwarkar, A.A. Surface-active potential of biosurfactants produced in curd whey by Pseudomonas aeruginosa strain-PP2 and Kocuria turfanesis strain-J at extreme environmental conditions. Bioresour. Technol. 2012, 126, 368–374. [Google Scholar] [CrossRef]
- Khopade, A.; Biao, R.; Liu, X.; Mahadik, K.; Zhang, L.; Kokare, C. Production and stability studies of the biosurfactant isolated from marine Nocardiopsis sp. B4. Desalination 2012, 285, 198–204. [Google Scholar] [CrossRef]
- Anyanwu, C.U.; Obi, S.K.C.; Okolo, B.N. Lipopeptide biosurfactant production by Serratia marcescens NSK-1 strain isolated from petroleum-contaminated soil. J. Appl. Sci. Res. 2011, 7, 79–87. [Google Scholar]
- Hou, J.N.; Zhang, C.S.; Shi, X.Y. Seasonal variations and distribution characteristics of petroleum hydrocarbons in northern Yellow Sea. Prog. Fish. Sci. 2011, 32, 121–125. [Google Scholar]
- Li, H.; Liu, Y.H.; Luo, N.; Zhang, X.Y.; Luan, T.G.; Hu, J.M.; Wang, Z.Y.; Wu, P.C.; Chen, M.J.; Lu, J.Q. Biodegradation of benzene and its derivatives by a psychrotolerant and moderately haloalkaliphilic Planococcus sp. strain ZD22. Res. Microbiol. 2006, 157, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Whyte, L.G.; Hawari, J.; Zhou, E.; Bourbonnière, L.; Inniss, W.E.; Greer, C.W. Biodegradation of variable-chain-length alkanes at low temperatures by apsychrotrophic Rhodococcus sp. Appl. Environ. Microbiol. 1998, 64, 2578–2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, P.; Xu, W.; Tang, S.; Cao, B.; Wei, D.; Zhang, M.; Lin, J.; Li, W. Isolation and Characterization of a Biosurfactant Producing Strain Planococcus sp. XW-1 from the Cold Marine Environment. Int. J. Environ. Res. Public Health 2022, 19, 782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020782
Guo P, Xu W, Tang S, Cao B, Wei D, Zhang M, Lin J, Li W. Isolation and Characterization of a Biosurfactant Producing Strain Planococcus sp. XW-1 from the Cold Marine Environment. International Journal of Environmental Research and Public Health. 2022; 19(2):782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020782
Chicago/Turabian StyleGuo, Ping, Weiwei Xu, Shi Tang, Binxia Cao, Danna Wei, Manxia Zhang, Jianguo Lin, and Wei Li. 2022. "Isolation and Characterization of a Biosurfactant Producing Strain Planococcus sp. XW-1 from the Cold Marine Environment" International Journal of Environmental Research and Public Health 19, no. 2: 782. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19020782