
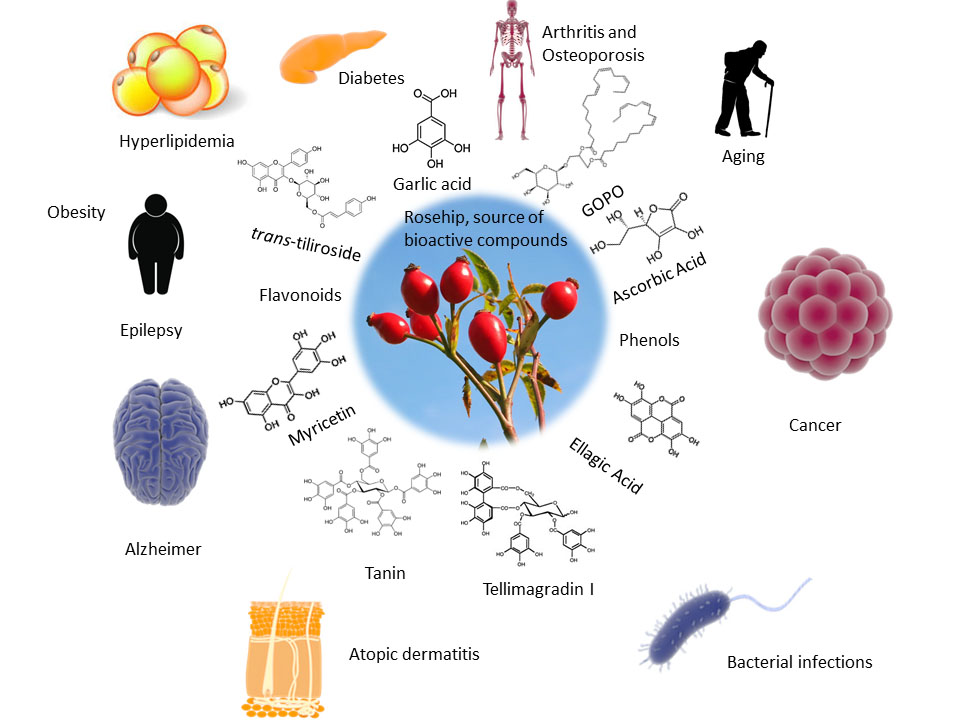
Therapeutic Applications of Rose Hips from Different Rosa Species

Abstract
:
1. Methodology
2. Introduction
2.1. Origin of Oxidation: The Importance of Natural Antioxidants
2.2. Presence of Antioxidants in Rose Hips Coming from Different Varieties of Rosa
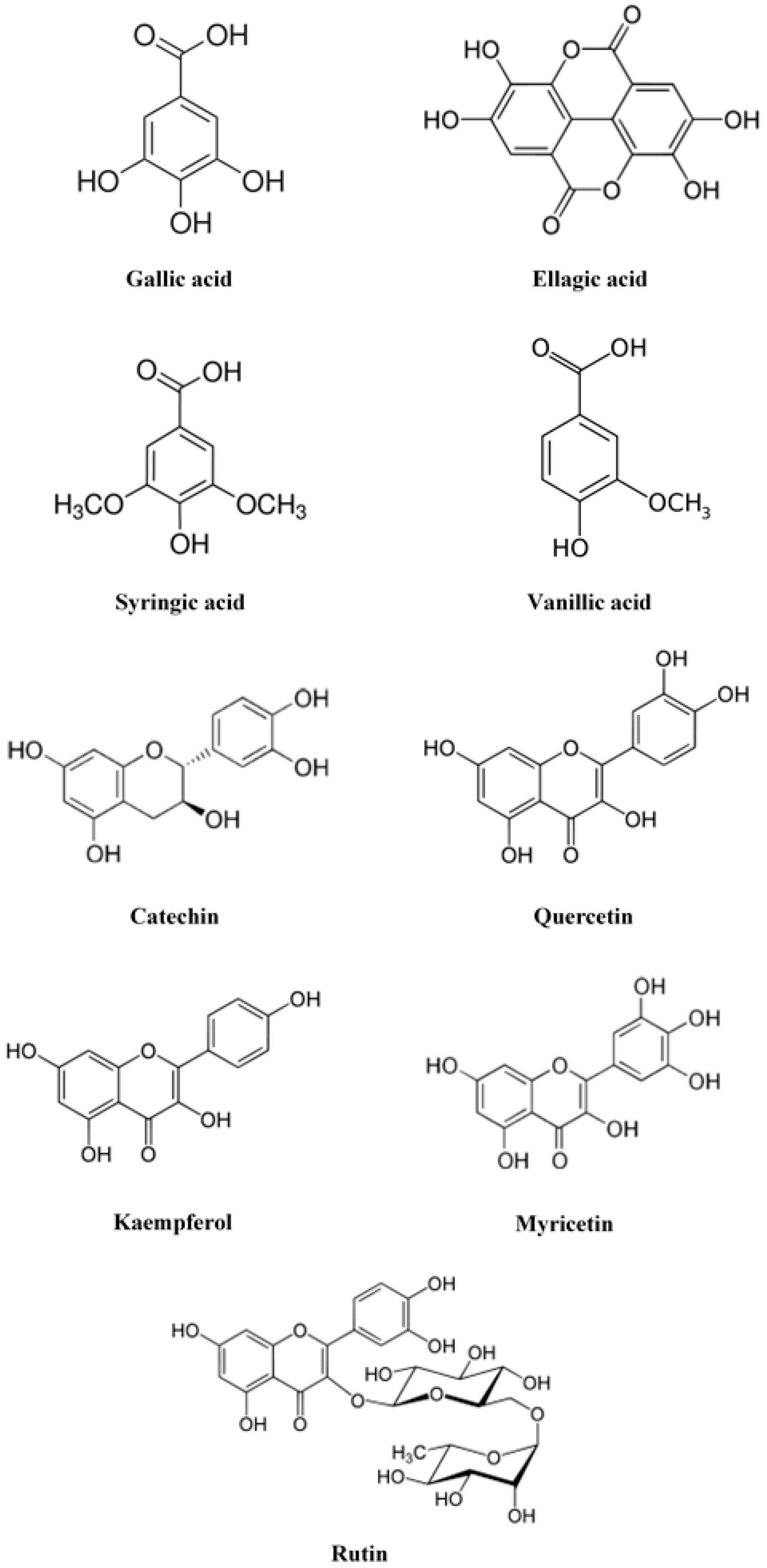
2.2.1. Hydrosoluble Antioxidants
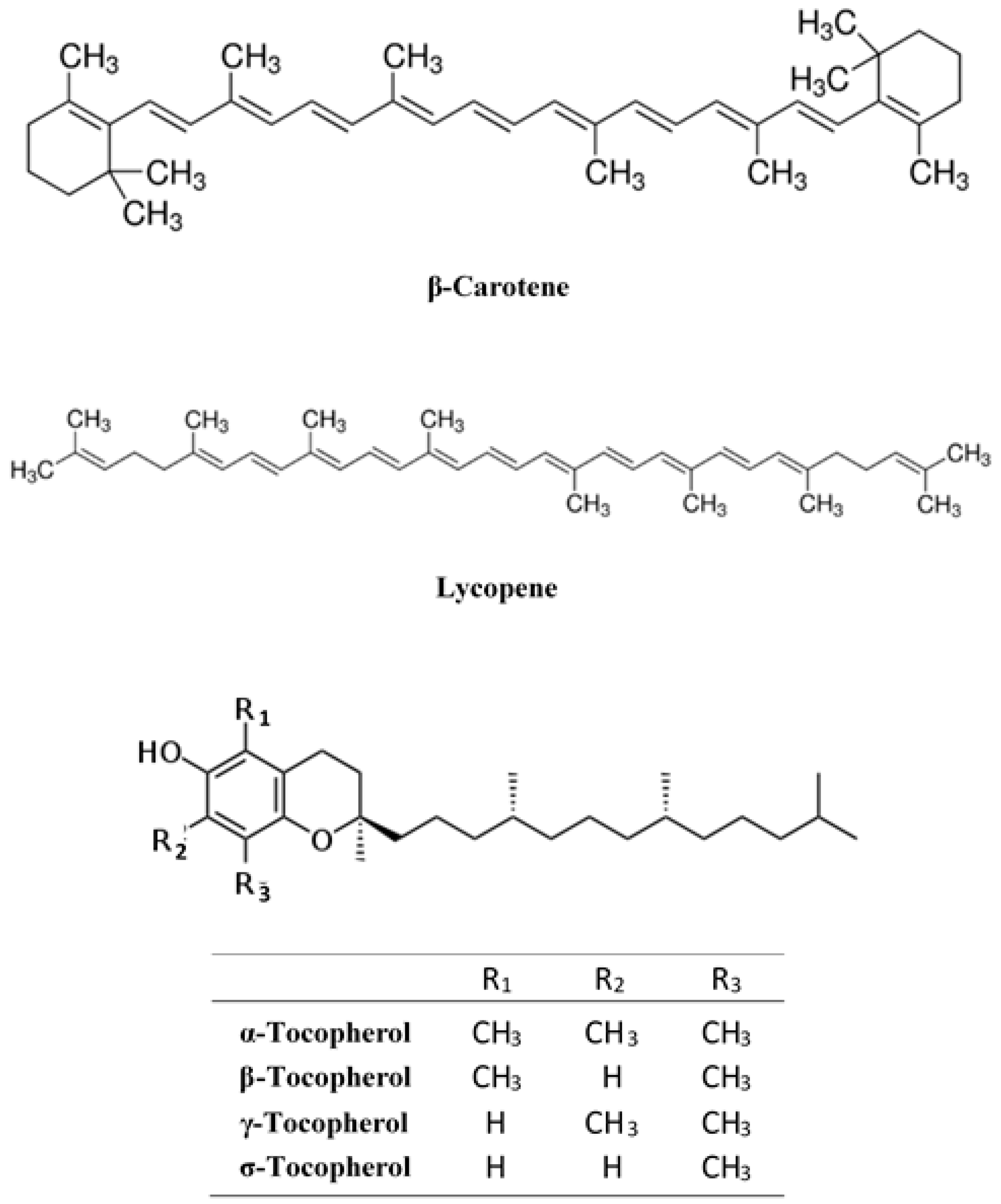
2.2.2. Lipid-Soluble Antioxidants
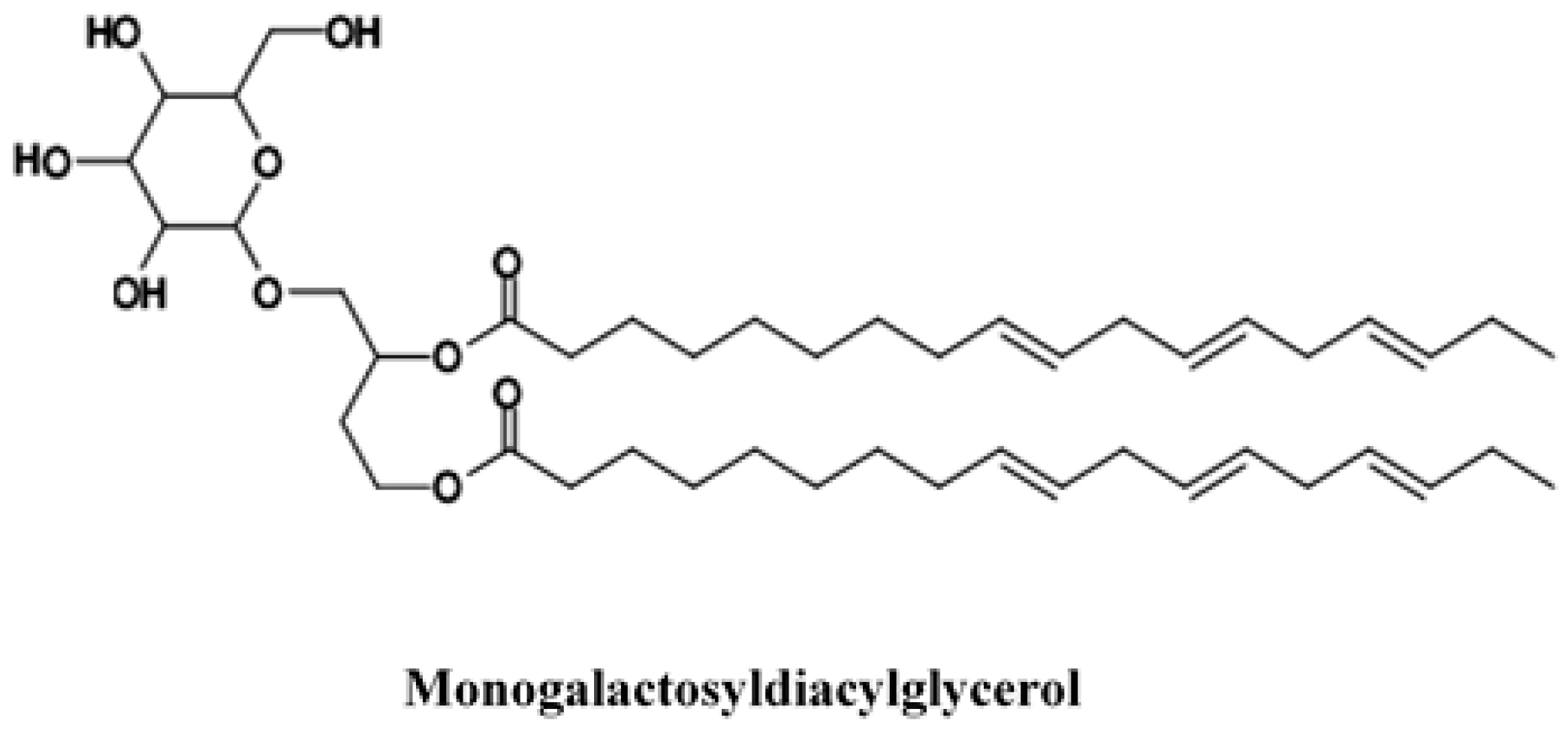
2.3. Presence of Other Active Compounds in Rose Hips
2.4. Factors Affecting the Concentration of Bioactive Compounds in Rose Hips. Utilization of These Compounds in Agro-Food Industry
3. Rose Hip and Disease
3.1. Anticancer Activity
3.2. Rheumatoid Arthritis
3.3. Osteoporosis
3.4. Diabetes
3.5. Hyperlipidaemia
3.6. Obesity
3.7. Renal Disturbances
3.8. Hepatotoxicity
3.9. Neuroprotective Activity
3.10. Skin Disorders and Aging
3.11. Diarrhoeal
3.12. Anti-Ulcerogenic Effect
3.13. Antimicrobial Effect
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gutteridge, J.M.; Halliwell, B. Free radicals and antioxidants in the year 2000. A historical look to the future. Ann. N. Y. Acad. Sci. 2000, 899, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Cheng, X.; Wang, L.; Wang, S.; Ren, G. A determination of potential alpha-glucosidase inhibitors from azuki beans (Vigna angularis). Int. J. Mol. Sci. 2011, 12, 6445–6451. [Google Scholar] [CrossRef] [PubMed]
- Willcox, J.K.; Ash, S.L.; Catignani, G.L. Antioxidants and prevention of chronic disease. Crit. Rev. Food Sci. Nutr. 2004, 44, 275–295. [Google Scholar] [CrossRef] [PubMed]
- Punchard, N.A.; Kelly, F.J. Free Radicals: A Practical Approach; IRL Press at Oxford University Press: Oxford, UK, 1996. [Google Scholar]
- Zhou, K.; Laux, J.J.; Yu, L. Comparison of swiss red wheat grain and fractions for their antioxidant properties. J. Agric. Food Chem. 2004, 52, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Charles, D.J. Antioxidant Properties of Spices, Herbs and Other Sources; Springer: New York, NY, USA, 2013. [Google Scholar]
- Okuda, T. Systematics and health effects of chemically distinct tannins in medicinal plants. Phytochemistry 2005, 66, 2012–2031. [Google Scholar] [CrossRef] [PubMed]
- Eberhardt, M.V.; Lee, C.Y.; Liu, R.H. Antioxidant activity of fresh apples. Nature 2000, 405, 903–904. [Google Scholar] [PubMed]
- Ganesan, K.; Kumar, K.S.; Rao, P.S. Comparative assessment of antioxidant activity in three edible species of green seaweed, enteromorpha from okha, northwest coast of India. Innov. Food Sci. Emerg. Technol. 2011, 12, 73–78. [Google Scholar] [CrossRef]
- Kaur, C.; Kapoor, H.C. Antioxidants in fruits and vegetables—The millennium’s health. Int. J. Food Sci. Technol. 2001, 36, 703–725. [Google Scholar] [CrossRef]
- Roberts, A.; Debener, T.; Gudin, S. Encyclopedia of Rose Science; Academic Press: Cambridge, MA, USA, 2003. [Google Scholar]
- Uggla, M.; Gustavsson, K.E.; Olsson, M.E.; Nybom, H. Changes in colour and sugar content in rose hips (Rosa dumalis L. and Rosa rubiginosa L.) during ripening. J. Hortic. Sci. Biotechnol. 2005, 80, 204–208. [Google Scholar] [CrossRef]
- Ercisli, S.; Orhan, E.; Esitken, A. Fatty acid composition of Rosa species seeds in turkey. Chem. Nat. Compd. 2007, 43, 605–606. [Google Scholar] [CrossRef]
- Tumbas, V.T.; Canadanovic-Brunet, J.M.; Cetojevic-Simin, D.D.; Cetkovic, G.S.; Ethilas, S.M.; Gille, L. Effect of rosehip (Rosa canina L.) phytochemicals on stable free radicals and human cancer cells. J. Sci. Food Agric. 2012, 92, 1273–1281. [Google Scholar] [CrossRef] [PubMed]
- Demir, N.; Yildiz, O.; Alpaslan, M.; Hayaloglu, A. Evaluation of volatiles, phenolic compounds and antioxidant activities of rose hip (Rosa L.) fruits in turkey. LWT Food Sci.Technol. 2014, 57, 126–133. [Google Scholar] [CrossRef]
- Nilsson, O.R. Flora of Turkey and the East Aegean Islands; University Press: Edinburgh, UK, 1997. [Google Scholar]
- Wissemann, V.; Gallenmüller, F.; Ritz, C.; Steinbrecher, T.; Speck, T. Inheritance of growth form and mechanical characters in reciprocal polyploid hybrids of rosa section caninae—implications for the ecological niche differentiation and radiation process of hybrid offspring. Trees 2006, 20, 340–347. [Google Scholar] [CrossRef]
- Özkan, G.; Sagdiç, O.; Baydar, N.; Baydar, N. Note: Antioxidant and antibacterial activities of Rosa damascena flower extracts. Food Sci. Tech. Int. 2004, 10, 277–281. [Google Scholar] [CrossRef]
- De la Iglesia, R.; Milagro, F.I.; Campion, J.; Boque, N.; Martinez, J.A. Healthy properties of proanthocyanidins. Biofactors 2010, 36, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Lopes, G.; Daletos, G.; Proksch, P.; Andrade, P.B.; Valentao, P. Anti-inflammatory potential of monogalactosyl diacylglycerols and a monoacylglycerol from the edible brown seaweed fucus Spiralis linnaeus. Mar. Drugs 2014, 12, 1406–1418. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, S.; Gascon, S.; Luquin, A.; Laguna, M.; Ancin-Azpilicueta, C.; Rodriguez-Yoldi, M.J. Rosa canina extracts have antiproliferative and antioxidant effects on Caco-2 human colon cancer. PLoS ONE 2016, 11, e0159136. [Google Scholar] [CrossRef] [PubMed]
- Demir, F.; Özcan, M. Chemical and technological properties of rose (Rosa canina L.) fruits grown wild in Turkey. J. Food Eng. 2001, 47, 333–336. [Google Scholar] [CrossRef]
- Olsson, M.E.; Andersson, S.; Werlemark, G.; Uggla, M.; Gustavsson, K.E. Carotenoids and phenolic in rose hips. Acta Hort. 2005, 490, 249–253. [Google Scholar] [CrossRef]
- Adamczak, A.; Buchwald, W.; Zieliński, J.; Mielcarek, S. Flavonoid and organic acid content in rose hips (Rosa L., sect. Caninae dc. Em. Christ.). Acta Biol. Cracov. Ser. Bot. 2012, 54, 105–112. [Google Scholar]
- Roman, I.; Stanila, A.; Stanila, S. Bioactive compounds and antioxidant activity of Rosa canina L. Biotypes from spontaneous flora of Transylvania. Chem. Cent. J. 2013, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Harrison, F.E. A critical review of vitamin c for the prevention of age-related cognitive decline and Alzheimer’s disease. J. Alzheimers Dis. 2012, 29, 711–726. [Google Scholar] [PubMed]
- Czyzowska, A.; Klewicka, E.; Pogorzelski, E.; Nowak, A. Polyphenols, vitamin C and antioxidant activity in wines from Rosa canina L. and Rosa rugosa thunb. J. Food Comp. Anal. 2015, 39, 62–68. [Google Scholar] [CrossRef]
- Türkben, C.; Uylaser, V.; Incedayi, B.; Çelikkol, I. Effects of different maturity periods and processes on nutritional components of rose hip (Rosa canina L.). J. Food Agric. Environ. 2010, 8, 26–30. [Google Scholar]
- Festa, F.; Aglitti, T.; Duranti, G.; Ricordy, R.; Perticone, P.; Cozzi, R. Strong antioxidant activity of ellagic acid in mammalian cells in vitro revealed by the comet assay. Anticancer Res. 2001, 21, 3903–3908. [Google Scholar] [PubMed]
- Whitley, A.C.; Stoner, G.D.; Darby, M.V.; Walle, T. Intestinal epithelial cell accumulation of the cancer preventive polyphenol ellagic acid—extensive binding to protein and DNA. Biochem. Pharmacol. 2003, 66, 907–915. [Google Scholar] [CrossRef]
- Schieber, A.; Berardini, N.; Carle, R. Identification of flavonol and xanthone glycosides from mango (Mangifera indica L. Cv.“Tommy Atkins”) peels by high-performance liquid chromatography-electrospray ionization mass spectrometry. J. Agric. Food Chem. 2003, 51, 5006–5011. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.D.; Zhang, D.Y.; Gao, X.J.; Parry, J.; Liu, K.; Liu, B.L.; Wang, M. Quercetin and quercetin-3-O-glucuronide are equally effective in ameliorating endothelial insulin resistance through inhibition of reactive oxygen species-associated inflammation. Mol. Nutr. Food Res. 2013, 57, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Hosni, K.; Chrif, R.; Zahed, N.; Abid, I.; Medfei, W.; Sebei, H.; Brahim, N.B. Fatty acid and phenolic constituents of leaves, flowers and fruits of tunisian dog rose (Rosa canina L.). Riv. Ital. Sostanze Gr. 2010, 87, 117–123. [Google Scholar]
- Fujii, T.; Saito, M. Inhibitory effect of quercetin isolated from rose hip (Rosa canina L.) against melanogenesis by mouse melanoma cells. Biosci. Biotechnol. Biochem. 2009, 73, 1989–1993. [Google Scholar] [CrossRef] [PubMed]
- Conceição de Oliveira, M.; Sichieri, R.; Sanchez Moura, A. Weight loss associated with a daily intake of three apples or three pears among overweight women. Nutrition 2003, 19, 253–256. [Google Scholar] [CrossRef]
- Ouerghemmi, S.; Sebei, H.; Siracusa, L.; Ruberto, G.; Saija, A.; Cimino, F.; Cristani, M. Comparative study of phenolic composition and antioxidant activity of leaf extracts from three wild Rosa species grown in different tunisia regions: Rosa canina L., Rosa moschata Herrm. And Rosa sempervirens L. Ind. Crops Prod. 2016, 94, 167–177. [Google Scholar] [CrossRef]
- Aladedunye, F.; Kersting, H.J.; Matthäus, B. Phenolic extract from wild rose hip with seed: Composition, antioxidant activity, and performance in canola oil. Eur. J. Lipid Sci. Technol. 2014, 116, 1025–1034. [Google Scholar] [CrossRef]
- Abdel-Hameed, E.-S.S.; Bazaid, S.A.; Shohayeb, M.M. Total phenolics and antioxidant activity of defatted fresh taif rose, Saudi Arabia. Br. J. Pharmacol. 2012, 2, 129–140. [Google Scholar] [CrossRef]
- Öztürk, N.; Tuncel, M.; Tuncel, N. Determination of phenolic acids by a modified HPLC: Its application to various plant materials. J. Liq. Chromatogr. Relat. Technol. 2007, 30, 587–596. [Google Scholar] [CrossRef]
- Denev, P.; Kratchanova, M.; Ciz, M.; Lojek, A.; Vasicek, O.; Nedelcheva, P.; Blazheva, D.; Toshkova, R.; Gardeva, E.; Yossifova, L.; et al. Biological activities of selected polyphenol-rich fruits related to immunity and gastrointestinal health. Food Chem. 2014, 157, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.B.; Haffner, K.; Baugerod, H.; Andersen, L.F.; et al. A systematic screening of total antioxidants in dietary plants. J. Nutr. 2002, 132, 461–471. [Google Scholar] [PubMed]
- Guimarães, R.; Barros, L.; Carvalho, A.M.; Ferreira, I.C. Studies on chemical constituents and bioactivity of Rosa micrantha: An alternative antioxidants source for food, pharmaceutical, or cosmetic applications. J. Agric. Food Chem. 2010, 58, 6277–6284. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Carvalho, A.M.; Ferreira, I.C. Exotic fruits as a source of important phytochemicals: Improving the traditional use of Rosa canina fruits in portugal. Food Res. Int. 2011, 44, 2233–2236. [Google Scholar] [CrossRef]
- Hodisan, T.; Socaciu, C.; Ropan, I.; Neamtu, G. Carotenoid composition of Rosa canina fruits determined by thin-layer chromatography and high-performance liquid chromatography. J. Pharm. Biomed. Anal. 1997, 16, 521–528. [Google Scholar] [CrossRef]
- Hornero-Mendez, D.; Minguez-Mosquera, M.I. Carotenoid pigments in Rosa mosqueta hips, an alternative carotenoid source for foods. J. Agric. Food Chem. 2000, 48, 825–828. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Henriksson, E.; Ström, K.; Alenfall, J.; Göransson, O.; Holm, C. Rose hip exerts antidiabetic effects via a mechanism involving downregulation of the hepatic lipogenic program. Am. J. Physiol. Endocrinol. Metab. 2011, 300, E111–E121. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T. β-carotene, carotenoids, and disease prevention in humans. FASEB J. 1996, 10, 690–701. [Google Scholar] [PubMed]
- Woodson, K.; Stewart, C.; Barrett, M.; Bhat, N.K.; Virtamo, J.; Taylor, P.R.; Albanes, D. Effect of vitamin intervention on the relationship between GSTM1, smoking, and lung cancer risk among male smokers. Cancer Epidemiol. Biomark. Prev. 1999, 8, 965–970. [Google Scholar]
- Chan, J.M.; Stampfer, M.J.; Ma, J.; Rimm, E.B.; Willett, W.C.; Giovannucci, E.L. Supplemental vitamin E intake and prostate cancer risk in a large cohort of men in the united states. Cancer Epidemiol. Biomark. Prev. 1999, 8, 893–899. [Google Scholar]
- Stone, W.L.; Papas, A.M. Tocopherols and the etiology of colon cancer. J. Nat. Cancer Inst. 1997, 89, 1006–1014. [Google Scholar] [CrossRef] [PubMed]
- Fromm, M.; Bayha, S.; Kammerer, D.R.; Carle, R. Identification and quantitation of carotenoids and tocopherols in seed oils recovered from different Rosaceae species. J. Agric. Food Chem. 2012, 60, 10733–10742. [Google Scholar] [CrossRef] [PubMed]
- Nowak, R. Fatty acids composition in fruits of wild rose species. Acta Soc. Bot. Pol. 2005, 74, 3. [Google Scholar] [CrossRef]
- Grajzer, M.; Prescha, A.; Korzonek, K.; Wojakowska, A.; Dziadas, M.; Kulma, A.; Grajeta, H. Characteristics of rose hip (Rosa canina L.) cold-pressed oil and its oxidative stability studied by the differential scanning calorimetry method. Food Chem. 2015, 188, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Kazaz, S.; Baydar, H.; Erbas, S. Variations in chemical compositions of Rosa damascena Mill. And Rosa canina L. fruits. Czech. J. Food Sci. Vol. 2009, 27, 178–184. [Google Scholar]
- Franco, D.; Pinelo, M.; Sineiro, J.; Núñez, M.J. Processing of Rosa rubiginosa: Extraction of oil and antioxidant substances. Bioresource Technol. 2007, 98, 3506–3512. [Google Scholar] [CrossRef] [PubMed]
- Szentmihályi, K.; Vinkler, P.; Lakatos, B.; Illés, V.; Then, M. Rose hip (Rosa canina L.) oil obtained from waste hip seeds by different extraction methods. Bioresource Technol. 2002, 82, 195–201. [Google Scholar] [CrossRef]
- Gaude, N.; Tippmann, H.; Flemetakis, E.; Katinakis, P.; Udvardi, M.; Dormann, P. The galactolipid digalactosyldiacylglycerol accumulates in the peribacteroid membrane of nitrogen-fixing nodules of soybean and lotus. J. Biol. Chem. 2004, 279, 34624–34630. [Google Scholar] [CrossRef] [PubMed]
- Somerville, C.J.B.; Jaworski, J.G.; Ohlrogge, J.B. Lipids; American Society of Plant Physiologists: Rockville, MD, USA, 2000. [Google Scholar]
- Maeda, N.; Hada, T.; Yoshida, H.; Mizushina, Y. Inhibitory effect on replicative DNA polymerases, human cancer cell proliferation, and in vivo anti-tumor activity by glycolipids from spinach. Curr. Med. Chem. 2007, 14, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Kokai, Y.; Hada, T.; Yoshida, H.; Mizushina, Y. Oral administration of monogalactosyl diacylglycerol from spinach inhibits colon tumor growth in mice. Exp. Ther. Med. 2013, 5, 17–22. [Google Scholar] [PubMed]
- Larsen, E.; Kharazmi, A.; Christensen, L.P.; Christensen, S.B. An antiinflammatory galactolipid from rose hip (Rosa canina) that inhibits chemotaxis of human peripheral blood neutrophils in vitro. J. Nat. Prod. 2003, 66, 994–995. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M. Rosehip—An evidence based herbal medicine for inflammation and arthritis. Aust. Fam. Physician 2012, 41, 495–498. [Google Scholar] [PubMed]
- Christensen, L.P. Galactolipids as potential health promoting compounds in vegetable foods. Recent Pat. Food Nutr. Agric. 2009, 1, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.-C.; Chen, Y.-P.; Wu, J.-H.; Huang, C.-C.; Wang, S.-Y.; Yang, N.-S.; Shyur, L.-F. A galactolipid possesses novel cancer chemopreventive effects by suppressing inflammatory mediators and mouse B16 melanoma. Cancer Res. 2007, 67, 6907–6915. [Google Scholar] [CrossRef] [PubMed]
- McCutcheon, A.; Roberts, T.; Gibbons, E.; Ellis, S.; Babiuk, L.; Hancock, R.; Towers, G. Antiviral screening of british columbian medicinal plants. J. Ethnopharmacol. 1995, 49, 101–110. [Google Scholar] [CrossRef]
- Orhan, D.D.; Hartevioğlu, A.; Küpeli, E.; Yesilada, E. In vivo anti-inflammatory and antinociceptive activity of the crude extract and fractions from Rosa canina L. Fruits. J. Ethnopharmacol. 2007, 112, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Dogan, A.; Celik, F.; Gundogdu, M.; Kazankaya, A.; Gazioglu Sensoy, R. Changes in seed mineral matter contents of rose hip species grown naturally in Turkey. Acta Hortic. 2015. [Google Scholar] [CrossRef]
- Dogan, A.; Kazankaya, A. Fruit properties of Rose hip species grown in lake Van Basin (Eastern Anatolia Region). Asian J. Plant. Sci. 2006, 5, 120–122. [Google Scholar]
- Strålsjö, L.; Alklint, C.; Olsson, M.E.; Sjöholm, I. Total folate content and retention in rosehips (Rosa ssp.) after drying. J. Agric. Food Chem. 2003, 51, 4291–4295. [Google Scholar] [CrossRef] [PubMed]
- Baiano, A.; Del Nobile, M.A. Antioxidant compounds from vegetable matrices: Biosynthesis, occurrence, and extraction systems. Crit. Rev. Food. Sci. Nutr. 2016, 56, 2053–2068. [Google Scholar] [CrossRef] [PubMed]
- Cunja, V.; Mikulic-Petkovsek, M.; Zupan, A.; Stampar, F.; Schmitzer, V. Frost decreases content of sugars, ascorbic acid and some quercetin glycosides but stimulates selected carotenes in Rosa canina hips. J. Plant. Physiol. 2015, 178, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, S.; Jiménez-Moreno, N.; Luquin, A.; Laguna, M.; Rodríguez-Yoldi, M.J.; Ancín-Azpilicueta, C. Chemical composition of rosehips from different Rosa species: An alternative source of antioxidants for food industry. Food Add. Contam. Part A 2017. [Google Scholar] [CrossRef] [PubMed]
- Vossen, E.; Utrera, M.; de Smet, S.; Morcuende, D.; Estévez, M. Dog rose (Rosa canina L.) as a functional ingredient in porcine frankfurters without added sodium ascorbate and sodium nitrite. Meat Sci. 2012, 92, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Romero, N.; Robert, P.; Masson, L.; Ortiz, J.; González, K.; Tapia, K.; Dobaganes, C. Effect of α-tocopherol, α-tocotrienol and Rosa mosqueta shell extract on the performance of antioxidant-stripped canola oil (Brassica sp.) at high temperature. Food Chem. 2007, 104, 383–389. [Google Scholar] [CrossRef]
- Utrera, M.; Morcuende, D.; Ganhão, R.; Estévez, M. Role of phenolics extracting from Rosa canina L. on meat protein oxidation during frozen storage and beef patties processing. Food Bioprocess. Tech. 2015, 8, 854–864. [Google Scholar] [CrossRef]
- Zizka, A.; Thiombiano, A.; Dressler, S.; Nacoulma, B.M.; Ouédraogo, A.; Ouédraogo, I.; Ouédraogo, O.; Zizka, G.; Hahn, K.; Schmidt, M. Traditional plant use in burkina faso (West Africa): A national-scale analysis with focus on traditional medicine. J. Ethnobiol. Ethnomed. 2015, 11, 9. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S.J. The role of traditional medicine practice in primary health care within aboriginal australia: A review of the literature. J. Ethnobiol. Ethnomed. 2013, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Nejatbakhsh, F.; Shirbeigi, L.; Rahimi, R.; Abolhassani, H. Review of local herbal compounds found in the iranian traditional medicine known to optimise male fertility. Andrologia 2016, 48, 850–859. [Google Scholar] [CrossRef] [PubMed]
- Deb, L.; Laishram, S.; Khumukcham, N.; Ningthoukhongjam, D.; Nameirakpam, S.S.; Dey, A.; Moirangthem, D.S.; Talukdar, N.C.; Ningthoukhongjam, T.R. Past, present and perspectives of manipur traditional medicine: A major health care system available for rural population in the North-East India. J. Ethnopharmacol. 2015, 169, 387–400. [Google Scholar] [CrossRef] [PubMed]
- De Fátima, Â.; Silva Terra, B.; Moreira da Silva, C.; Leite da Silva, D.; Pereira Araujo, D.; da Silva Neto, L.; Anderson Nascimento de Aquino, R. From nature to market: Examples of natural products that became drugs. Recent Pat. Biotechnol. 2014, 8, 76–88. [Google Scholar] [CrossRef] [PubMed]
- Thanan, R.; Oikawa, S.; Hiraku, Y.; Ohnishi, S.; Ma, N.; Pinlaor, S.; Yongvanit, P.; Kawanishi, S.; Murata, M. Oxidative stress and its significant roles in neurodegenerative diseases and cancer. Int. J. Mol. Sci. 2014, 16, 193–217. [Google Scholar] [CrossRef] [PubMed]
- Barrera, G. Oxidative stress and lipid peroxidation products in cancer progression and therapy. ISRN Oncol. 2012, 2012, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Chaudhary, A.; Jha, U.; Yadav, U.C. Oxidative stress and antioxidants in disease and cancer. Asian Pac. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef]
- Roleira, F.M.; Tavares-da-Silva, E.J.; Varela, C.L.; Costa, S.C.; Silva, T.; Garrido, J.; Borges, F. Plant derived and dietary phenolic antioxidants: Anticancer properties. Food Chem. 2015, 183, 235–258. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, R.; Barros, L.; Calhelha, R.C.; Carvalho, A.M.; Queiroz, M.J.R.; Ferreira, I.C. Bioactivity of different enriched phenolic extracts of wild fruits from northeastern portugal: A comparative study. Plant. Foods Hum. Nutr. 2014, 69, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Bonner, M.Y.; Arbiser, J.L. The antioxidant paradox: What are antioxidants and how should they be used in a therapeutic context for cancer. Future Med. Chem. 2014, 6, 1413–1422. [Google Scholar] [CrossRef] [PubMed]
- Uifălean, A.; Schneider, S.; Gierok, P.; Ionescu, C.; Iuga, C.A.; Lalk, M. The impact of soy isoflavones on MCF-7 and MDA-MB-231 breast cancer cells using a global metabolomic approach. Int. J. Mol. Sci. 2016, 17, 1443. [Google Scholar] [CrossRef] [PubMed]
- Zhamanbayeva, G.T.; Aralbayeva, A.N.; Murzakhmetova, M.K.; Tuleukhanov, S.T.; Danilenko, M. Cooperative antiproliferative and differentiation-enhancing activity of medicinal plant extracts in acute myeloid leukemia cells. Biomed. Pharmacother. 2016, 82, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Corcoran, A.; Nadkarni, S.; Yasuda, K.; Sakaki, T.; Brown, G.; Kutner, A.; Marcinkowska, E. Biological evaluation of double point modified analogues of 1,25-dihydroxyvitamin D2 as potential anti-leukemic agents. Int. J. Mol. Sci. 2016, 17, 91. [Google Scholar] [CrossRef] [PubMed]
- Baurska, H.; Kiełbiński, M.; Biecek, P.; Haus, O.; Jaźwiec, B.; Kutner, A.; Marcinkowska, E. Monocytic differentiation induced by side-chain modified analogs of vitamin D in ex vivo cells from patients with acute myeloid leukemia. Leuk. Res. 2014, 38, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Jung, M.G.; Kang, H.B.; Choi, K.-C.; Haam, S.; Jun, W.; Kim, Y.-J.; Cho, H.Y.; Yoon, H.-G. Effect of anti-histone acetyltransferase activity from Rosa rugosa thunb. (Rosaceae) extracts on androgen receptor-mediated transcriptional regulation. J. Ethnopharmacol. 2008, 118, 412–417. [Google Scholar] [CrossRef] [PubMed]
- Takayama, K.i.; Inoue, S. Transcriptional network of androgen receptor in prostate cancer progression. Int. J. Urol. 2013, 20, 756–768. [Google Scholar] [CrossRef] [PubMed]
- Li, L.-C.; Carroll, P.R.; Dahiya, R. Epigenetic changes in prostate cancer: Implication for diagnosis and treatment. J. Natl. Cancer Inst. 2005, 97, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Li, S.-Y.; Huang, X.-E.; Cui, J.-J.; Zhao, T.; Zhang, H. Inhibition of tumor growth in vitro by a combination of extracts from Rosa roxburghii Tratt and fagopyrum cymosum. Asian Pac. J. Cancer Prev. 2012, 13, 2409–2414. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Z.-J.; Liu, J.; Liu, L.-K.; Zhang, E.-S.; Li, W.-L. Inhibition of metastasis and invasion of ovarian cancer cells by crude polysaccharides from Rosa roxburghii Tratt in vitro. J. Cancer Prev. 2013, 15, 10351–10354. [Google Scholar] [CrossRef]
- Guo, F.; Tian, J.; Cui, M.; Fang, M.; Yang, L. Downregulation of Matrix Metalloproteinase 9 by small interfering rna inhibits the tumor growth of ovarian epithelial carcinoma in vitro and in vivo. Mol. Med. Rep. 2015, 12, 753–759. [Google Scholar] [PubMed]
- Lee, K.R.; Lee, J.S.; Kim, Y.R.; Song, I.G.; Hong, E.K. Polysaccharide from inonotus obliquus inhibits migration and invasion in B16-F10 cells by suppressing MMP-2 and MMP-9 via downregulation of NF-κB signaling pathway. Oncol. Rep. 2014, 31, 2447–2453. [Google Scholar] [CrossRef] [PubMed]
- Barbour, K.E. Prevalence of doctor-diagnosed arthritis at state and county levels—United States, 2014. MMWR Morb. Mortal. Wkly. Rep. 2016, 65, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.C. The infectious etiology of rheumatoid arthritis. Arthritis Rheum. 1978, 21, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Brooks, P.M. The burden of musculoskeletal disease: A global perspective. Clin. Rheumatol. 2006, 25, 778–781. [Google Scholar] [CrossRef] [PubMed]
- Kirkeskov, B.; Christensen, R.; Bügel, S.; Bliddal, H.; Danneskiold-Samsøe, B.; Christensen, L.P.; Andersen, J.R. The effects of rose hip (Rosa canina) on plasma antioxidative activity and c-reactive protein in patients with rheumatoid arthritis and normal controls: A prospective cohort study. Phytomedicine 2011, 18, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Kharazmi, A.; Winther, K. Rose hip inhibits chemotaxis and chemiluminescence of human peripheral blood neutrophils in vitro and reduces certain inflammatory parameters in vivo. Inflammopharmacology 1999, 7, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Warholm, O.; Skaar, S.; Hedman, E.; Mølmen, H.M.; Eik, L. The effects of a standardized herbal remedy made from a subtype of Rosa canina in patients with osteoarthritis: A double-blind, randomized, placebo-controlled clinical trial. Curr. Ther. Res. 2003, 64, 21–31. [Google Scholar] [CrossRef]
- Rein, E.; Kharazmi, A.; Winther, K. A herbal remedy, hyben vital (stand. Powder of a subspecies of Rosa canina fruits), reduces pain and improves general wellbeing in patients with osteoarthritis—a double-blind, placebo-controlled, randomised trial. Phytomedicine 2004, 11, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Winther, K.; Rein, E.; Kharazmi, A. The anti-inflammatory properties of rose-hip. Inflammopharmacology 1999, 7, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhu, Y.; Zhang, H.-J.; Jiao, W.-H.; Han, B.-N.; Liu, Z.-X.; Qiu, F.; Chen, W.-S.; Lin, H.-W. Anti-Inflammatory secondary metabolites from the leaves of Rosa laevigata. Bioorg. Med. Chem. 2013, 21, 3290–3297. [Google Scholar] [CrossRef] [PubMed]
- Schwager, J.; Hoeller, U.; Wolfram, S.; Richard, N. Rose hip and its constituent galactolipids confer cartilage protection by modulating cytokine, and chemokine expression. BMC Complement. Altern. Med. 2011, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Schwager, J.; Richard, N.; Schoop, R.; Wolfram, S. A novel rose hip preparation with enhanced anti-inflammatory and chondroprotective effects. Mediators Inflamm. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jäger, A.K.; Eldeen, I.M.; van Staden, J. COX-1 and-2 activity of rose hip. Phytother. Res. 2007, 21, 1251–1252. [Google Scholar] [CrossRef] [PubMed]
- Wenzig, E.; Widowitz, U.; Kunert, O.; Chrubasik, S.; Bucar, F.; Knauder, E.; Bauer, R. Phytochemical composition and in vitro pharmacological activity of two rose hip (Rosa canina L.) preparations. Phytomedicine 2008, 15, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-S.; Kim, S.-H. Inhibitory effect of astragalin on expression of lipopolysaccharide-induced inflammatory mediators through NF-κB in macrophages. Arch. Pharm. Res. 2011, 34, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Kim, M.J.; Heo, H.J.; Kim, H.K.; Hong, B.; Kim, C.-J.; Kim, B.-G.; Shin, D.H. Protective effect of Rosa laevigata against amyloid β peptide-induced oxidative stress. Amyloid 2006, 13, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Xu, Y.; Han, X.; Liang, C.; Yin, L.; Xu, L.; Qi, Y.; Zhao, Y.; Peng, J.; Sun, C. Potent effects of flavonoid-rich extract from Rosa laevigata michx fruit against hydrogen peroxide-induced damage in PC12 cells via attenuation of oxidative stress, inflammation and apoptosis. Molecules 2014, 19, 11816–11832. [Google Scholar] [CrossRef] [PubMed]
- Makarov, S.S. NF-κβ in rheumatoid arthritis: A pivotal regulator of inflammation, hyperplasia, and tissue destruction. Arthritis Res.Ther. 2001, 3, 200. [Google Scholar] [CrossRef] [PubMed]
- Whang, W.K.; Park, H.S.; Ham, I.; Oh, M.; Namkoong, H.; Kim, H.K.; Hwang, D.W.; Hur, S.Y.; Kim, T.E.; Park, Y.G. Methyl gallate and chemicals structurally related to methyl gallate protect human umbilical vein endothelial cells from oxidative stress. Exp. Mol. Med. 2005, 37, 343. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.C.; Lee, Y.H.; Jung, M.G.; Kwon, S.H.; Kim, M.J.; Jun, W.J.; Lee, J.; Lee, J.M.; Yoon, H.G. Gallic acid suppresses lipopolysaccharide-induced nuclear factor-kappab signaling by preventing rela acetylation in A549 lung cancer cells. Mol. Cancer Res. 2009, 7, 2011–2021. [Google Scholar] [CrossRef] [PubMed]
- An, H.-J.; Kim, I.-T.; Park, H.-J.; Kim, H.-M.; Choi, J.-H.; Lee, K.-T. Tormentic acid, a triterpenoid saponin, isolated from Rosa rugosa, inhibited LPS-induced INOS, COX-2, and TNF-α expression through inactivation of the nuclear factor-κB pathway in raw 264.7 macrophages. Int. Immunopharmacol. 2011, 11, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Takayanagi, H. Osteoclasts, rheumatoid arthritis, and osteoimmunology. Curr. Opin. Rheumatol. 2006, 18, 419–426. [Google Scholar] [CrossRef] [PubMed]
- Kurt, A.; Bjoern, F.; Bernd, W.; Eduardo, M. Rose hip special extract FB9440 inhibited osteoclastogenesis via NF-κB pathway. Planta Med. 2013, 79. [Google Scholar] [CrossRef]
- Cheng, B.C.Y.; Fu, X.-Q.; Guo, H.; Li, T.; Wu, Z.-Z.; Chan, K.; Yu, Z.-L. The genus Rosa and arthritis: Overview on pharmacological perspectives. Pharmacol. Res. 2016, 114, 219–234. [Google Scholar] [CrossRef] [PubMed]
- Bernabei, R.; Martone, A.M.; Ortolani, E.; Landi, F.; Marzetti, E. Screening, diagnosis and treatment of osteoporosis: A brief review. Clin. Cases Miner. Bone Metab. 2014, 11, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Banfi, G.; Iorio, E.L.; Corsi, M.M. Oxidative stress, free radicals and bone remodeling. Clin. Chem. Lab. Med. 2008, 46, 1550–1555. [Google Scholar] [CrossRef] [PubMed]
- Sheweita, S.; Khoshhal, K. Calcium metabolism and oxidative stress in bone fractures: Role of antioxidants. Curr. Drug Metab. 2007, 8, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Devareddy, L.; Hooshmand, S.; Collins, J.K.; Lucas, E.A.; Chai, S.C.; Arjmandi, B.H. Blueberry prevents bone loss in ovariectomized rat model of postmenopausal osteoporosis. J. Nutr. Biochem. 2008, 19, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemical profiles and antioxidant activity of wheat varieties. J. Agric. Food Chem. 2003, 51, 7825–7834. [Google Scholar] [CrossRef] [PubMed]
- Peterkofsky, B. Ascorbate requirement for hydroxylation and secretion of procollagen: Relationship to inhibition of collagen synthesis in scurvy. Am. J. Clin. Nutr. 1991, 54, 1135S–1140S. [Google Scholar] [PubMed]
- Xing, W.; Singgih, A.; Kapoor, A.; Alarcon, C.M.; Baylink, D.J.; Mohan, S. Nuclear Factor-E2-Related Factor-1 mediates ascorbic acid induction of osterix expression via interaction with antioxidant-responsive element in bone cells. J. Biol. Chem. 2007, 282, 22052–22061. [Google Scholar] [CrossRef] [PubMed]
- Hubert, P.A.; Lee, S.G.; Lee, S.-K.; Chun, O.K. Dietary polyphenols, berries, and age-related bone loss: A review based on human, animal, and cell studies. Antioxidants 2014, 3, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Blair, M. Diabetes mellitus review. Urol. Nurs. 2016, 36, 27–36. [Google Scholar] [PubMed]
- Gojka, R. WHO Global report on diabetes: A summary. Int. J. Noncommun. Dis. 2016, 1, 3–8. [Google Scholar]
- Ceriello, A. New insights on oxidative stress and diabetic complications may lead to a “causal” antioxidant therapy. Diabetes Care 2003, 26, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Westphal, S.A.; Palumbo, P.J. Insulin and oral hypoglycemic agents should not be used in combination in the treatment of type 2 diabetes. Arch. Intern. Med. 2003, 163, 1783–1785. [Google Scholar] [CrossRef] [PubMed]
- Orhan, N.; Aslan, M.; Hosbas, S.; Deliorman, O.D. Antidiabetic effect and antioxidant potential of Rosa canina fruits. Pharmacogn. Mag. 2009, 5, 309. [Google Scholar] [CrossRef]
- Taghizadeh, M.; Rashidi, A.A.; Taherian, A.A.; Vakili, Z.; Sajad Sajadian, M.; Ghardashi, M. Antidiabetic and antihyperlipidemic effects of ethanol extract of Rosa canina L. Fruit on diabetic rats: An experimental study with histopathological evaluations. J. Evid. Based Complement. Altern. Med. 2016, 21, NP25–NP30. [Google Scholar] [CrossRef] [PubMed]
- Sagdullaev, B.; Sagdullaev, S.S.; Aripov, K.N. Aqueous extraction of the fruit of Rosa canina. Chem. Nat. Compd. 1997, 33, 449–452. [Google Scholar] [CrossRef]
- Can, A.; Akev, N.; Sütlüpinar, N. Effect of rose hips on blood glucose level. Acta Pharm. Turc. 1992, 34, 17–22. [Google Scholar]
- Fattahi, A.; Niyazi, F.; Shahbazi, B.; Farzaei, M.H.; Bahrami, G. Antidiabetic mechanisms of Rosa canina fruits. J. Evid. Based Complement. Altern. Med. 2017, 22, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Ghadyale, V.; Takalikar, S.; Haldavnekar, V.; Arvindekar, A. Effective control of postprandial glucose level through inhibition of intestinal alpha glucosidase by Cymbopogon martinii (roxb.). J. Evid. Based Complement. Altern. Med. 2011, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Kumar, R.; Laloo, D.; Hemalatha, S. Diabetes mellitus: An overview on its pharmacological aspects and reported medicinal plants having antidiabetic activity. Asian Pac. J. Trop. Biomed. 2012, 2, 411–420. [Google Scholar] [CrossRef]
- Ninomiya, K.; Matsuda, H.; Kubo, M.; Morikawa, T.; Nishida, N.; Yoshikawa, M. Potent anti-obese principle from Rosa canina: Structural requirements and mode of action of trans-tiliroside. Bioorg. Med. Chem. Lett. 2007, 17, 3059–3064. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Miller, M.; Tighe, A.P.; Davidson, M.H.; Schaefer, E.J. Omega-3 fatty acids and coronary heart disease risk: Clinical and mechanistic perspectives. Atherosclerosis 2008, 197, 12–24. [Google Scholar] [CrossRef] [PubMed]
- Raiciu, A.D.; Popescu, M.; Ivopol, G.C.; Bordei, N.; Alexandru, G.; Crisan, I.; Manea, S.; Dima, S.O. Therapeutic applications of vegetable oils and GC-MS evaluation of ω-3, ω-6 and ω-9 amounts in six oleaginous plants. Rev. Chim. 2016, 67, 2449–2453. [Google Scholar]
- Yoshikawa, T.; Shimano, H.; Yahagi, N.; Ide, T.; Amemiya-Kudo, M.; Matsuzaka, T.; Nakakuki, M.; Tomita, S.; Okazaki, H.; Tamura, Y. Polyunsaturated fatty acids suppress Sterol Regulatory Element-Binding Protein 1C promoter activity by inhibition of liver x receptor (LXR) binding to lxr response elements. J. Biol. Chem. 2002, 277, 1705–1711. [Google Scholar] [CrossRef] [PubMed]
- Adkins, Y.; Kelley, D.S. Mechanisms underlying the cardioprotective effects of ω-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2010, 21, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Morgen, C.S.; Sørensen, T.I. Obesity: Global trends in the prevalence of overweight and obesity. Nat. Rev. Endocrinol. 2014, 10, 513–514. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.Y. Obesity-related digestive diseases and their pathophysiology. Gut Liver 2017, 11, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Lastra, G.; Sowers, J.R. Obesity and cardiovascular disease: Role of adipose tissue, inflammation, and the renin-angiotensin-aldosterone system. Horm. Mol. Biol. Clin. Investig. 2013, 15, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Wensveen, F.M.; Valentić, S.; Šestan, M.; Turk Wensveen, T.; Polić, B. The “big bang” in obese fat: Events initiating obesity—induced adipose tissue inflammation. Eur. J. Immunol. 2015, 45, 2446–2456. [Google Scholar] [CrossRef] [PubMed]
- Connaughton, R.M.; McMorrow, A.M.; McGillicuddy, F.C.; Lithander, F.E.; Roche, H.M. Impact of anti-inflammatory nutrients on obesity-associated metabolic-inflammation from childhood through to adulthood. Proc. Nutr. Soc. 2016, 75, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Grygiel-Gorniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical implications—A review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, A.; Nishida, N.; Matsuura, Y.; Shibata, N. Rosehip extract inhibits lipid accumulation in white adipose tissue by suppressing the expression of Peroxisome Proliferator-Activated Receptor Gamma. Prev. Nutr. Food Sci. 2013, 18, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, A.; Nishida, N.; Fukuhara, I.; Noro, A.; Kozai, Y.; Sato, H.; Matsuura, Y. Daily intake of rosehip extract decreases abdominal visceral fat in preobese subjects: A randomized, double-blind, placebo-controlled clinical trial. Diabetes Metab. Syndr. Obes. 2015, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Zuk, A.; Bonventre, J.V. Acute kidney injury. Annu. Rev. Med. 2016, 67, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Ashtiyani, S.C.; Najafi, H.; Jalalvandi, S.; Hosseinei, F. Protective effects of Rosa canina L fruit extracts on renal disturbances induced by reperfusion injury in rats. Iran. J. Kidney Dis. 2013, 7, 290. [Google Scholar]
- Zhao, L.; Xu, L.; Tao, X.; Han, X.; Yin, L.; Qi, Y.; Peng, J. Protective effect of the total flavonoids from Rosa laevigata michx fruit on renal ischemia-reperfusion injury through suppression of oxidative stress and inflammation. Molecules 2016, 21, 952. [Google Scholar] [CrossRef] [PubMed]
- Kashihara, N.; Haruna, Y.; K Kondeti, V.; S Kanwar, Y. Oxidative stress in diabetic nephropathy. Curr. Med. Chem. 2010, 17, 4256–4269. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Liao, Q.; Luo, Y.; Qing, Z.; Zhang, Q.; He, G. Renal protective effect of Rosa laevigata michx. By the inhibition of oxidative stress in streptozotocin-induced diabetic rats. Mol. Med. Rep. 2012, 5, 1548–1554. [Google Scholar] [PubMed]
- Sakhaee, K. Recent advances in the pathophysiology of nephrolithiasis. Kidney Int. 2009, 75, 585–595. [Google Scholar] [CrossRef] [PubMed]
- Tracy, C.R.; Pearle, M.S. Update on the medical management of stone disease. Curr. Opin. Urol. 2009, 19, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Tayefi-Nasrabadi, H.; Sadigh-Eteghad, S.; Aghdam, Z. The effects of the hydroalcohol extract of Rosa canina l. Fruit on experimentally nephrolithiasic wistar rats. Phytother. Res. 2012, 26, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Kale, I.; Khan, M.A.; Irfan, Y.; Veerana, G.A. Hepatoprotective potential of ethanolic and aqueous extract of flowers of Sesbania grandiflora (linn) induced by ccl4. Asian Pac. J. Trop. Biomed. 2012, 2, S670–S679. [Google Scholar] [CrossRef]
- Pareek, A.; Godavarthi, A.; Issarani, R.; Nagori, B.P. Antioxidant and hepatoprotective activity of Fagonia schweinfurthii (Hadidi) hadidi extract in carbon tetrachloride induced hepatotoxicity in HEPG2 cell line and rats. J. Ethnopharmacol. 2013, 150, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, H.; Hosseinzadeh, S.; Touri, M.A.; Ghavamzadeh, M.; Barmak, M.J. Hepatoprotective effect of Rosa canina fruit extract against carbon tetrachloride induced hepatotoxicity in rat. Avicenna J. Phytomed. 2016, 6, 181. [Google Scholar] [PubMed]
- Ringman, J.M.; Goate, A.; Masters, C.L.; Cairns, N.J.; Danek, A.; Graff-Radford, N.; Ghetti, B.; Morris, J.C.; Network, D.I.A. Genetic heterogeneity in alzheimer disease and implications for treatment strategies. Curr. Neurol. Neurosci. Rep. 2014, 14, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.H.; Wang, X.; Zhu, X. Mitochondrial defects and oxidative stress in alzheimer disease and parkinson disease. Free Radic. Biol. Med. 2013, 62, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Esfandiary, E.; Karimipour, M.; Mardani, M.; Ghanadian, M.; Alaei, H.A.; Mohammadnejad, D.; Esmaeili, A. Neuroprotective effects of Rosa damascena extract on learning and memory in a rat model of amyloid-β-induced alzheimer’s disease. Adv. Biomed. Res. 2015, 4, 131. [Google Scholar] [PubMed]
- Semwal, D.K.; Semwal, R.B.; Combrinck, S.; Viljoen, A. Myricetin: A dietary molecule with diverse biological activities. Nutrients 2016, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- DeToma, A.S.; Choi, J.S.; Braymer, J.J.; Lim, M.H. Myricetin: A naturally occurring regulator of Metal-Induced Amyloid-β aggregation and neurotoxicity. ChemBioChem 2011, 12, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, C.C.T.; Almeida, A.B.; Araújo, P.V.P.; de Abreu, R.N.D.C.; Chaves, E.M.C.; de Vale, O.C.; Macêdo, D.S.; Woods, D.J.; de Franca Fonteles, M.M.; Vasconcelos, S.M.M. Oxidative stress and epilepsy: Literature review. Oxid. Med. Cell. Longev. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Diniz, T.C.; Silva, J.C.; de Lima-Saraiva, S.R.G.; de Almeida Ribeiro, F.P.R.; Pacheco, A.G.M.; de Freitas, R.M.; Quintans-Júnior, L.J.; de Souza Siqueira Quintans, J.; Mendes, R.L.; da Silva Almeida, J.R.G. The role of flavonoids on oxidative stress in epilepsy. Oxid. Med. Cell. Longev. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Homayoun, M.; Seghatoleslam, M.; Pourzaki, M.; Shafieian, R.; Hosseini, M.; Bideskan, A.E. Anticonvulsant and neuroprotective effects of Rosa damascena hydro-alcoholic extract on rat hippocampus. Avicenna J. Phytomed. 2015, 5, 260. [Google Scholar] [PubMed]
- Uttara, B.; Singh, A.V.; Zamboni, P.; Mahajan, R. Oxidative stress and neurodegenerative diseases: A review of upstream and downstream antioxidant therapeutic options. Curr. Neuropharmacol. 2009, 7, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Vidaña, D.I.; Ngai, S.P.-C.; He, W.; Chow, J.K.-W.; Lau, B.W.-M.; Tsang, H.W.-H. The effectiveness of aromatherapy for depressive symptoms: A systematic review. J. Evid. Based Complement. Altern. Med. 2017, 2017, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hongratanaworakit, T. Relaxing effect of rose oil on humans. Nat. Prod. Commun. 2009, 4, 291–296. [Google Scholar] [PubMed]
- Nazıroğlu, M.; Kozlu, S.; Yorgancıgil, E.; Uğuz, A.C.; Karakuş, K. Rose oil (from Rosa damascena Mill.) vapor attenuates depression-induced oxidative toxicity in rat brain. J. Nat. Med. 2013, 67, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Beck, L.A. Immunologic targets in atopic dermatitis and emerging therapies: An update. Am. J. Clin. Dermatol. 2016, 17, 425–443. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Jeong, M.S.; Park, K.J.; Choi, Y.W.; Seo, S.J.; Lee, M.W. Topical application of Rosa multiflora root extract improves atopic dermatitis-like skin lesions induced by mite antigen in NC/NGA mice. Biol. Pharm. Bull. 2014, 37, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Rittié, L.; Fisher, G.J. Natural and sun-induced aging of human skin. Cold Spring Harb. Perspect. Med. 2015, 5, a015370. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzyme Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Ikeda, K.; Saito, M. Inhibitory effect of rose hip (Rosa canina L.) on melanogenesis in mouse melanoma cells and on pigmentation in brown guinea pigs. Biosci. Biotechnol. Biochem. 2011, 75, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Phetcharat, L.; Wongsuphasawat, K.; Winther, K. The effectiveness of a standardized rose hip powder, containing seeds and shells of Rosa canina, on cell longevity, skin wrinkles, moisture, and elasticity. Clin. Interv. Aging 2015, 10, 1849. [Google Scholar] [PubMed]
- Kim, J.K.; Mun, S.; Kim, M.S.; Kim, M.B.; Sa, B.K.; Hwang, J.K. 5,7-dimethoxyflavone, an activator of PPARα/γ, inhibits uvb-induced mmp expression in human skin fibroblast cells. Exp. Dermatol. 2012, 21, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Kim, D.H.; Nho, Y.-H.; Park, J.-E.; Kim, S.-N.; Choi, E.H. A mixture of extracts of kochia scoparia and Rosa multiflora with PPARα/γ dual agonistic effects prevents photoaging in hairless mice. Int. J. Mol. Sci. 2016, 17, 1919. [Google Scholar] [CrossRef] [PubMed]
- Altemus, M.; Rao, B.; Dhabhar, F.S.; Ding, W.; Granstein, R.D. Stress-induced changes in skin barrier function in healthy women. J. Investig. Dermatol. 2001, 117, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Fukada, M.; Kano, E.; Miyoshi, M.; Komaki, R.; Watanabe, T. Effect of “rose essential oil” inhalation on stress-induced skin-barrier disruption in rats and humans. Chem. Sens. 2012, 37, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Malfertheiner, P.; Chan, F.K.; McColl, K.E. Peptic ulcer disease. Lancet 2009, 374, 1449–1461. [Google Scholar] [CrossRef]
- Gürbüz, I.; Üstün, O.; Yesilada, E.; Sezik, E.; Kutsal, O. Anti-Ulcerogenic activity of some plants used as folk remedy in Turkey. J. Ethnopharmacol. 2003, 88, 93–97. [Google Scholar] [CrossRef]
- Rao, A.; Gurfinkel, D. The bioactivity of saponins: Triterpenoid and steroidal glycosides. Drug Metabol. Drug Interact. 2000, 17, 211–236. [Google Scholar] [CrossRef] [PubMed]
- Carlo, G.D.; Mascolo, N.; Izzo, A.A.; Capasso, F. Effects of quercetin on the gastrointestinal tract in rats and mice. Phytother. Res. 1994, 8, 42–45. [Google Scholar] [CrossRef]
- Meli, R.; Di Carlo, G.; Capasso, F. Inhibitory action of quercetin on intestinal transit in mice. Phytother. Res. 1990, 4, 201–202. [Google Scholar] [CrossRef]
- Lattanzio, F.; Greco, E.; Carretta, D.; Cervellati, R.; Govoni, P.; Speroni, E. In vivo anti-inflammatory effect of Rosa canina L. extract. J. Ethnopharmacol. 2011, 137, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Talib, W.H.; Mahasneh, A.M. Antimicrobial, cytotoxicity and phytochemical screening of jordanian plants used in traditional medicine. Molecules 2010, 15, 1811–1824. [Google Scholar] [CrossRef] [PubMed]
- Yi, O.; Jovel, E.M.; Towers, G.N.; Wahbe, T.R.; Cho, D. Antioxidant and antimicrobial activities of native Rosa sp. from British Columbia, Canada. Int. J. Food Sci. Nutr. 2007, 58, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Kumarasamy, Y.; Cox, P.J.; Jaspars, M.; Nahar, L.; Sarker, S.D. Screening seeds of scottish plants for antibacterial activity. J. Ethnopharmacol. 2002, 83, 73–77. [Google Scholar] [CrossRef]
- Olech, M.; Nowak, R.; Pecio, Ł.; Łoś, R.; Malm, A.; Rzymowska, J.; Oleszek, W. Multidirectional characterisation of chemical composition and health-promoting potential of Rosa rugosa hips. Nat. Prod. Res. 2016, 31, 667–671. [Google Scholar] [CrossRef] [PubMed]
- Frey, F.M.; Meyers, R. Antibacterial activity of traditional medicinal plants used by haudenosaunee peoples of New York state. BMC Complement. Altern. Med. 2010, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Miyasaki, Y.; Rabenstein, J.D.; Rhea, J.; Crouch, M.L.; Mocek, U.M.; Kittell, P.E.; Morgan, M.A.; Nichols, W.S.; Van Benschoten, M.M.; Hardy, W.D.; et al. Isolation and characterization of antimicrobial compounds in plant extracts against multidrug-resistant acinetobacter baumannii. PLoS ONE 2013, 8, e61594. [Google Scholar] [CrossRef] [PubMed]
- Sarabhai, S.; Harjai, K.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula retz. Increase the susceptibility of pseudomonas aeruginosa to stress by inhibiting polyphosphate kinase. J. Appl. Microbiol. 2015, 118, 817–825. [Google Scholar] [CrossRef] [PubMed]
- Shiota, S.; Shimizu, M.; Sugiyama, J.; Morita, Y.; Mizushima, T.; Tsuchiya, T. Mechanisms of action of corilagin and tellimagrandin I that remarkably potentiate the activity of β-lactams against methicillin-resistant staphylococcus aureus. Microbiol. Immunol. 2004, 48, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Horváth, G.; Molnár, P.; Radó-Turcsi, E.; Deli, J.; Kawase, M.; Satoh, K.; Tanaka, T.; Tani, S.; Sakagami, H.; Gyémánt, N. Carotenoid composition and in vitro pharmacological activity of rose hips. Acta Biochim. Pol. 2012, 59, 129–132. [Google Scholar] [PubMed]
- Jang, S.H.; Lim, J.W.; Morio, T.; Kim, H. Lycopene inhibits helicobacter pylori-induced ATM/ATR-dependent DNA damage response in gastric epithelial ags cells. Free Radic. Biol. Med. 2012, 52, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Flores-Mireles, A.L.; Walker, J.N.; Caparon, M.; Hultgren, S.J. Urinary tract infections: Epidemiology, mechanisms of infection and treatment options. Nat. Rev. Microbiol. 2015, 13, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Miano, R.; Germani, S.; Vespasiani, G. Stones and urinary tract infections. Urol Int. 2007, 79, 32–36. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.; Bharti, P.; Seasotiya, L.; Malik, A.; Dalal, S. In vitro screening and evaluation of some indian medicinal plants for their potential to inhibit jack bean and bacterial ureases causing urinary infections. Pharm. Biol. 2015, 53, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.T.; Žemlička, M. Plant-derived urease inhibitors as alternative chemotherapeutic agents. Arch. Phar. 2016, 349, 507–522. [Google Scholar] [CrossRef] [PubMed]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M.J. Colorectal carcinoma: A general overview and future perspectives in colorectal cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Kamijo, M.; Kanazawa, T.; Funaki, M.; Nishizawa, M.; Yamagishi, T. Effects of Rosa rugosa petals on intestinal bacteria. Biosci. Biotechnol. Biochem. 2008, 72, 773–777. [Google Scholar] [CrossRef] [PubMed]
- Puupponen-Pimiä, R.; Nohynek, L.; Alakomi, H.-L.; Oksman-Caldentey, K.-M. Bioactive berry compounds—Novel tools against human pathogens. Appl. Microbiol. Biotechnol. 2005, 67, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Tamura, S.; Yang, G.-M.; Yasueda, N.; Matsuura, Y.; Komoda, Y.; Murakami, N. Tellimagrandin I, HCV invasion inhibitor from Rosae rugosae Flos. Bioorg. Med. Chem. Lett. 2010, 20, 1598–1600. [Google Scholar] [CrossRef] [PubMed]
- Granich, R.; Gupta, S.; Hersh, B.; Williams, B.; Montaner, J.; Young, B.; Zuniga, J.M. Trends in aids deaths, new infections and art coverage in the top 30 countries with the highest aids mortality burden; 1990–2013. PLoS ONE 2015, 10, e0131353. [Google Scholar] [CrossRef] [PubMed]
- Pau, A.K.; George, J.M. Antiretroviral therapy: Current drugs. Infect. Dis. Clin. North. Am. 2014, 28, 371–402. [Google Scholar] [CrossRef] [PubMed]
- Fu, M.; Ng, T.; Jiang, Y.; Pi, Z.; Liu, Z.; Li, L.; Liu, F. Compounds from rose (Rosa rugosa) flowers with human immunodeficiency virus type 1 reverse transcriptase inhibitory activity. J. Pharm. Pharmacol. 2006, 58, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.-Q.; Yang, X.-Z.; Miao, J.-H.; Tang, C.-P.; Ke, C.-Q.; Zhang, J.-B.; Ma, X.-J.; Ye, Y. New triterpene glucosides from the roots of Rosa laevigata Michx. Molecules 2008, 13, 2229–2237. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rose Species | Cancer Cell Line | Reference |
|---|---|---|
| Rosa canina | Cervix epithelioid carcinoma (HeLa) | Tumbas et al. [14] |
| Colon carcinoma (HT-29) | ||
| Rosa canina | Non-small cell lung cancer (NCI-H460) | Guimarães et al. [85] |
| Colon carcinoma (HCT-15) | ||
| Hepatocellular carcinoma (HepG2) | ||
| Rosa canina | Colon carcinoma (Caco-2) | Jiménez et al. [21] |
| Activity | Active Compound | Mechanism of Action | References |
|---|---|---|---|
| Anti-inflammatory | GOPO (1,2-di-O-α-linolenoyl-3-O-β-d-galactopyranosyl-sn-glycerol) | Reduction in peripheral blood polymorphonuclear leukocytes, neutrophils and monocytes migration Reduction in the levels of C-reactive protein (CRP) | Larsen et al. [61] Winther et al. [105] Kharazmi et al. [102] |
| Fatty acids (triterpenoic acids, ursolic acid, oleanolic acid and betulinic acid) | Inhibition of COX-1 and COX-2 | Jäger et al. [109] Wenzing et al. [110] | |
| Reduction in pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, (IFN)-γ, IL-12) and chemokines CCL5 (RANTES), IP-10 (CXCL10) production | Yan et al. [106] Schwager et al. [107,108] | ||
| Inhibition of NF-κB related inflammatory response | Phytosterols β-sitosterol | Attenuation of NF-κB phosphorylation | Whang et al. [115] |
| Gallic acid | Reestablishment IκBα and NF-κB association | Choi et al. [116] | |
| Astragalin and tormentic acid | Inhibition of IkBα phosphorylation and degradation | An et al. [117] Cheng et al. [120] | |
| Antioxidant | Antioxidants (vitamin C, vitamin E, carotenoids, polyphenols and antioxidant enzymes) | Reduction of ROS production | Kirkeskov et al. [101] Schwager et al. [107] Jimenez et al. [21] |
| Inhibition of NO release from macrophages | Kirkeskov et al. [101] Schwager et al. [107] Kim et al. [111] | ||
| Protection against cell apoptosis, DNA and mitochondrial H2O2-induced damage and amyloid β peptide-induced oxidative injury | Choi et al. [112] Liu et al. [113] | ||
| Aqueous extract | Inhibition of RANKL-induced osteoclastogenesis | Cheng et al. [120] |
| Rosa Species | Bacteria Strain | Mean Diameter of Inhibition Zone (mm) | Reference |
|---|---|---|---|
| R. multiflora | E. coli | 9.0 | Frey et al. [196] |
| S. typhimurium | 8.1 | ||
| R. nutkana | S. aureus | 15.4 | Yi et al. [193] |
| MRSA | 17 | ||
| E. faecalis | 18 | ||
| B. subtilis | 8 | ||
| R. woodsii | S. aureus | 10 | |
| MRSA | 9.8 | ||
| E. faecalis | 9 | ||
| R. psiocarpa | S. aureus | 9.4 | |
| MRSA | 7.4 | ||
| E. faecalis | 8.2 |
| Rosa Species | Bacteria Strain | MIC (mg/mL) | Reference |
|---|---|---|---|
| R. canina | E. coli 8110 | 0.1 | Kumarasamy et al. [194] |
| R. rugosa | S. epidermis | 1.25 | Olech et al. [195] |
| S. aureus | 1.25 | ||
| B. subtilis | 1.25 | ||
| M. luteus | 0.625 | ||
| E. coli | 1.25 | ||
| K. pneumoniae | 1.25 | ||
| P. aeruginosa | 1.25 | ||
| P. mirabilis | 1.25 |
| Rosa Species | Bacteria Strain | % Inhibition | Reference |
|---|---|---|---|
| R. damascena | P. aeruginosa | 80.49 | Talib et al. [192] |
| E. coli | 60.69 | ||
| S. typhimurium | 100.82 | ||
| B. cereus | 101.09 | ||
| MRSA | 95.75 |
| Rosa Species | Yeast Strain | Mean Diameter of Inhibition Zone (mm) | Reference |
|---|---|---|---|
| R. nutkana | C. albicans | 25 | Yi et al. [197] |
| R. woodsii | 22.8 | ||
| R. pisocarpa | 10 |
| Rosa Species | Yeast Strain | MIC (mg/mL) | Reference |
|---|---|---|---|
| R. rugosa | C. albicans | 0.156 | Olech et al. [195] |
| C. parapsilosis | 0.156 |
| Rosa Species | Yeast Strain | % Inhibition | Reference |
|---|---|---|---|
| R. damascena | C. albicans | 98.87 | Talib et al. [192] |
| A. niger | 30.72 |
| Phytochemical | Disease of Application | Mechanism of Action | References |
|---|---|---|---|
| Ascorbic acid | Cancer | Anti-inflammatory Antioxidant | Jiménez et al. [21] |
| Aging | Antioxidant and anti-inflammatory effect Contribution to skin and collagen formation | Phetcharat et al. [181] | |
| Arthritis rheumatoid | Inhibition of NO release from macrophages | Kirkeskov et al. [101] Kim et al. [111] Schwager et al. [108] | |
| Osteoporosis | Stimulation osteoblast differentiation and matrix synthesis | Devareddy et al. [124] Adom et al. [125] | |
| Polysaccharides | Cancer | Induction of cytotoxicity in reduction of cancer metastasis by reducing the levels of matrix metalloprotease-9 (MMP-9) | Guo et al. [96] Lee et al. [97] |
| Polysaccharide-peptide complex and other polymers | Retroviral infection | Inhibition of viral retrotranscriptase | Fu et al. [212] |
| Myricetin | Alzheimer | Inhibition of amyloid β formation and tau-antagonist activity | Semwal et al. [167] DeToma et al. [168] |
| Quercetin | Age spots | Inhibition of tyrosinase | Fujii et al. [180] |
| Gallic acid | Arthritis Rheumatoid | Reestablishment IκBα and NF-κB association | Choi et al. [116] |
| Diabetes | Inhibition of α-amylase and α-glucosidase | Ghadyale et al. [138] | |
| Ellagic acid | Cancer | Anti-mutagenic and anti-carcinogenic effect | Festa et al. [29] Whitley et al. [30] |
| Bacterial infections | Downregulation of polyphosphatase kinase 1 (PPK1) | Sarabhai et al. [198] | |
| Tellimagrandin I | Bacterial infections | Inhibition of penicillin binding protein 2′ (PBP2) and enhancement of β-lactam antibiotics effect | Shiota et al. [199] |
| 1,25-Dihydroxyvitamin D3 analogues | Acute myeloid leukemia | Induction of differentiation | Zhamanbayeva et al. [88] Corcoran et al. [89] Baurska et al. [90] |
| Tannin RM-3 | Atopic dermatitis | Anti-inflammatory activity (reduction of the levels of cyclooxygenase 2 mediators and inducible nitric oxide synthase) Suppression Th2-polarized immune system | Wang et al. [176] Park et al. [177] |
| Unidentified phenols | Bacterial infections | Membrane permeabilisation, extracellular enzymes inhibition, nutrient and energy deprivation by an hyperacidification of plasma membrane interface that disrupts H+-ATPase | Olech et al. [195] |
| Dysbiosis | Specific inhibition of pathogenic bacteria growth | Kamijo et al. [207] | |
| Prostate cancer | Anti-histone acetyltransferase activity | Lee et al. [91] Takayama et al. [92] | |
| Urinary tract infections (UTIs) and pyelonephritis | Urease inhibitor | Hassan et al. [205] Bai et al. [204] | |
| Isoflavonoid phytoestrogen | Cancer | Stimulation of growing oestrogen dependent cells | Uifălean et al. [87] |
| Flavonoids | Obesity | Reduction of PPARγ expression, and prevention of lipid accumulation | Ninomiya et al. [140] Nagatomo et al. [151] |
| Epilepsy | Reduction of lipid peroxidation and anticonvulsive effect | Diniz et al. [170] Homayoun et al. [171] Uttara et al. [172] | |
| Triterpenoic acids, ursolic acid, oleanolic acid and betulinic acid | Arthritis Reumatoid | Inhibition of COX-1 and COX-2 | Jäger et al. [109] Wenzing et al. [110] |
| Lycopene and other carotenes | Bacterial infections | Anti-H. pylori activity | Horváth et al. [200] |
| trans-Tiliroside | Hyperlipidaemia | Reduction of reduced plasma and liver triglyceride free fatty acid (FFA) levels. Induction of peroxisome proliferator-activated receptor α (PPAR-α) expression | Ninomiya et al. [140] |
| Polyunsaturated fatty acid ω-3, 6 and 9 | Hyperlipidaemia | Reduction of TG synthesis, reduction of the expression of hepatic very low-density lipoprotein (VLDL)-TG, hepatic lipase and Apo CIII and increase Apo CII and VLDL-receptor | Adkins et al. [144] |
| Galactolipids GOPO (1,2-di-O-α-linolenoyl-3-O-β-d-galactopyranosyl-sn-glycerol) | Arthritis Rheumatoid | Reduction in peripheral blood polymorphonuclear leukocytes, neutrophils and monocytes migration Reduction in the levels of C-reactive protein (CRP) | Larsen et al. [61] Kharazmi et al. [102] Whinter et al. [105] |
| Unidentified compounds from whole extract | Arthritis rheumatoid | Inhibition of RANKL-induced osteoclastogenesis | Cheng et al. [120] |
| Diabetes | Hypoglycemic effect and induction of β-cell proliferation | Taghizadeh et al. [134] Fattahi et al. [137] | |
| Acute kidney injury Diabetic nephropathy | Anti-inflammatory effect Reduction of ROS levels | Zhao et al. [155] Zhou et al. [157] | |
| Peptic ulcer | Antioxidant effect | Nazıroğlu et al. [175] | |
| Depression | Antioxidant effect | Nazıroğlu et al. [175] | |
| Antiviral | Inhibition of viral invasion capacity | McCutcheon et al. [65] | |
| Unidentified components from fruit extract | Hepatic injuries | Reduction of peroxidation of unsaturated fatty acids | Carlo et al. [187] Meli et al. [188] |
| Unidentified components from root extract | Atopic dermatitis | Anti-inflammatory (reduction of cyclooxygenase 2 inducible nitric oxide synthase levels) Suppression Th2-polarized immune system | Park et al. [177] |
| Unidentified components from leaf extract | Diarrhoeal | Anti-secretory activity, inhibition of acetylcholine and histamine activity | Carlo et al. [187] Meli et al. [188] Rao et al. [186] |
| Unidentified components from rose powder | Wrinkles | Antioxidant effect: scavenging of reactive oxygen species produced by UV-radiation and decrease of matrix metalloprotease proteins levels | Phetcharat et al. [181] Jeo et al. [183] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mármol, I.; Sánchez-de-Diego, C.; Jiménez-Moreno, N.; Ancín-Azpilicueta, C.; Rodríguez-Yoldi, M.J. Therapeutic Applications of Rose Hips from Different Rosa Species. Int. J. Mol. Sci. 2017, 18, 1137. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061137
Mármol I, Sánchez-de-Diego C, Jiménez-Moreno N, Ancín-Azpilicueta C, Rodríguez-Yoldi MJ. Therapeutic Applications of Rose Hips from Different Rosa Species. International Journal of Molecular Sciences. 2017; 18(6):1137. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061137
Chicago/Turabian StyleMármol, Inés, Cristina Sánchez-de-Diego, Nerea Jiménez-Moreno, Carmen Ancín-Azpilicueta, and María Jesús Rodríguez-Yoldi. 2017. "Therapeutic Applications of Rose Hips from Different Rosa Species" International Journal of Molecular Sciences 18, no. 6: 1137. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18061137