
Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
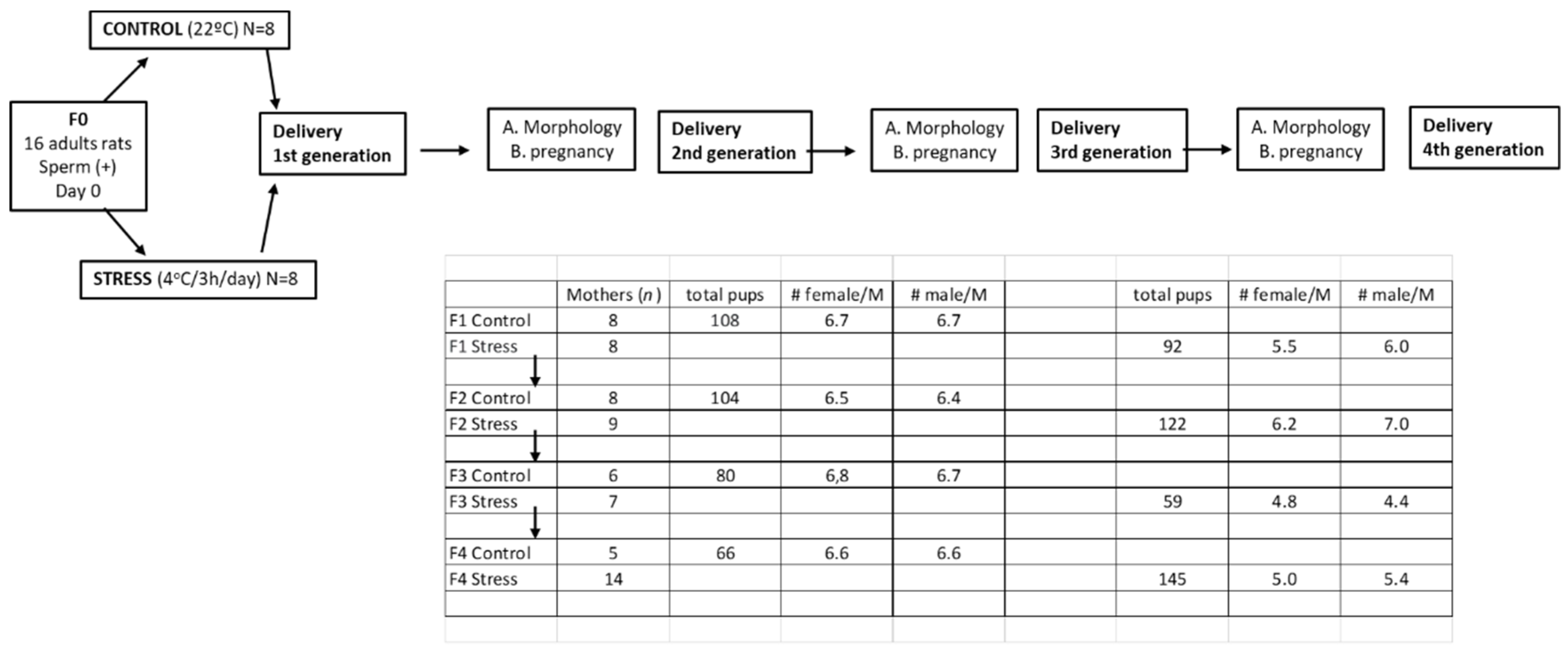
2.1. Animals and Experimental Design
2.2. Gestational Stress Induction
2.3. Studies with Generations
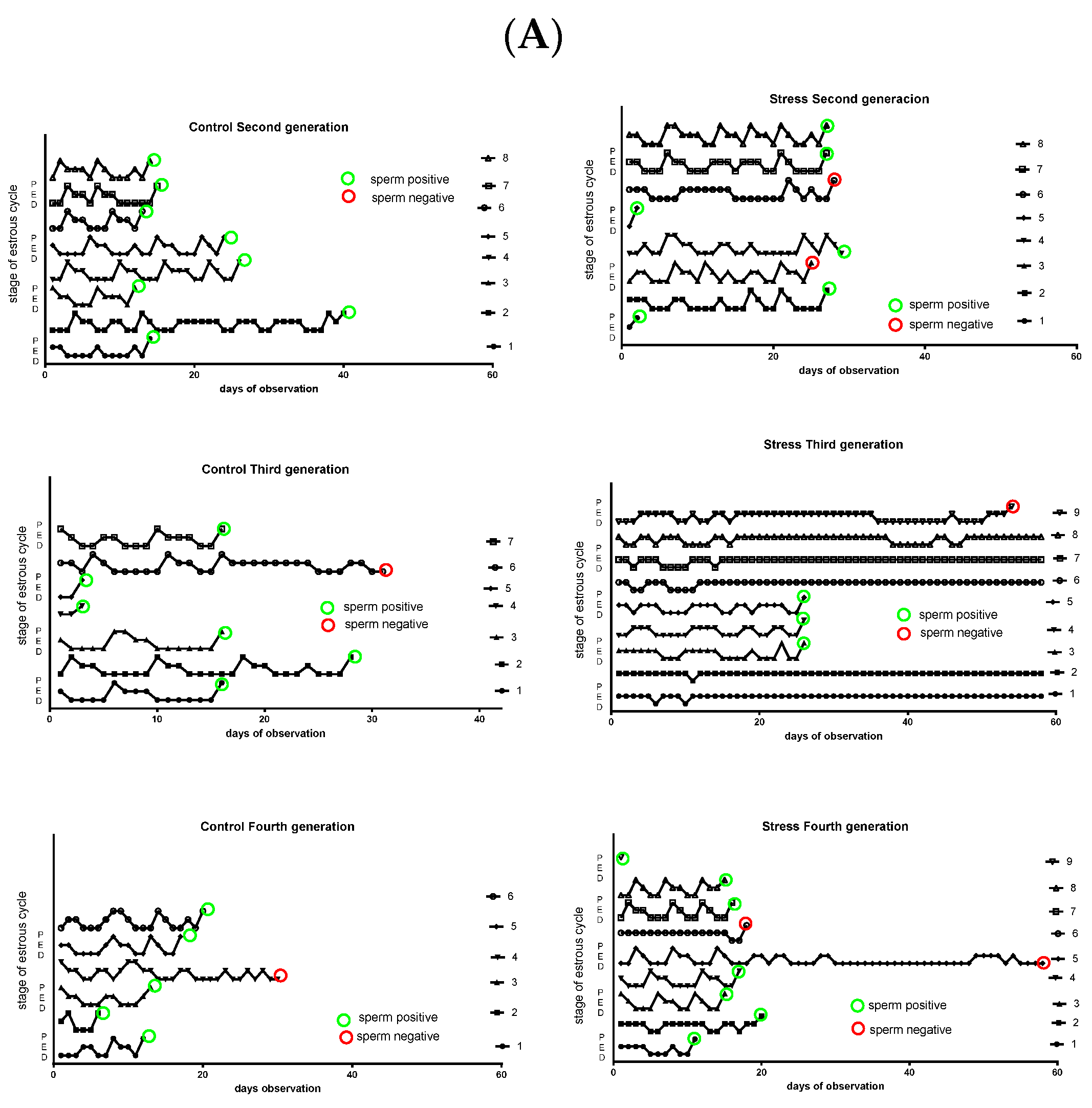
2.4. Estrous Cycling Activities of the Rats
2.5. Morphometric Analysis
2.6. Statistical Analysis
3. Results
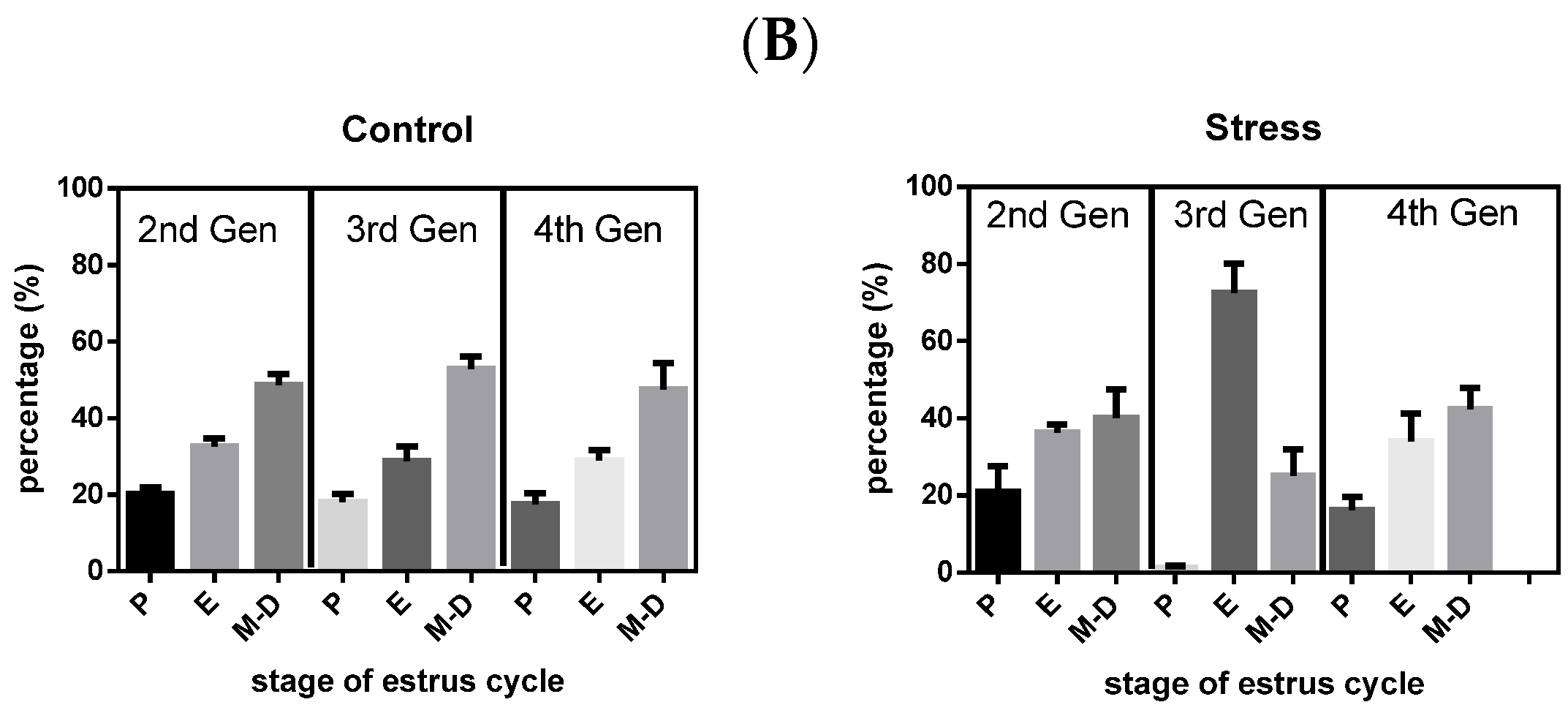
3.1. Effect of Gestational Stress on the Estrus Cycling Activity of the Rats of the Second, Third, and Fourth Generation
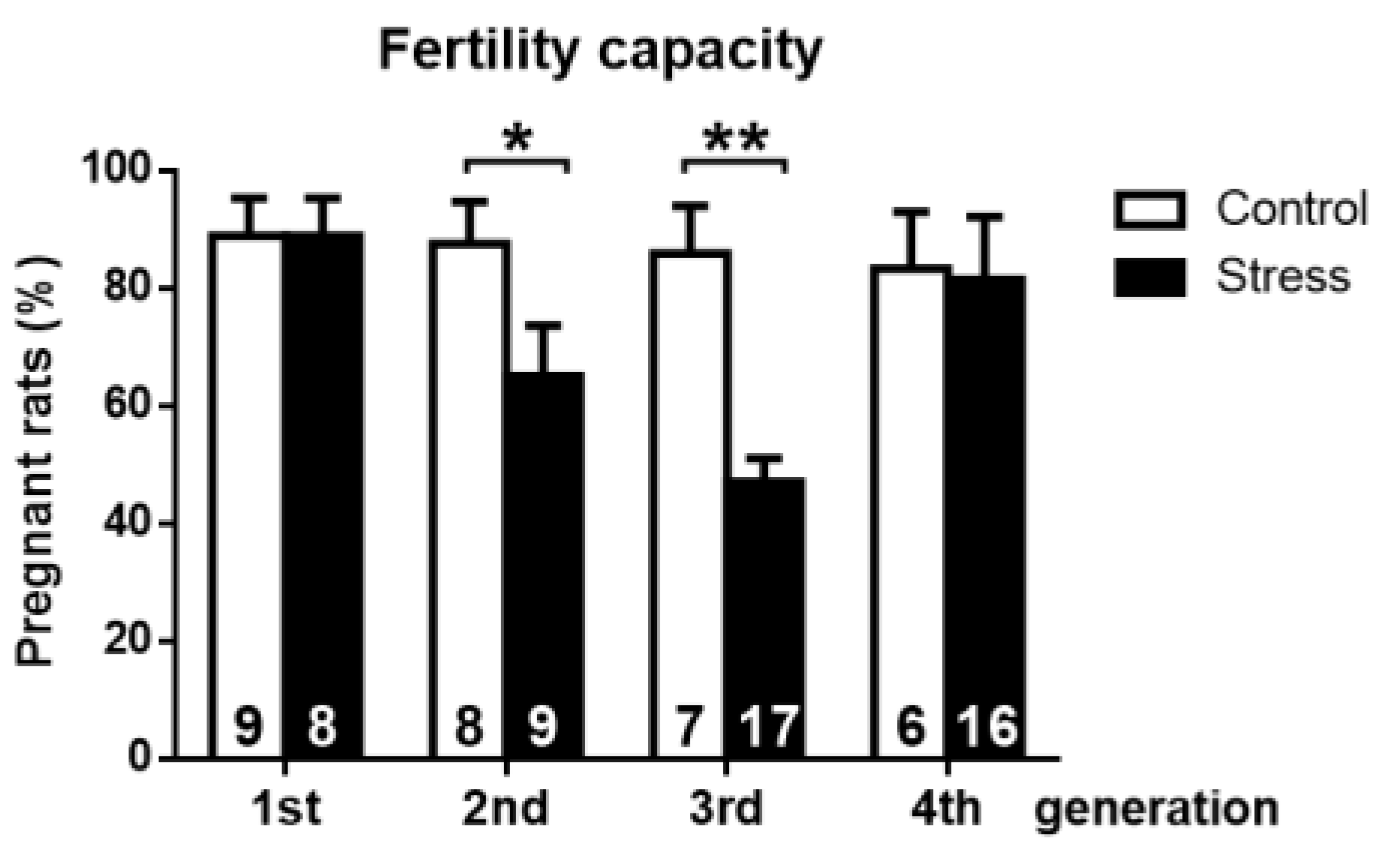
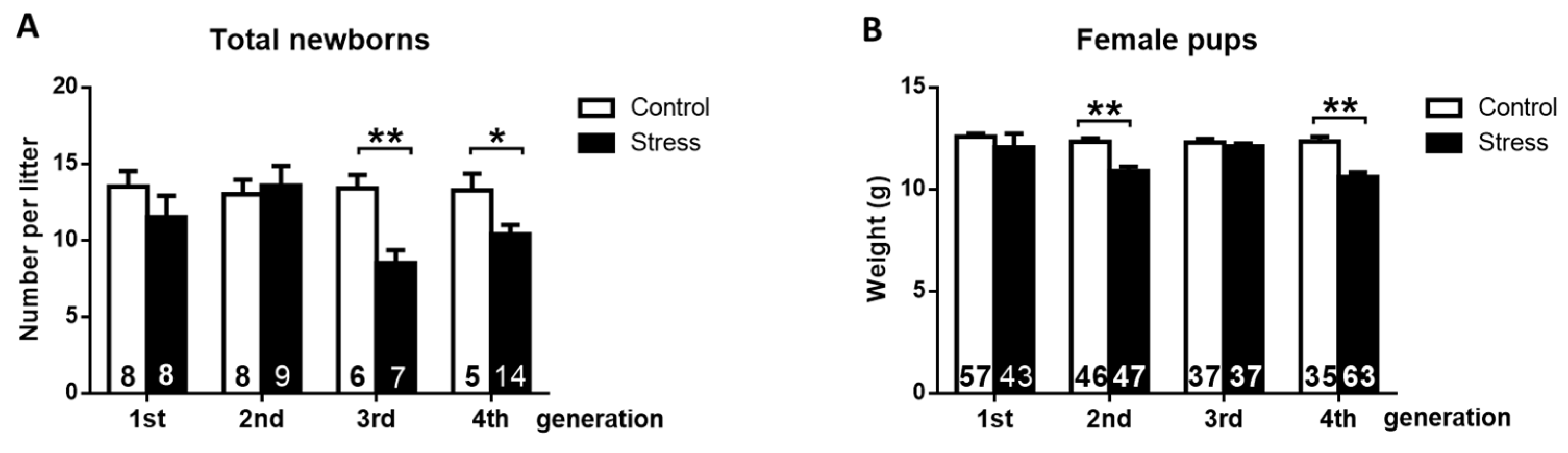
3.2. Effect of Gestational Stress on the Fertile Capacity
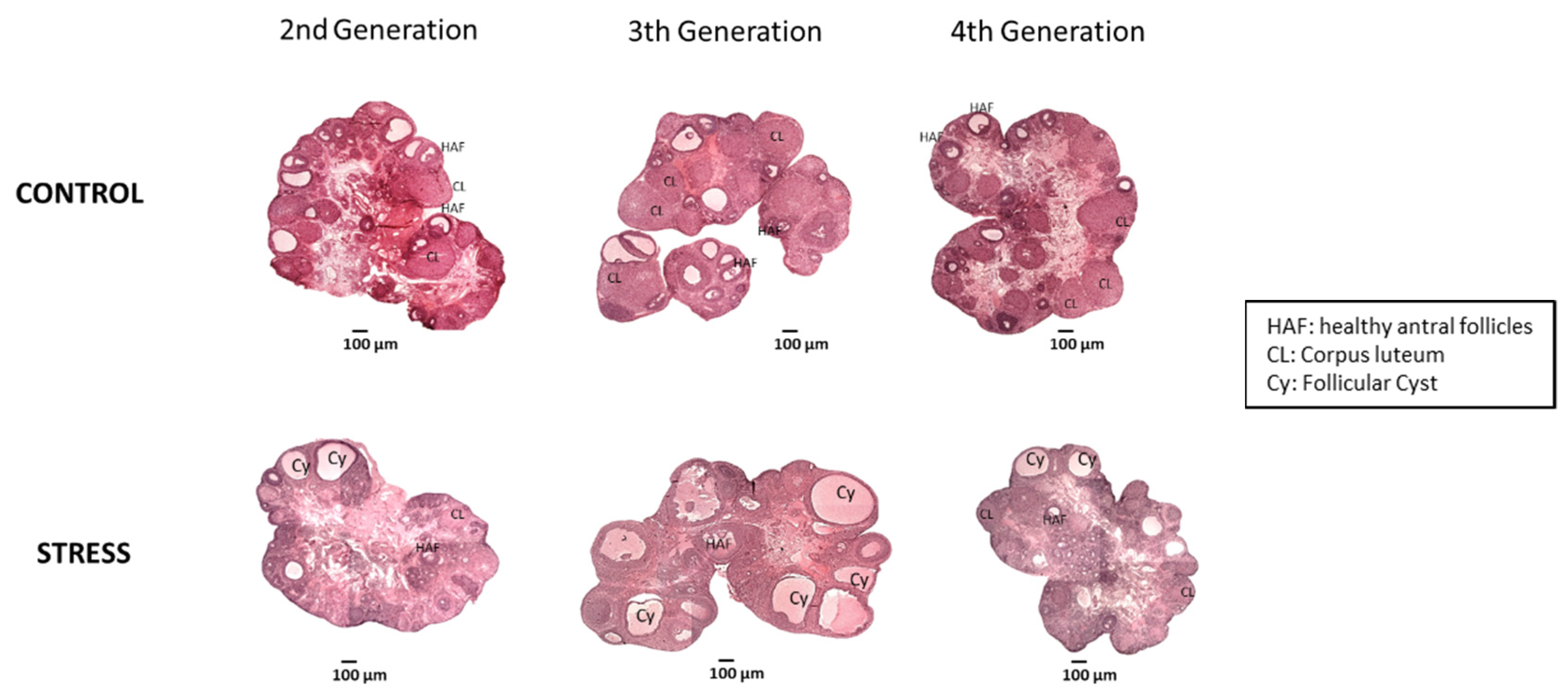
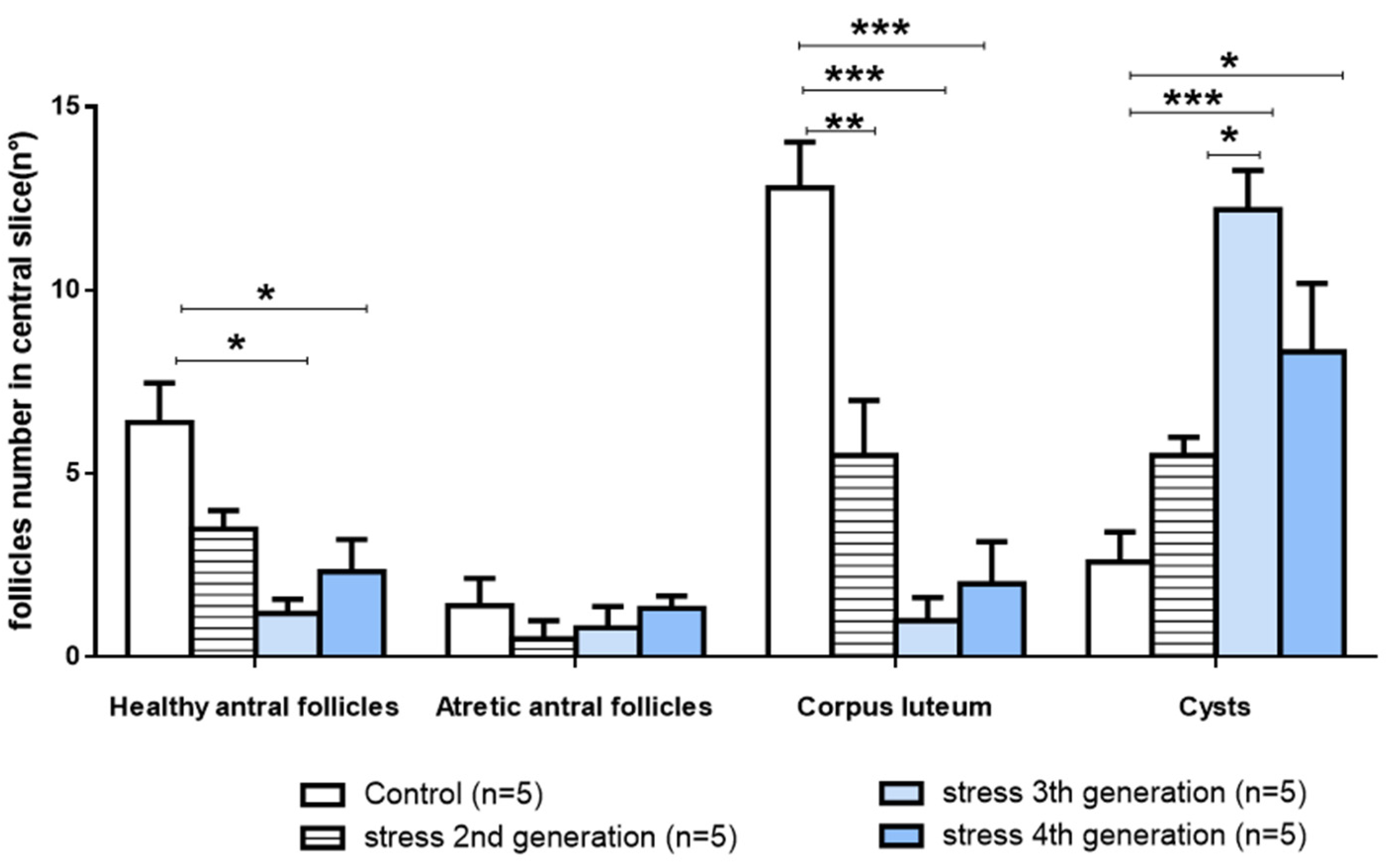
3.3. Effect of Gestational Stress in Follicular Dynamics of Ovary Follicular Development
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cottrell, E.C.; Seckl, J.R. Prenatal stress, glucocorticoids and the programming of adult disease. Front. Behav. Neurosci. 2009, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chales, N.; Barker, D.J. The thrifty phenotype hypothesis: Type 2 diabetes. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef] [Green Version]
- Kajantie, E. Fetal origins of stress-related adult disease. Ann. N. Y. Acad. Sci. 2006, 1083, 11–27. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.I.; Jones, A.; Goulden, P.A. Birth weight, stress, and the metabolic syndrome in adult life. Ann. N. Y. Acad. Sci. 2006, 1083, 28–36. [Google Scholar] [CrossRef]
- Godfrey, K.M.; Barker, D.J. Fetal programming and adult health. Public Health Nutr. 2001, 4, 611–624. [Google Scholar] [CrossRef]
- Goldstein, D.S.; Kopin, I.J. Adrenomedullary, adrenocortical, and sympathoneural responses to stressors: A meta-analysis. Endocr. Regul. 2008, 42, 111–119. [Google Scholar]
- Pacak, K.; Baffi, J.S.; Kvetnansky, R.; Goldstein, D.S.; Palkovits, M. Stressor-specific activation of catecholaminergic systems: Implications for stress-related hypothalamic-pituitary-adrenocortical responses. Adv. Pharmacol. 1998, 42, 561–564. [Google Scholar]
- Benedict, C.R.; Fillenz, M.; Stanford, C. Noradrenaline release in rats during prolonged cold-stress and repeated swim-stress. Br. J. Pharm. 1979, 66, 521–524. [Google Scholar] [CrossRef] [Green Version]
- Lansdown, A.; Rees, D.A. The sympathetic nervous system in polycystic ovary syndrome: A novel therapeutic target? Clin. Endocrinol. 2012, 77, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Medina, G.; Maliqueo, M.; Crisosto, N.; Echiburu, B.; Sir-Petermann, T.; Lara, H.E. Metformin increases norepinephrine transporter expression in placenta of patients with polycystic ovary syndrome. Eur. Rev. Med. Pharm. Sci. 2017, 21, 3482–3489. [Google Scholar]
- Sverrisdottir, Y.B.; Mogren, T.; Kataoka, J.; Janson, P.O.; Stener-Victorin, E. Is polycystic ovary syndrome associated with high sympathetic nerve activity and size at birth? Am. J. Physiol. Endocrinol. Metab. 2008, 294, E576–E581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barria, A.; Leyton, V.; Ojeda, S.R.; Lara, H.E. Ovarian steroidal response to gonadotropins and beta-adrenergic stimulation is enhanced in polycystic ovary syndrome: Role of sympathetic innervation. Endocrinology 1993, 133, 2696–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piquer, B.; Fonseca, J.L.; Lara, H.E. Gestational stress, placental norepinephrine transporter and offspring fertility. Reproduction 2017, 153, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, M.; Arancibia, S.; Fiedler, J.L.; Lara, H.E. Chronic intermittent cold stress activates ovarian sympathetic nerves and modifies ovarian follicular development in the rat. Biol. Reprod. 2003, 68, 2038–2043. [Google Scholar] [CrossRef] [Green Version]
- Dorfman, M.; Ramirez, V.D.; Stener-Victorin, E.; Lara, H.E. Chronic-intermittent cold stress in rats induces selective ovarian insulin resistance. Biol. Reprod. 2009, 80, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Barra, R.; Cruz, G.; Mayerhofer, A.; Paredes, A.; Lara, H.E. Maternal sympathetic stress impairs follicular development and puberty of the offspring. Reproduction 2014, 148, 137–145. [Google Scholar] [CrossRef] [Green Version]
- AVMA. American Veterinary Medical Association Guidelines for the Euthanasia of Animals; 2020 Edition; American Veterinary Medical Association 1931 N.: Schaumburg, IL, USA, 2020. [Google Scholar]
- Urra, J.; Blohberger, J.; Tiszavari, M.; Mayerhofer, A.; Lara, H.E. In vivo blockade of acetylcholinesterase increases intraovarian acetylcholine and enhances follicular development and fertility in the rat. Sci. Rep. 2016, 6, 30129. [Google Scholar] [CrossRef] [Green Version]
- CHubscher, H.; Brooks, D.L.; Johnson, J.R. A quantitative method for assessing stages of the rat estrous cycle. Biotech. Histochem. 2005, 80, 79–87. [Google Scholar] [CrossRef]
- Paccola, C.G.R.C.C.; Stumpp, T.; Miraglia, S.M.; Cipriano, I. The rat estrous cycle revisited: A quantitative and qualitative analysis. Anim. Reprod. 2013, 10, 677–683. [Google Scholar]
- del Campo, M.; Piquer, B.; Witherington, J.; Sridhar, A.; Lara, H.E. Effect of Superior Ovarian Nerve and Plexus Nerve Sympathetic Denervation on Ovarian-Derived Infertility Provoked by Estradiol Exposure to Rats. Front. Physiol. 2019, 10, 349. [Google Scholar] [CrossRef] [Green Version]
- Riquelme, R.; Ruz, F.; Mayerhofer, A.; Lara, H.E. Role of ovarian sympathetic nerves and cholinergic local system during cold stress. J. Endocrinol. 2019, 242, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, A.N.; Midgley, A.R., Jr. Morphometric analysis of follicular development in the rat. Biol. Reprod. 1978, 19, 597–605. [Google Scholar] [CrossRef] [Green Version]
- Lara, H.E.; Dissen, G.A.; Leyton, V.; Paredes, A.; Fuenzalida, H.; Fiedler, J.L.; Ojeda, S.R. An increased intraovarian synthesis of nerve growth factor and its low affinity receptor is a principal component of steroid-induced polycystic ovary in the rat. Endocrinology 2000, 141, 1059–1072. [Google Scholar] [CrossRef] [PubMed]
- Brawer, J.R.; Munoz, M.; Farookhi, R. Development of the polycystic ovarian condition (PCO) in the estradiol valerate-treated rat. Biol. Reprod. 1986, 35, 647–655. [Google Scholar] [CrossRef] [PubMed]
- Zar, J. Biostatistical Analysis, 2nd ed.; Prentice Hall: Hoboken, NJ, USA, 1984. [Google Scholar]
- Pacak, K.; Palkovits, M.; Yadid, G.; Kvetnansky, R.; Kopin, I.J.; Goldstein, D.S. Heterogeneous neurochemical responses to different stressors: A test of Selye’s doctrine of nonspecificity. Am. J. Physiol. 1998, 275, R1247–R1255. [Google Scholar] [CrossRef]
- Bernuci, M.P.; Leite, C.M.; Barros, P.; Kalil, B.; Leoni, G.B.; Bianco-Borges, B.D.; Franci, C.R.; Szawka, R.E.; Lara, H.E.; Anselmo-Franci, J.A. Transitory activation of central and ovarian norepinephrine systems during cold stress-induced polycystic ovary in rats. J. Neuroendocr. 2013, 25, 23–33. [Google Scholar] [CrossRef]
- Bernuci, M.P.; Szawka, R.E.; Helena, C.V.; Leite, C.M.; Lara, H.E.; Anselmo-Franci, J.A. Locus coeruleus mediates cold stress-induced polycystic ovary in rats. Endocrinology 2008, 149, 2907–2916. [Google Scholar] [CrossRef] [Green Version]
- Jimmy, P.D.; Neill, D. Physiology of Reproduction; Elsevier Academic Press: Birmingham, Alabama, 2006. [Google Scholar]
- Toufexis, D.; Rivarola, M.A.; Lara, H.; Viau, V. Stress and the reproductive axis. J. Neuroendocr. 2014, 26, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Ajayi, A.F.; Akhigbe, R.E. Staging of the estrous cycle and induction of estrus in experimental rodents: An update. Fertil. Res. Pr. 2020, 6, 5. [Google Scholar] [CrossRef] [Green Version]
- TBrown, J.; Moore, M.J.; Blaustein, J.D. Maintenance of progesterone-facilitated sexual behavior in female rats requires continued hypothalamic protein synthesis and nuclear progestin receptor occupation. Endocrinology 1987, 121, 298–304. [Google Scholar] [CrossRef]
- Mani, S.K.; Blaustein, J.D.; Allen, J.M.; Law, S.W.; O’Malley, B.W.; Clark, J.H. Inhibition of rat sexual behavior by antisense oligonucleotides to the progesterone receptor. Endocrinology 1994, 135, 1409–1414. [Google Scholar] [CrossRef] [PubMed]
- Jara, P.; Rage, F.; Dorfman, M.; Grouselle, D.; Barra, R.; Arancibia, S.; Lara, H.E. Cold-induced glutamate release in vivo from the magnocellular region of the paraventricular nucleus is involved in ovarian sympathetic activation. J. Neuroendocrinol. 2010, 22, 979–986. [Google Scholar] [CrossRef] [PubMed]
- Mayila, Y.; Matsuzaki, T.; Iwasa, T.; Tungalagsuvd, A.; Munkhzaya, M.; Yano, K.; Yanagihara, R.; Tokui, T.; Minato, S.; Takeda, A.; et al. The reduction in sexual behavior of adult female rats exposed to immune stress in the neonatal period is associated with reduced hypothalamic progesterone receptor expression. Gen. Comp. Endocrinol. 2020, 288, 113360. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, J.; Jara, P.; Luza, S.; Dorfman, M.; Grouselle, D.; Rage, F.; Lara, H.E.; Arancibia, S. Cold stress induces metabolic activation of thyrotrophin-releasing hormone-synthesising neurones in the magnocellular division of the hypothalamic paraventricular nucleus and concomitantly changes ovarian sympathetic activity parameters. J. Neuroendocrinol. 2006, 18, 367–376. [Google Scholar] [CrossRef]
- Beyer, C.; Gonzalez-Flores, O.; Gonzalez-Mariscal, G. Ring a reduced progestins potently stimulate estrous behavior in rats: Paradoxical effect through the progesterone receptor. Physiol. Behav. 1995, 58, 985–993. [Google Scholar] [CrossRef]
- Martinez-Pinto, J.; Piquer, B.; Tiszavari, M.; Lara, H.E. Neonatal exposure to estradiol valerate reprograms the rat ovary androgen receptor and anti-Mullerian hormone to a polycystic ovary phenotype. Reprod. Toxicol. 2018, 75, 127–135. [Google Scholar] [CrossRef]
- Wang, D.; Cheng, X.; Fang, H.; Ren, Y.; Li, X.; Ren, W.; Xue, B.; Yang, C. Effect of cold stress on ovarian & uterine microcirculation in rats and the role of endothelin system. Reprod. Biol. Endocrinol. 2020, 18, 29. [Google Scholar] [CrossRef] [Green Version]
- Paredes, A.; Galvez, A.; Leyton, V.; Aravena, G.; Fiedler, J.L.; Bustamante, D.; Lara, H.E. Stress promotes development of ovarian cysts in rats: The possible role of sympathetic nerve activation. Endocrine 1998, 8, 309–315. [Google Scholar] [CrossRef]
- Sir-Petermann, T.; Codner, E.; Maliqueo, M.; Echiburu, B.; Hitschfeld, C.; Crisosto, N.; Perez-Bravo, F.; Recabarren, S.E.; Cassorla, F. Increased anti-Mullerian hormone serum concentrations in prepubertal daughters of women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2006, 91, 3105–3109. [Google Scholar] [CrossRef]
- Sir-Petermann, T.; Hitchsfeld, C.; Codner, E.; Maliqueo, M.; Iniguez, G.; Echiburu, B.; Sanchez, F.; Crisosto, N.; Cassorla, F. Gonadal function in low birth weight infants: A pilot study. J. Pediatr. Endocrinol. Metab. 2007, 20, 405–414. [Google Scholar] [CrossRef]
- Sir-Petermann, T.; Maliqueo, M.; Angel, B.; Lara, H.E.; Perez-Bravo, F.; Recabarren, S.E. Maternal serum androgens in pregnant women with polycystic ovarian syndrome: Possible implications in prenatal androgenization. Hum. Reprod. 2002, 17, 2573–2579. [Google Scholar] [CrossRef] [PubMed]
- Sir-Petermann, T.; Maliqueo, M.; Codner, E.; Echiburu, B.; Crisosto, N.; Perez, V.; Perez-Bravo, F.; Cassorla, F. Early metabolic derangements in daughters of women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2007, 92, 4637–4642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumesic, D.A.; Padmanabhan, V.; Chazenbalk, G.D.; Abbott, D.H. Polycystic ovary syndrome as a plausible evolutionary outcome of metabolic adaptation. Reprod. Biol. Endocrinol. 2022, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Mimouni, N.E.H.; Paiva, I.; Barbotin, A.L.; Timzoura, F.E.; Plassard, D.; Le Gras, S.; Ternier, G.; Pigny, P.; Catteau-Jonard, S.; Simon, V.; et al. Polycystic ovary syndrome is transmitted via a transgenerational epigenetic process. Cell Metab. 2021, 33, 513–530.e518. [Google Scholar] [CrossRef]
- Nilsson, E.E.; Sadler-Riggleman, I.; Skinner, M.K. Environmentally induced epigenetic transgenerational inheritance of disease. Environ. Epigenet. 2018, 4, dvy016. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piquer, B.; Ruz, F.; Barra, R.; Lara, H.E. Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study. Int. J. Environ. Res. Public Health 2022, 19, 3044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19053044
Piquer B, Ruz F, Barra R, Lara HE. Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study. International Journal of Environmental Research and Public Health. 2022; 19(5):3044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19053044
Chicago/Turabian StylePiquer, Beatriz, Freddy Ruz, Rafael Barra, and Hernan E. Lara. 2022. "Gestational Sympathetic Stress Programs the Fertility of Offspring: A Rat Multi-Generation Study" International Journal of Environmental Research and Public Health 19, no. 5: 3044. https://0-doi-org.brum.beds.ac.uk/10.3390/ijerph19053044