Co-Administration of Melatonin Effectively Enhances the Therapeutic Effects of Pioglitazone on Mesenchymal Stem Cells Undergoing Indoxyl Sulfate-Induced Senescence through Modulation of Cellular Prion Protein Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
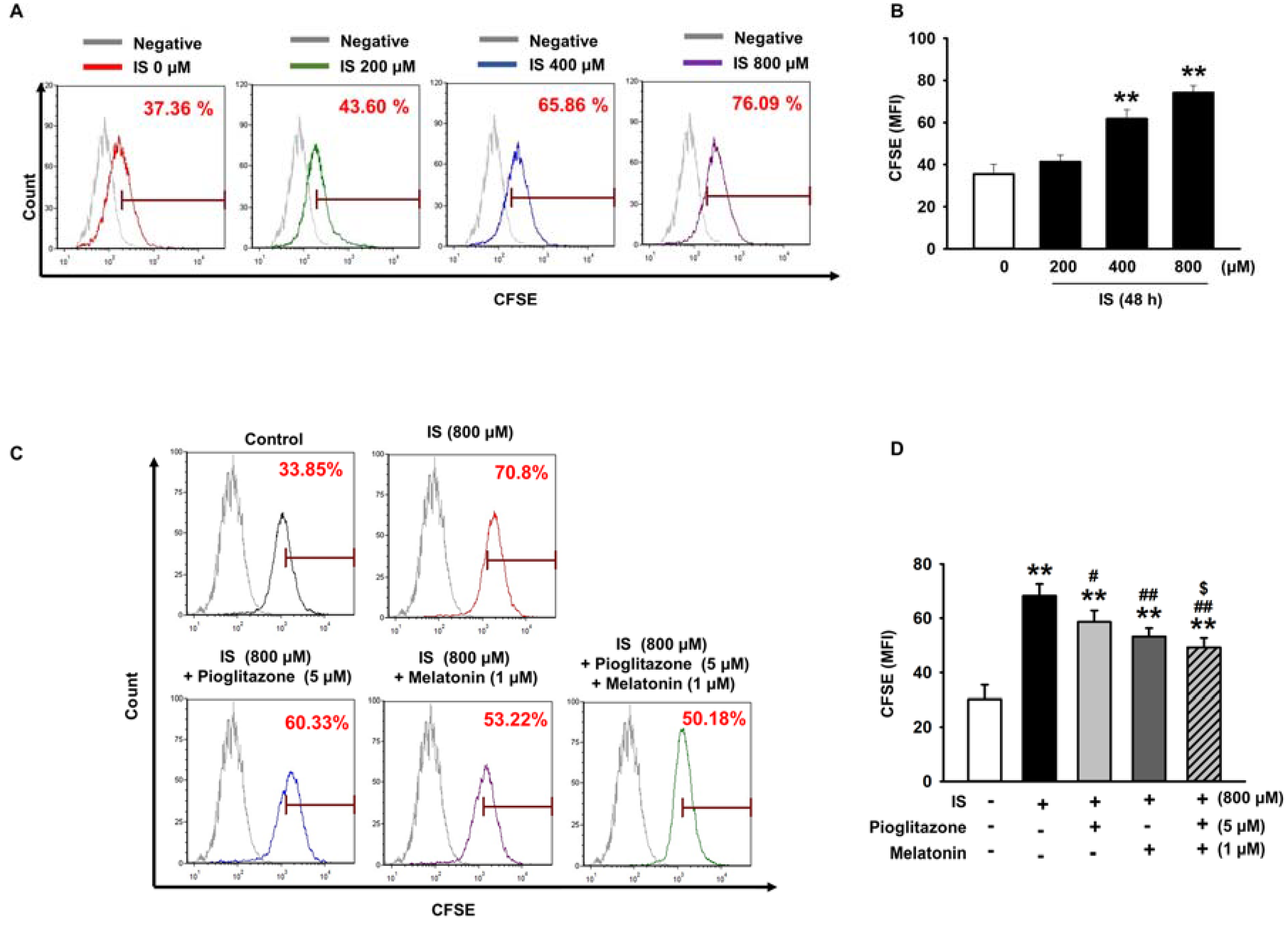
2.1. Pioglitazone and Melatonin Protected from Uremic Toxin-Induced Senescence
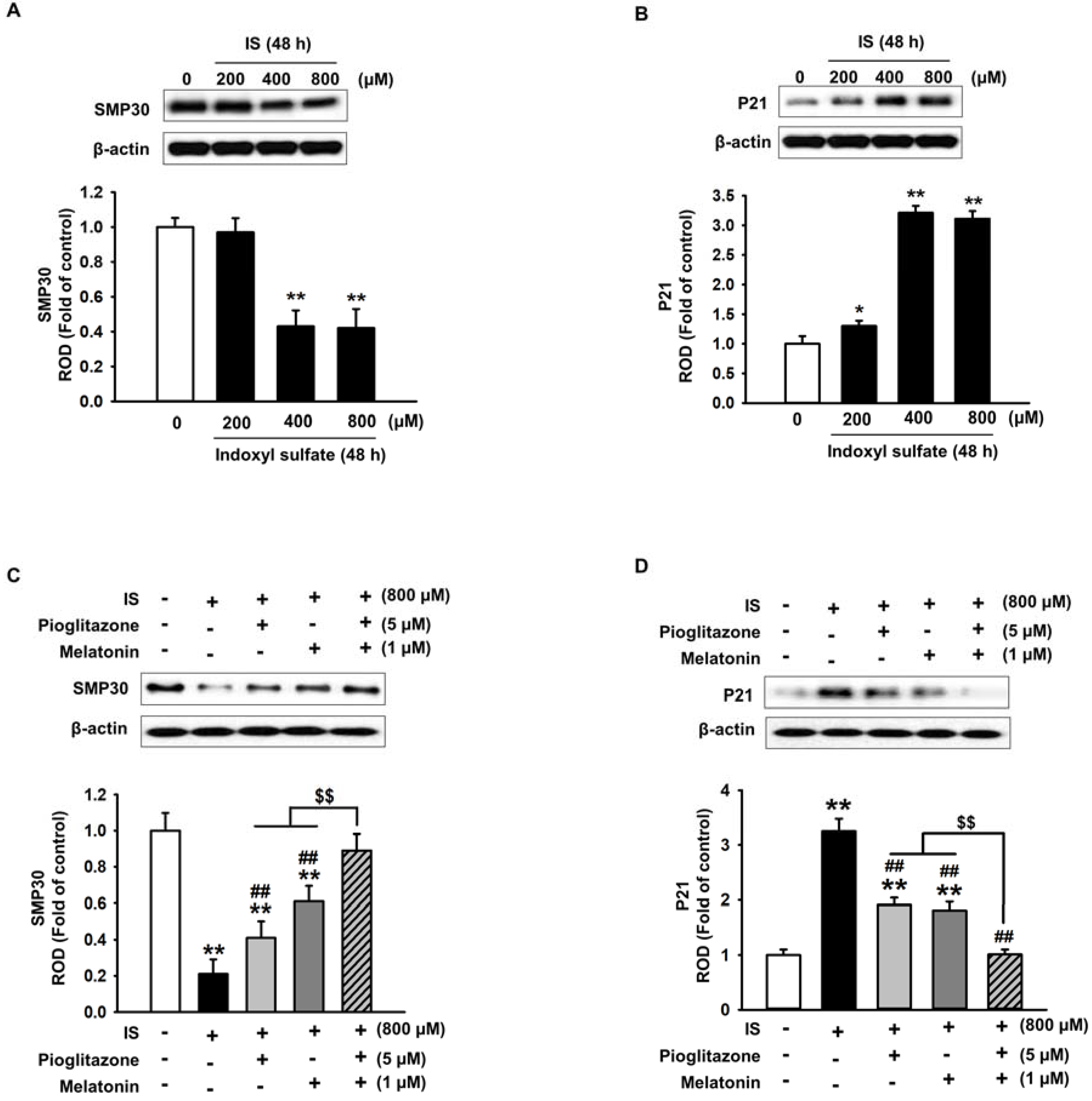
2.2. Pioglitazone and Melatonin Regulate Cell Senescence-Related Protein Expression
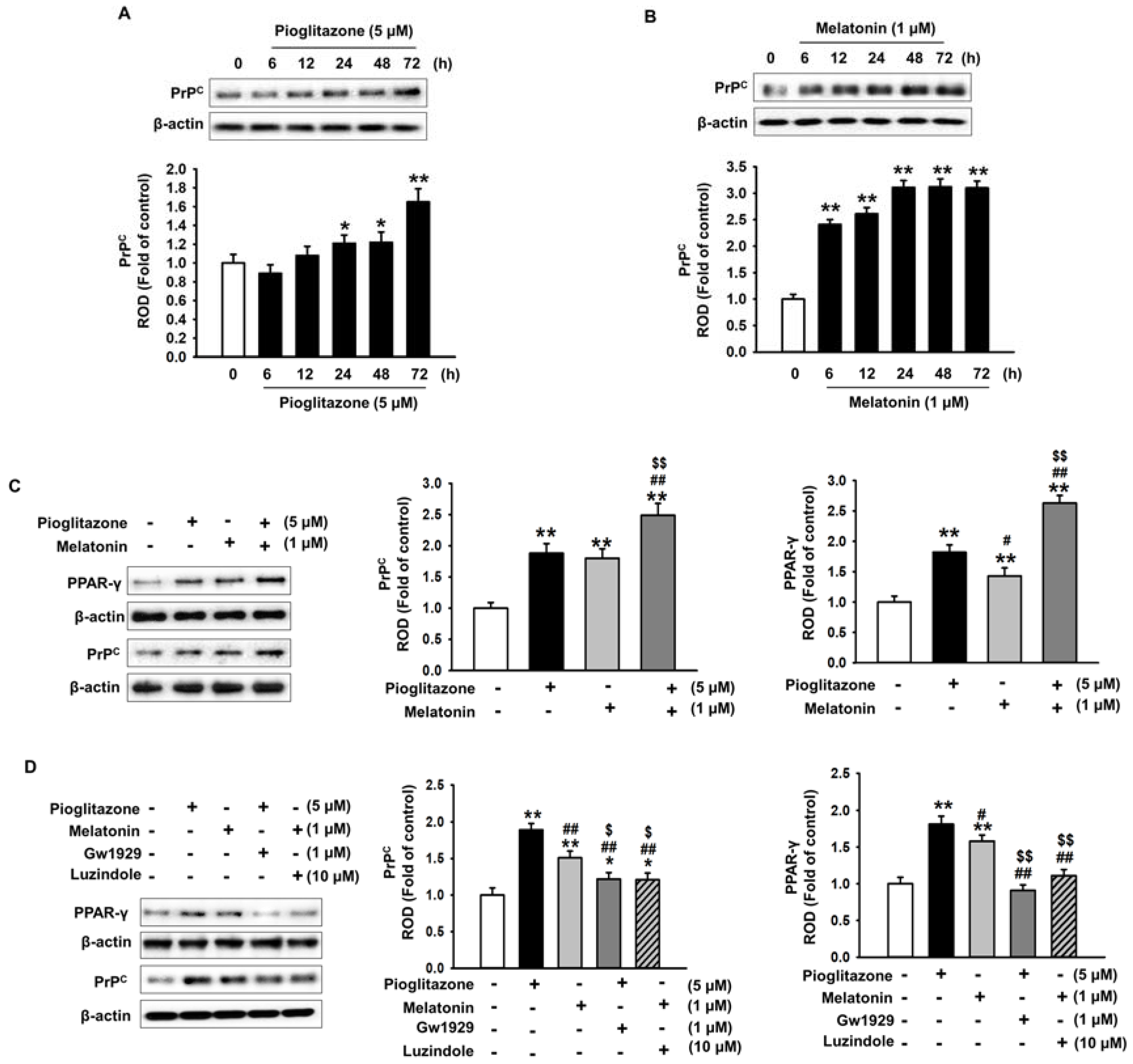
2.3. Pioglitazone and Melatonin Increase Cellular Prion Protein Expression
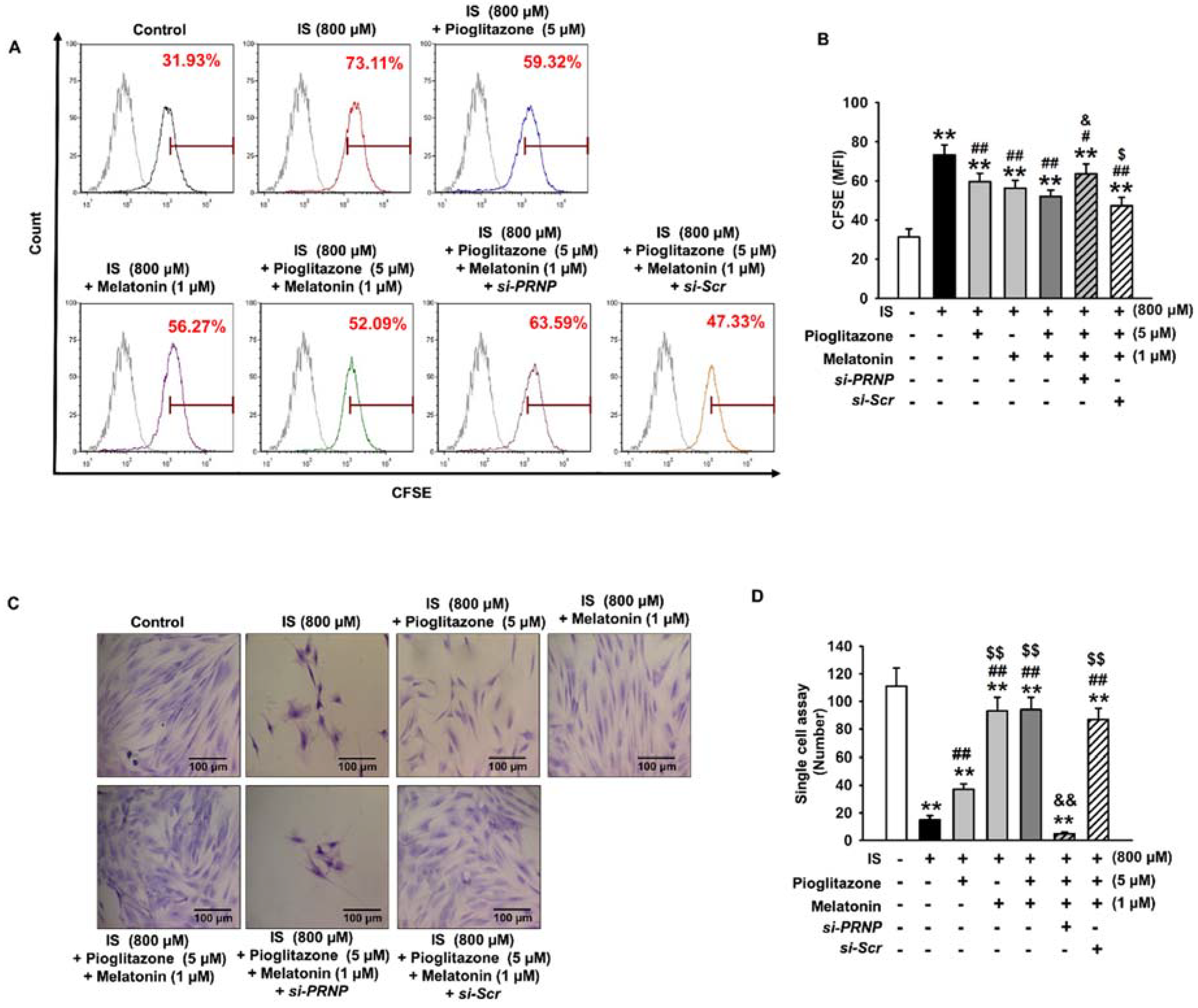
2.4. Inhibition of Senescence through the Expression of Cellular Prion Protein
3. Discussion
4. Materials and Methods
4.1. Mesenchymal Stem Cell Culture
4.2. BrdU Incorporation Assay
4.3. Beta-Galactosidase Staining Assay
4.4. Western Blot Analysis
4.5. CFSE Staining Assay
4.6. siRNA Transfection
4.7. Single-Cell Cultivation Assay
4.8. Statistical Analyses
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Wu, I.W.; Hsu, K.H.; Lee, C.C.; Sun, C.Y.; Hsu, H.J.; Tsai, C.J.; Tzen, C.Y.; Wang, Y.C.; Lin, C.Y.; Wu, M.S. P-cresyl sulphate and indoxyl sulphate predict progression of chronic kidney disease. Nephrol. Dial. Transplant. 2011, 26, 938–947. [Google Scholar] [CrossRef] [PubMed]
- Jha, V.; Garcia-Garcia, G.; Iseki, K.; Li, Z.; Naicker, S.; Plattner, B.; Saran, R.; Wang, A.Y.; Yang, C.W. Chronic kidney disease: Global dimension and perspectives. Lancet 2013, 382, 260–272. [Google Scholar] [CrossRef]
- Saran, R.; Li, Y.; Robinson, B.; Abbott, K.C.; Agodoa, L.Y.; Ayanian, J.; Bragg-Gresham, J.; Balkrishnan, R.; Chen, J.L.; Cope, E.; et al. US Renal Data System 2015 annual data report: Epidemiology of kidney disease in the united states. Am. J. Kidney Dis. 2016, 67, S1–S305. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, A.A.; Segel, J.E.; Zhuo, X.; Hoerger, T.J.; Imai, K.; Williams, D. Medical costs of CKD in the medicare population. J. Am. Soc. Nephrol. 2013, 24, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, Y.; Hamano, T.; Nakano, C.; Obi, Y.; Matsui, I.; Kusunoki, Y.; Mori, D.; Oka, T.; Hashimoto, N.; Takabatake, Y.; et al. Association between density of coronary artery calcification and serum magnesium levels among patients with chronic kidney disease. PLoS ONE 2016, 11, e0163673. [Google Scholar] [CrossRef] [PubMed]
- Santana, A.C.; Degaspari, S.; Catanozi, S.; Delle, H.; de Sa Lima, L.; Silva, C.; Blanco, P.; Solez, K.; Scavone, C.; Noronha, I.L. Thalidomide suppresses inflammation in adenine-induced CKD with uraemia in mice. Nephrol. Dial. Transplant. 2013, 28, 1140–1149. [Google Scholar] [CrossRef] [PubMed]
- Sarnak, M.J.; Levey, A.S.; Schoolwerth, A.C.; Coresh, J.; Culleton, B.; Hamm, L.L.; McCullough, P.A.; Kasiske, B.L.; Kelepouris, E.; Klag, M.J.; et al. Kidney disease as a risk factor for development of cardiovascular disease: A statement from the American heart association councils on kidney in cardiovascular disease, high blood pressure research, clinical cardiology, and epidemiology and prevention. Hypertension 2003, 42, 1050–1065. [Google Scholar] [CrossRef] [PubMed]
- Tsuruya, K.; Yoshida, H.; Kuroki, Y.; Nagata, M.; Mizumasa, T.; Mitsuiki, K.; Yoshiura, T.; Hirakawa, M.; Kanai, H.; Hori, K.; et al. Brain atrophy in peritoneal dialysis and CKD stages 3–5: A cross-sectional and longitudinal study. Am. J. Kidney Dis. 2015, 65, 312–321. [Google Scholar] [CrossRef] [PubMed]
- Vanholder, R.; Schepers, E.; Pletinck, A.; Nagler, E.V.; Glorieux, G. The uremic toxicity of indoxyl sulfate and p-cresyl sulfate: A systematic review. J. Am. Soc. Nephrol. 2014, 25, 1897–1907. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Ise, M. Indoxyl sulfate, a circulating uremic toxin, stimulates the progression of glomerular sclerosis. J. Lab. Clin. Med. 1994, 124, 96–104. [Google Scholar] [PubMed]
- Niwa, T. Removal of protein-bound uraemic toxins by haemodialysis. Blood Purif. 2013, 35 (Suppl. 2), 20–25. [Google Scholar] [CrossRef] [PubMed]
- Imasawa, T.; Utsunomiya, Y.; Kawamura, T.; Zhong, Y.; Nagasawa, R.; Okabe, M.; Maruyama, N.; Hosoya, T.; Ohno, T. The potential of bone marrow-derived cells to differentiate to glomerular mesangial cells. J. Am. Soc. Nephrol. 2001, 12, 1401–1409. [Google Scholar] [PubMed]
- Ito, T.; Suzuki, A.; Imai, E.; Okabe, M.; Hori, M. Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J. Am. Soc. Nephrol. 2001, 12, 2625–2635. [Google Scholar] [PubMed]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells-current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef] [PubMed]
- Eirin, A.; Lerman, L.O. Mesenchymal stem cell treatment for chronic renal failure. Stem Cell Res. Ther. 2014, 5, 83. [Google Scholar] [CrossRef] [PubMed]
- Aghajani Nargesi, A.; Lerman, L.O.; Eirin, A. Mesenchymal stem cell-derived extracellular vesicles for kidney repair: Current status and looming challenges. Stem Cell Res. Ther. 2017, 8, 273. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, X.; Wang, W.; Li, J.; Li, Y.; Li, L.; Wang, S.; Zhang, J.; Zhang, Y.; Huang, H. The effects of indoxyl sulfate on human umbilical cord-derived mesenchymal stem cells in vitro. Cell. Physiol. Biochem. 2016, 38, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Banfi, A.; Muraglia, A.; Dozin, B.; Mastrogiacomo, M.; Cancedda, R.; Quarto, R. Proliferation kinetics and differentiation potential of ex vivo expanded human bone marrow stromal cells: Implications for their use in cell therapy. Exp. Hematol. 2000, 28, 707–715. [Google Scholar] [CrossRef]
- Dou, L.; Jourde-Chiche, N.; Faure, V.; Cerini, C.; Berland, Y.; Dignat-George, F.; Brunet, P. The uremic solute indoxyl sulfate induces oxidative stress in endothelial cells. J. Thromb. Haemost. 2007, 5, 1302–1308. [Google Scholar] [CrossRef] [PubMed]
- Tumur, Z.; Niwa, T. Indoxyl sulfate inhibits nitric oxide production and cell viability by inducing oxidative stress in vascular endothelial cells. Am. J. Nephrol. 2009, 29, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Kolachalama, V.B.; Shashar, M.; Alousi, F.; Shivanna, S.; Rijal, K.; Belghasem, M.E.; Walker, J.; Matsuura, S.; Chang, G.H.; Gibson, C.M.; et al. Uremic solute-aryl hydrocarbon receptor-tissue factor axis associates with thrombosis after vascular injury in humans. J. Am. Soc. Nephrol. 2018, 29, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, H.; Bolati, D.; Adijiang, A.; Enomoto, A.; Nishijima, F.; Dateki, M.; Niwa, T. Senescence and dysfunction of proximal tubular cells are associated with activated p53 expression by indoxyl sulfate. Am. J. Physiol. Cell Physiol. 2010, 299, C1110–C1117. [Google Scholar] [CrossRef] [PubMed]
- Hickson, L.J.; Cosio, F.G.; El-Zoghby, Z.M.; Gloor, J.M.; Kremers, W.K.; Stegall, M.D.; Griffin, M.D.; Jaffe, A.S. Survival of patients on the kidney transplant wait list: Relationship to cardiac troponin T. Am. J. Transplant. 2008, 8, 2352–2359. [Google Scholar] [CrossRef] [PubMed]
- Hickson, L.J.; El-Zoghby, Z.M.; Lorenz, E.C.; Stegall, M.D.; Jaffe, A.S.; Cosio, F.G. Patient survival after kidney transplantation: Relationship to pretransplant cardiac troponin T levels. Am. J. Transplant. 2009, 9, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Schoonover, K.L.; Hickson, L.J.; Norby, S.M.; Hogan, M.C.; Chaudhary, S.; Albright, R.C., Jr.; Dillon, J.J.; McCarthy, J.T.; Williams, A.W. Risk factors for hospitalization among older, incident haemodialysis patients. Nephrology 2013, 18, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Maung, S.C.; El Sara, A.; Chapman, C.; Cohen, D.; Cukor, D. Sleep disorders and chronic kidney disease. World J. Nephrol. 2016, 5, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Nigam, G.; Camacho, M.; Chang, E.T.; Riaz, M. Exploring sleep disorders in patients with chronic kidney disease. Nat. Sci. Sleep 2018, 10, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.P.; Han, Y.S.; Lee, J.H.; Kim, S.M.; Lee, S.H. Melatonin rescues mesenchymal stem cells from senescence induced by the uremic toxin p-cresol via inhibiting mTOR-dependent autophagy. Biomol. Ther. 2017, 26, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.; Ordonez, R.; Reiter, R.J.; Gonzalez-Gallego, J.; Mauriz, J.L. Melatonin and endoplasmic reticulum stress: Relation to autophagy and apoptosis. J. Pineal Res. 2015, 59, 292–307. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, R.J.; Gonzalez-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Su, S.C.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62, e12370. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.H.; Ting, M.K.; Verma, S.; Kuo, L.T.; Yang, N.I.; Hsieh, I.C.; Wang, S.Y.; Hung, A.; Cherng, W.J. Pioglitazone increases the numbers and improves the functional capacity of endothelial progenitor cells in patients with diabetes mellitus. Am. Heart J. 2006, 152, 1051.e1–1051.e8. [Google Scholar] [CrossRef] [PubMed]
- Gensch, C.; Clever, Y.P.; Werner, C.; Hanhoun, M.; Bohm, M.; Laufs, U. The PPAR-γ agonist pioglitazone increases neoangiogenesis and prevents apoptosis of endothelial progenitor cells. Atherosclerosis 2007, 192, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Shinmura, D.; Togashi, I.; Miyoshi, S.; Nishiyama, N.; Hida, N.; Tsuji, H.; Tsuruta, H.; Segawa, K.; Tsukada, Y.; Ogawa, S.; et al. Pretreatment of human mesenchymal stem cells with pioglitazone improved efficiency of cardiomyogenic transdifferentiation and cardiac function. Stem Cells 2011, 29, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Kim, Y.S.; Hong, S.H.; Oh, Y.M. Therapeutic effects of adipose-derived stem cells pretreated with pioglitazone in an emphysema mouse model. Exp. Mol. Med. 2016, 48, e266. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, L.; Gao, H.; Li, Y. Effect of pioglitazone on transdifferentiation of preosteoblasts from rat bone mesenchymal stem cells into adipocytes. J. Huazhong Univ. Sci. Technol. Med. Sci. 2012, 32, 530–533. [Google Scholar] [CrossRef] [PubMed]
- Nishijima, K.; Ohno, T.; Amano, A.; Kishimoto, Y.; Kondo, Y.; Ishigami, A.; Tanaka, S. Bone degeneration and its recovery in smp30/gnl-knockout mice. J. Nutr. Health Aging 2017, 21, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.M.; Narita, M.; Lowe, S.W.; Livingston, D.M. The p400 e1a-associated protein is a novel component of the p53 --> p21 senescence pathway. Genes Dev. 2005, 19, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.J.; Kim, J.K.; Hwang, S.D. Mesenchymal stem cell therapy for chronic renal failure. Expert Opin. Biol. Ther. 2010, 10, 1217–1226. [Google Scholar] [CrossRef] [PubMed]
- Papazova, D.A.; Oosterhuis, N.R.; Gremmels, H.; van Koppen, A.; Joles, J.A.; Verhaar, M.C. Cell-based therapies for experimental chronic kidney disease: A systematic review and meta-analysis. Dis. Models Mech. 2015, 8, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Hickson, L.J.; Eirin, A.; Lerman, L.O. Challenges and opportunities for stem cell therapy in patients with chronic kidney disease. Kidney Int. 2016, 89, 767–778. [Google Scholar] [CrossRef] [PubMed]
- Idziak, M.; Pedzisz, P.; Burdzinska, A.; Gala, K.; Paczek, L. Uremic toxins impair human bone marrow-derived mesenchymal stem cells functionality in vitro. Exp. Toxicol. Pathol. 2014, 66, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Kramann, R.; Couson, S.K.; Neuss, S.; Kunter, U.; Bovi, M.; Bornemann, J.; Knuchel, R.; Jahnen-Dechent, W.; Floege, J.; Schneider, R.K. Exposure to uremic serum induces a procalcific phenotype in human mesenchymal stem cells. Arterioscler. Thromb. Vasc. Biol. 2011, 31, e45–e54. [Google Scholar] [CrossRef] [PubMed]
- Tran, U.M.; Rajarajacholan, U.; Soh, J.; Kim, T.S.; Thalappilly, S.; Sensen, C.W.; Riabowol, K. LincRNA-p21 acts as a mediator of ing1b-induced apoptosis. Cell Death Dis. 2015, 6, e1668. [Google Scholar] [CrossRef] [PubMed]
- Ki, M.R.; Lee, H.R.; Park, J.K.; Hong, I.H.; Han, S.Y.; You, S.Y.; Lee, E.M.; Kim, A.Y.; Lee, S.S.; Jeong, K.S. Ascorbate promotes carbon tetrachloride-induced hepatic injury in senescence marker protein 30-deficient mice by enhancing inflammation. J. Nutr. Biochem. 2011, 22, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Wang, L.; Hou, J.; Guo, T.; Xing, Y.; Zheng, S.; Zhou, C.; Huang, H.; Long, H.; Zhong, T.; et al. Peroxisome proliferator-activated receptor γ promotes mesenchymal stem cells to express connexin43 via the inhibition of TGF-β1/smads signaling in a rat model of myocardial infarction. Stem Cell Rev. 2015, 11, 885–899. [Google Scholar] [CrossRef] [PubMed]
- Tian-Liang, Z.; Xue-Rui, W.; Run, W.; Chuan, W. Effects of PrPc on DF-1 cells’ biological processes and RNA-seq-based analysis of differential genes. J. Cell. Physiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Han, Y.S.; Yoon, Y.M.; Yun, C.W.; Yun, S.P.; Kim, S.M.; Kwon, H.Y.; Jeong, D.; Baek, M.J.; Lee, H.J.; et al. Role of hspa1l as a cellular prion protein stabilizer in tumor progression via HIF-1α/GP78 axis. Oncogene 2017, 36, 6555–6567. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Yasutaka, Y.; Nishioku, T.; Kusakabe, S.; Futagami, K.; Yamauchi, A.; Kataoka, Y. Involvement of the cellular prion protein in the migration of brain microvascular endothelial cells. Neurosci. Lett. 2011, 496, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Morel, E.; Fouquet, S.; Strup-Perrot, C.; Pichol Thievend, C.; Petit, C.; Loew, D.; Faussat, A.M.; Yvernault, L.; Pincon-Raymond, M.; Chambaz, J.; et al. The cellular prion protein PrPc is involved in the proliferation of epithelial cells and in the distribution of junction-associated proteins. PLoS ONE 2008, 3, e3000. [Google Scholar] [CrossRef]
- Lee, J.H.; Han, Y.S.; Lee, S.H. Potentiation of biological effects of mesenchymal stem cells in ischemic conditions by melatonin via upregulation of cellular prion protein expression. J. Pineal Res. 2017, 62, e12385. [Google Scholar] [CrossRef] [PubMed]
- Vanholder, R.; de Smet, R.; Glorieux, G.; Argiles, A.; Baurmeister, U.; Brunet, P.; Clark, W.; Cohen, G.; De Deyn, P.P.; Deppisch, R.; et al. Review on uremic toxins: Classification, concentration, and interindividual variability. Kidney Int. 2003, 63, 1934–1943. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ryu, J.M.; Han, Y.S.; Zia, M.F.; Kwon, H.Y.; Noh, H.; Han, H.J.; Lee, S.H. Fucoidan improves bioactivity and vasculogenic potential of mesenchymal stem cells in murine hind limb ischemia associated with chronic kidney disease. J. Mol. Cell. Cardiol. 2016, 97, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Noh, H.; Yu, M.R.; Kim, H.J.; Jang, E.J.; Hwang, E.S.; Jeon, J.S.; Kwon, S.H.; Han, D.C. Uremic toxin p-cresol induces Akt-pathway-selective insulin resistance in bone marrow-derived mesenchymal stem cells. Stem Cells 2014, 32, 2443–2453. [Google Scholar] [CrossRef] [PubMed]






© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, Y.S.; Kim, S.M.; Lee, J.H.; Lee, S.H. Co-Administration of Melatonin Effectively Enhances the Therapeutic Effects of Pioglitazone on Mesenchymal Stem Cells Undergoing Indoxyl Sulfate-Induced Senescence through Modulation of Cellular Prion Protein Expression. Int. J. Mol. Sci. 2018, 19, 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051367
Han YS, Kim SM, Lee JH, Lee SH. Co-Administration of Melatonin Effectively Enhances the Therapeutic Effects of Pioglitazone on Mesenchymal Stem Cells Undergoing Indoxyl Sulfate-Induced Senescence through Modulation of Cellular Prion Protein Expression. International Journal of Molecular Sciences. 2018; 19(5):1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051367
Chicago/Turabian StyleHan, Yong Seok, Sang Min Kim, Jun Hee Lee, and Sang Hun Lee. 2018. "Co-Administration of Melatonin Effectively Enhances the Therapeutic Effects of Pioglitazone on Mesenchymal Stem Cells Undergoing Indoxyl Sulfate-Induced Senescence through Modulation of Cellular Prion Protein Expression" International Journal of Molecular Sciences 19, no. 5: 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19051367