Nitric Oxide Regulates Seedling Growth and Mitochondrial Responses in Aged Oat Seeds

Abstract
:1. Introduction
2. Results
2.1. Changes in Seed Germination and Seedling Growth in Aged Oat Seeds under Nitric Oxide Treatments
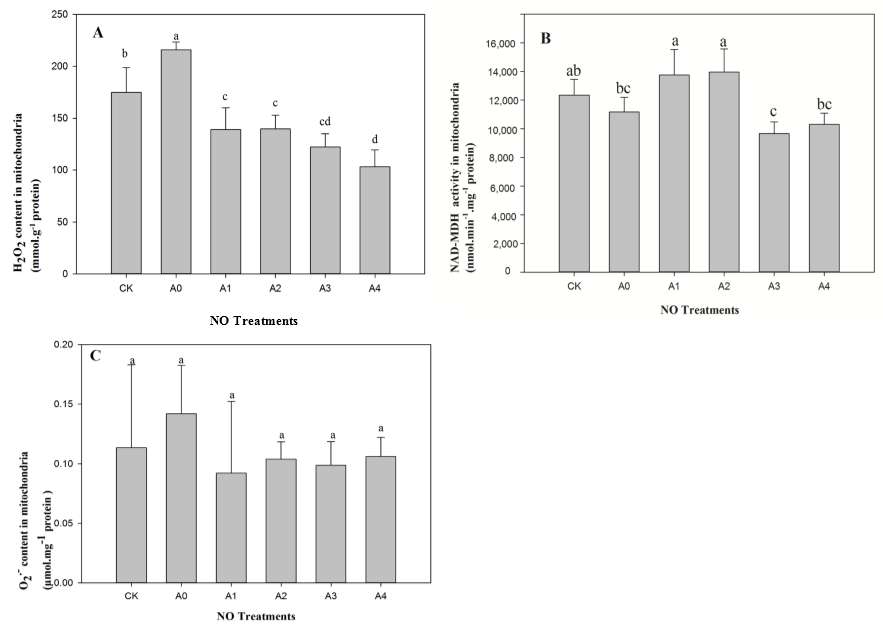
2.2. Changes in Mitochondrial H2O2 Content, Malate Dehydrogenase (NAD-MDH), and O2− Content in Aged Oat Seeds under NO Treatments
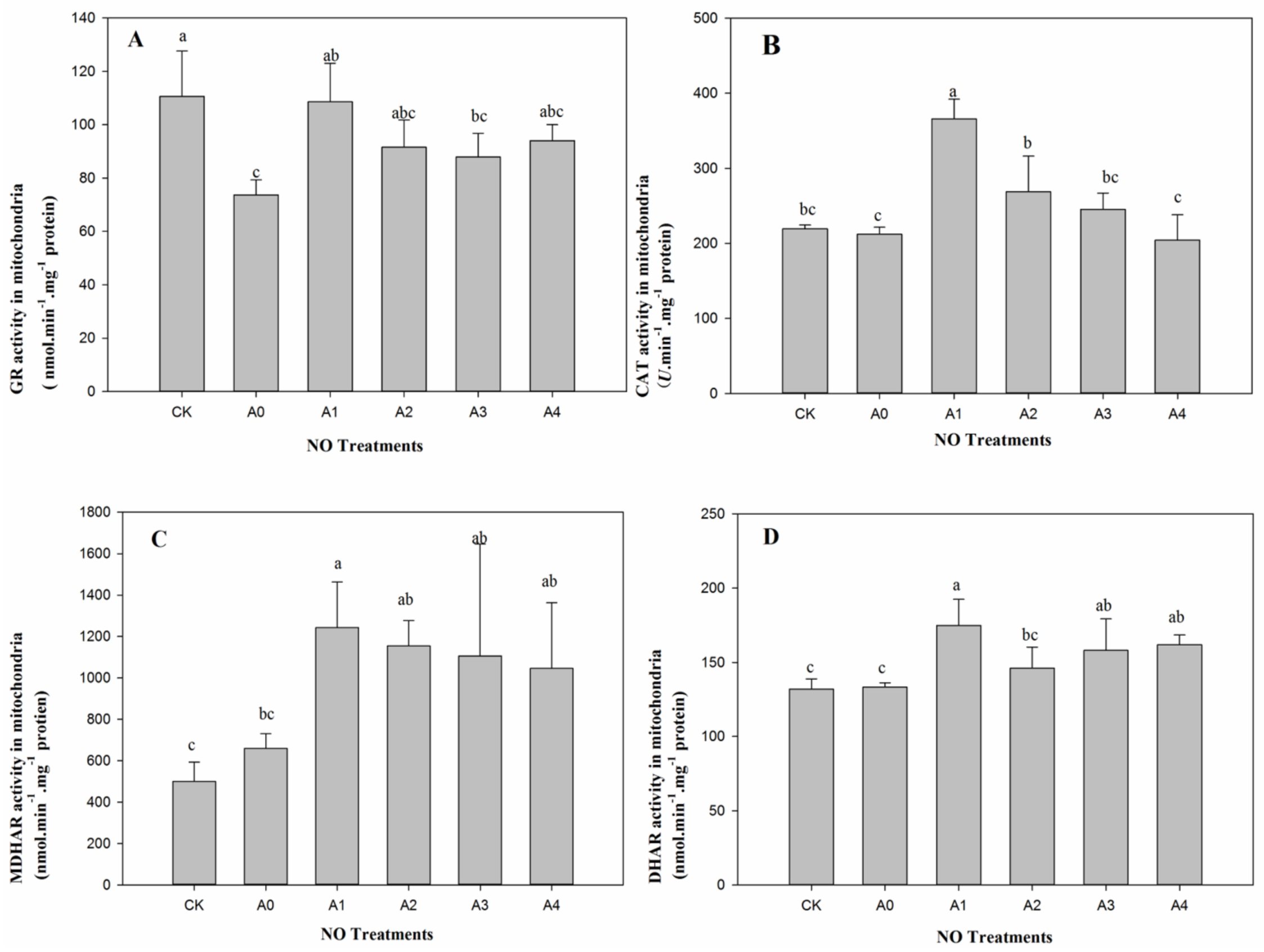
2.3. Changes in Mitochondrial Antioxidant Enzymes in Aged Oat Seeds under Nitric Oxide Treatments
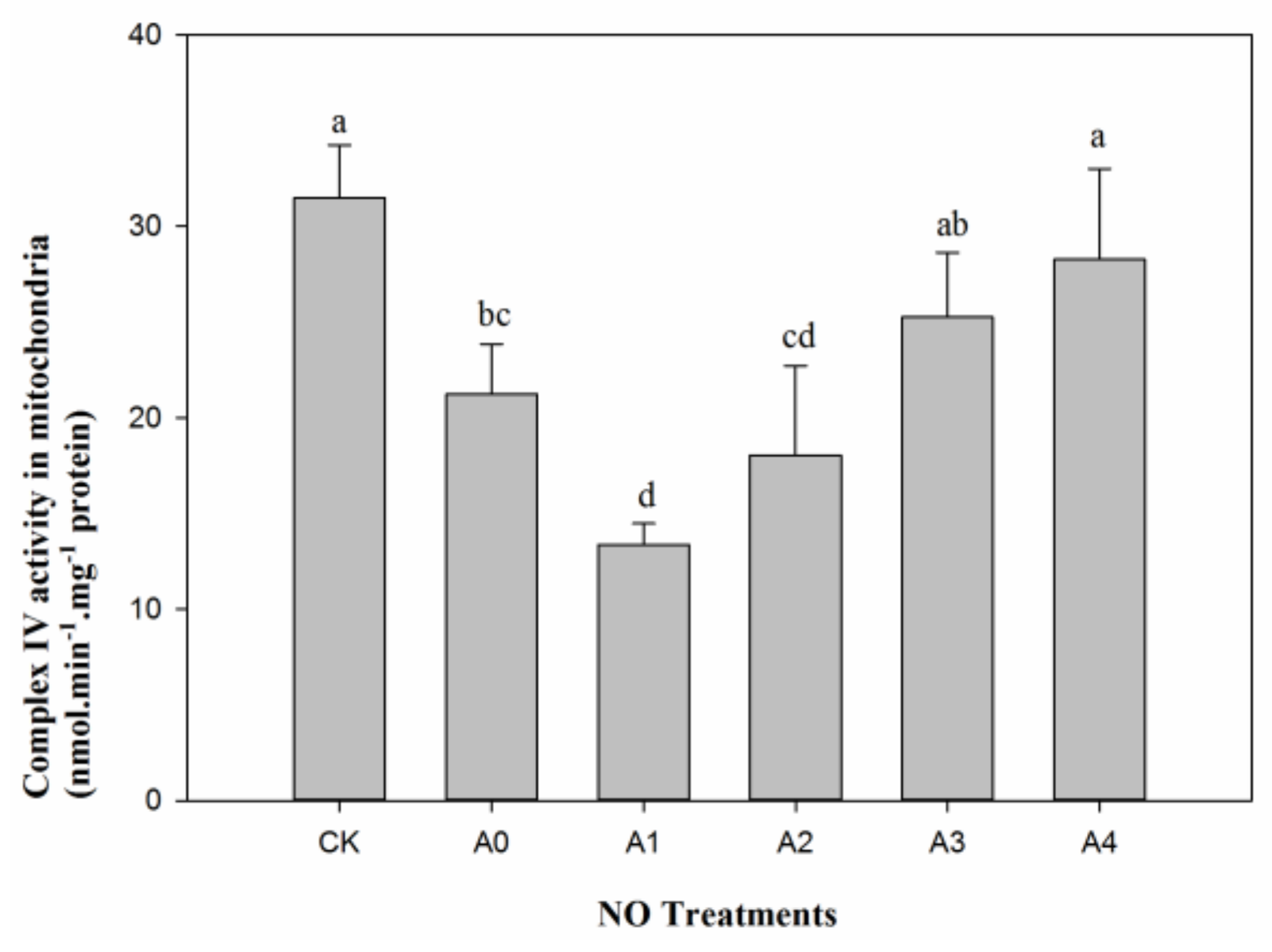
2.4. Changes in Mitochondrial Complex IV in Aged Oat Seeds under Nitric Oxide Treatments
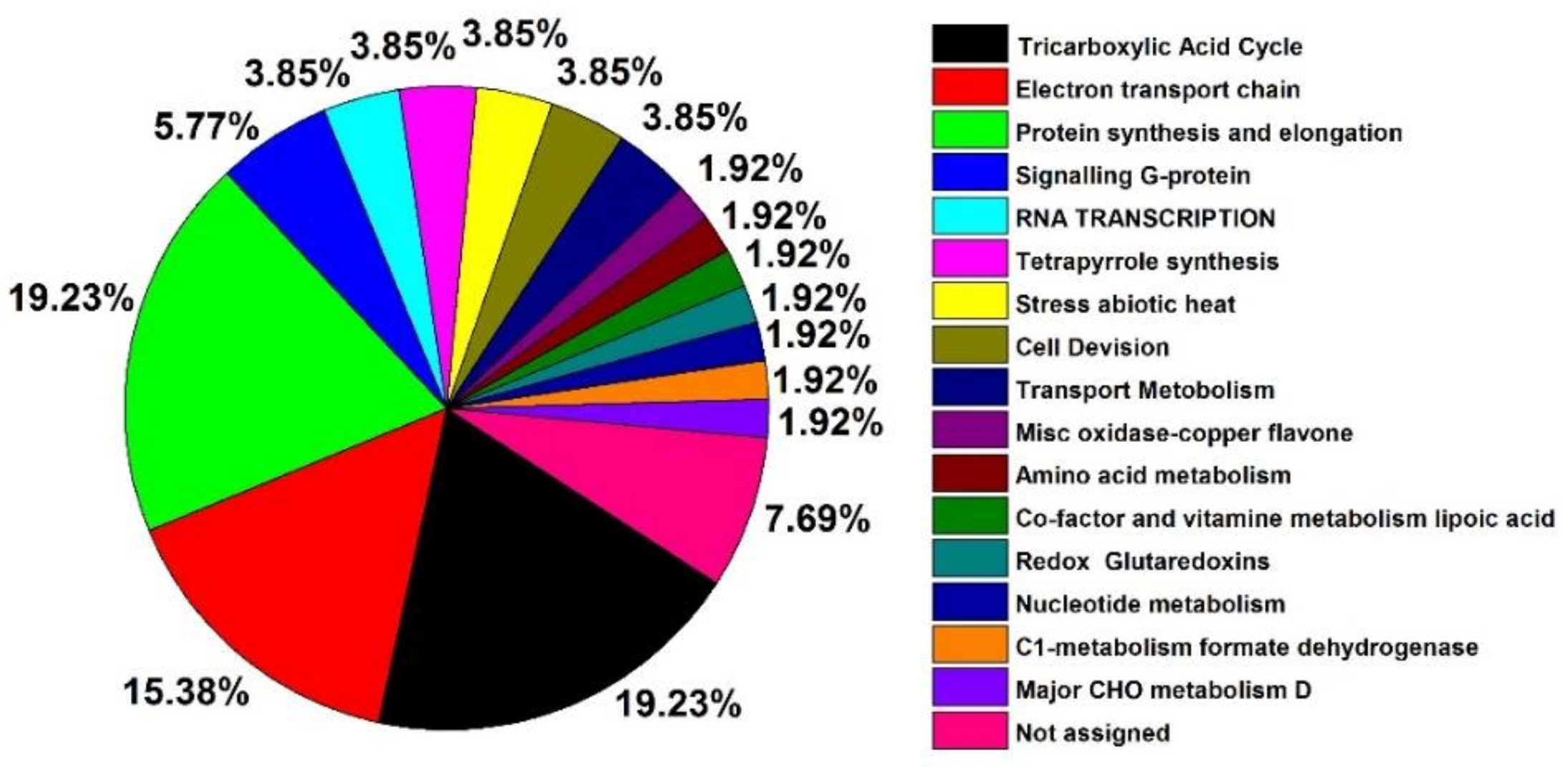
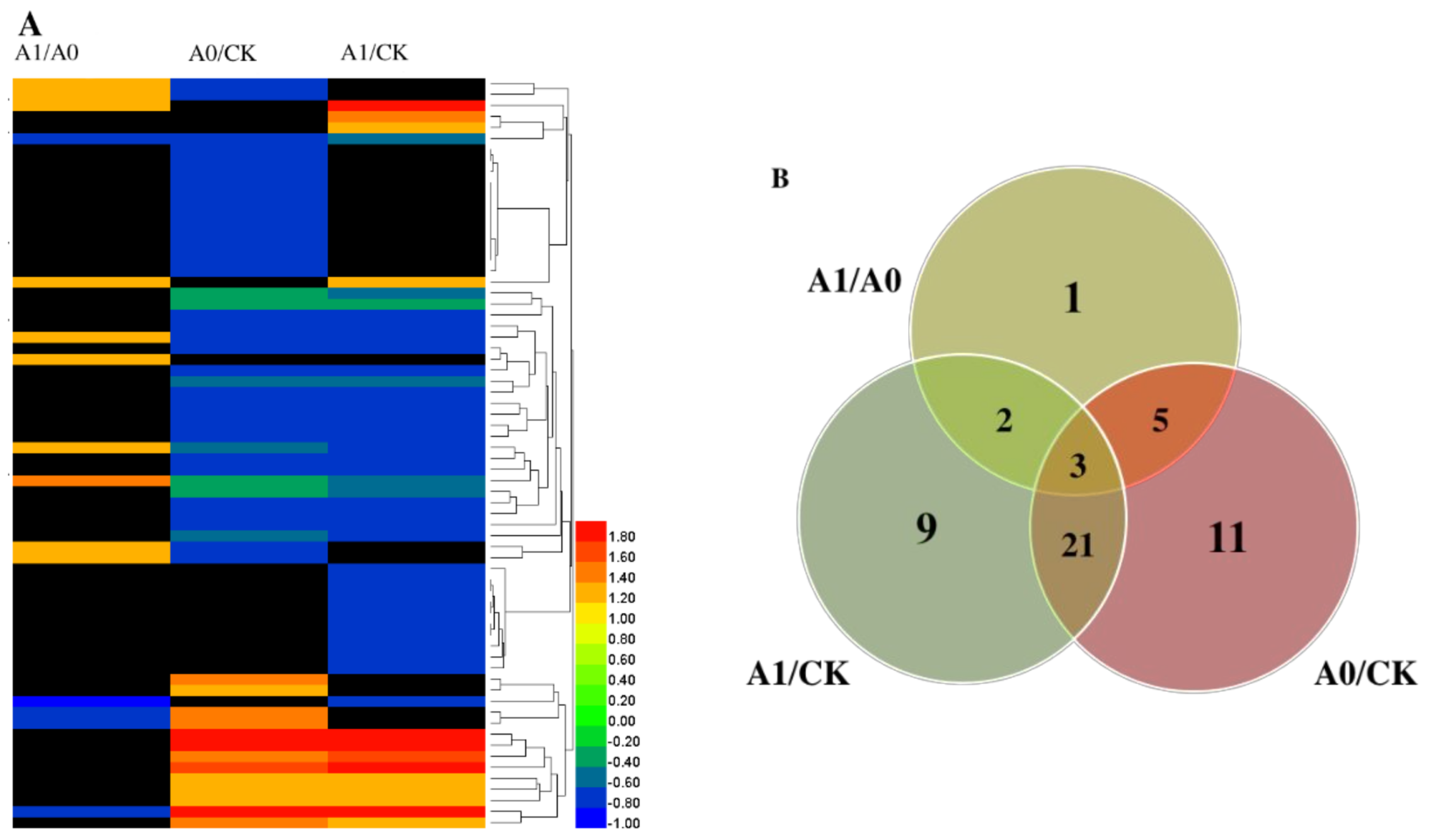
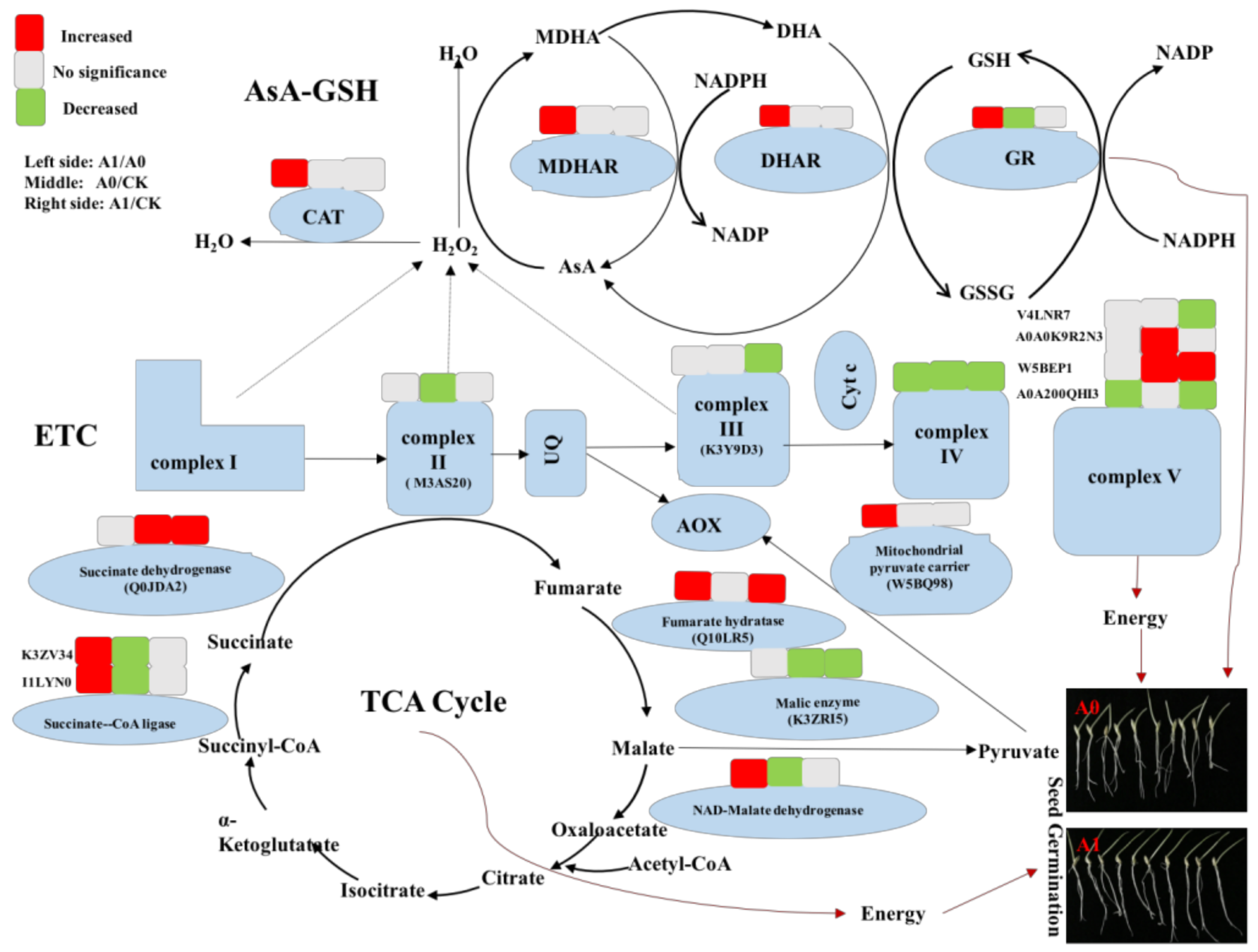
2.5. Changes in Mitochondrial Proteins in Aged Oat Seeds under Nitric Oxide Treatments
3. Discussion
4. Materials and Methods
4.1. Seed Materials
4.2. Determination of Seed Moisture Content
4.3. Adjusting the Seed Moisture Content
4.4. Seed Aging Treatments
4.5. NO Treatment and Germination Test
4.6. Isolation and Purification of Mitochondria
4.7. Determination of H2O2 Content
4.8. Enzyme Assays
4.9. Protein Quantification and Digestion
4.10. Mass Spectrometry Method and Data Analysis
4.10.1. DDA Sample Acquisition
4.10.2. Spectral Library Generation
4.10.3. DIA Sample Acquisition
4.10.4. DIA Data Analysis
4.10.5. Identification of Mitochondrial Proteins
4.11. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CAT | Catalase |
| DHAR | Dehydroascorbate reductase |
| ETC | Electron transport chain |
| GR | Glutathione reductase |
| H2O2 | Hydrogen peroxide |
| MGT | Mean germination time |
| MDHAR | Monodehydroascorbate reductase |
| NAD-MDH | NAD-malate dehydrogenase |
| TCA | Tricarboxylic acid |
References
- EI-Maarouf-Bouteau, H.; Mazuy, C.; Corbineau, F.; Bailly, C. DNA alteration and programmed cell death during ageing of sunflower seed. J. Exp. Bot. 2011, 62, 5003–5011. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.S.; Wang, M.Y.; Li, M.L.; Mao, P.S. Mitochondrial structural and antioxidant system responses to aging in oat (Avena sativa L.) seeds with different moisture contents. Plant Physiol. Biochem. 2015, 94, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.F.; Mao, P.S.; Sun, Y.; Li, M.L. Impacts of ascorbic acid on germination, antioxidant enzymes and ultrastructure of embryo cells of aged elymus sibiricus seeds with different moisture contents. Int. J. Agric. Biol. 2016, 18, 176–183. [Google Scholar] [CrossRef]
- McDonald, M.B. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol. 1999, 27, 177–237. [Google Scholar]
- Baily, C.; El-maarouf-bouteau, H.; Corbineau, F. From intracellular signaling networks to cell death: The dual of reactive oxygen species in seed physiology. CR Biol. 2008, 331, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Kibinza, S.; Vinel, D.; Côme, D.; Bailly, C.; Corbineau, F. Sunflower seed deterioration as related to moisture content during aging, energy metabolism and active oxygen species scavenging. Physiol. Plant. 2006, 128, 496–506. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Xue, H.; Pritchard, H.W.; Wang, X.F. Changes in the mitochondrial protein profile due to ROS eruption during ageing of elm (ulmus pumila L.) seeds. Plant Physiol. Biochem. 2017, 114, 72–87. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M. Plant mitochondria and oxidative stress: Electron transport, NADPH turnover, and metabolism of reactive oxygen species. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 561–591. [Google Scholar] [CrossRef] [PubMed]
- Fleury, C.; Mignotte, B.; Vayssière, J.L. Mitochondrial reactive oxygen species in cell death signaling. Biochimie 2002, 84, 131–141. [Google Scholar] [CrossRef]
- Rhoads, D.M.; Umbach, A.L.; Subbaiah, C.C.; Siedow, J.N. Mitochondrial reactive oxygen species, contribution to oxidative stress and interorganellar signaling. Plant Physiol. 2006, 141, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C. Alternative oxidase: A mitochondrial respiratory pathway to maintain metabolic and signaling homeostasis during abiotic and biotic stress in plants. Int. J. Mol. Sci. 2013, 14, 6805–6847. [Google Scholar] [CrossRef] [PubMed]
- Navrot, N.; Rouhier, N.; Gelhaye, E.; Jacquot, J.P. Reactive pxygen species generation and antioxidant systems in plant mitochondria. Physiol. Plant. 2007, 129, 185–195. [Google Scholar] [CrossRef]
- Affourtit, C.; Krab, K.; Moore, A.L. Control of plant mitochondrial respiration. Biochim. Biophys. Acta 2001, 1504, 58–69. [Google Scholar] [CrossRef]
- Wu, Q.; Feng, H.Q.; Li, H.Y.; Wang, D.S.; Liang, H.G. Effects of drought stress on cyanide-resistance respiration and metabolism of reactive oxygen in wheat seedling. J. Plant Physiol. Mol. Biol. 2006, 32, 217–224. [Google Scholar]
- Fiorani, F.; Umbach, A.L.; Siedow, J.N. The alternative oxidase of plant mitochondria is involved in the acclimation of shoot growth at low temperature. A study of Arabidopsis AOX1a transgenic plants. Plant Physiol. 2005, 139, 1790–1805. [Google Scholar] [CrossRef] [PubMed]
- Bailly, C. Active oxygen species and antioxidants in seed biology. Seed Sci. Res. 2004, 14, 93–107. [Google Scholar] [CrossRef]
- Xin, X.; Tian, Q.; Yin, G.K.; Chen, X.L.; Zhang, J.M.; Sophia, N.G.; Lu, X.X. Reduced mitochondrial and ascorbate-glutathione activity after artificial ageing in soybean seed. J. Plant Physiol. 2014, 177, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environment stress in plants. Front. Environ. Sci. 2014, 2, 1–13. [Google Scholar] [CrossRef]
- Krasuska, U.; Gniazdowska, A. Nitric oxide and hydrogen cyanide as regulating factors of enzymatic antioxidant system in germination apple embryos. Acta Physiol. Plant. 2012, 34, 683–692. [Google Scholar] [CrossRef]
- Zheng, C.F.; Jiang, D.; Liu, F.L.; Dai, T.B.; Liu, W.C.; Jing, Q.; Cao, W.X. Exogenous nitric oxide improves seed germination in wheat against mitochondrial oxidative damage induced by high salinity. Environ. Exp. Bot. 2009, 67, 222–227. [Google Scholar] [CrossRef]
- Lin, Y.; Liu, Z.Z.; Shi, Q.; Wei, M.; Yang, F.J. Exogenous nitric oxide (NO) increased antioxidant capativity of cucumber hypocotyl and radicle under salt stress. Sci. Hortic. 2012, 142, 118–127. [Google Scholar] [CrossRef]
- Hu, K.D.; Hu, L.Y.; Li, Y.H.; Zhang, F.Q.; Zhang, H. Protective roles of nitric oxide on germination and antioxidant metabolism in wheat seeds under copper stress. Plant Growth Regul. 2007, 53, 173–183. [Google Scholar] [CrossRef]
- Millar, A.H.; Day, D.A. Nitric oxide inhibits the cytochrome oxidase but not the alternative oxidase of plant mitochondria. FEBS Lett. 1996, 398, 155–158. [Google Scholar] [CrossRef]
- Royo, B.; Moran, J.F.; Ratcliffe, R.G.; Gupta, K.J. Nitric oxide induces the alternative oxidase pathway in Arabidopsis seedlings deprived of inorganic phosphate. J. Exp. Bot. 2015, 60, 6273–6280. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.K.; Whelan, J.; Wu, S.H.; Zhou, J.; Chen, B.; Chen, X.L.; Zhang, J.M.; He, J.J.; Xin, X.; Lu, X.X. Comprehensive mitochondrial metabolic shift during the critical node of seed ageing in rice. PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Lin, X.H.; Zhou, Y.C.; Chen, X.L.; Liu, X.; Lu, X.X. Proteome analysis of maize seeds: The effect of artificial ageing. Physiol. Plant. 2011, 143, 126–138. [Google Scholar] [CrossRef] [PubMed]
- Belibni, M.V.; Lamattina, L. Nitric oxide stimulates seed germination and de-etiolation and inhibits hypocotyl elongation, three light-inducible responses in plants. Planta 2000, 210, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Mattews, S.; Noli, E.; Demir, I.; Khajeh-Hosseini, M.; Wagner, M.H. Evaluation of seed quality: From physiology to international standardization. Seed Sci. Res. 2012, 22, S69–S73. [Google Scholar] [CrossRef]
- Law, S.R.; Narsai, R.; Whelan, J. Mitochondrial biogenesis in plants during seed germination. Mitochondrion 2014, 19, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Jardim-messeder, D.; Caverzan, A.; Rauber, R.; Ferreira, E.S.; Margis-Pinheiro, M.; Galina, A. Succinate dehydrogenase (mitochondrial complex II) is a source of reactive oxygen species in plants and regulates sevelopment and stress responnses. New Phytol. 2015, 208, 776–789. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.C. Nitric oxide regulates mitochondrial respiration and cell function by inhibiting cytochrome oxidase. FEBS Lett. 1995, 369, 136–139. [Google Scholar] [CrossRef]
- Brown, G.C. Regulation of mitochondrial respiration by nitric oxide inhibition of cytochrome c oxidase. Biochim. Biophys. Acta 2001, 1504, 46–57. [Google Scholar] [CrossRef]
- Johnson, J.D.; Mehus, J.G.; Tews, K.; Milavetz, B.L.; Lambeth, D.O. Genetic evidence for the expression of ATP- and GTP-specific Succinyl-CoA synthetases in multicellular eucaryotes. J. Biochem. 1998, 273, 27580–27586. [Google Scholar] [CrossRef]
- Maurino, V.G.; Gerrard Wheeler, M.C.; Andreo, C.S.; Drincovich, M.F. Redoundancy is sometimes seen only by the uncritical: Does Arabidopsis need six malic enzyme isoform. Plant Sci. 2009, 176, 715–721. [Google Scholar] [CrossRef]
- Millar, A.H.; Wiskich, J.T.; Whelan, J.; Day, D.A. Organic acid activation of the alternative oxidase of plant mitochondria. FEBS Lett. 1993, 329, 259–262. [Google Scholar] [CrossRef]
- Day, A.A.; Millar, A.H.; Wiskich, J.T.; Whelan, J. Regulation of alternative oxidase by pyruvate in soybean mitochondria. Plant Physiol. 1994, 106, 1421–1427. [Google Scholar] [CrossRef] [PubMed]
- Vanlerberghe, G.C.; Day, D.A.; Wiskich, J.T.; Vanlerberghe, A.E.; Mclntosh, L. Alternative oxidase activity in tobacco leaf mitochondria (Dependence on trucarboxylic acid cycle-mediated redox regulation and pyruvate activation). Plant Physiol. 1995, 109, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Oliver, S.N.; Lunn, J.E.; Urbanzcyk-Wochniak, E.; Lytovchenko, A.; Van Dongen, J.T.; Faix, B.; Schmalzlin, E.; Fernie, A.R.; Geigenberger, P. Decreased expression of cytosolic pyruvate kinase in potato tubers lead to a decline in pyruvate resulting in an in vivo repression of the alternative oxidase. Plant Physiol. 2008, 148, 1640–1654. [Google Scholar] [CrossRef] [PubMed]
- Raha, S.; Robinson, B.H. Mitochondria, oxygen free radicals, disease and ageing. Trends Biochem. Sci. 2000, 25, 502–508. [Google Scholar] [CrossRef]
- Maxwell, D.P.; Wang, Y.; Mclotosh, L. The alternative oxidase lowers mitochondrial reactive oxygen production in plant cells. Proc. Natl. Acad. Sci. USA 1999, 96, 8271–8276. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Rad, U.V.; Durner, J. Nitric oxide induces transcriptional activation of the nitric oxide-tolerant alternative oxidase in Arabidopsis suspension cells. Planta 2002, 215, 914–923. [Google Scholar] [CrossRef] [PubMed]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Basserdorf, Awitzerland, 2015. [Google Scholar]
- Ellis, R.A.; Roberts, E.H. The quantification of ageing and survival in orthodox seeds. Seed Sci. Res. 1981, 9, 373–409. [Google Scholar]
- Spivak, M.; Weston, J.; Bottou, L.; Kall, L.; Noble, W.S. Improvements to the percolator algorithm for peptide identification from shotgun proteomics data sets. J. Proteom. Res. 2009, 8, 3737–3745. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Germination Percentage (%) | Mean Germination Time (d) | Root Length (cm) | Shoot Length (cm) |
|---|---|---|---|---|
| CK | 99 ± 1.2 a | 1.9 ± 0.04 d | 6.2 ± 0.30 a | 4.7 ± 0.38 a |
| A0 | 68 ± 5.9 b,c | 3.4 ± 0.14 b | 3.9 ± 0.50 c | 1.9 ± 0.23 c |
| A1 | 78 ± 4.3 b | 3.0 ± 0.04 c | 5.1 ± 0.54 b | 2.4 ± 0.46 b |
| A2 | 75 ± 8.3 b,c | 3.3 ± 0.20 b | 4.1 ± 0.40 c | 2.0 ± 0.27 b,c |
| A3 | 70 ± 8.2 b,c | 3.4 ± 0.29 b | 2.0 ± 0.39 d | 0.7 ± 0.08 d |
| A4 | 65 ± 6.2 c | 3.9 ± 0.19 a | 1.7 ± 0.05 d | 0.6 ± 0.08 d |
| Hit Number | Accession No. | Protein Name (Species) | Fold Change | ||
|---|---|---|---|---|---|
| A1/A0 | A0/CK | A1/CK | |||
| TCA cycle | |||||
| I1LYN0 | AT2G20420 | Succinate-CoA ligase [ADP-forming] subunit beta, mitochondrial Glycine max | 1.25 | 0.67 | ns |
| K3ZV34 | AT5G23250 | Succinate-CoA ligase [ADP-forming] subunit alpha, mitochondrial Setaria italica | 1.33 | 0.76 | ns |
| Q10LR5 | AT5G50950 | Fumarate hydratase 1, mitochondrial, putative, expressed Oryza sativa subsp. japonica | 1.25 | ns | 2.56 |
| R4X771 | AT5G50950 | Fumarate hydratase, mitochondrial Taphrina deformans | ns | 0.40 | 0.49 |
| K3ZRI5 | AT2G13560 | Malic enzyme Setaria italica | ns | 0.70 | 0.71 |
| K3YRJ0 | AT1G79440 | Aldehyde dehydrogenase Setaria italica | ns | 0.77 | 0.73 |
| B4FRH5 | AT2G20420 | Succinate-CoA ligase [ADP-forming] subunit beta, mitochondrial Zea mays | ns | 0.79 | ns |
| W5C4B7 | AT5G23250 | Succinate-CoA ligase [ADP-forming] subunit alpha, mitochondrial Triticum aestivum | ns | 0.67 | ns |
| V4KMJ1 | AT5G66760 | Succinate dehydrogenase [ubiquinone] flavoprotein subunit, mitochondrial Eutrema salsugineum | ns | 1.52 | ns |
| M3AS20 | AT5G40650 | Succinate dehydrogenase [ubiquinone] iron-sulfur subunit, mitochondrial Pseudocercosporafijiensis | ns | 0.74 | ns |
| Mitochondrial electron transport | |||||
| A0A200QHI3 | AT3G52300 | ATP synthase subunit d, mitochondrial Macleaya cordata | 0.80 | ns | 0.67 |
| W5BEP1 | AT3G52300 | ATP synthase subunit d, mitochondrial Triticum aestivum | ns | 2.20 | 2.37 |
| A0A0K9R2N3 | AT3G52300 | ATP synthase subunit d, mitochondrial Spinacia oleracea | ns | 1.24 | ns |
| V4LNR7 | AT3G52300 | ATP synthase subunit d, mitochondrial Eutrema salsugineum | ns | ns | 0.70 |
| A0A1J3KAQ0 | AT5G08530 | NADH dehydrogenase [ubiquinone] flavoprotein 1, mitochondrial Noccaea caerulescens | ns | 0.72 | 0.72 |
| W5I0L9 | AT3G03070 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 6, mitochondrial Triticum aestivum | ns | ns | 0.68 |
| Q9SKT7 | AT5G13430 | External alternative NAD(P)H-ubiquinone oxidoreductase B4, mitochondrial Arabidopsis thaliana | ns | 3.93 | 4.23 |
| K3Y9D3 | AT5G13430 | Cytochrome b-c1 complex subunit Rieske, mitochondrial Setaria italica | ns | ns | 0.78 |
| Protein synthesis and elongation | |||||
| A0A0D3H000 | AT4G11120 | Elongation factor Ts, mitochondrial Oryza barthii | 1.35 | 0.65 | ns |
| B6T7S2 | AT5G47320 | 40S ribosomal protein S19 mitochondrial Zea mays | ns | 0.59 | 0.58 |
| F2DG12 | AT2G45030 | Elongation factor G, mitochondrial Hordeum vulgare subsp. vulgare | 1.33 | 0.46 | 0.66 |
| Q5JNL6 | AT1G51980 | Mitochondrial processing peptidase Oryza sativa subsp. japonica | ns | ns | 0.68 |
| B6U5I0 | AT2G29530 | Mitochondrial import inner membrane translocase subunit Tim10 Zea mays | ns | 0.72 | 0.62 |
| Q6EN45 | AT5G53140 | Probable protein phosphatase 2C member 13, mitochondrial Oryza sativa subsp. japonica | ns | 0.42 | 0.62 |
| A0A0D3GV84 | AT5G26860 | Lon protease homolog, mitochondrial Oryza barthii | ns | 0.66 | 0.75 |
| Q8L7B5 | AT2G33210 | Chaperonin CPN60-like 1, mitochondrial Arabidopsis thaliana | ns | ns | 1.49 |
| F2EDF6 | AT4G11120 | Elongation factor Ts, mitochondrial Hordeum vulgare subsp. vulgare | ns | 0.74 | ns |
| W5C618 | AT5G26860 | Lon protease homolog, mitochondrial Triticum aestivum | ns | 0.76 | ns |
| Signalling, G-protein | |||||
| F2CSX0 | AT5G27540 | Mitochondrial Rho GTPase Hordeum vulgare subsp. vulgare | ns | ns | 0.75 |
| A0A1Q3B5G5 | AT5G27540 | Mitochondrial Rho GTPase Cephalotus follicularis | ns | 0.40 | 0.42 |
| F2E3Y6 | AT5G39900 | Translation factor GUF1 homolog, mitochondrial Hordeum vulgare subsp. vulgare | ns | 0.76 | ns |
| RNA transcription | |||||
| Q7X745-2 | AT5G39840 | Isoform 2 of ATP-dependent RNA helicase SUV3L, mitochondrial Oryza sativa subsp. japonica | 1.33 | 0.62 | ns |
| Q6K7E2 | AT2G44020 | Mitochondrial transcription termination factor-like Oryza sativa subsp. japonica | ns | 0.73 | ns |
| Tetrapyrrole synthesis, protoporphyrin IX oxidase | |||||
| W5GSR7 | AT1G48520 | Glutamyl-tRNA(Gln) amidotransferase subunit B, chloroplastic/mitochondrial Triticum aestivum | ns | 0.77 | 0.77 |
| K3Y6C6 | AT5G14220 | Protoporphyrinogen oxidase Setaria italica | 0.75 | 2.43 | 1.83 |
| Stress, abiotic, heat | |||||
| G2X6B5 | AT5G22060 | Mitochondrial protein import protein MAS5 Verticillium dahliae | ns | 1.56 | 1.66 |
| A0A1J3J8H0 | AT5G22060 | Heat shock 70 kDa protein 10, mitochondrial (Fragment) Noccaea caerulescens | ns | 0.23 | 0.28 |
| Cell devision | |||||
| F2DZF0 | AT3G57090 | Mitochondrial fission 1 protein Hordeum vulgare subsp. vulgare | ns | 1.30 | 1.25 |
| W4ZR59 | AT3G57090 | Mitochondrial fission 1 protein Triticum aestivum | ns | 0.73 | 0.68 |
| Transport metobolism | |||||
| Q5NAJ0 | AT1G14560 | Graves disease mitochondrial solute carrier protein-like Oryza sativa subsp. japonica | ns | 1.30 | 1.25 |
| Q10QM8 | AT5G64970 | Mitochondrial carrier protein, expressed Oryza sativa subsp. japonica | ns | ns | 0.68 |
| Misc, oxidase-copper, flavone | |||||
| B4G146 | AT5G06580 | d-lactate dehydrogenase [cytochrome] mitochondrial Zea mays | 0.69 | 1.58 | ns |
| Amino acid metabolism | |||||
| B6SWZ4 | AT4G34030 | Methylcrotonoyl-CoA carboxylase beta chain mitochondrial Zea mays | ns | ns | 1.21 |
| Co-factor and vitamine metabolism, lipoic acid | |||||
| U5H066 | AT5G08415 | Lipoyl synthase, mitochondrial Microbotryum lychnidis-dioicae | ns | 0.73 | 0.72 |
| Redox, Glutaredoxins | |||||
| Q0JQ97 | AT3G15660 | Monothiol glutaredoxin-S1, mitochondrial Oryza sativa subsp. japonica | ns | 0.78 | ns |
| Nucleotide metabolism | |||||
| K3ZE81 | AT5G23300 | Dihydroorotate dehydrogenase (quinone), mitochondrial Setaria italica | ns | ns | 0.75 |
| C1-metabolism formate dehydrogenase | |||||
| A0A0D3GGT7 | AT5G14780 | Formate dehydrogenase, mitochondrial Oryza barthii | ns | 0.72 | 0.67 |
| Major CHO metabolism, Degradation, sucrose, Invertases, nautral | |||||
| Q10MC0 | AT1G56560 | Neutral/alkaline invertase 1, mitochondrial Oryza sativa subsp. japonica | ns | 0.76 | ns |
| Not assigned | |||||
| W5BQ98 | AT4G22310 | Mitochondrial pyruvate carrier Triticum aestivum | 1.31 | ns | ns |
| Q6ZGV8 | AT3G52140 | Clustered mitochondria protein homolog Oryza sativa subsp. japonica | ns | 1.68 | 1.91 |
| Q0JDA2 | AT1G47420 | Succinate dehydrogenase subunit 5, mitochondrial Oryza sativa subsp. japonica | ns | 1.32 | 1.23 |
| B6SPH3 | AT5G08040 | Mitochondrial import receptor subunit TOM5-like protein Zea mays | 1.45 | 0.33 | 0.45 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mao, C.; Zhu, Y.; Cheng, H.; Yan, H.; Zhao, L.; Tang, J.; Ma, X.; Mao, P. Nitric Oxide Regulates Seedling Growth and Mitochondrial Responses in Aged Oat Seeds. Int. J. Mol. Sci. 2018, 19, 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041052
Mao C, Zhu Y, Cheng H, Yan H, Zhao L, Tang J, Ma X, Mao P. Nitric Oxide Regulates Seedling Growth and Mitochondrial Responses in Aged Oat Seeds. International Journal of Molecular Sciences. 2018; 19(4):1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041052
Chicago/Turabian StyleMao, Chunli, Yanqiao Zhu, Hang Cheng, Huifang Yan, Liyuan Zhao, Jia Tang, Xiqing Ma, and Peisheng Mao. 2018. "Nitric Oxide Regulates Seedling Growth and Mitochondrial Responses in Aged Oat Seeds" International Journal of Molecular Sciences 19, no. 4: 1052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19041052