
Porcine Adipose-Derived Mesenchymal Stem Cells Retain Their Stem Cell Characteristics and Cell Activities While Enhancing the Expression of Liver-Specific Genes after Acute Liver Failure


Abstract
:1. Introduction
2. Results
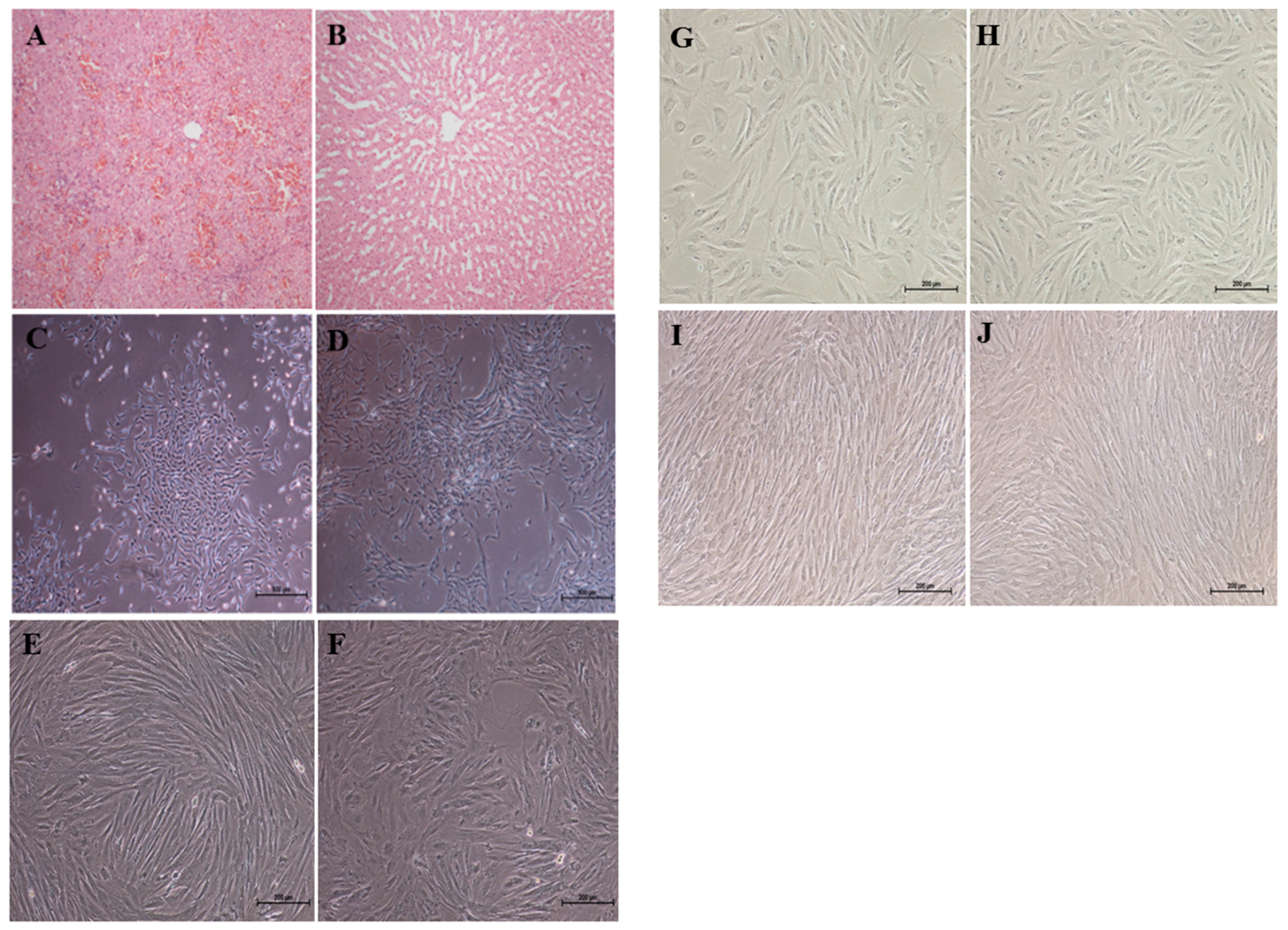
2.1. Cell Morphology
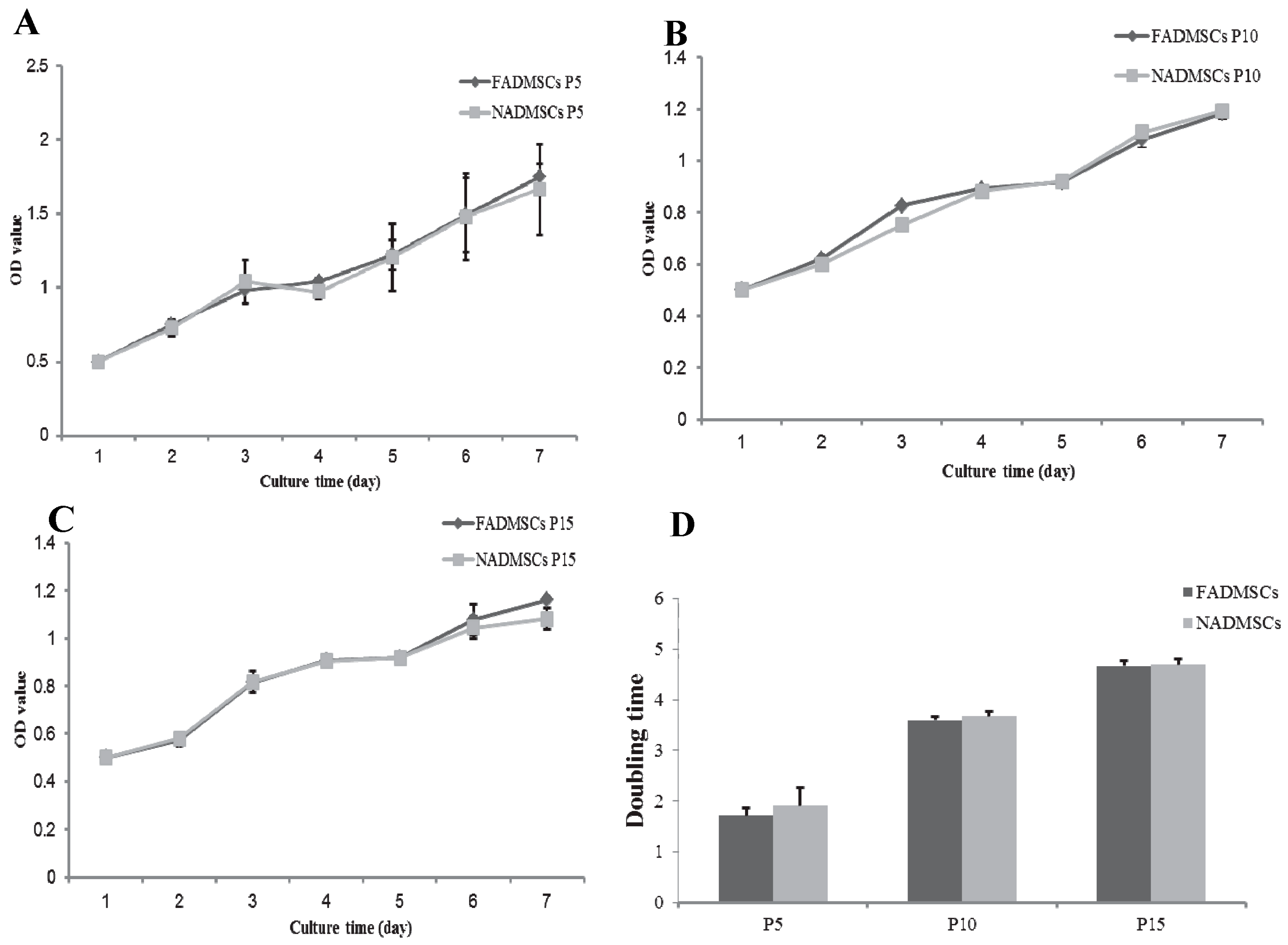
2.2. Cell Proliferative Capacity


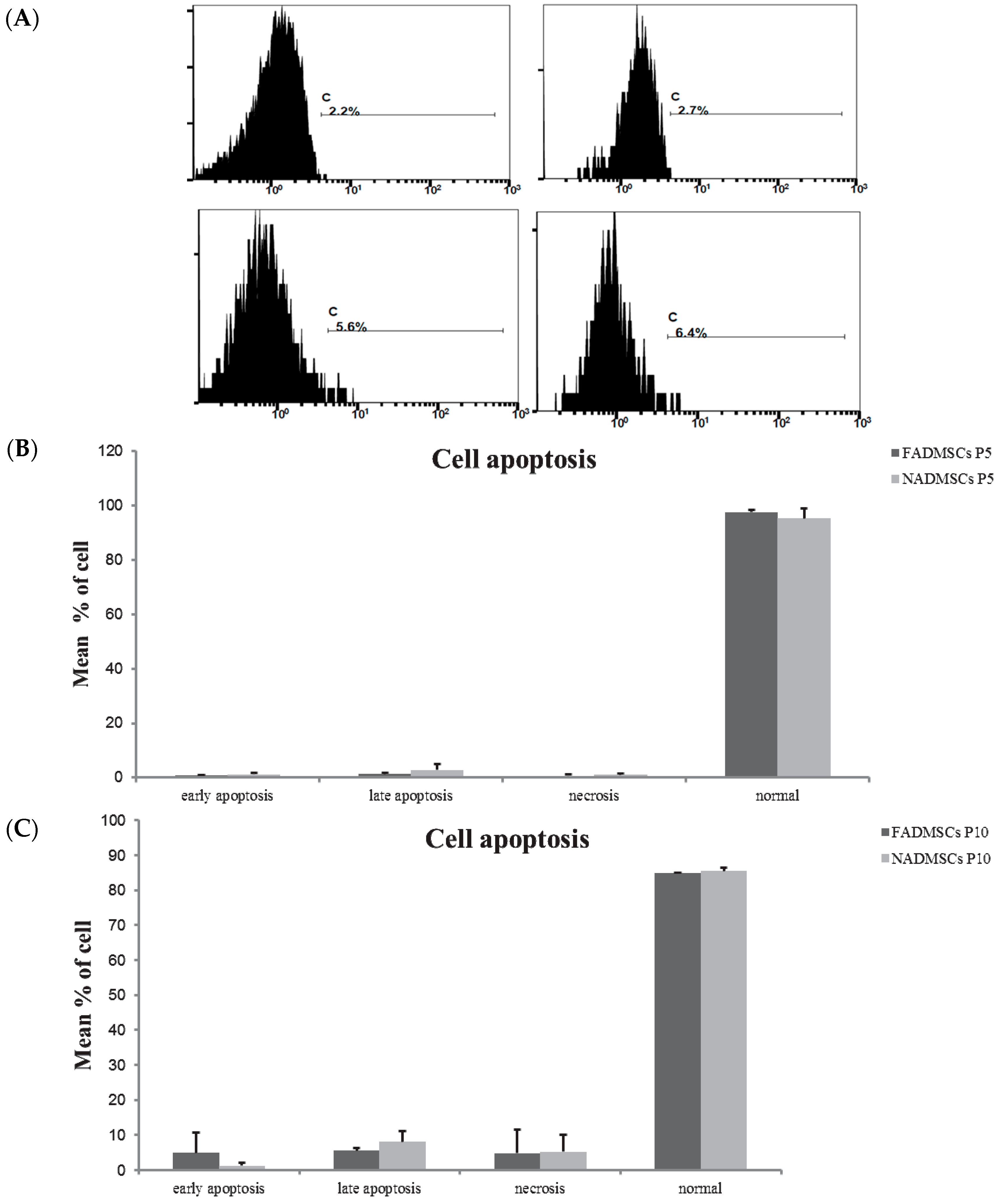
2.3. Cell Apoptosis

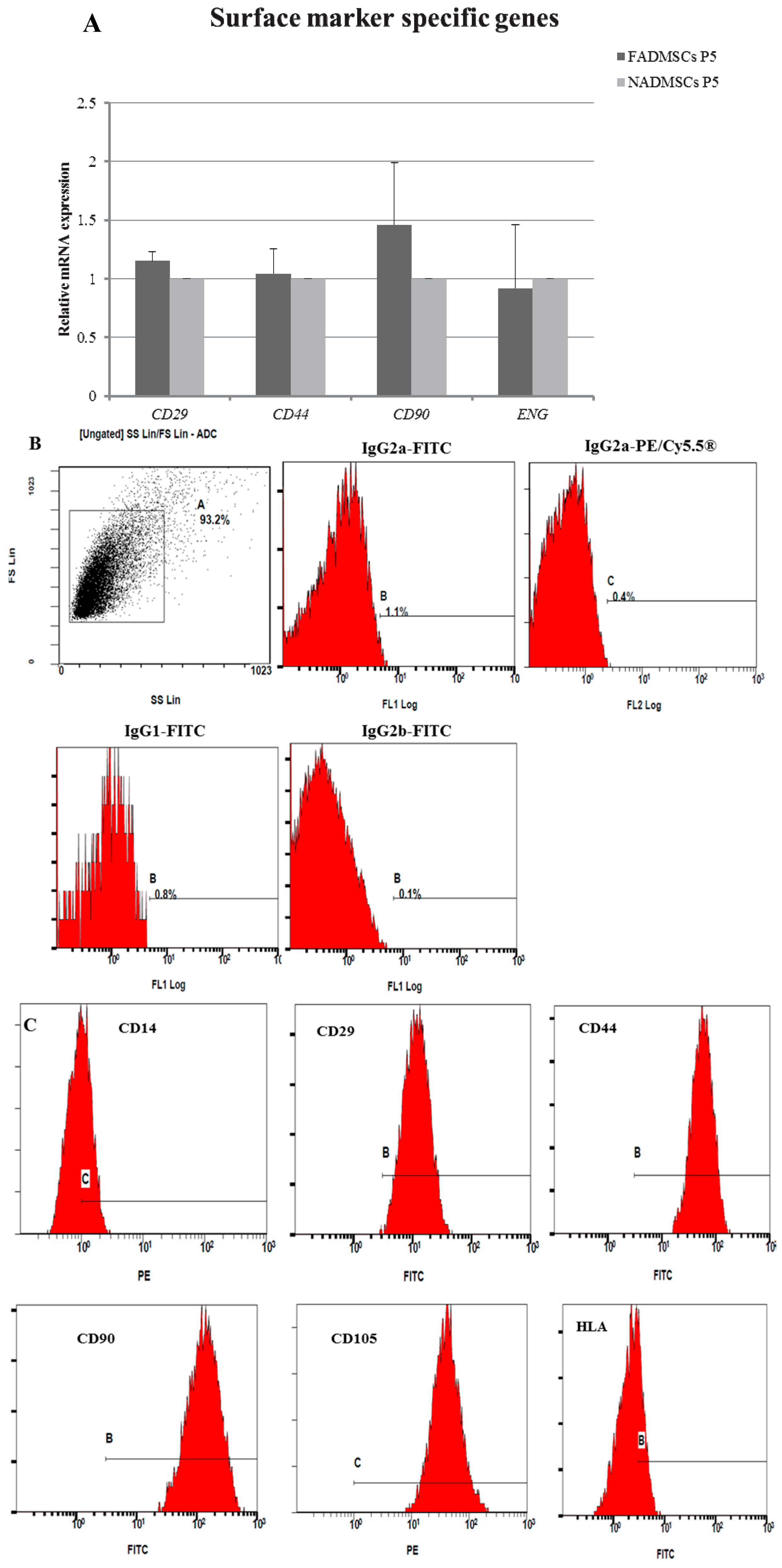
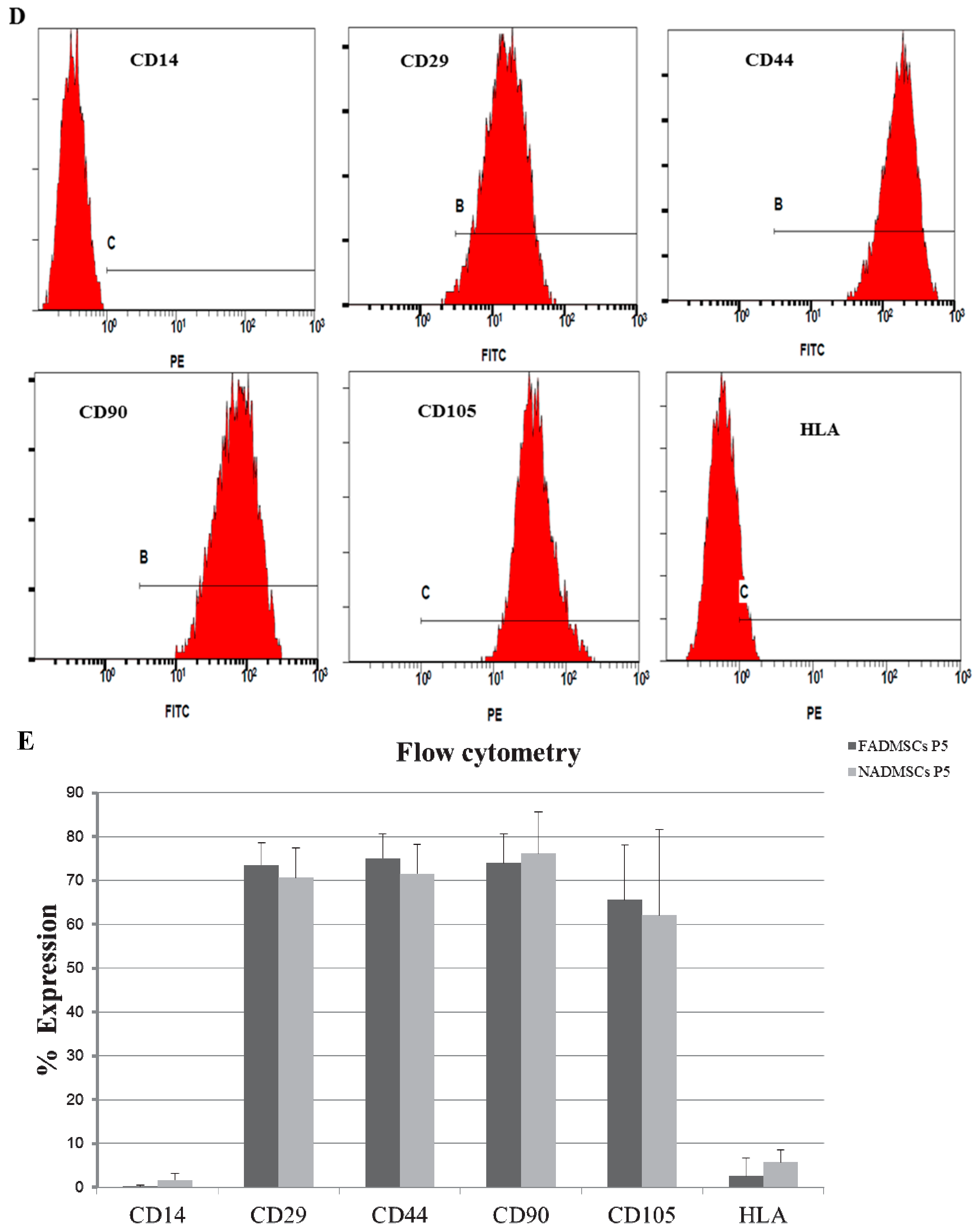
2.4. Surface Antigen Expression


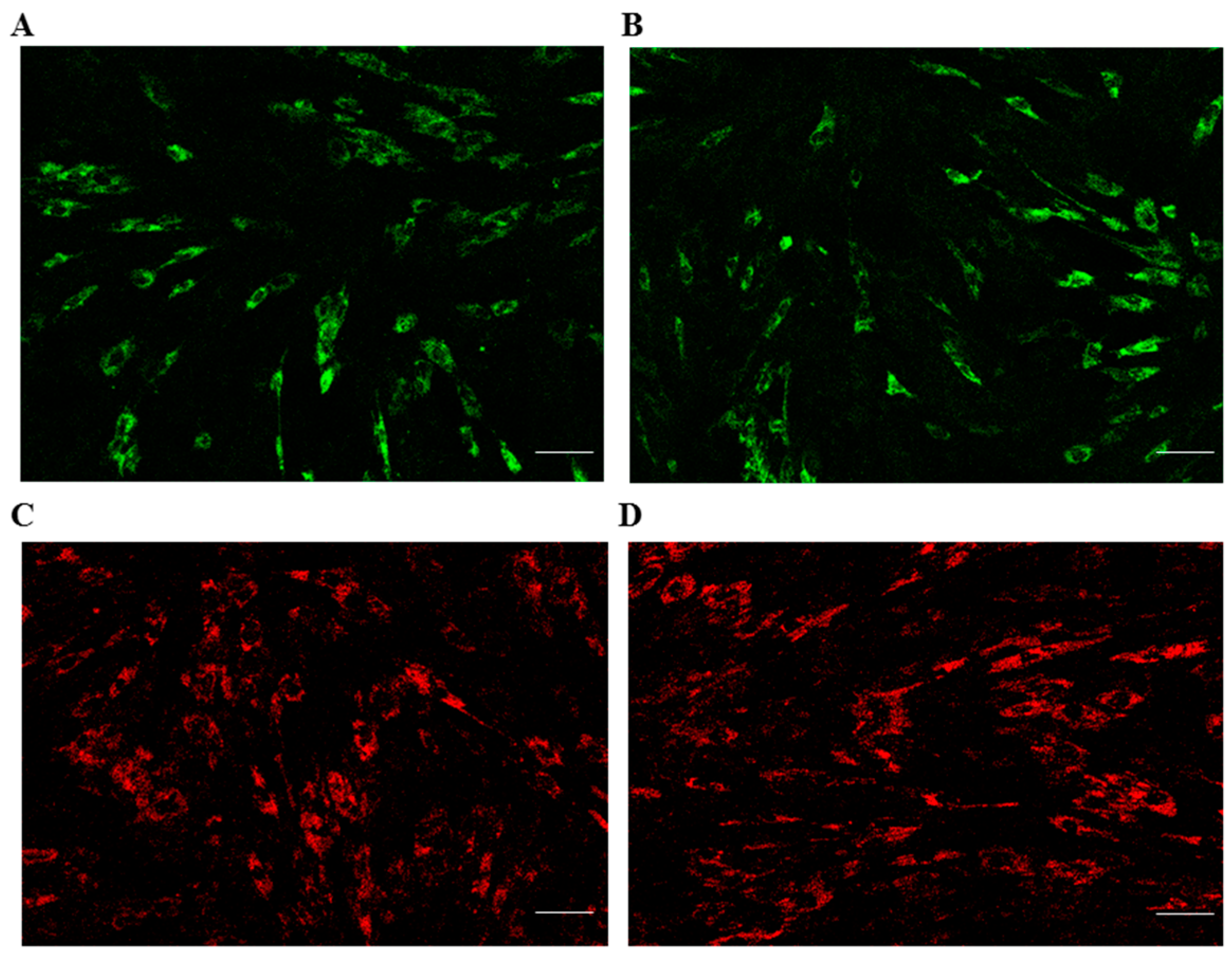
2.5. Mitochondrial and Lysosomal Activities

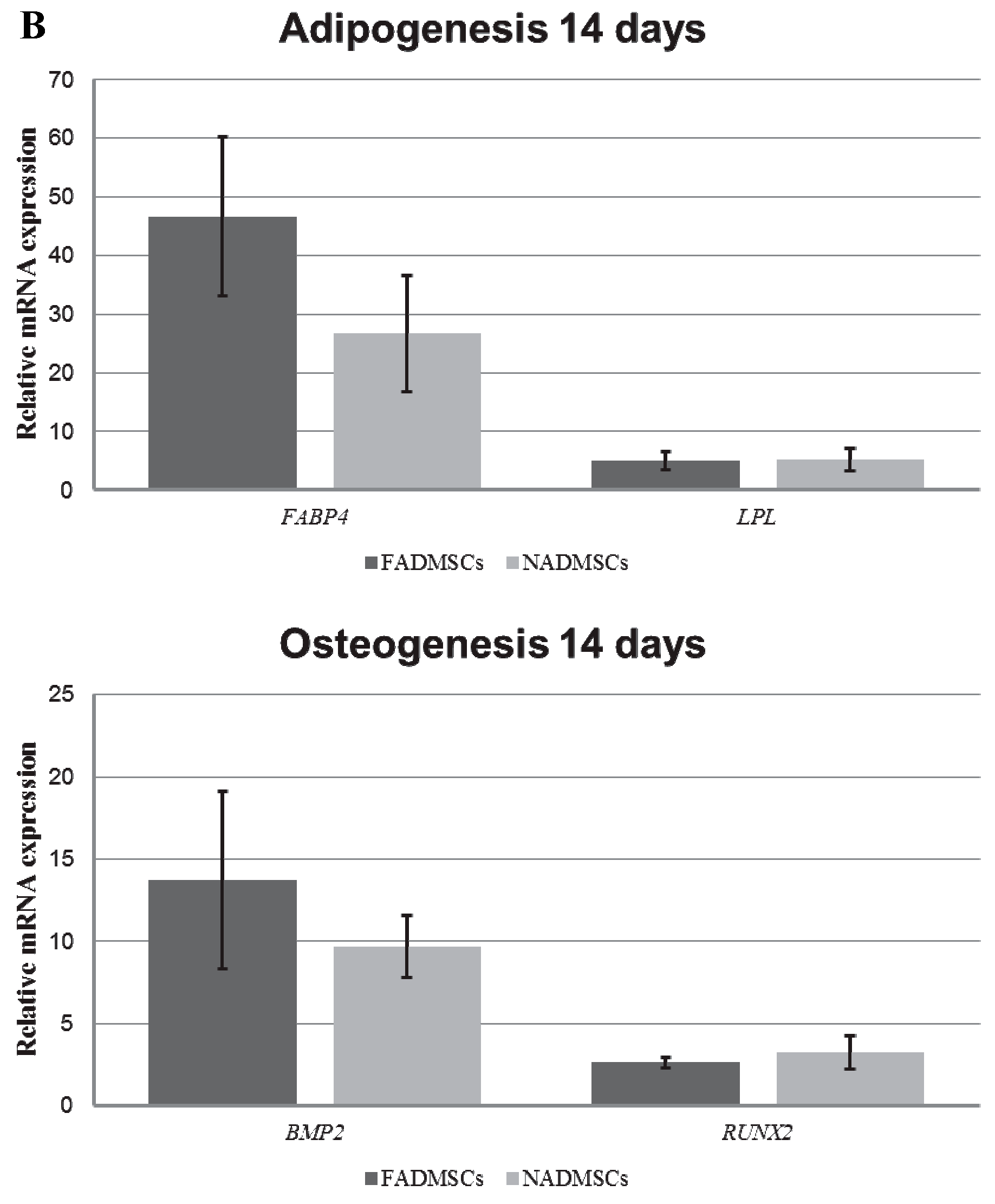
2.6. Multilineage Potency


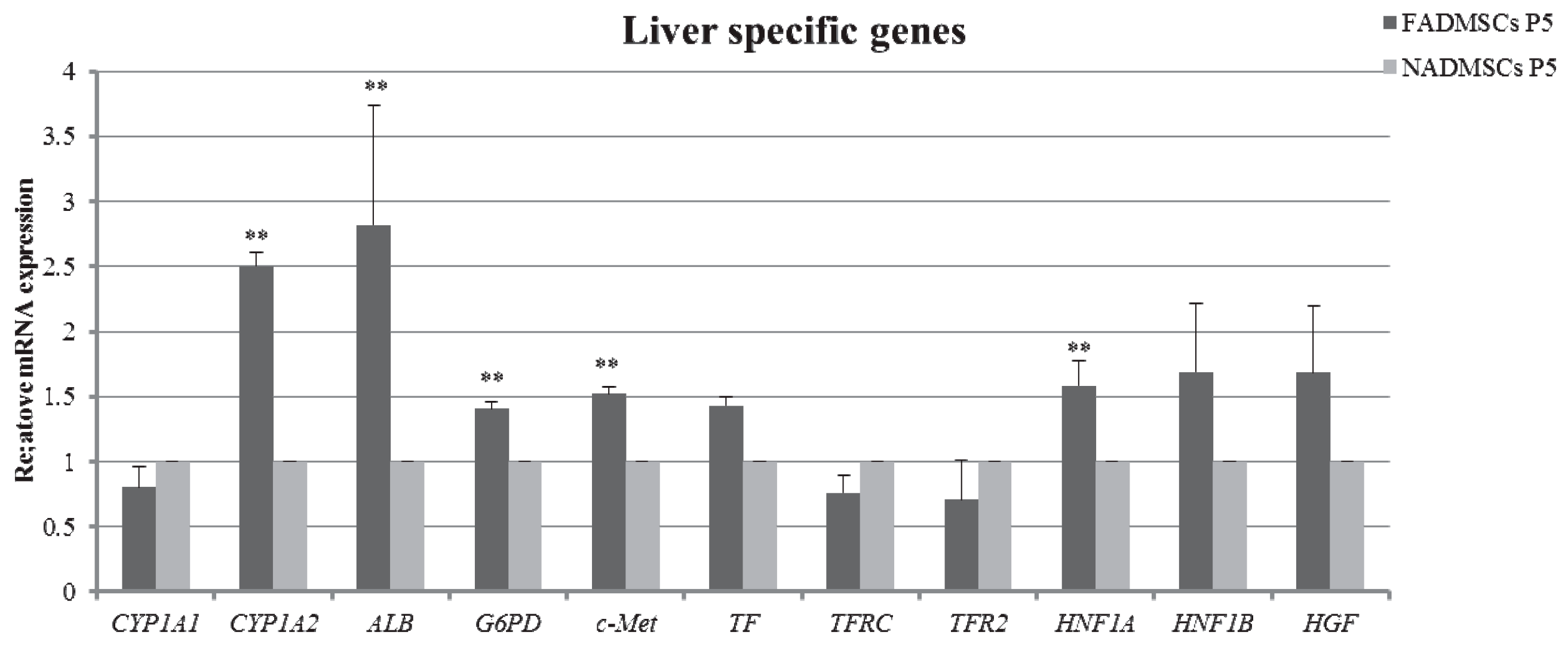
2.7. Liver-Specific Gene Expression

3. Discussion
4. Experimental Section
4.1. Animals
4.2. d-Galactosamine-Induced ALF Model and Immunohistochemistry
4.3. Isolation, in Vitro Culture, and Morphology of ADMSCs
4.4. Analysis of Cell Proliferation
4.5. Detection of Cellular Apoptosis
4.6. Flow Cytometry Analysis
4.7. Measurement of Mitochondrial and Lysosomal Fluorescence
4.8. Adipogenic Differentiation and Osteogenic Differentiation
4.9. RNA Extraction, Reverse Transcription (RT) and Real-Time Quantitative-PCR (RT-QPCR)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | NCBI Accession No. | Product Size (bp) | Forward | Reverse |
|---|---|---|---|---|
| CD29 | NM_213968.1 | 131 | 5′-CACTGCTGCTCATTTGGAAG-3′ | 5′-GGTTGTCACGGCACTCTTAT-3′ |
| CD44 | XM_003122867.4 | 109 | 5′-TGGAAGAGAGAAAGCCAAGC-3′ | 5′-GCCGTCATAAACTGGTCTGG-3′ |
| CD90 | NM_001146129.1 | 125 | 5′-GGCATCGCTCTCTTGCTAAC-3′ | 5′-GGCAGGTTGGTGGTATTCTC-3′ |
| ENG | NM_214031.1| | 121 | 5′-AGGTTTCTGAGGGCTGTGTG-3′ | 5′-TTTGGGTTGGTCATCTGGAC-3′ |
| CYP3A4 | NM_214423.1 | 220 | 5′-TTACACTTACCTGCCCTTTGG-3′ | 5′-TCCACTTACGGTCCCATCTC-3′ |
| ALB | NM_001005208.1 | 241 | 5′-GCCTCTTGTGGATGAGCCTA-3′ | 5′-GTTCAGGACCAGGGACAGAT-3′ |
| G6PD | XM_003360515.3 | 104 | 5′-GATCTACCGCATCGACCACT-3′ | 5′-TGTTGTCTCGGTTCCAGATG-3′ |
| c-MET | NM_001038008.1 | 113 | 5′-GTGGCTGGGACTTTGGATT-3′ | 5′-CGTGTTTGTCGTGCTCTCAC-3′ |
| TF | NM_001244653.1 | 101 | 5′-GAGGCCAATAAGTGCTCCAG-3′ | 5′-ATGCAATCCAGGTAGGAGGA-3′ |
| TFRC | NM_214001.1 | 111 | 5′-TCCCTCAAACACCTCGCTTA-3′ | 5′-GGGCATAATCTTCATTCAGCA-3′ |
| TFR2 | XM_003124374.3 | 108 | 5′-TGATAAGTTCCACGCCAAGA-3′ | 5′-ACCTGCTCGTAAAGGGTCTG-3′ |
| HNF1A | NM_001032388.1 | 103 | 5′-AGGCTCGTGATTCTGCACTT-3′ | 5′-TTTGGCCTTACTGCCTTCTG-3′ |
| HNF1B | NM_213956.1 | 106 | 5′-TGTCTACCTTGTGCTCCTTCG-3′ | 5′-CAGTGTGTTTGGCTCAGTTCA-3′ |
| HGF | XM_003130222.3 | 104 | 5′-ATGCGAGGGAGATTATGGTG-3′ | 5′-GACGATTTGGAATGGCACA-3′ |
| GAPDH | NM_001206359.1 | 154 | 5′-ATGGTGAAGGTCGGAGTGAA-3′ | 5′-CGTGGGTGGAATCATACTGG-3′ |
4.10. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Forbes, S.J.; Gupta, S.; Dhawan, A. Cell therapy for liver disease: From liver transplantation to cell factory. J. Hepatol. 2015, 62, S157–S169. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Li, L. In vitro and in vivo hepatic differentiation of adult somatic stem cells and extraembryonic stem cells for treating end stage liver diseases. Stem Cells Int. 2015, 2015, 871972. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Piao, J.; Jin, L.; Zhou, Y. Does pretreatment of bone marrow mesenchymal stem cells with 5-azacytidine or double intravenous infusion improve their therapeutic potential for dilated cardiomyopathy? Med. Sci. Monit. Basic Res. 2013, 19, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Fraser, J.K.; Zhu, M.; Wulur, I.; Alfonso, Z. Adipose-derived stem cells. Methods Mol. Biol. 2008, 449, 59–67. [Google Scholar] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Seki, T.; Yokoyama, Y.; Nagasaki, H.; Kokuryo, T.; Nagino, M. Adipose tissue-derived mesenchymal stem cell transplantation promotes hepatic regeneration after hepatic ischemia-reperfusion and subsequent hepatectomy in rats. J. Surg. Res. 2012, 178, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Seki, A.; Sakai, Y.; Komura, T.; Nasti, A.; Yoshida, K.; Higashimoto, M.; Honda, M.; Usui, S.; Takamura, M.; Takamura, T.; et al. Adipose tissue-derived stem cells as a regenerative therapy for a mouse steatohepatitis-induced cirrhosis model. Hepatology 2013, 58, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Park, K.C.; Lee, J.U.; Kim, K.J.; Kim, D.G. Therapeutic potential of adipose tissue-derived stem cells for liver failure according to the transplantation routes. J. Korean Surg. Soc. 2011, 81, 176–186. [Google Scholar] [CrossRef] [PubMed]
- Kubo, N.; Narumi, S.; Kijima, H.; Mizukami, H.; Yagihashi, S.; Hakamada, K.; Nakane, A. Efficacy of adipose tissue-derived mesenchymal stem cells for fulminant hepatitis in mice induced by concanavalin A. J. Gastroenterol. Hepatol. 2012, 27, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Usunier, B.; Benderitter, M.; Tamarat, R.; Chapel, A. Management of fibrosis: The mesenchymal stromal cells breakthrough. Stem Cells Int. 2014, 2014, 340257. [Google Scholar] [CrossRef] [PubMed]
- Swiderska-Syn, M.; Syn, W.K.; Xie, G.; Kruger, L.; Machado, M.V.; Karaca, G.; Michelotti, G.A.; Choi, S.S.; Premont, R.T.; Diehl, A.M. Myofibroblastic cells function as progenitors to regenerate murine livers after partial hepatectomy. Gut 2014, 63, 1333–1344. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Qu, B.; Huo, J.H.; Wu, S.L.; Zhang, M.Y.; Wang, Z.R. Serum from radiofrequency-injured livers induces differentiation of bone marrow stem cells into hepatocyte-like cells. J. Surg. Res. 2009, 155, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Yannam, G.R.; Han, B.; Setoyama, K.; Yamamoto, T.; Ito, R.; Brooks, J.M.; Guzman-Lepe, J.; Galambos, C.; Fong, J.V.; Deutsch, M.; et al. A nonhuman primate model of human radiation-induced venocclusive liver disease and hepatocyte injury. Int. J. Radiat. Oncol. Biol. Phys. 2014, 88, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Vainshtein, J.M.; Kabarriti, R.; Mehta, K.J.; Roy-Chowdhury, J.; Guha, C. Bone marrow-derived stromal cell therapy in cirrhosis: Clinical evidence, cellular mechanisms, and implications for the treatment of hepatocellular carcinoma. Int. J. Radiat. Oncol. Biol. Phys. 2014, 89, 786–803. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Mejia, V.; Bueno, C.; Roldan, M.; Sanchez, L.; Ligero, G.; Menendez, P.; Martin, M. The adaptation of human embryonic stem cells to different feeder-free culture conditions is accompanied by a mitochondrial response. Stem Cells Dev. 2012, 21, 1145–1155. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.Y.; Liou, C.W.; Chen, S.D.; Hsu, T.Y.; Chuang, J.H.; Wang, P.W.; Huang, S.T.; Tiao, M.M.; Chen, J.B.; Lin, T.K.; et al. Mitochondrial transfer from Wharton’s jelly-derived mesenchymal stem cells to mitochondria-defective cells recaptures impaired mitochondrial function. Mitochondrion 2015, 22, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Ohkuma, S.; Moriyama, Y.; Takano, T. Identification and characterization of a proton pump on lysosomes by fluorescein-isothiocyanate-dextran fluorescence. Proc. Natl. Acad. Sci. USA 1982, 79, 2758–2762. [Google Scholar] [CrossRef] [PubMed]
- Fogel, J.L.; Thein, T.Z.; Mariani, F.V. Use of LysoTracker to detect programmed cell death in embryos and differentiating embryonic stem cells. J. Vis. Exp. 2012, 68. [Google Scholar] [CrossRef] [PubMed]
- Rallapalli, S.; Bishi, D.K.; Verma, R.S.; Cherian, K.M.; Guhathakurta, S. A multiplex PCR technique to characterize human bone marrow derived mesenchymal stem cells. Biotechnol. Lett. 2009, 31, 1843–1850. [Google Scholar] [CrossRef] [PubMed]
- Radcliffe, C.H.; Flaminio, M.J.; Fortier, L.A. Temporal analysis of equine bone marrow aspirate during establishment of putative mesenchymal progenitor cell populations. Stem Cells Dev. 2010, 19, 269–282. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, S.; Tautenhahn, H.M.; Winkler, S.; Stock, P.; Jonas, S.; Dollinger, M.; Christ, B. Isolation and hepatocyte differentiation of mesenchymal stem cells from porcine bone marrow—“Surgical waste” as a novel MSC source. Transplant. Proc. 2013, 45, 2056–2058. [Google Scholar] [CrossRef] [PubMed]
- Ranera, B.; Lyahyai, J.; Romero, A.; Vazquez, F.J.; Remacha, A.R.; Bernal, M.L.; Zaragoza, P.; Rodellar, C.; Martin-Burriel, I. Immunophenotype and gene expression profiles of cell surface markers of mesenchymal stem cells derived from equine bone marrow and adipose tissue. Vet. Immunol. Immunopathol. 2011, 144, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Noort, W.A.; Oerlemans, M.I.; Rozemuller, H.; Feyen, D.; Jaksani, S.; Stecher, D.; Naaijkens, B.; Martens, A.C.; Buhring, H.J.; Doevendans, P.A.; et al. Human versus porcine mesenchymal stromal cells: Phenotype, differentiation potential, immunomodulation and cardiac improvement after transplantation. J. Cell. Mol. Med. 2012, 16, 1827–1839. [Google Scholar] [CrossRef] [PubMed]
- Wanet, A.; Arnould, T.; Najimi, M.; Renard, P. Connecting mitochondria, metabolism, and stem cell fate. Stem Cells Dev. 2015, 24, 1957–1971. [Google Scholar] [CrossRef] [PubMed]
- Prigione, A.; Adjaye, J. Modulation of mitochondrial biogenesis and bioenergetic metabolism upon in vitro and in vivo differentiation of human ES and iPS cells. Int. J. Dev. Biol. 2010, 54, 1729–1741. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Lindgren, A.G.; Srivastava, A.S.; Clark, A.T.; Banerjee, U. Mitochondrial function controls proliferation and early differentiation potential of embryonic stem cells. Stem Cells 2011, 29, 486–495. [Google Scholar] [CrossRef] [PubMed]
- Bukowiecki, R.; Adjaye, J.; Prigione, A. Mitochondrial function in pluripotent stem cells and cellular reprogramming. Gerontology 2014, 60, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Tao, R.; Wu, W.; Cao, H.; Xin, J.; Guo, J.; Jiang, L.; Hong, X.; Demetriou, A.A.; Farkas, D.; et al. Transcriptional profiling and hepatogenic potential of acute hepatic failure-derived bone marrow mesenchymal stem cells. Differentiation 2010, 80, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, W.; Xin, J.; Guo, J.; Jiang, L.; Tao, R.; Cao, H.; Hong, X.; Li, L. Acute hepatic failure-derived bone marrow mesenchymal stem cells express hepatic progenitor cell genes. Cells Tissues Organs 2011, 194, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Mohsin, S.; Shams, S.; Ali Nasir, G.; Khan, M.; Javaid Awan, S.; Khan, S.N.; Riazuddin, S. Enhanced hepatic differentiation of mesenchymal stem cells after pretreatment with injured liver tissue. Differentiation 2011, 81, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yuan, Y.; Qin, H.; Ying, C.; Liu, W.; Zhang, J.; He, Y.; Liu, Z. Serum from hepatectomized rats induces the differentiation of adipose tissue mesenchymal stem cells into hepatocyte-like cells and upregulates the expression of hepatocyte growth factor and interleukin-6 in vitro. Int. J. Mol. Med. 2013, 31, 667–675. [Google Scholar] [PubMed]
- Katoonizadeh, A.; Nevens, F.; Verslype, C.; Pirenne, J.; Roskams, T. Liver regeneration in acute severe liver impairment: A clinicopathological correlation study. Liver Int. 2006, 26, 1225–1233. [Google Scholar] [CrossRef] [PubMed]
- Krampera, M.; Cosmi, L.; Angeli, R.; Pasini, A.; Liotta, F.; Andreini, A.; Santarlasci, V.; Mazzinghi, B.; Pizzolo, G.; Vinante, F.; et al. Role for interferon-gamma in the immunomodulatory activity of human bone marrow mesenchymal stem cells. Stem Cells 2006, 24, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Phinney, D.G.; Prockop, D.J. Concise review: Mesenchymal stem/multipotent stromal cells: The state of transdifferentiation and modes of tissue repair—Current views. Stem Cells 2007, 25, 2896–2902. [Google Scholar] [CrossRef] [PubMed]
- Bruckner, S.; Tautenhahn, H.M.; Winkler, S.; Stock, P.; Dollinger, M.; Christ, B. A fat option for the pig: Hepatocytic differentiated mesenchymal stem cells for translational research. Exp. Cell Res. 2014, 321, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Liu, G.; Wu, X.; Wang, Y.; Tong, M.; Liu, B.; Wang, K.; Peng, Y.; Kong, X. Adipose derived mesenchymal stem cells efficiently rescue carbon tetrachloride-induced acute liver failure in mouse. Sci. World J. 2014, 2014, 103643. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Yang, J.; Yu, J.; Pan, Q.; Li, J.; Zhou, P.; Li, Y.; Pan, X.; Wang, Y.; Li, L. Therapeutic potential of transplanted placental mesenchymal stem cells in treating Chinese miniature pigs with acute liver failure. BMC Med. 2012, 10. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; La Rosa, F.G.; Genova, E.E.; Huber, K.; Schaack, J.; Degregori, J.; Serkova, N.J.; Li, Y.; Su, L.J.; Kessler, E.; et al. Simultaneous activation of Kras and inactivation of p53 induces soft tissue sarcoma and bladder urothelial hyperplasia. PLoS ONE 2013, 8, e74809. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.; Zhou, N.; Li, J.; Shi, D.; Cao, H.; Li, J.; Li, L. Porcine Adipose-Derived Mesenchymal Stem Cells Retain Their Stem Cell Characteristics and Cell Activities While Enhancing the Expression of Liver-Specific Genes after Acute Liver Failure. Int. J. Mol. Sci. 2016, 17, 62. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010062
Hu C, Zhou N, Li J, Shi D, Cao H, Li J, Li L. Porcine Adipose-Derived Mesenchymal Stem Cells Retain Their Stem Cell Characteristics and Cell Activities While Enhancing the Expression of Liver-Specific Genes after Acute Liver Failure. International Journal of Molecular Sciences. 2016; 17(1):62. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010062
Chicago/Turabian StyleHu, Chenxia, Ning Zhou, Jianzhou Li, Ding Shi, Hongcui Cao, Jun Li, and Lanjuan Li. 2016. "Porcine Adipose-Derived Mesenchymal Stem Cells Retain Their Stem Cell Characteristics and Cell Activities While Enhancing the Expression of Liver-Specific Genes after Acute Liver Failure" International Journal of Molecular Sciences 17, no. 1: 62. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17010062