The Venom of the Spine-Bellied Sea Snake (Hydrophis curtus): Proteome, Toxin Diversity and Intraspecific Variation


Abstract
:1. Introduction
2. Results
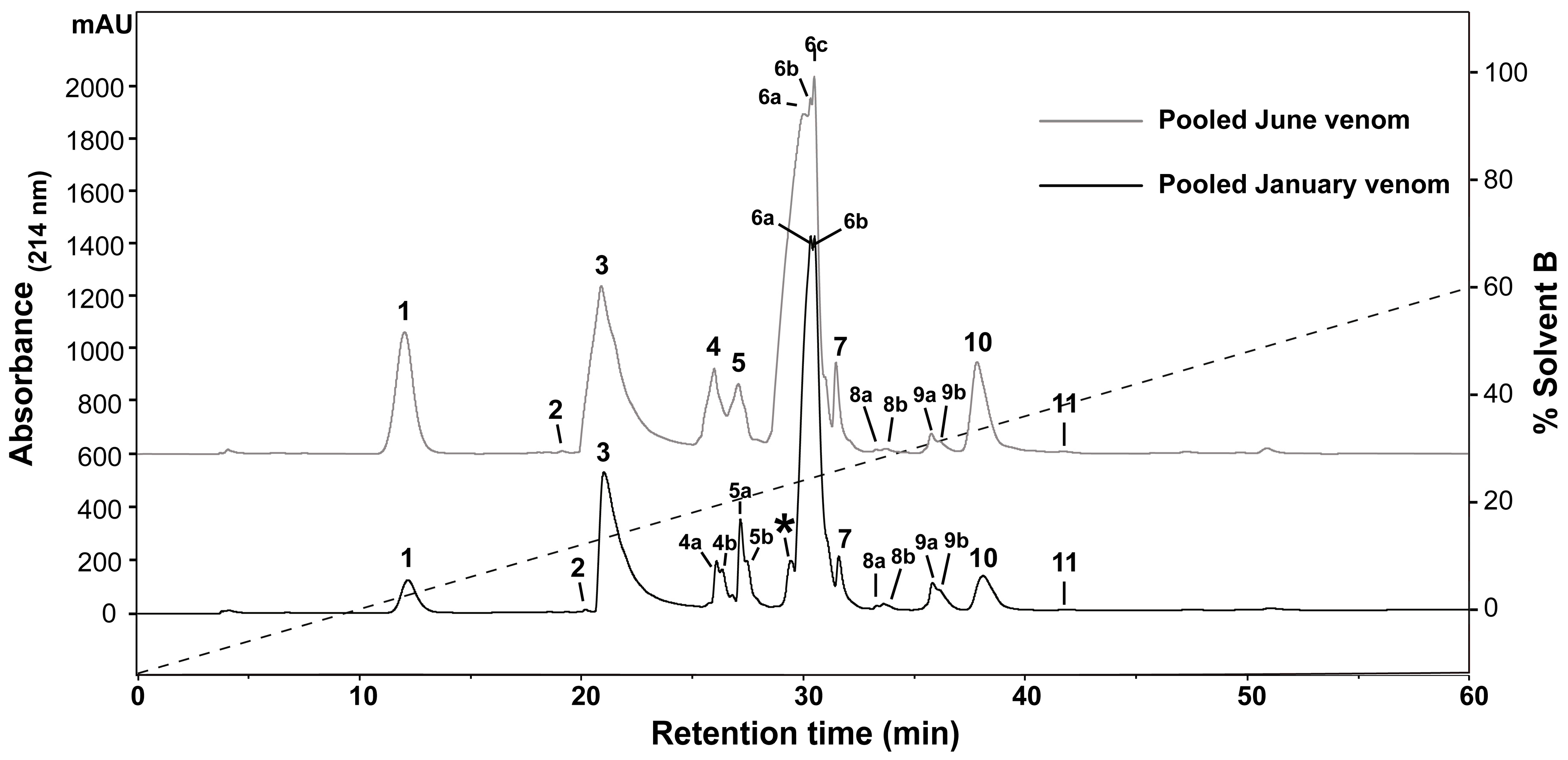
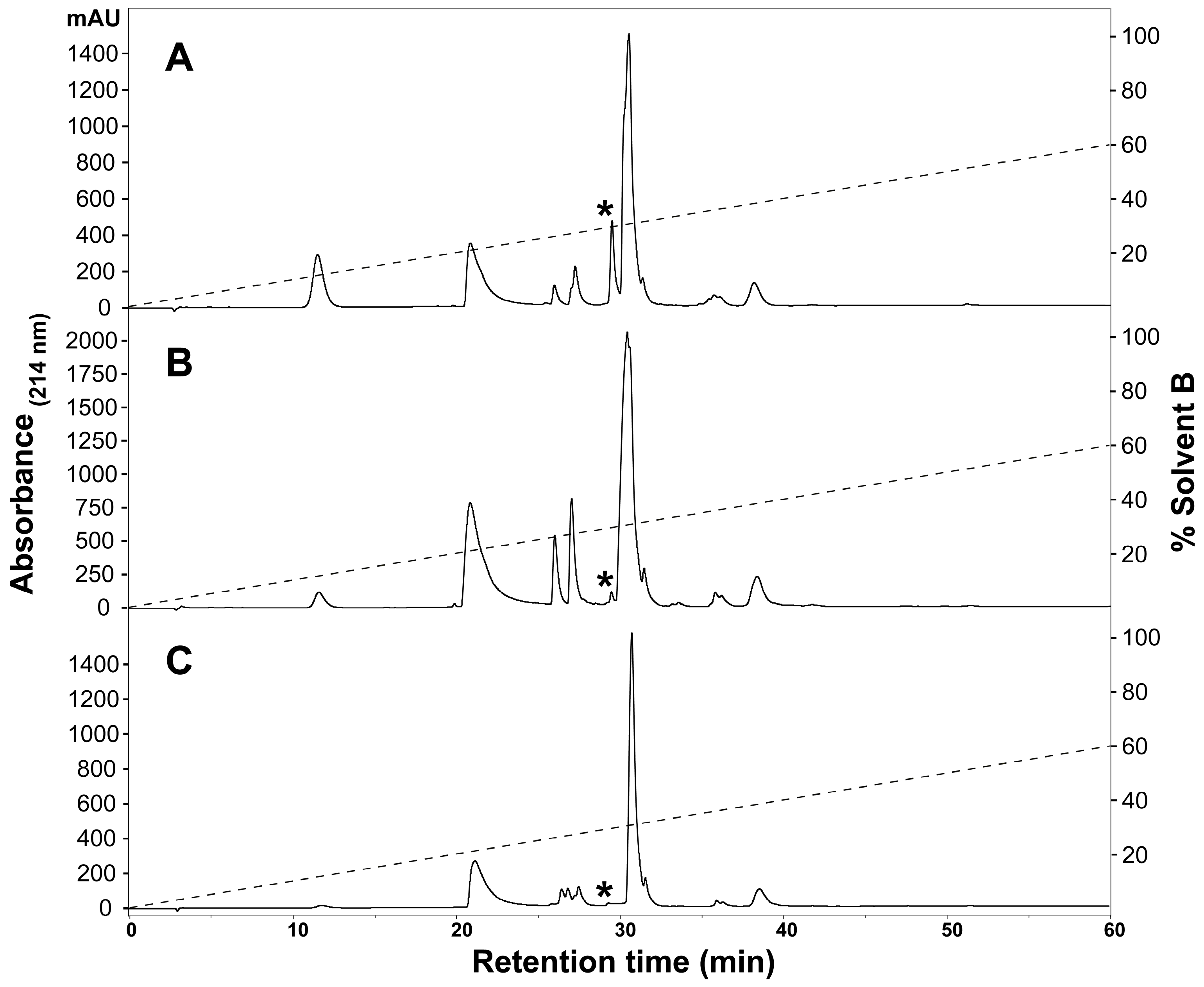
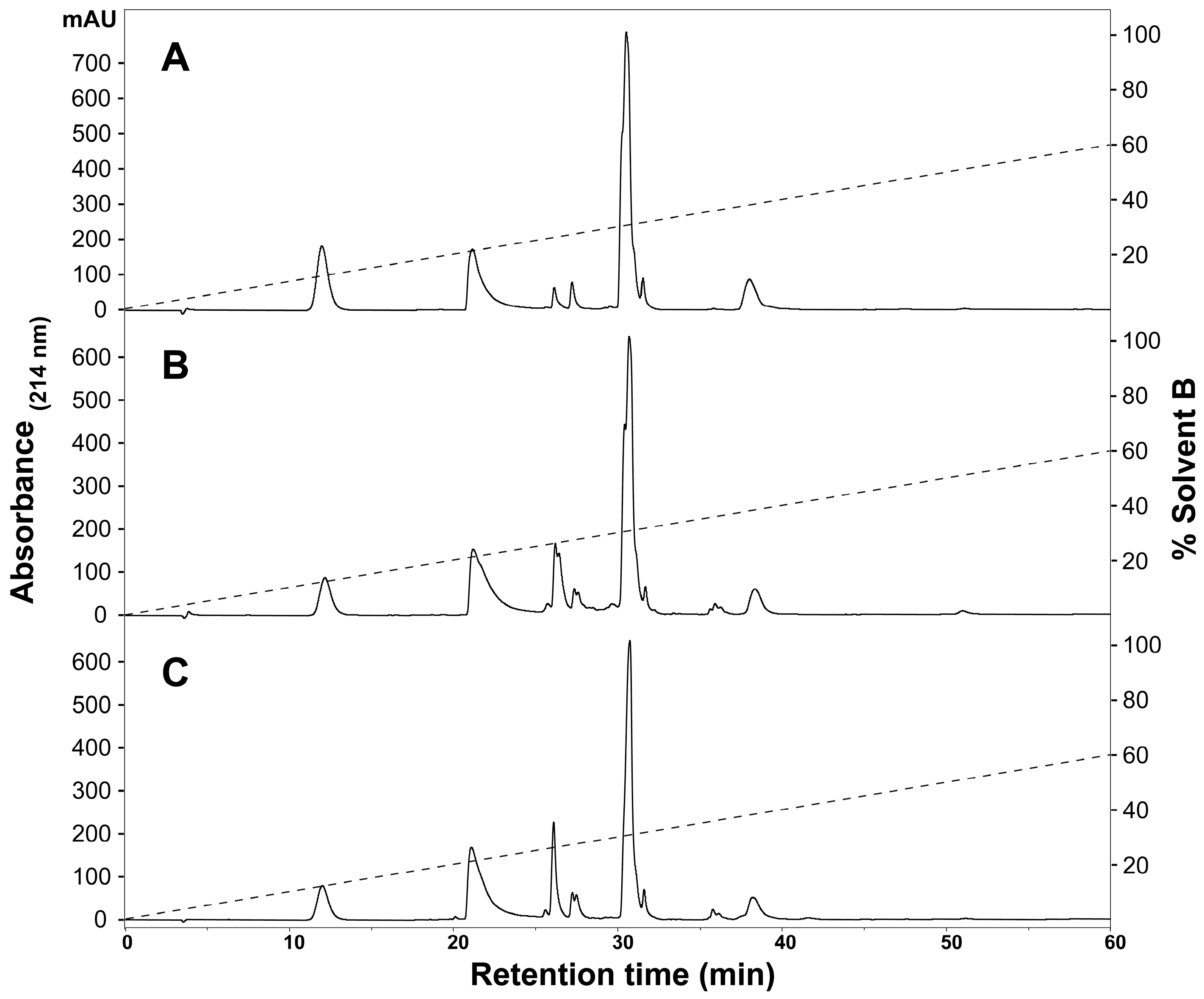
2.1. Assessment of Variation in H. curtus Venom
2.2. Protein Identification with ProteinPilot and MASCOT
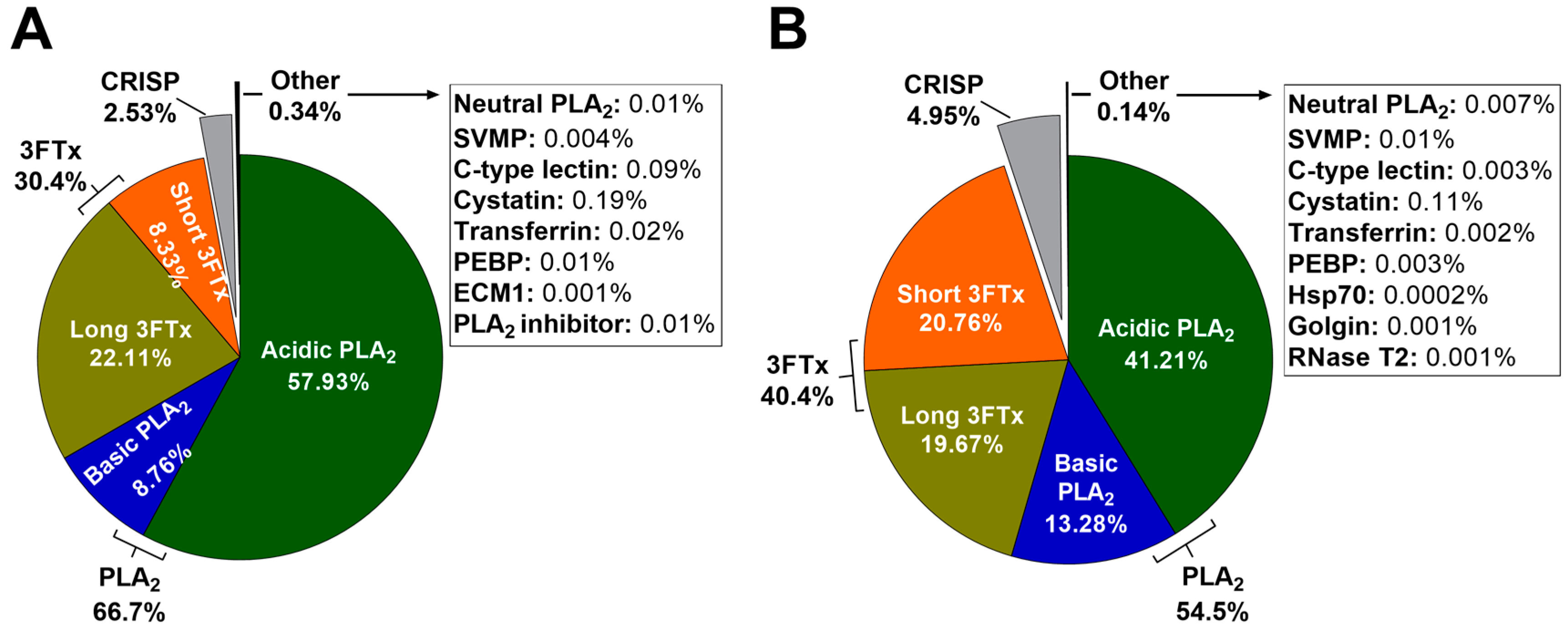
2.3. Relative Protein Abundance Estimations Using MASCOT emPAI
3. Discussion
3.1. Assessment of Variation in H. curtus Venom
3.2. Protein Identification and Relative Abundance Estimations with ProteinPilot and MASCOT
3.3. Toxin Diversity in H. curtus Venom
4. Materials and Methods
4.1. Venom Samples
4.2. Pooling Venom Samples
4.3. Venom Fractionation and Comparative Profiling with Reversed-Phase HPLC
4.4. Venom ‘Fingerprint’ Analysis Using LC-MS
4.5. In-Solution Trypsin Digestion
4.6. LC-ESI-MS/MS Tandem Mass Spectrometry
4.7. Protein Identification with ProteinPilot and MASCOT
4.8. Relative Protein Abundance Estimations
4.9. Calculating Molecular Weights from Existing H. curtus Sequences
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 3FTx | Three-Finger Toxin |
| ACN | Acetonitrile |
| CRISP | Cysteine-Rich Secretory Protein |
| CTL | C-Type Lectin |
| Cys | Cystatin |
| DTT | 1,4-Dithiothreitol |
| ECM1 | Extra-Cellular Matrix Protein |
| FA | Formic Acid |
| FDR | False Discovery Rate |
| Hsp70 | Heat Shock Protein |
| LC-ESI-MS | Liquid Chromatography Electrospray Ionisation Mass Spectrometry |
| MALDI-TOF | Matrix-Assisted Laser Desorption Ionisation-Time of Flight |
| MQ | Milli-Q |
| nAChR | Nicotinic Acetylcholine Receptor |
| Nucleo | Nucleobindin |
| PC1A | Peptidase C1A |
| PEBP | Phosphatidylethanolamine-Binding Protein |
| PLA2 | Phospholipase A2 |
| RNase T2 | Ribonuclease T2 |
| RP-HPLC | Reversed-Phase High-Performance Liquid Chromatography |
| SVMP | Snake Venom Metalloproteinase |
| SVSP | Snake Venom Serine Protease |
| TFA | Trifluoroacetic Acid |
| Trans | Transferrin |
References
- Sanders, K.L.; Lee, M.S.Y.; Mumpuni; Bertozzi, T.; Rasmussen, A.R. Multilocus phylogeny and recent rapid radiation of the viviparous sea snakes (Elapidae: Hydrophiinae). Mol. Phylogenet. Evol. 2013, 66, 575–591. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A.R.; Murphy, J.C.; Ompi, M.; Gibbons, J.W.; Uetz, P. Marine reptiles. PLoS ONE 2011, 6, e27373. [Google Scholar] [CrossRef] [PubMed]
- White, J. Clinical toxicology of sea snakebites. In Handbook of Clinical Toxicology of Animal Venoms and Poisons; Meier, J., White, J., Eds.; CRC Press: Boca Raton, FL, USA, 1995; pp. 159–170. [Google Scholar]
- Voris, H.K.; Voris, H.H. Feeding strategies in marine snakes: An analysis of evolutionary, morphological, behavioral and ecological relationships. Am. Zool. 1983, 23, 411–425. [Google Scholar] [CrossRef]
- Fry, B.G.; Wüster, W.; Ryan Ramjan, S.F.; Jackson, T.; Martelli, P.; Kini, R.M. Analysis of Colubroidea snake venoms by liquid chromatography with mass spectrometry: Evolutionary and toxinological implications. Rapid Commun. Mass Spectrom. 2003, 17, 2047–2062. [Google Scholar] [CrossRef] [PubMed]
- Laustsen, A.H.; Gutiérrez, J.M.; Rasmussen, A.R.; Engmark, M.; Gravlund, P.; Sanders, K.L.; Lohse, B.; Lomonte, B. Danger in the reef: Proteome, toxicity, and neutralization of the venom of the olive sea snake, Aipysurus laevis. Toxicon 2015, 107, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvete, J.J.; Ghezellou, P.; Paiva, O.; Matainaho, T.; Ghassempour, A.; Goudarzi, H.; Kraus, F.; Sanz, L.; Williams, D.J. Snake venomics of two poorly known Hydrophiinae: Comparative proteomics of the venoms of terrestrial Toxicocalamus longissimus and marine Hydrophis cyanocinctus. J. Proteom. 2012, 75, 4091–4101. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Pla, D.; Sasa, M.; Tsai, W.C.; Solórzano, A.; Ureña-Díaz, J.M.; Fernández-Montes, M.L.; Mora-Obando, D.; Sanz, L.; Gutiérrez, J.M.; et al. Two color morphs of the pelagic yellow-bellied sea snake, Pelamis platura, from different locations of Costa Rica: Snake venomics, toxicity, and neutralization by antivenom. J. Proteom. 2014, 103, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.H.; Tan, K.Y.; Lim, S.E.; Tan, N.H. Venomics of the beaked sea snake, Hydrophis schistosus: A minimalist toxin arsenal and its cross-neutralization by heterologous antivenoms. J. Proteom. 2015, 126, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Dunson, W.A. Adaptations of sea snakes. In Biology of Sea Snakes; Dunson, W.A., Ed.; University Park Press: Baltimore, MD, USA, 1975; pp. 3–19. [Google Scholar]
- Reid, H.A. Myoglobinuria and sea-snake-bite poisoning. Br. Med. J. 1961, 1, 1284–1289. [Google Scholar] [CrossRef] [PubMed]
- Reid, H.A. Epidemiology and clinical aspects of sea snake bites. In Biology of Sea Snakes; Dunson, W.A., Ed.; University Park Press: Baltimore, MD, USA, 1975; pp. 417–462. [Google Scholar]
- Tu, A.T. Sea snake venoms and neurotoxins. J. Agric. Food Chem. 1974, 22, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Ukuwela, K.D.B.; Silva, A.; Mumpuni; Fry, B.G.; Sanders, K.L. Multilocus phylogeography of the sea snake Hydrophis curtus reveals historical vicariance and cryptic lineage diversity. Zool. Scr. 2014, 43, 472–484. [Google Scholar] [CrossRef]
- Warrell, D.A. Sea snake bites in the Asia-Pacific region. In Sea Snake Toxinology; Gopalakrishnakone, P., Ed.; Singapore University Press: Singapore, 1994; pp. 1–37. [Google Scholar]
- Vijayaraghavan, B.; Ganesh, S.R. Venomous snakes and snakebites in India. In Clinical Toxinology in Asia Pacific and Africa; Faiz, A., Fernando, R., Gnanathasan, C.A., Habib, A.G., Yang, C.-C., Eds.; Springer Science + Business Media: Dordrecht, The Netherlands, 2015. [Google Scholar]
- Cogger, H.G. Reptiles and Amphibians of Australia, 7th ed.; CSIRO Publishing: Collingwood, Australia, 2014. [Google Scholar]
- Heatwole, H. Attacks by sea snakes on divers. In Biology of Sea Snakes; Dunson, W.A., Ed.; University Park Press: Baltimore, MD, USA, 1975; pp. 503–516. [Google Scholar]
- Tan, T.; Xiang, X.; Qu, H.; Zhu, S.; Bi, Q. The study on venom proteins of Lapemis hardwickii by cDNA phage display. Toxicol. Lett. 2011, 206, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.L.; Wei, J.W.; Zhong, X.F.; Zhao, G.J.; Peng, L.S.; Wu, W.Y.; Xu, A.L. Diversity of PLA2 genes from sea snake Lapemis hardwickii Gray venom. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao (Shanghai) 2001, 33, 345–350. [Google Scholar] [PubMed]
- Fox, J.; Tu, A.T. Conformational analysis of a snake neurotoxin by prediction from sequence, circular dichroism, and Raman spectroscopy. Arch. Biochem. Biophys. 1979, 193, 407–414. [Google Scholar] [CrossRef]
- Zhong, X.F.; Peng, L.S.; Wu, W.Y.; Wei, J.W.; Yang, H.; Yang, Y.Z.; Xu, A.L. Identification and functional characterization of three postsynaptic short-chain neurotoxins from Hydrophiinae, Lapemis hardwickii Gray. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao 2001, 33, 457–462. [Google Scholar] [PubMed]
- Wei, J.W.; Zhong, X.F.; Peng, L.S.; Wu, W.Y.; Xu, A.L. Cloning and sequence analysis of full-length cDNA encoding cysteine-rich venom protein from Lapemis hardwickii. Acta Sci. Nat. Univ. Sunyatseni 2001, 40, 5–8. [Google Scholar]
- He, J.; Chen, S.; Gu, J. Identification and characterization of Harobin, a novel fibrino(geno)lytic serine protease from a sea snake (Lapemis hardwickii). FEBS Lett. 2007, 581, 2965–2973. [Google Scholar] [CrossRef] [PubMed]
- Chetty, N.; Du, A.; Hodgson, W.C.; Winkel, K.; Fry, B.G. The in vitro neuromuscular activity of Indo-Pacific sea-snake venoms: Efficacy of two commercially available antivenoms. Toxicon 2004, 44, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, R.; Karthigayan, S.; Balasubashini, M.S.; Vijayalakshmi, S.; Balasubramanian, T. Histopathological changes induced in mice after intramuscular and intraperitoneal injections of venom from spine-bellied sea snake, Lapemis curtus (Shaw, 1802). J. Pharmacol. Toxicol. 2007, 2, 307–318. [Google Scholar]
- Tan, N.-H.; Ponnudurai, G. A comparative study of the biological properties of some sea snake venoms. Comp. Biochem. Physiol. 1991, 99, 351–354. [Google Scholar] [CrossRef]
- Pahari, S.; Bickford, D.; Fry, B.G.; Kini, R.M. Expression pattern of three-finger toxin and phospholipase A2 genes in the venom glands of two sea snakes, Lapemis curtus and Acalyptophis peronii: Comparison of evolution of these toxins in land snakes, sea kraits and sea snakes. BMC Evol. Biol. 2007, 7, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Calvete, J.J. Proteomic tools against the neglected pathology of snake bite envenoming. Expert Rev. Proteom. 2011, 8, 739–758. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P.; Williams, V.; White, J. Snake venom variability: Methods of study, results and interpretation. Toxicon 1991, 29, 1279–1303. [Google Scholar] [CrossRef]
- Alape-Girón, A.; Sanz, L.; Escolano, J.; Flores-Díaz, M.; Madrigal, M.; Sasa, M.; Calvete, J.J. Snake venomics of the lancehead pitviper Bothrops asper: Geographic, individual, and ontogenetic variations. J. Proteome Res. 2008, 7, 3556–3571. [Google Scholar] [CrossRef] [PubMed]
- Furtado, M.F.D.; Maruyama, M.; Kamiguti, A.S.; Antonio, L.C. Comparative study of nine Bothrops snake venoms from adult female snakes and their offspring. Toxicon 1991, 29, 219–226. [Google Scholar] [CrossRef]
- Mackessy, S.P. Venom ontogeny in the Pacific rattlesnakes Crotalus viridis helleri and C. v. oreganus. Copeia 1988, 1988, 92–101. [Google Scholar] [CrossRef]
- Zelanis, A.; Tashima, A.K.; Pinto, A.F.M.; Paes Leme, A.F.; Stuginski, D.R.; Furtado, M.F.; Sherman, N.E.; Ho, P.L.; Fox, J.W.; Serrano, S.M.T. Bothrops jararaca venom proteome rearrangement upon neonate to adult transition. Proteomics 2011, 11, 4218–4228. [Google Scholar] [CrossRef] [PubMed]
- Lomonte, B.; Fernandez, J.; Sanz, L.; Angulo, Y.; Sasa, M.; Gutiérrez, J.M.; Calvete, J.J. Venomous snakes of Costa Rica: Biological and medical implications of their venom proteomic profiles analyzed through the strategy of snake venomics. J. Proteom. 2014, 105, 323–339. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Guidelines for the Production, Control and Regulation of Snake Antivenom Immunoglobulins; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Menezes, M.C.; Furtado, M.F.; Travaglia-Cardoso, S.R.; Camargo, A.C.M.; Serrano, S.M.T. Sex-based individual variation of snake venom proteome among eighteen Bothrops jararaca siblings. Toxicon 2006, 47, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, D.C.; Prezoto, B.C.; Konno, K.; Melo, R.L.; Furtado, M.F.; Camargo, A.C.M.; Serrano, S.M.T. Mass spectrometric analysis of the individual variability of Bothrops jararaca venom peptide fraction. Evidence for sex-based variation among the bradykinin-potentiating peptides. Rapid Commun. Mass Spectrom. 2007, 21, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Ishihama, Y.; Oda, Y.; Tabata, T.; Sato, T.; Nagasu, T.; Rappsilber, J.; Mann, M. Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell. Proteom. 2005, 4, 1265–1272. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J.; Alewood, P.F. Modern venom profiling: Mining into scorpion venom biodiversity. In Scorpion Venoms; Possani, L.D., Schwartz, E.F., Rodríguez de la Vega, R.C., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 547–561. [Google Scholar]
- Barber, C.M.; Isbister, G.K.; Hodgson, W.C. Alpha neurotoxins. Toxicon 2013, 66, 47–58. [Google Scholar] [CrossRef] [PubMed]
- White, J. A Clinician’s Guide to Australian Venomous Bites and Stings: Incorporating the Updated CSL Antivenom Handbook; bioCSL: Parkville, Australia, 2013. [Google Scholar]
- Harris, J.B. Snake venoms in science and clinical medicine 3. Neuropharmacological aspects of the activity of snake venoms. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 745–747. [Google Scholar] [CrossRef]
- Brook, G.A.; Torres, L.F.; Gopalakrishnakone, P.; Duchen, L.W. Effects of phospholipase of Enhydrina schistosa venom on nerve, motor end-plate and muscle of the mouse. Q. J. Exp. Physiol. 1987, 72, 571–591. [Google Scholar] [CrossRef] [PubMed]
- Geh, S.L.; Toh, H.T. Ultrastructural changes in skeletal muscle caused by a phospholipase A2 fraction isolated from the venom of a sea snake, Enhydrina schistosa. Toxicon 1978, 16, 633–643. [Google Scholar] [CrossRef]
- Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Cellular pathology induced by snake venom phospholipase A2 myotoxins and neurotoxins: Common aspects of their mechanisms of action. Cell. Mol. Life Sci. 2008, 65, 2897–2912. [Google Scholar] [CrossRef] [PubMed]
- Verheij, H.M.; Boffa, M.C.; Rothen, C.; Bryckaert, M.C.; Verger, R.; de Haas, G.H. Correlation of enzymatic activity and anticoagulant properties of phospholipase A2. Eur. J. Biochem. 1980, 112, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Gutiérrez, J.M.; Angulo, Y.; Sanz, L.; Juárez, P.; Calvete, J.J.; Lomonte, B. Isolation of an acidic phospholipase A2 from the venom of the snake Bothrops asper of Costa Rica: Biochemical and toxicological characterization. Biochimie 2010, 92, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Jackson, T.; Koludarov, I.; Ali, S.; Dobson, J.; Zdenek, C.; Dashevsky, D.; op den Brouw, B.; Masci, P.; Nouwens, A.; Josh, P.; et al. Rapid radiations and the race to redundancy: An investigation of the evolution of Australian elapid snake venoms. Toxins 2016, 8, 309. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Gao, J.-F.; Lin, L.; Ma, X.; Ji, X. Age-related variation in snake venom: Evidence from two snakes (Naja atra and Deinagkistrodon acutus) in Southeastern China. Asian Herpetol. Res. 2014, 5, 119–127. [Google Scholar]
- Modahl, C.M.; Mukherjee, A.K.; Mackessy, S.P. An analysis of venom ontogeny and prey-specific toxicity in the monocled cobra (Naja kaouthia). Toxicon 2016, 119, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Petras, D.; Sanz, L.; Segura, A.; Herrera, M.; Villalta, M.; Solano, D.; Vargas, M.; León, G.; Warrell, D.A.; Theakston, R.D.G.; et al. Snake venomics of African spitting cobras: Toxin composition and assessment of congeneric cross-reactivity of the pan-African EchiTAb-Plus-ICP antivenom by antivenomics and neutralization approaches. J. Proteome Res. 2011, 10, 1266–1280. [Google Scholar] [CrossRef] [PubMed]
- Herrera, M.; Fernández, J.; Vargas, M.; Villalta, M.; Segura, Á.; León, G.; Angulo, Y.; Paiva, O.; Matainaho, T.; Jensen, S.D.; et al. Comparative proteomic analysis of the venom of the taipan snake, Oxyuranus scutellatus, from Papua New Guinea and Australia: Role of neurotoxic and procoagulant effects in venom toxicity. J. Proteom. 2012, 75, 2128–2140. [Google Scholar] [CrossRef] [PubMed]
- NCBI Non-Redundant Serpentes Database. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/protein/?term=serpentes (accessed on 9 March 2017).
- UniProtKB Serpentes Database. Available online: http://www.uniprot.org/uniprot/?query=Serpentes&sort=score (accessed on 9 March 2017).
- UniProtKB Database. search term ‘Lapemis hardwickii’. Available online: http://www.uniprot.org (accessed on 3 November 2017).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Peak Area % | June MW (Da) | January MW (Da) | Peptides (95%) | Score | Cov % (95%) | Protein Family | Related protein (exPASy MW) | Species (Accession) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 10.1 | 6,688.5 6,745.3 6728.2 | 6,746.8 6,688.4 6,800.8 | 43 | 16.0 | 58.0 | 3FTx | Short neurotoxin 1 (6689.5) | Hydrophis hardwickii (P68416) |
| 2 | 0.05 | 6,688.1 6,747.1 6,727.9 6,720.2 7,732.8 6,811.8 | 6,747.7 6,688.4 6,800.8 6,353.0 6,720.8 6,811.4 7,734.8 | 16 | 25.6 | 33.6 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) |
| 8 | 13.5 | 52.2 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 13 | 12.0 | 58.0 | 3FTx | Short neurotoxin 1 (6689.5) | Hydrophis hardwickii (P68416) | ||||
| 6 | 10.2 | 58.1 | 3FTx | Long neurotoxin 2 | Hydrophis hardwickii (A3FM53) | ||||
| 16 | 9.1 | 28.6 | CRISP | Cysteine-rich venom protein pseudechetoxin-like | Notechis scutatus scutatus (Q3SB04) | ||||
| 17 | 2.0 | 29.4 | CRISP | Cysteine-rich secretory protein | Cryptophis nigrescens (B5G6G3) | ||||
| 3 | 22.4 | 7,632.4 7,574.4 7,556.7 7,614.7 | 7,632.7 7,574.6 7,686.5 7,614.3 7,556.1 | 48 | 54.7 | 79.4 | 3FTx | Alpha-elapitoxin-Lh2a (7633.7) | Hydrophis hardwickii (Q8UW28) |
| 13 | 16.0 | 57.6 | 3FTx | Long neurotoxin 1 | Hydrophis hardwickii (Q8UW29) | ||||
| 4 | 6.4 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 9 | 6.0 | 58.1 | 3FTx | Long neurotoxin 2 | Hydrophis hardwickii (A3FM53) | ||||
| 4 | 4.2 | 13.5 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 2 | 3.1 | 7.5 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 8 | 2.2 | 20.3 | 3FTx | Toxin Lc a | Laticauda colubrina (P0C8R7) | ||||
| 4 | 4.5 | 13,252.0 13,307.7 13,325.1 7,632.3 7,574.6 15,670.2 7,733.5 | 13,336.1 13,308.8 7,632.6 13,328.7 7,574.1 13,251.7 13,389.7 7,614.6 7,686.5 15,726.2 15,672.7 | 113 | 93.9 | 77.4 | PLA2 | Basic phospholipase A2 73 (13,310.0) | Hydrophis hardwickii (Q8UW30) |
| 42 | 31.2 | 79.4 | 3FTx | α-elapitoxin-Lh2a (7633.7) | Hydrophis hardwickii (Q8UW28) | ||||
| 17 | 18.8 | 62.0 | 3FTx | Long neurotoxin 1 | Hydrophis hardwickii (Q8UW29) | ||||
| 61 | 12.2 | 74.7 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 9 | 11.7 | 29.4 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 4 | 7.9 | 58.0 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 14 | 4.2 | 29.0 | PLA2 | Acidic phospholipase A2 57 | Hydrophis hardwickii (Q8UW31) | ||||
| 9 | 4.0 | 59.1 | 3FTx | Long neurotoxin 2 | Hydrophis hardwickii (A3FM53) | ||||
| 6 | 2.0 | 28.3 | PLA2 | PLA-2-Den-2 | Denisonia devisi (R4G2S8) | ||||
| 4–5 | – | – | – | 154 | 86.9 | 76.7 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) |
| 117 | 21.4 | 75.3 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 29 | 18.7 | 69.6 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 15 | 12.3 | 62.0 | 3FTx | Long neurotoxin 1 | Hydrophis hardwickii (Q8UW29) | ||||
| 4 | 7.4 | 58.0 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 29 | 5.3 | 43.4 | PLA2 | Acidic phospholipase A2 57 | Hydrophis hardwickii (Q8UW31) | ||||
| 7 | 3.2 | 12.6 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 29 | 2.2 | 26.7 | PLA2 | Acidic phospholipase A2 S5-32M | Austrelaps superbus (Q9PUH4) | ||||
| 3 | 2.1 | 4.2 | SVMP | Nigrescease-1 | Cryptophis nigrescens (B5KFV8) | ||||
| 8 | 2.0 | 17.7 | PLA2 | Putative phospholipase A2 | Austrelaps labialis (B2BRS9) | ||||
| 4 | 2.0 | 20.3 | 3FTx | Toxin Lc b | Laticauda colubrina (P0C8R8) | ||||
| 5 | 3.3 | 13,344.9 13,401.7 7,632.5 7,574.4 13,269.8 13,364.8 13,326.8 | 13,344.7 13,401.4 7,632.4 7,574.1 | 116 | 122.7 | 76.0 | PLA2 | Basic phospholipase A2 (13,346.0) | Hydrophis hardwickii (Q8UW08) |
| 25 | 26.7 | 69.6 | 3FTx | α-elapitoxin-Lh2a (7633.7) | Hydrophis hardwickii (Q8UW28) | ||||
| 20 | 26.0 | 60.5 | PLA2 | Acidic phospholipase A2 57 | Hydrophis hardwickii (Q8UW31) | ||||
| 73 | 20.3 | 67.1 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 8 | 16.0 | 22.7 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 9 | 12.1 | 15.2 | SVMP | Zinc metalloproteinase-disintegrin-like MTP9 | Drysdalia coronoides (F8RKV9) | ||||
| 4 | 8.0 | 57.6 | 3FTx | Long neurotoxin 1 | Hydrophis hardwickii (Q8UW29) | ||||
| 3 | 6.0 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 16 | 4.0 | 34.8 | PLA2 | Basic phospholipase A2 3 | Laticauda semifasciata (P00612) | ||||
| 2 | 2.0 | 20.3 | 3FTx | Toxin Lc a | Laticauda colubrina (P0C8R7) | ||||
| * | 2.5 | – | 14,157.2 7,632.1 13,401.6 13,345.2 7,574.6 | 144 | 136.6 | 77.6 | PLA2 | Acidic phospholipase A2 57 | Hydrophis hardwickii (Q8UW31) |
| 56 | 55.9 | 72.6 | PLA2 | Basic phospholipase A2 (13,346.0) | Hydrophis hardwickii (Q8UW08) | ||||
| 19 | 18.6 | 64.1 | 3FTx | α-elapitoxin-Lh2a (7633.7) | Hydrophis hardwickii (Q8UW28) | ||||
| 46 | 15.5 | 71.9 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 3 | 6.0 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 13 | 6.0 | 20.4 | PLA2 | PLA2-Bra-11 | Brachyurophis roperi (R4G2E0) | ||||
| 2 | 4.1 | 7.6 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 2 | 2.7 | 20.3 | 3FTx | Toxin Lc a | Laticauda colubrina (P0C8R7) | ||||
| 8 | 2.1 | 29.7 | PLA2 | Pa-18 | Pseudechis australis (Q45Z21) | ||||
| 9 | 2.0 | 17.1 | PLA2 | Acidic phospholipase A2 2 | Ophiophagus hannah (Q9DF33) | ||||
| 6a, 6b, 6c | 47.9 | 14,215.4 14,314.8 7,632.6 14,269.3 7,574.8 13,113.8 13,402.4 13,170.5 7,685.9 13,325.3 7,615.0 | 14,215.5 14,314.6 7,632.4 7,574.3 | 116 | 115.2 | 73.7 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) |
| 26 | 20.1 | 56.2 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 8 | 14.0 | 64.1 | 3FTx | α-elapitoxin-Lh2a (7633.7) | Hydrophis hardwickii (Q8UW28) | ||||
| 22 | 10.0 | 65.8 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 4 | 6.8 | 58.0 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 2 | 4.0 | 7.6 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 12 | 2.1 | 47.7 | PLA2 | PLA-2 | Notechis scutatus (Q45Z32) | ||||
| 5 | 1.5 | 19.0 | PLA2 | Basic phospholipase A2 2 (Fragment) | Bungarus caeruleus (Q6SLM1) | ||||
| 7 | 2.7 | 14,215.4 14,269.6 14,295.5 14,315.1 13,946.5 | 14,215.4 14,295.5 14,269.9 13,514.1 14,315.2 13,457.4 | 157 | 130.3 | 82.2 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) |
| 17 | 10.3 | 56.9 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 6 | 6.8 | 34.8 | 3FTx | α elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 19 | 6.0 | 55.5 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 3 | 5.6 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 16 | 4.0 | 45.5 | PLA2 | Putative phospholipase A2 | Austrelaps labialis (B2BRS7) | ||||
| 9 | 4.0 | 45.2 | PLA2 | Acidic phospholipase A2 2 | Ophiophagus hannah (Q9DF33) | ||||
| 4 | 3.2 | 12.2 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 12 | 2.0 | 46.7 | PLA2 | PLA2 Hs-1 | Hoplocephalus stephensii (A6MJG3) | ||||
| 8a, 8b | 0.23 | 14,328.8 14,428.3 14,214.9 14,381.2 14,267.9 | 14,328.2 14,216.0 14,052.6 14,382.3 14,269.9 | 76 | 87.6 | 78.3 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) |
| 16 | 16.4 | 56.9 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 12 | 12.9 | 62.0 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 4 | 6.6 | 16.8 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 3 | 6.4 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 18 | 6.0 | 55.5 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 6 | 4.0 | 31.8 | PLA2 | PLA-2 | Notechis scutatus (Q45Z32) | ||||
| 9a | 0.7 | 15,045.6 15,144.9 14,216.4 14,975.0 15,102.3 14,847.0 14,934.0 15,003.3 15,074.1 | 15,046.2 15,005.0 15,144.2 15,101.5 14,846.2 14,975.0 14,886.8 15,370.2 | 64 | 65.6 | 80.9 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) |
| 17 | 12.6 | 73.3 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 8 | 7.8 | 52.2 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 18 | 6.0 | 61.0 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 3 | 5.6 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 7 | 3.1 | 27.3 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 9b | 0.5 | 15,044.9 15,004.8 14,845.6 14,934.0 14,215.6 20,615.9 | 15,045.6 15,004.0 14,846.8 14,214.6 14,933.4 15,099.5 14,973.8 15,145.5 15,073.4 14,886.3 20,614.6 | 103 | 99.3 | 88.2 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) |
| 20 | 16.5 | 47.1 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 16 | 13.6 | 59.0 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 6 | 6.9 | 24.3 | PEBP | Phosphatidylethanolamine-binding protein 4 | Micrurus fulvius (U3FZ77) | ||||
| 17 | 5.6 | 61.0 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 4 | 5.4 | 58.0 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 5 | 5.0 | 51.1 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 2 | 1.7 | 3.8 | Hsp70 | 78 kDa glucose-regulated protein | Ophiophagus hannah (V8NEC1) | ||||
| 16 | 1.4 | 37.0 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) | ||||
| 10 | 7.5 | 24,321.2 24,420.2 24,223.2 24,521.0 24,058.7 14,215.2 | 24,320.5 24,419.6 24,520.7 24,222.4 24,058.3 14,215.2 | 86 | 77.4 | 75.2 | CRISP | Cysteine-rich venom protein 2 (24,522.8) | Hydrophis hardwickii (Q8UW11) |
| 12 | 9.4 | 68.4 | PLA2 | Acidic phospholipase A2 57 (14,217.0) | Hydrophis hardwickii (Q8UW31) | ||||
| 6 | 7.6 | 37.7 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 4 | 5.7 | 28.4 | Cys | Cystatin | Hoplocephalus stephensii (E3P6P0) | ||||
| 3 | 5.4 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 50 | 5.3 | 63.0 | CRISP | Cysteine-rich venom protein 1 (24,334.8) | Hydrophis hardwickii (Q8UW25) | ||||
| 23 | 4.9 | 36.1 | CRISP | Cysteine-rich venom protein pseudechetoxin | Pseudechis australis (Q8AVA4) | ||||
| 7 | 4.7 | 47.3 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 6 | 2.7 | 52.2 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 11 | 0.13 | 43,732.5 | 43,730.9 43,414.4 | 32 | 51.2 | 60.1 | CRISP | Cysteine-rich venom protein 2 | Hydrophis hardwickii (Q8UW11) |
| 13 | 26.1 | 67.1 | PLA2 | Acidic phospholipase A2 57 | Hydrophis hardwickii (Q8UW31) | ||||
| 30 | 14.5 | 52.5 | CRISP | Cysteine-rich venom protein 1 | Hydrophis hardwickii (Q8UW25) | ||||
| 9 | 12.6 | 45.2 | PLA2 | Basic phospholipase A2 | Hydrophis hardwickii (Q8UW08) | ||||
| 5 | 10.7 | 5.0 | Golgin | Golgi apparatus protein 1 (Fragment) | Ophiophagus hannah (V8NSK8) | ||||
| 5 | 10.7 | 5.8 | Trans | Transferrin | Boiga irregularis (A0A0B8RPR5) | ||||
| 5 | 7.8 | 51.1 | 3FTx | α-elapitoxin-Lh2a | Hydrophis hardwickii (Q8UW28) | ||||
| 3 | 6.1 | 43.2 | 3FTx | Short neurotoxin 1 | Hydrophis hardwickii (P68416) | ||||
| 10 | 6.0 | 52.7 | PLA2 | Basic phospholipase A2 73 | Hydrophis hardwickii (Q8UW30) | ||||
| 5 | 6.0 | 10.5 | Nucleo | Nucleobindin-2 | Micrurus fulvius (U3FBZ2) | ||||
| 2 | 4.2 | 11.6 | CTL | C-type lectin 1 | Hydrophis hardwickii (A3FM55) | ||||
| 2 | 4.1 | 7.6 | PC1A | Cathepsin B | Micrurus fulvius (U3FD65) | ||||
| 12 | 4.0 | 30.7 | CRISP | Cysteine-rich venom protein pseudechetoxin | Pseudechis australis (Q8AVA4) | ||||
| 3 | 2.2 | 6.0 | Trans | Transferrin | Boaedon fuliginosus (Q1EL74) |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neale, V.; Sotillo, J.; Seymour, J.E.; Wilson, D. The Venom of the Spine-Bellied Sea Snake (Hydrophis curtus): Proteome, Toxin Diversity and Intraspecific Variation. Int. J. Mol. Sci. 2017, 18, 2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122695
Neale V, Sotillo J, Seymour JE, Wilson D. The Venom of the Spine-Bellied Sea Snake (Hydrophis curtus): Proteome, Toxin Diversity and Intraspecific Variation. International Journal of Molecular Sciences. 2017; 18(12):2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122695
Chicago/Turabian StyleNeale, Vanessa, Javier Sotillo, Jamie E. Seymour, and David Wilson. 2017. "The Venom of the Spine-Bellied Sea Snake (Hydrophis curtus): Proteome, Toxin Diversity and Intraspecific Variation" International Journal of Molecular Sciences 18, no. 12: 2695. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms18122695