
CTHRSSVVC Peptide as a Possible Early Molecular Imaging Target for Atherosclerosis

Abstract
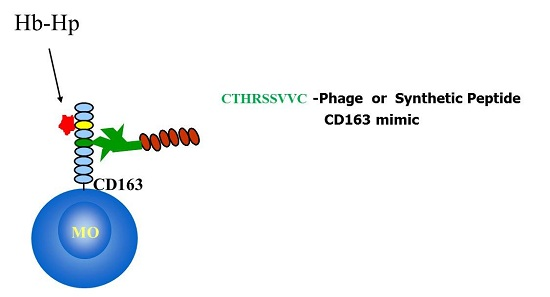
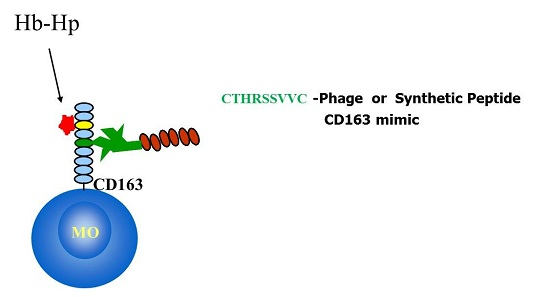
:
1. Introduction
2. Results
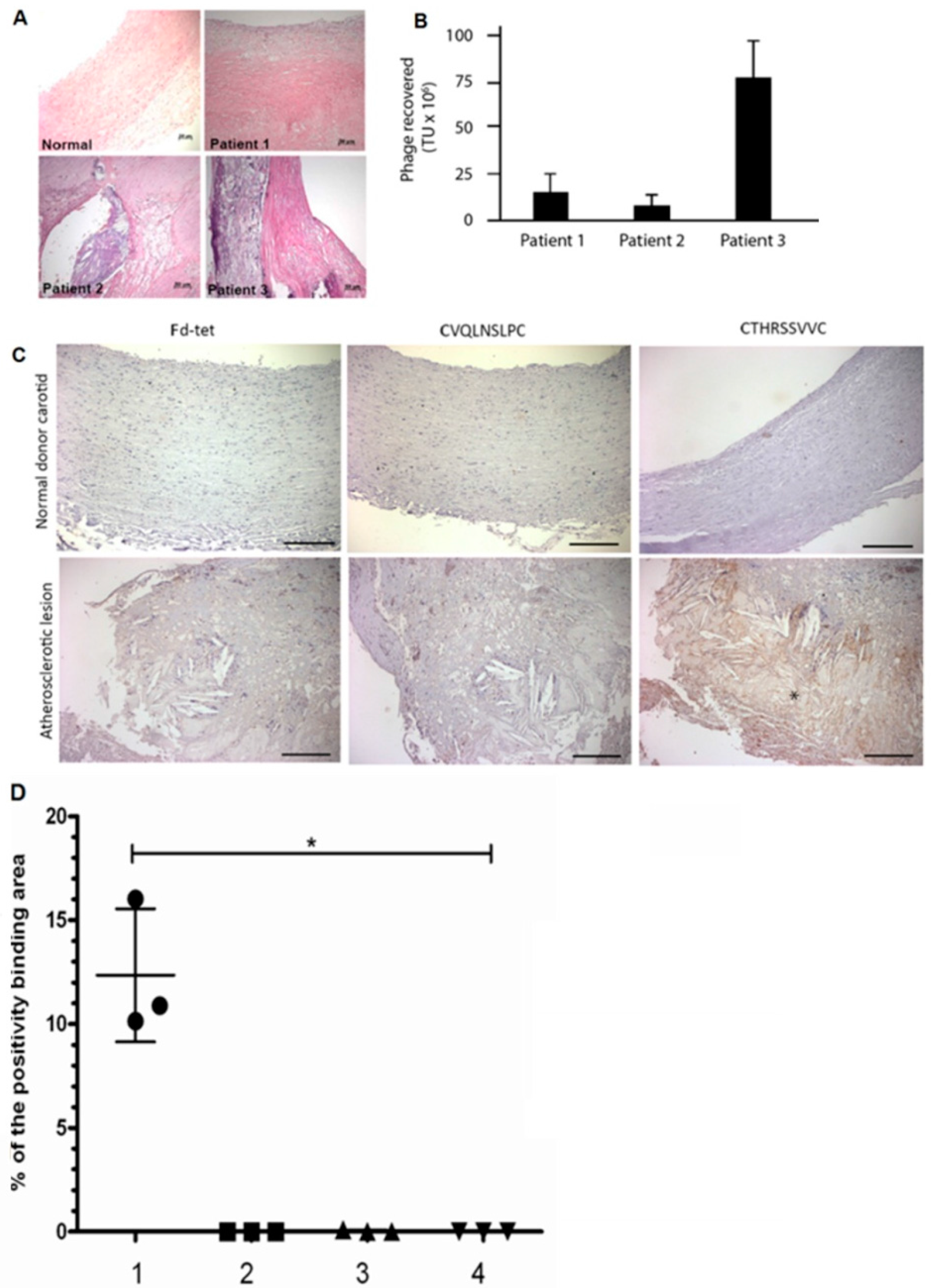
2.1. Selection of Phages Displaying Peptides Bound to Atherosclerotic Lesions
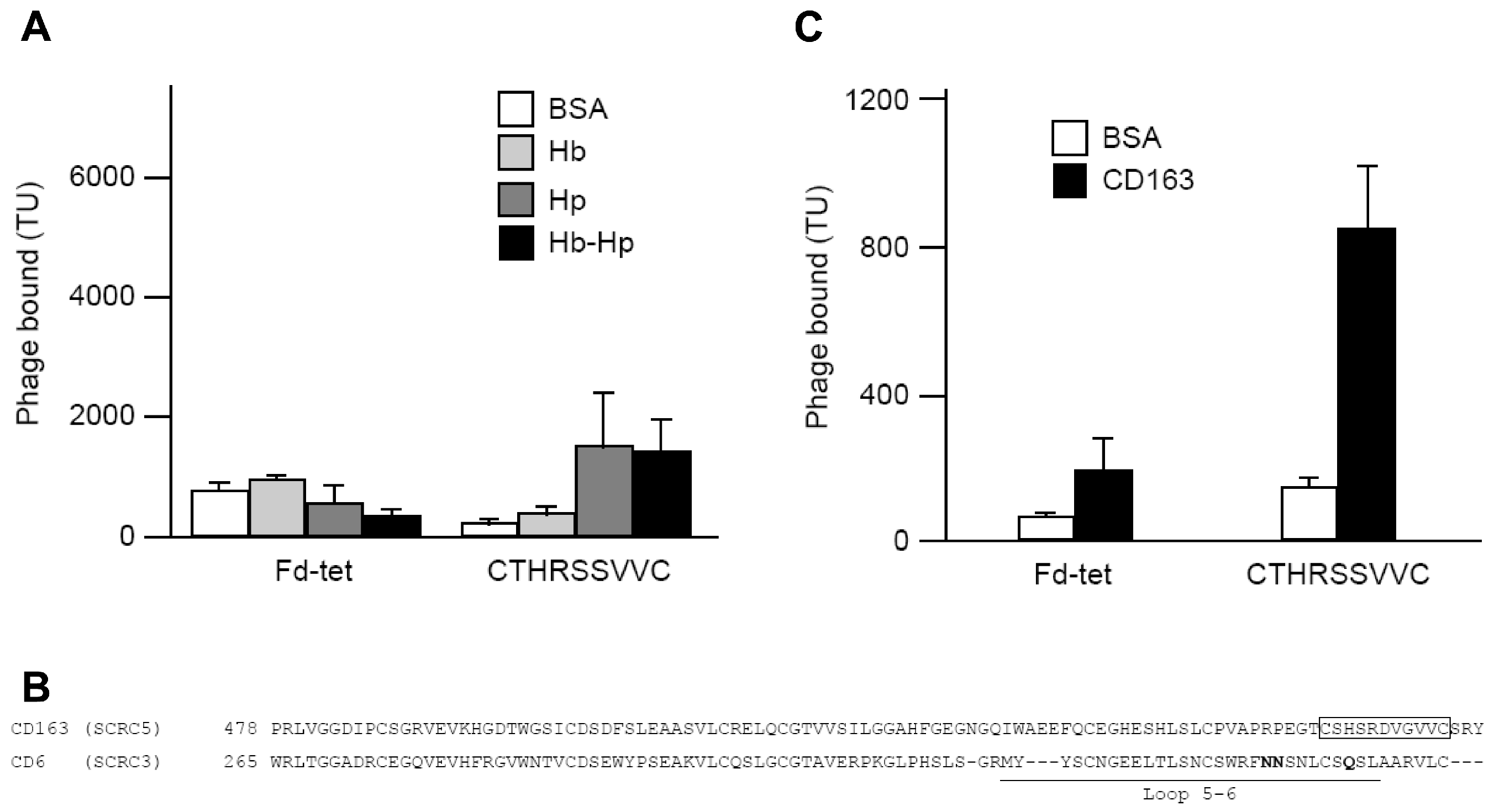
2.2. Peptide Selection and Receptor Identification
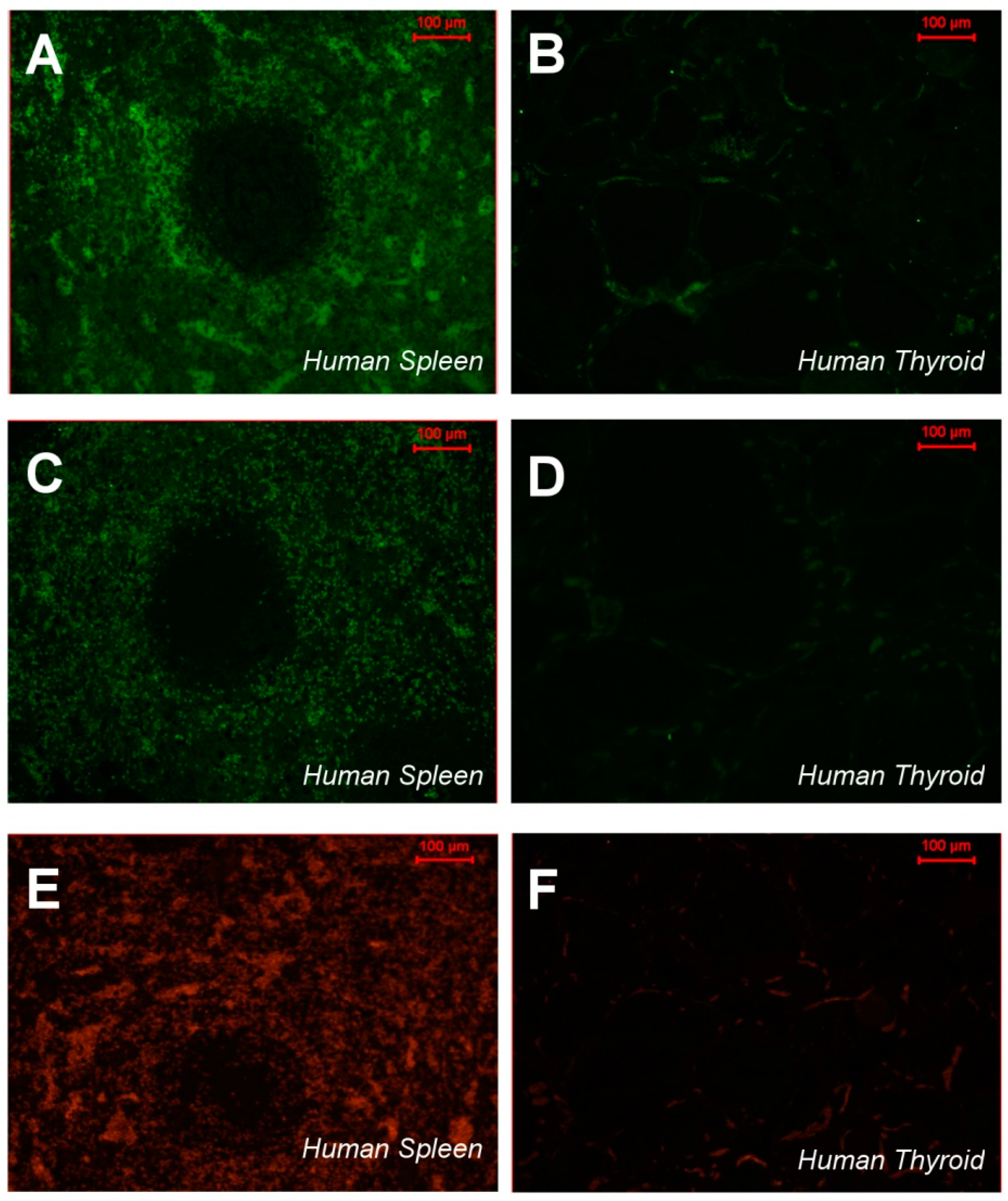
2.3. CTHRSSVVC-Phage Targets for Tissues
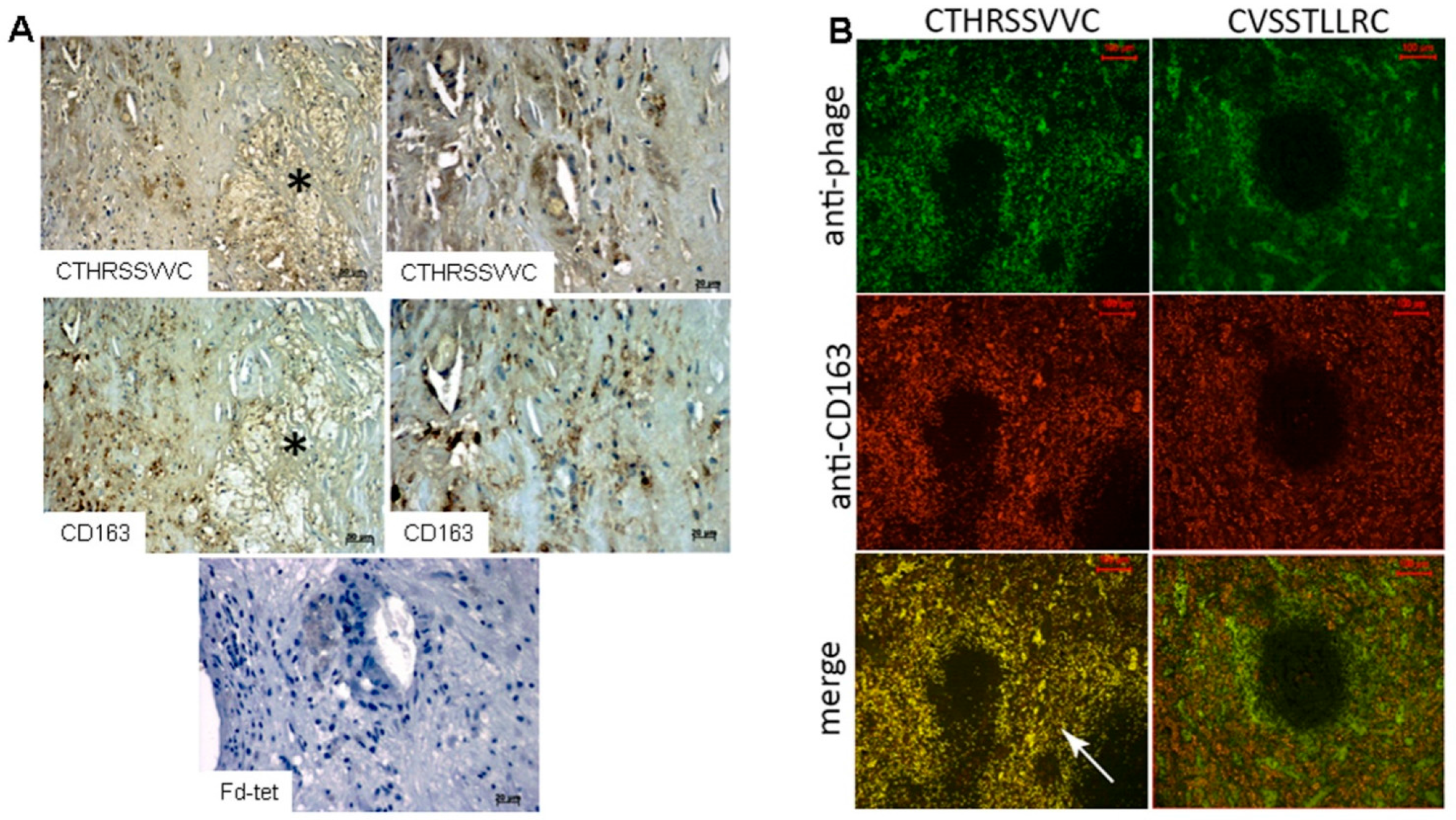
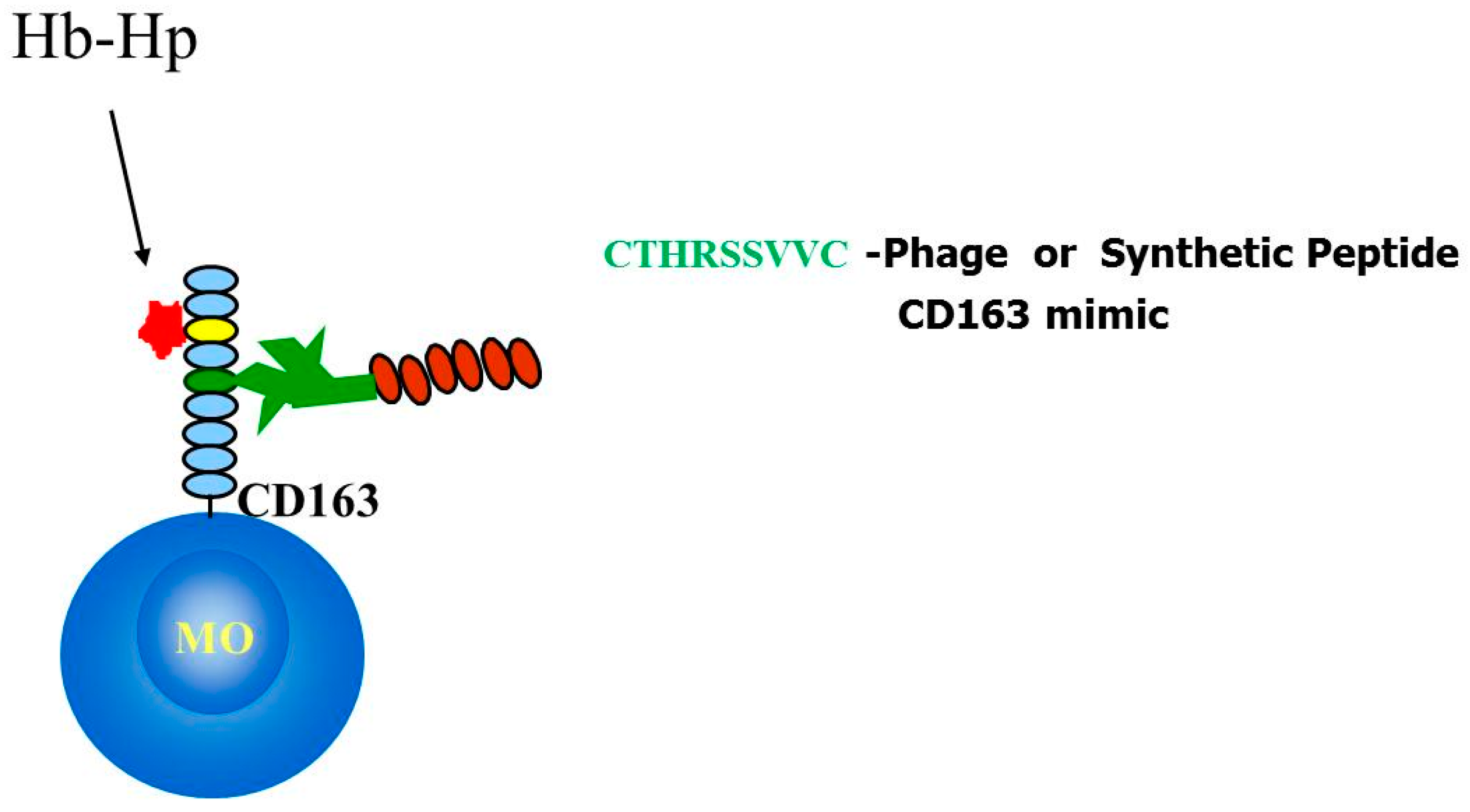
2.4. Identification and Characterization of a CD163 Binding Peptide
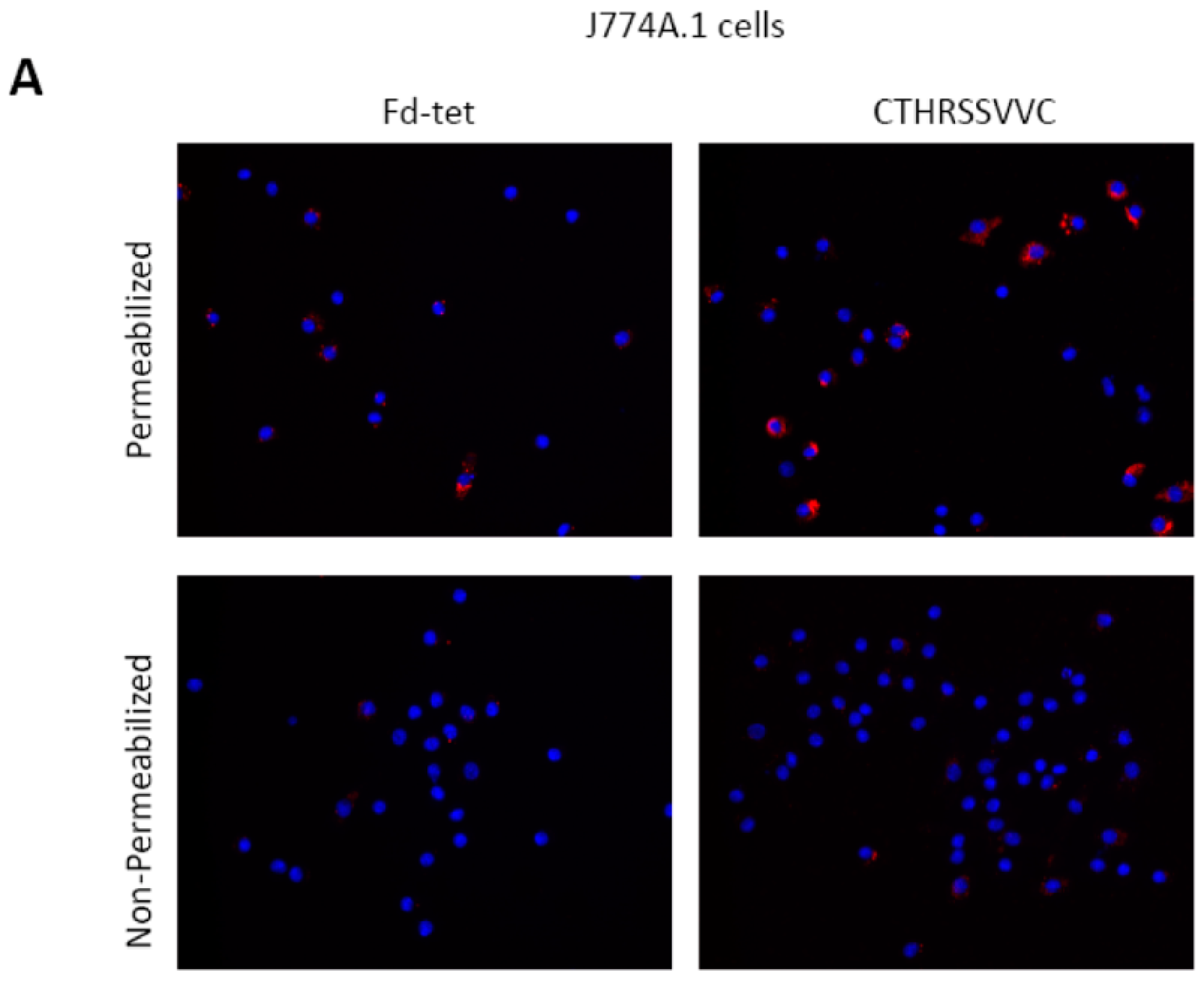
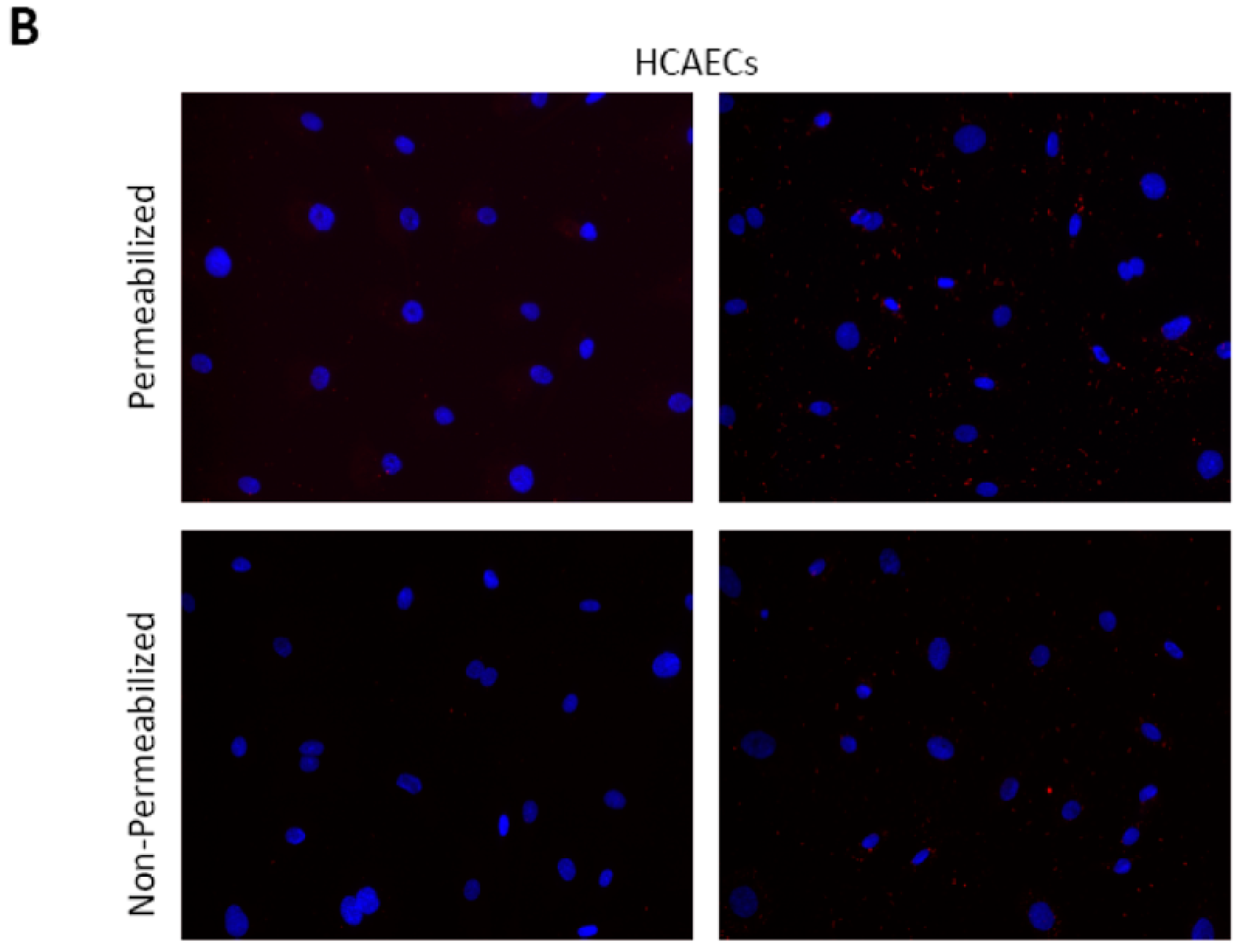
2.5. CTHRSSVVC-Phage Is Internalized by CD163 Expressing Cells
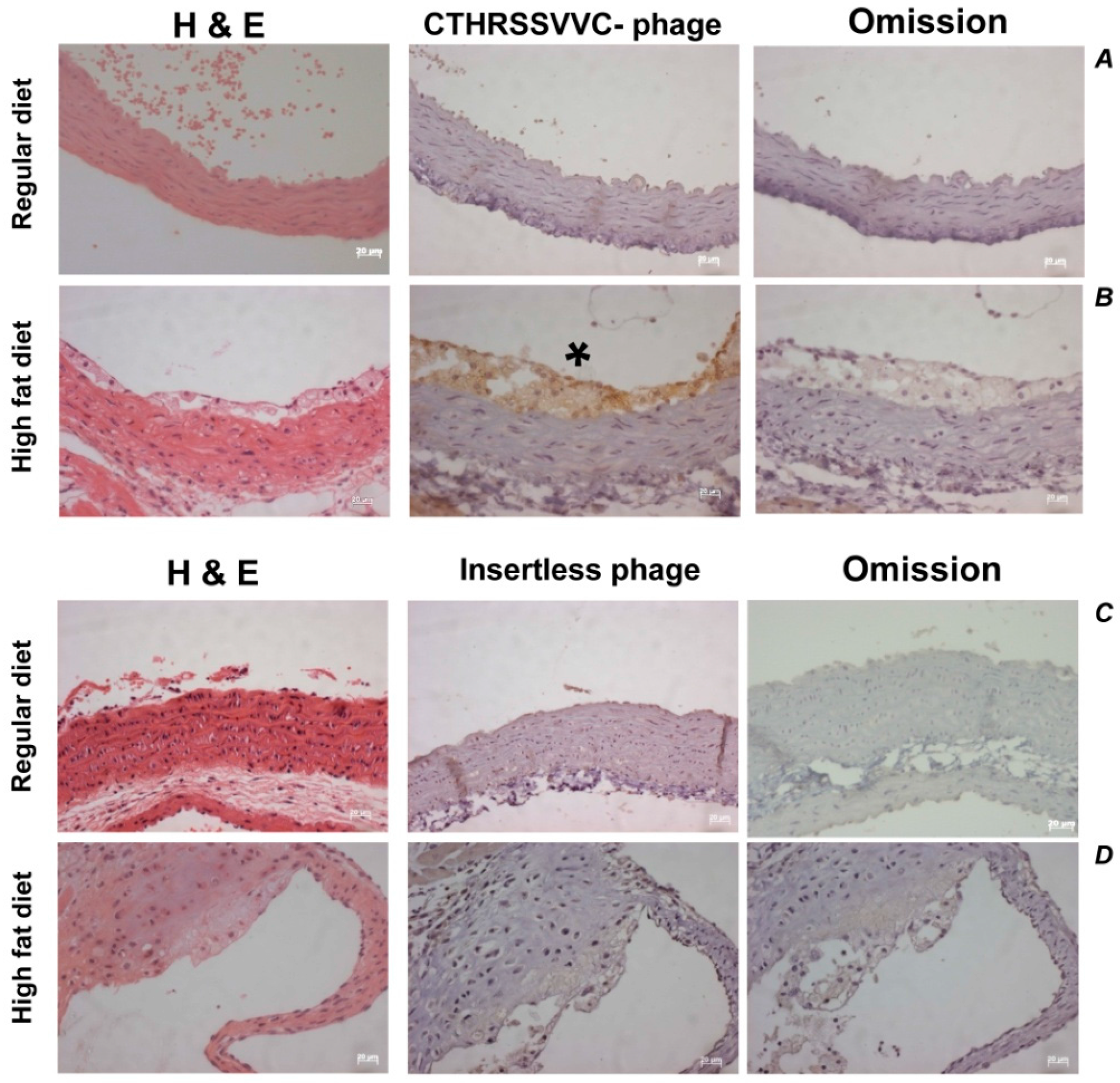
2.6. CTHRSSVVC-Phage Targets Atherosclerotic Lesions in Vivo
2.7. 111In-DOTA-CTHRSSVVC Peptide Preparation and Atheroma/Normal Carotid Binding Studies
3. Discussion
4. Materials and Methods
4.1. General Procedures
4.1.1. Ethics Statement
4.1.2. Reagents
4.1.3. Atheroma Plaques and Normal Carotid Specimens
4.1.4. Phage Display Selection
4.1.5. Phage Amplification
4.1.6. Phage Overlay Assay
4.1.7. Quantification of Phage Positivity
4.1.8. Phage Binding Assay
4.1.9. Phage Internalization
4.1.10. 111In-DOTA-CTHRSSVVC Peptide Preparation and Atheroma/Normal Carotid Binding Studies
4.2. CTHRSSVVC-Phage Targeting Atherosclerotic Lesions in Vivo
4.2.1. Animal Model (LDLr−/−)
4.2.2. CTHRSSVVC-Phage Homing in LDLr−/− Mice
4.2.3. Immunohistochemistry
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vinchi, F.; Muckenthaler, M.U.; Da Silva, M.C.; Balla, G.; Balla, J.; Jeney, V. Atherogenesis and iron: From epidemiology to cellular level. Front. Pharmacol. 2014, 5, 94. [Google Scholar] [CrossRef] [PubMed]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Chinetti-Gbaguidi, G.; Colin, S.; Staels, B. Macrophage subsets in atherosclerosis. Nat. Rev. Cardiol. 2015, 12, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Boyle, J.J.; Harrington, H.A.; Piper, E.; Elderfield, K.; Stark, J.; Landis, R.C.; Haskard, D.O. Coronary intraplaque hemorrhage evokes a novel atheroprotective macrophage phenotype. Am. J. Pathol. 2009, 174, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Madsen, M.; Moller, H.J.; Nielsen, M.J.; Jacobsen, C.; Graversen, J.H.; van den, B.T.; Moestrup, S.K. Molecular characterization of the haptoglobin.hemoglobin receptor CD163. Ligand binding properties of the scavenger receptor cysteine-rich domain region. J. Biol. Chem. 2004, 279, 51561–51567. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, M.; Graversen, J.H.; Jacobsen, C.; Sonne, O.; Hoffman, H.J.; Law, S.K.; Moestrup, S.K. Identification of the haemoglobin scavenger receptor. Nature 2001, 409, 198–201. [Google Scholar] [CrossRef] [PubMed]
- Staquicini, F.I.; Moeller, B.J.; Arap, W.; Pasqualini, R. Combinatorial vascular targeting in translational medicine. Proteom. Clin. Appl. 2010, 4, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Arap, W.; Kolonin, M.G.; Trepel, M.; Lahdenranta, J.; Cardo-Vila, M.; Giordano, R.J.; Mintz, P.J.; Ardelt, P.U.; Yao, V.J.; Vidal, C.I.; et al. Steps toward mapping the human vasculature by phage display. Nat. Med. 2002, 8, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Giordano, R.J.; Cardo-Vila, M.; Salameh, A.; Anobom, C.D.; Zeitlin, B.D.; Hawke, D.H.; Valente, A.P.; Almeida, F.C.; Nor, J.E.; Sidman, R.L.; et al. From combinatorial peptide selection to drug prototype (I): Targeting the vascular endothelial growth factor receptor pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 5112–5117. [Google Scholar] [CrossRef] [PubMed]
- Perschinka, H.; Wellenzohn, B.; Parson, W.; van der, Z.R.; Willeit, J.; Kiechl, S.; Wick, G. Identification of atherosclerosis-associated conformational heat shock protein 60 epitopes by phage display and structural alignment. Atherosclerosis 2007, 194, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.Y.; Lee, H.Y.; Kwak, W.; Yoo, J.; Na, M.H.; So, I.S.; Kwon, T.H.; Park, H.S.; Huh, S.; Oh, G.T.; et al. Phage display selection of peptides that home to atherosclerotic plaques: IL-4 receptor as a candidate target in atherosclerosis. J. Cell. Mol. Med. 2008, 12, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.G.; Zurita, A.J.; as-Neto, E.; Nunes, D.N.; Sidman, R.L.; Gelovani, J.G.; Arap, W.; Pasqualini, R. Beyond receptor expression levels: The relevance of target accessibility in ligand-directed pharmacodelivery systems. Trends Cardiovasc. Med. 2008, 18, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Giordano, R.J.; Edwards, J.K.; Tuder, R.M.; Arap, W.; Pasqualini, R. Combinatorial ligand-directed lung targeting. Proc. Am. Thorac. Soc. 2009, 6, 411–415. [Google Scholar] [CrossRef] [PubMed]
- Giordano, R.J.; Cardo-Vila, M.; Lahdenranta, J.; Pasqualini, R.; Arap, W. Biopanning and rapid analysis of selective interactive ligands. Nat. Med. 2001, 7, 1249–1253. [Google Scholar] [CrossRef] [PubMed]
- Giordano, R.J.; Lahdenranta, J.; Zhen, L.; Chukwueke, U.; Petrache, I.; Langley, R.R.; Fidler, I.J.; Pasqualini, R.; Tuder, R.M.; Arap, W. Targeted induction of lung endothelial cell apoptosis causes emphysema-like changes in the mouse. J. Biol. Chem. 2008, 283, 29447–29460. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.A.; Dejouvencel, T.; Labreuche, J.; Smadja, D.M.; Dussiot, M.; Martin-Ventura, J.L.; Egido, J.; Gaussem, P.; Emmerich, J.; Michel, J.B.; et al. Peripheral artery disease is associated with a high CD163/TWEAK plasma ratio. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Sainz, J.; Sata, M. Is PlGF a plaque growth factor? Cardiovasc. Res. 2010, 86, 4–5. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, M.J.; Moller, H.J.; Moestrup, S.K. Hemoglobin and heme scavenger receptors. Antioxid. Redox Signal. 2010, 12, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Moestrup, S.K.; Moller, H.J. CD163: A regulated hemoglobin scavenger receptor with a role in the anti-inflammatory response. Ann. Med. 2004, 36, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Aristoteli, L.P.; Moller, H.J.; Bailey, B.; Moestrup, S.K.; Kritharides, L. The monocytic lineage specific soluble CD163 is a plasma marker of coronary atherosclerosis. Atherosclerosis 2006, 184, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, M.M.; Tensen, C.P.; van As, J.H.; van den Berg, T.K.; Fluitsma, D.M.; Dijkstra, C.D.; Dopp, E.A.; Droste, A.; van Gaalen, F.A.; Sorg, C.; et al. Regulation of CD 163 on human macrophages: Cross-linking of CD163 induces signaling and activation. J. Leukoc. Biol. 1999, 66, 858–866. [Google Scholar] [PubMed]
- Graversen, J.H.; Madsen, M.; Moestrup, S.K. CD163: A signal receptor scavenging haptoglobin-hemoglobin complexes from plasma. Int. J. Biochem. Cell Biol. 2002, 34, 309–314. [Google Scholar] [CrossRef]
- Bowen, M.A.; Aruffo, A.A.; Bajorath, J. Cell surface receptors and their ligands: In vitro analysis of CD6-CD166 interactions. Proteins 2000, 40, 420–428. [Google Scholar] [CrossRef]
- Zurita, A.J.; Troncoso, P.; Cardo-Vila, M.; Logothetis, C.J.; Pasqualini, R.; Arap, W. Combinatorial screenings in patients: The interleukin-11 receptor alpha as a candidate target in the progression of human prostate cancer. Cancer Res. 2004, 64, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, M.L.; Moore, K.J.; Freeman, M.W.; Reed, G.L. Lipopolysaccharide induces scavenger receptor a expression in mouse macrophages: A divergent response relative to human THP-1 monocyte/macrophages. J. Immunol. 2000, 164, 2692–2700. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, R.; Ruoslahti, E. Organ targeting in vivo using phage display peptide libraries. Nature 1996, 380, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Arap, W.; Pasqualini, R.; Ruoslahti, E. Cancer treatment by targeted drug delivery to tumor vasculature in a mouse model. Science 1998, 279, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Segers, F.M.; den, A.B.; Bot, I.; van der Graaf, L.M.; van, D.V.; Gonzalez, W.; Raynal, I.; de Winther, M.; Wodzig, W.K.; Poelmann, R.E.; et al. Scavenger receptor-AI-targeted iron oxide nanoparticles for in vivo MRI detection of atherosclerotic lesions. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1812–1819. [Google Scholar] [CrossRef] [PubMed]
- De Vries, H.E.; Moor, A.C.; Dubbelman, T.M.; van Berkel, T.J.; Kuiper, J. Oxidized low-density lipoprotein as a delivery system for photosensitizers: Implications for photodynamic therapy of atherosclerosis. J. Pharmacol. Exp. Ther. 1999, 289, 528–534. [Google Scholar] [PubMed]
- Lipinski, M.J.; Amirbekian, V.; Frias, J.C.; Aguinaldo, J.G.; Mani, V.; Briley-Saebo, K.C.; Fuster, V.; Fallon, J.T.; Fisher, E.A.; Fayad, Z.A. MRI to detect atherosclerosis with gadolinium-containing immunomicelles targeting the macrophage scavenger receptor. Magn. Reson. Med. 2006, 56, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Bhattacharjee, G.; Boisvert, W.; Dilley, R.; Edgington, T. In vivo interrogation of the molecular display of atherosclerotic lesion surfaces. Am. J. Pathol. 2003, 163, 1859–1871. [Google Scholar] [CrossRef]
- Houston, P.; Goodman, J.; Lewis, A.; Campbell, C.J.; Braddock, M. Homing markers for atherosclerosis: Applications for drug delivery, gene delivery and vascular imaging. FEBS Lett. 2001, 492, 73–77. [Google Scholar] [CrossRef]
- Molenaar, T.J.; Twisk, J.; de Haas, S.A.; Peterse, N.; Vogelaar, B.J.; van Leeuwen, S.H.; Michon, I.N.; van Berkel, T.J.; Kuiper, J.; Biessen, E.A. P-selectin as a candidate target in atherosclerosis. Biochem. Pharmacol. 2003, 66, 859–866. [Google Scholar] [CrossRef]
- Kelly, K.A.; Nahrendorf, M.; Yu, A.M.; Reynolds, F.; Weissleder, R. In vivo phage display selection yields atherosclerotic plaque targeted peptides for imaging. Mol. Imaging Biol. 2006, 8, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, S.F.; Sandholt, B.V.; Keller, S.H.; Hansen, A.E.; Clemmensen, A.E.; Sillesen, H.; Højgaard, L.; Ripa, R.S.; Kjær, A. 64Cu-DOTATATE PET/MRI for Detection of Activated Macrophages in Carotid Atherosclerotic Plaques: Studies in Patients Undergoing Endarterectomy. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1696–1703. [Google Scholar] [CrossRef] [PubMed]
- Owen, A.; Dufès, C.; Moscatelli, D.; Mayes, E.; Lovell, J.F.; Katti, K.V.; Sokolov, K.; Mazza, M.; Fontaine, O.; Rannard, S.; et al. The application of nanotechnology in medicine: Treatment and diagnostics. Nanomedicine 2014, 9, 1291–1294. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Data Descriptions | Histopathology | Controls | Selected Phages | ||||
|---|---|---|---|---|---|---|---|
| Peptides | NPA | Fd | CVSSTLLRC | CVQLNSLPC | CQAYKLGSC | CTHRSSVVC | |
| Similarity | PlGF | Galectin 4 | Collagen α-1 (IX) chain | CD163 | |||
| Tissues/Diagnosis | |||||||
| Patient 1 Hypertension, dyslipidemia and aneurysm | Inflammation, without thrombus, calcium, no hemorrhage. | − | − | + FC | NP | NP | ++ LC |
| Patient 2 Diabetes and revascularization | Inflammation, thrombus, calcium, hemorrhage. | − | − | + FC | + FC | + FC | ++ LC, T |
| Patient 3 Revascularization, hypertension, dyslipidemia, stroke and abdominal aortic aneurysm | Inflammation, thrombus, calcium, no hemorrhage. | − | − | + FC | + FC | + FC | ++ LC, T |
| Non Atherosclerotic Carotid 3 | Normal | − | − | − | − | NP | − |
| Peptide | Motif | Blast Score * | Protein | GenBank |
|---|---|---|---|---|
| CVQLNSLPC | QLNSLP | 21.8 | Galectin 4 | AAH34750.1 |
| CQAYKLGSC | AYKLGS | 20.3 | Collagen α-1 (IX) chain | NP_001094312.1 |
| CTHRSSVVC | C+B/A+HSR-VVC | 38.8 | CD163 | CAB45233.1 |
| CVSSTLLRC | CVS LLRC | 18.9 | Placental growth factor | AAB30462.2 |
| CQALSNALC | QALSNAL | 20.6 | Angiomotion | AAI30295.1 |
| CDLLYNGVC | DLLYN+C | 23.1 | Ephrin type-B receptor 3 | NP004434.2 |
| DLL+NVC | 21.8 | Ephrin type-B receptor 6 | ABV55388.1 | |
| CGTQSGASC | T+SGASC | 21.0 | Tissue plasminogen activator | CAX11668.1 |
| CRREGVERC | R+GVERC | 24.0 | Emilin-1 | AAH09947.2 |
| CDGRFVRVC | RFVRV | 19.7 | Apolipoprotein L4 | CAQ08533.1 |
| CFVAGRVRC | FVA RVR | 21.4 | Inhibin β C | NP_005529.1 |
| CLEDSSWAC | CLED WA | 22.7 | C4b-binding protein β | NP_001017364.1 |
| CTSVVSSRC | TSVVSSR | 23.1 | Integrin α-11 | AAD51919 |
| Samples | * Atheroma CTHRSSVVC-Phage | Atheroma Insertless-Phage | Normal Carotid CTHRSSVVC-Phage | Normal Carotid Insertless-Phage |
|---|---|---|---|---|
| 1 | 10.14 | 0.01 | 0.01 | 0.00 |
| 2 | 10.88 | 0.00 | 0.09 | 0.00 |
| 3 | 16.02 | 0.00 | 0.01 | 0.00 |
| ±SEM | 12.35 | 0.00 | 0.04 | 0.00 |
| ±SD | 3.20 | 0.01 | 0.05 | 0.00 |
| Phages/Control | LDLr−/− High-Fat Diet | LDLr−/− Regular-Diet |
|---|---|---|
| CTHRSSVVC-phage * | 100% (n = 6) | 0% (n = 4) |
| Insertless phage (Fd) | 0% (n = 6) | 0% (n = 3) |
| Vehicle Total | 0% (n = 1) (n = 13) | 0% (n = 1) (n = 8) |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, R.A.; Giordano, R.J.; Gutierrez, P.S.; Rocha, V.Z.; Rudnicki, M.; Kee, P.; Abdalla, D.S.P.; Puech-Leão, P.; Caramelli, B.; Arap, W.; et al. CTHRSSVVC Peptide as a Possible Early Molecular Imaging Target for Atherosclerosis. Int. J. Mol. Sci. 2016, 17, 1383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091383
Silva RA, Giordano RJ, Gutierrez PS, Rocha VZ, Rudnicki M, Kee P, Abdalla DSP, Puech-Leão P, Caramelli B, Arap W, et al. CTHRSSVVC Peptide as a Possible Early Molecular Imaging Target for Atherosclerosis. International Journal of Molecular Sciences. 2016; 17(9):1383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091383
Chicago/Turabian StyleSilva, Rosemeire A., Ricardo J. Giordano, Paulo S. Gutierrez, Viviane Z. Rocha, Martina Rudnicki, Patrick Kee, Dulcinéia S. P. Abdalla, Pedro Puech-Leão, Bruno Caramelli, Wadih Arap, and et al. 2016. "CTHRSSVVC Peptide as a Possible Early Molecular Imaging Target for Atherosclerosis" International Journal of Molecular Sciences 17, no. 9: 1383. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms17091383