Anti-Metastatic Effects of Antrodan with and without Cisplatin on Lewis Lung Carcinomas in a Mouse Xenograft Model

Abstract
:1. Introduction
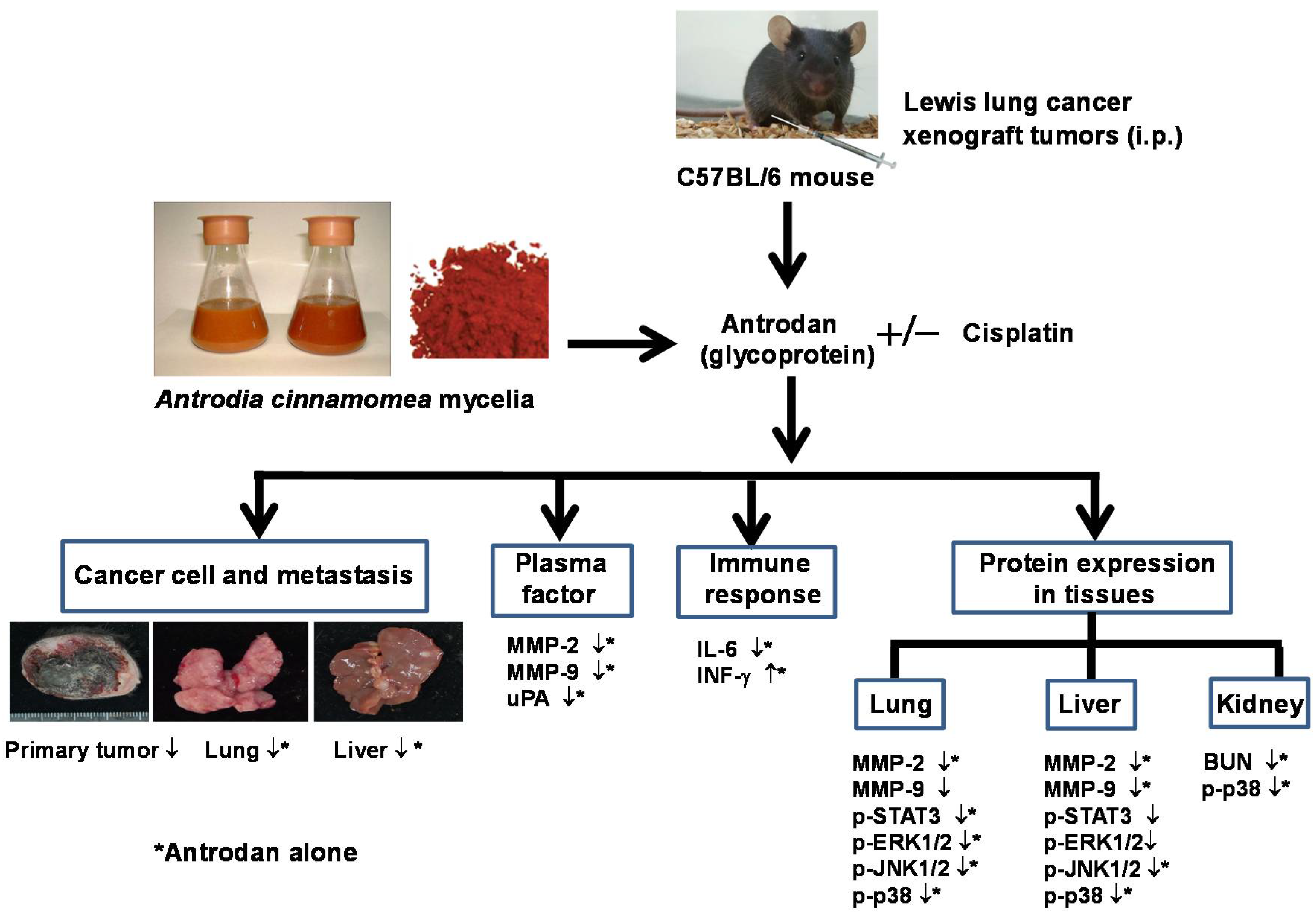
2. Results
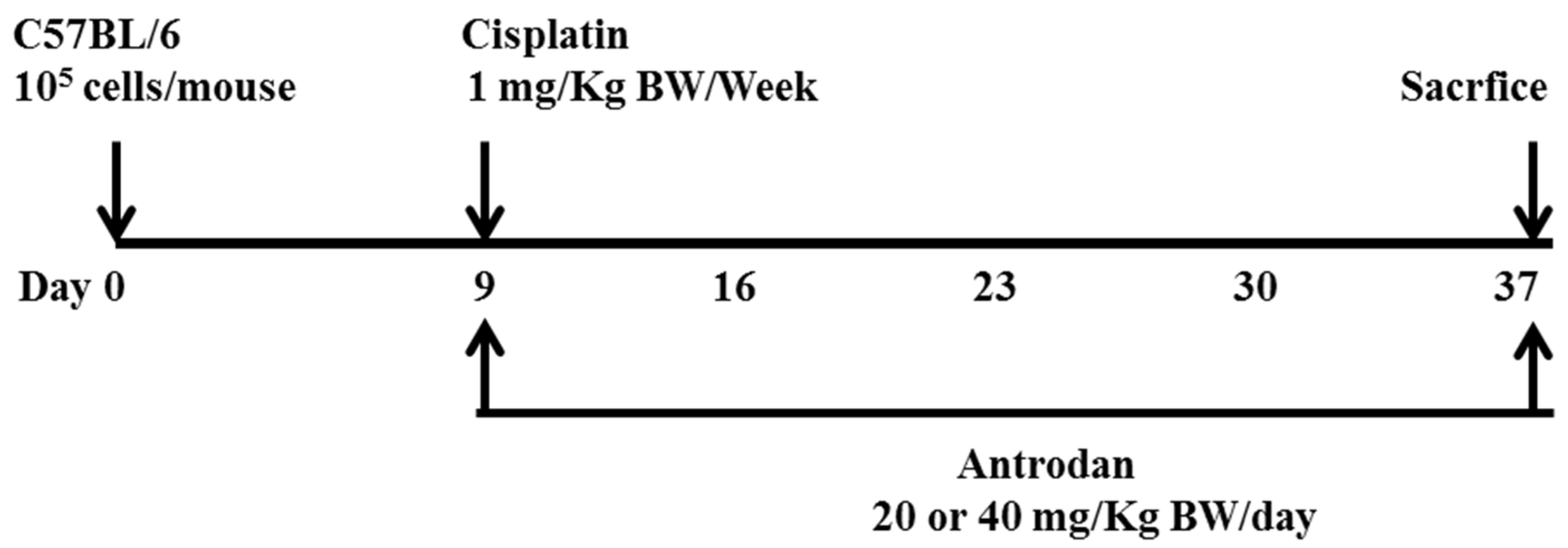
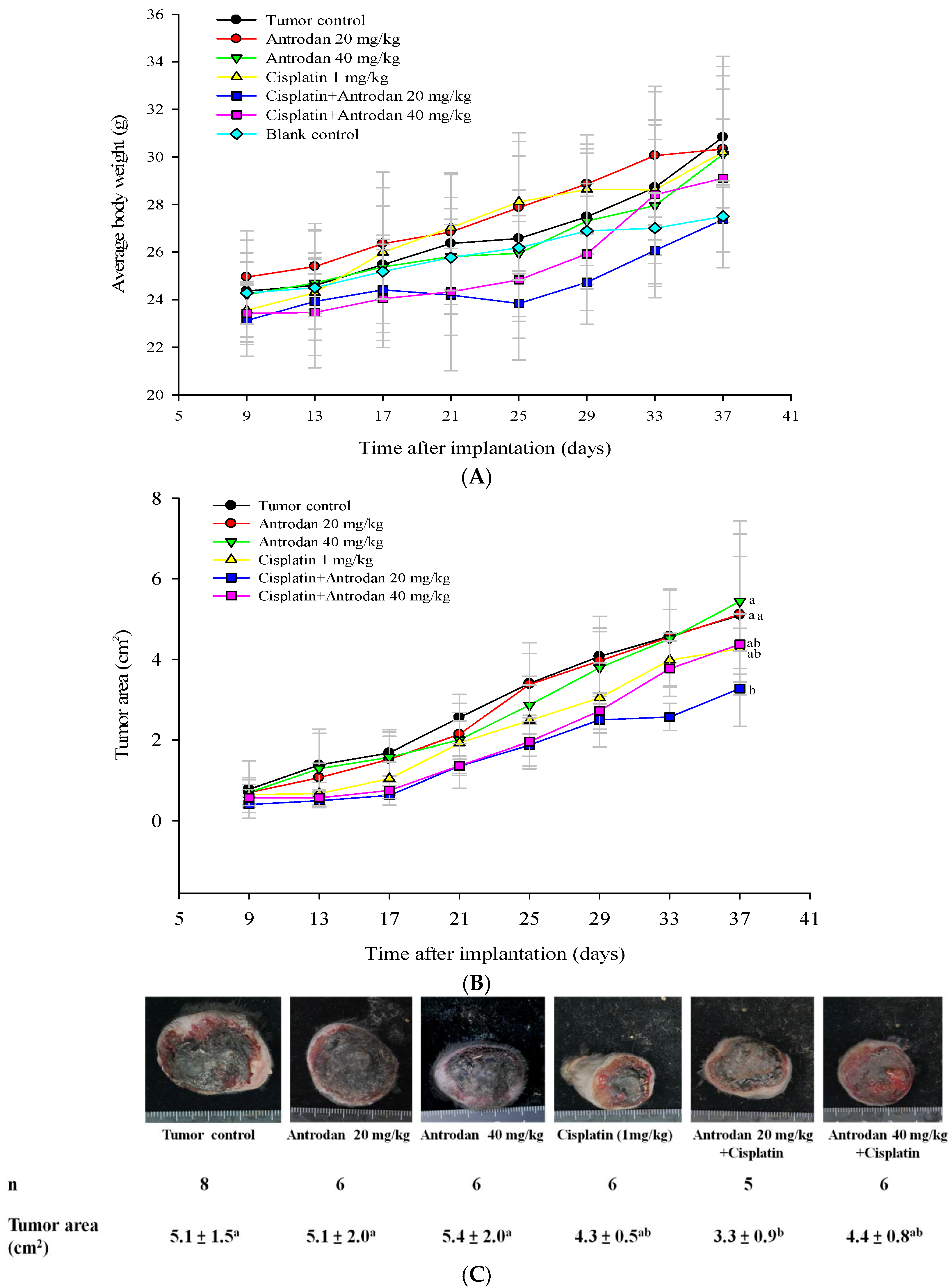
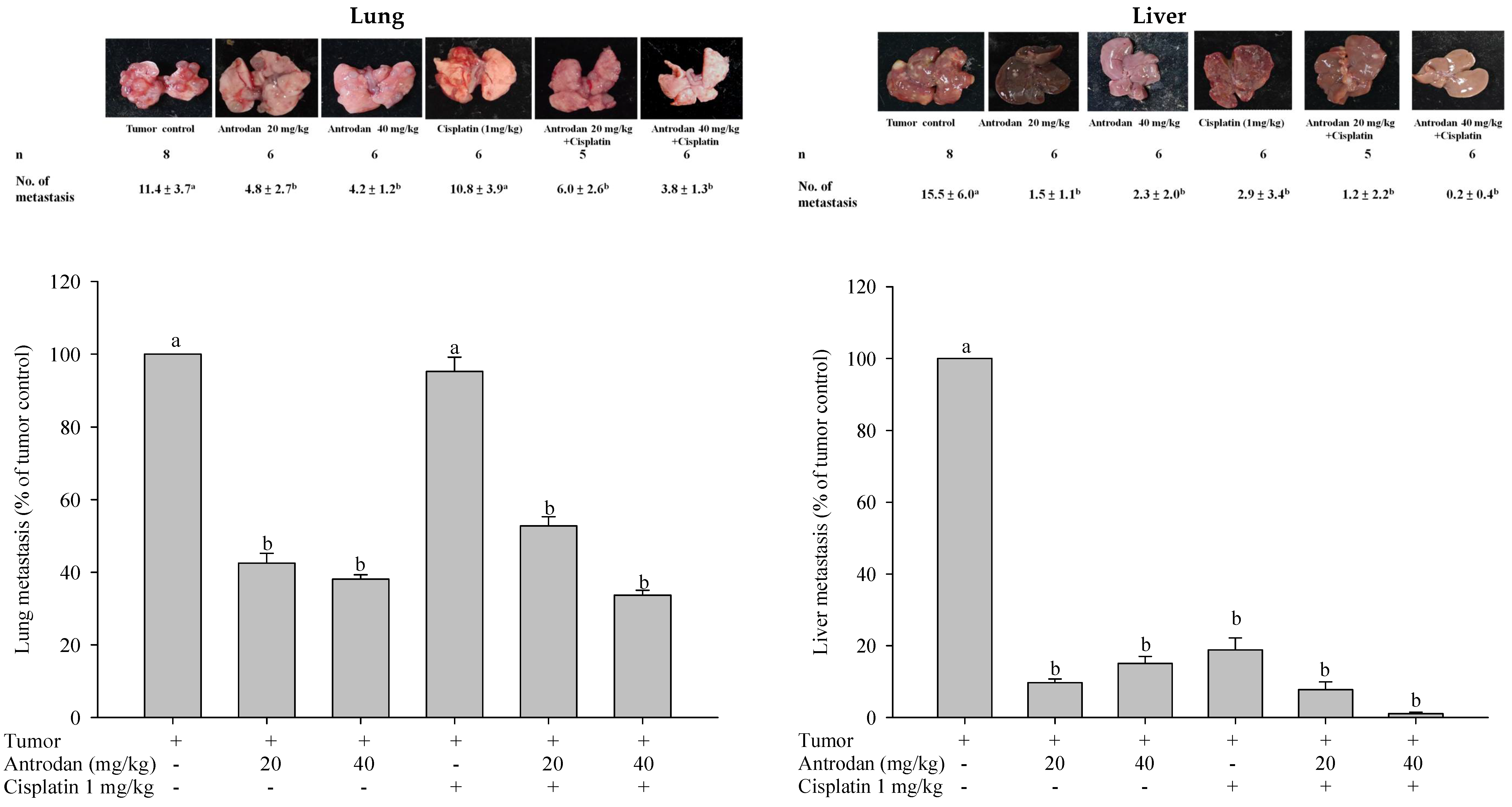
2.1. Effects of Antrodan Only, Cisplatin Only, and Their Combined Treatment on Body Weight, and Primary Tumor Growth and Metastasis
2.2. Effects of Treatment with Antrodan Only, and Its Combined Treatment with Cisplatin on IL-6 and IFN-γ Levels
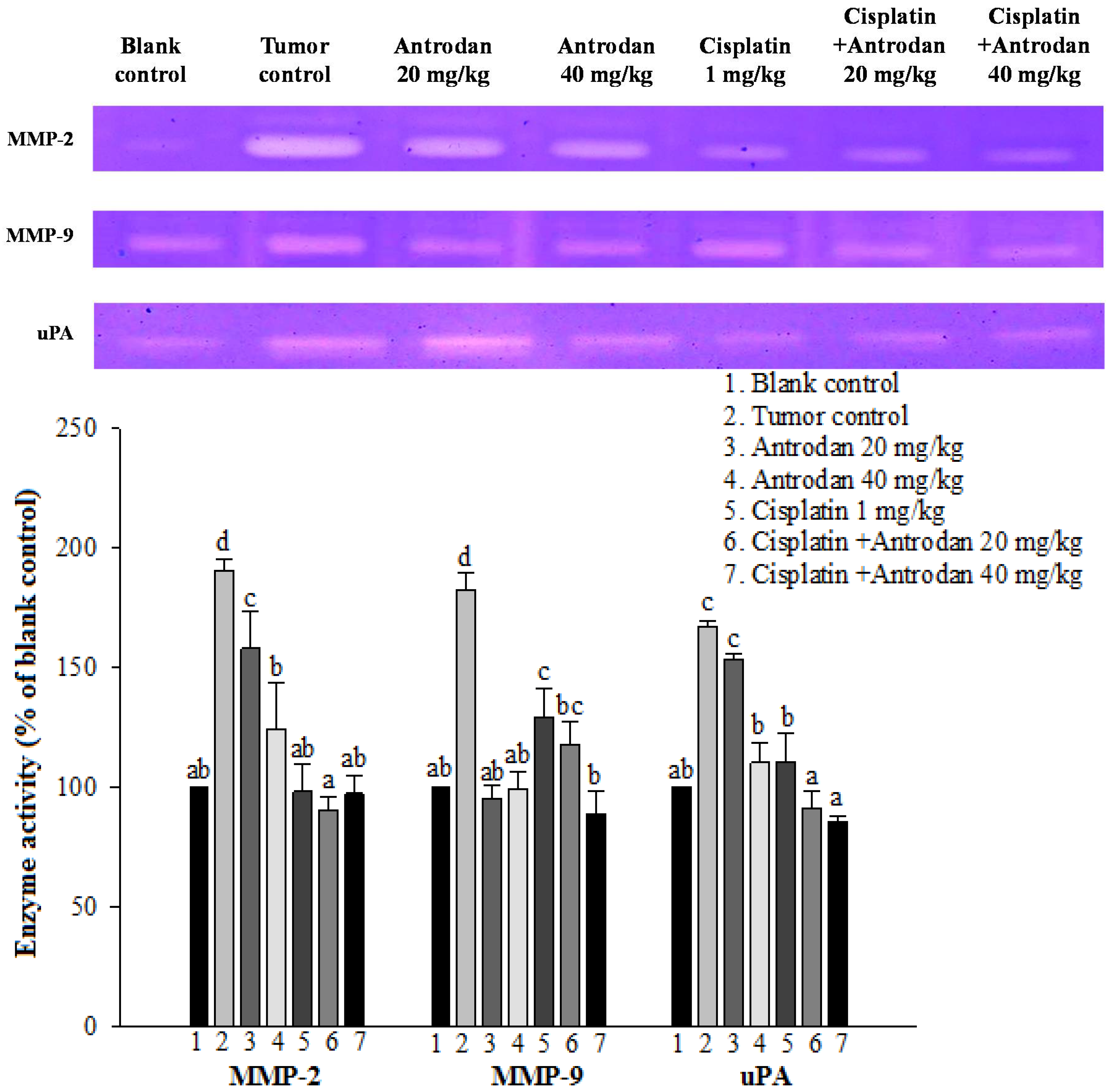
2.3. Effects of Treatment with Antrodan Only, Cisplatin Only, and a Combination of Both on Protein Activities of MMP-2, MMP-9, and uPA in Plasma of LLC-Bearing Mice
2.4. Effects of Treatment with Antrodan Only, Cisplatin Only, and a Combination of both on Protein Expression of MMP-2/9, STAT3, ERK1/2, JNK1/2, and p38, as well as on Phosphorylation of STAT3, ERK1/2, JNK1/2, and p38 in Lung and Liver Tissues of LLC-Bearing Mice
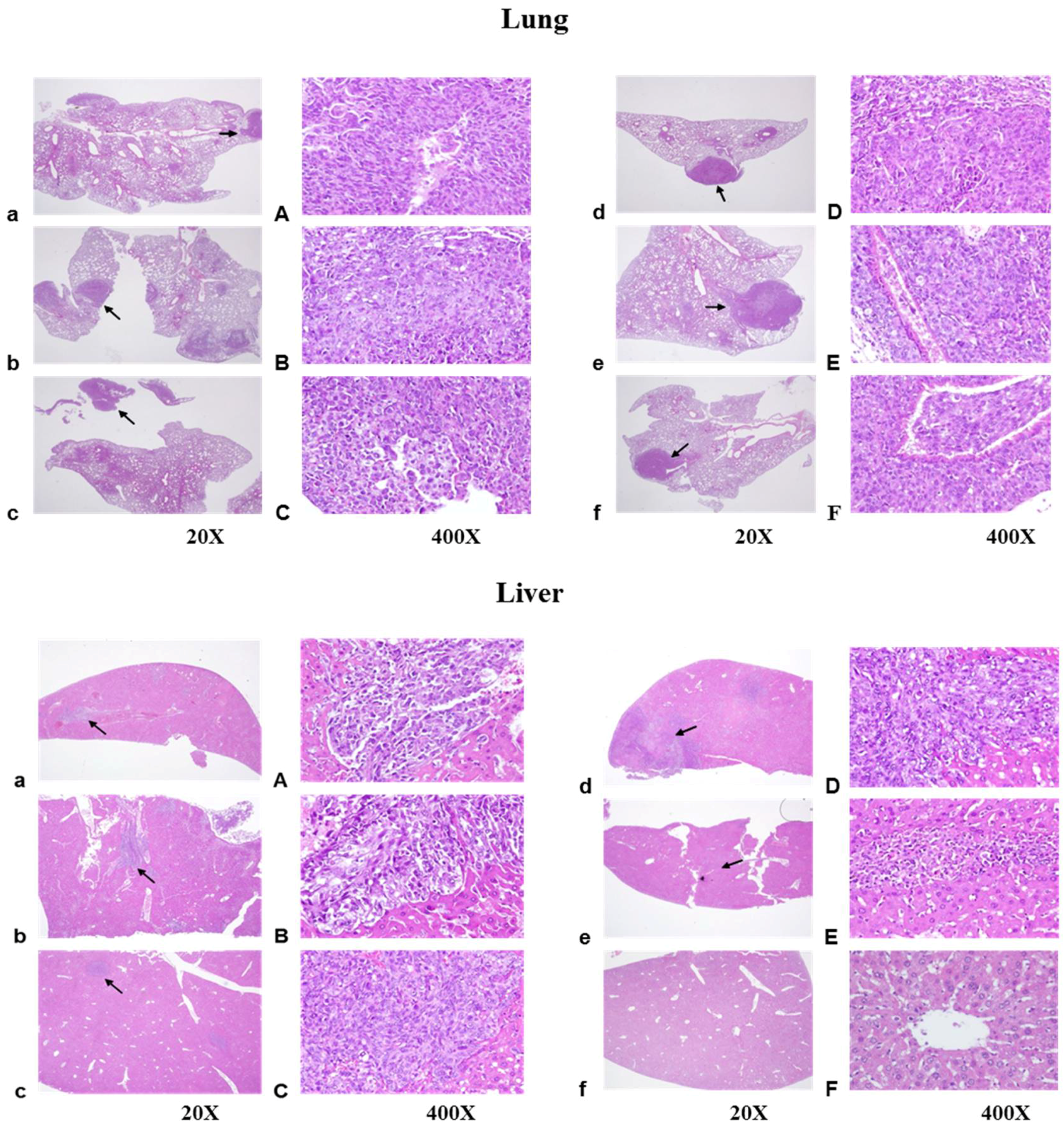
2.5. Histopathological Findings
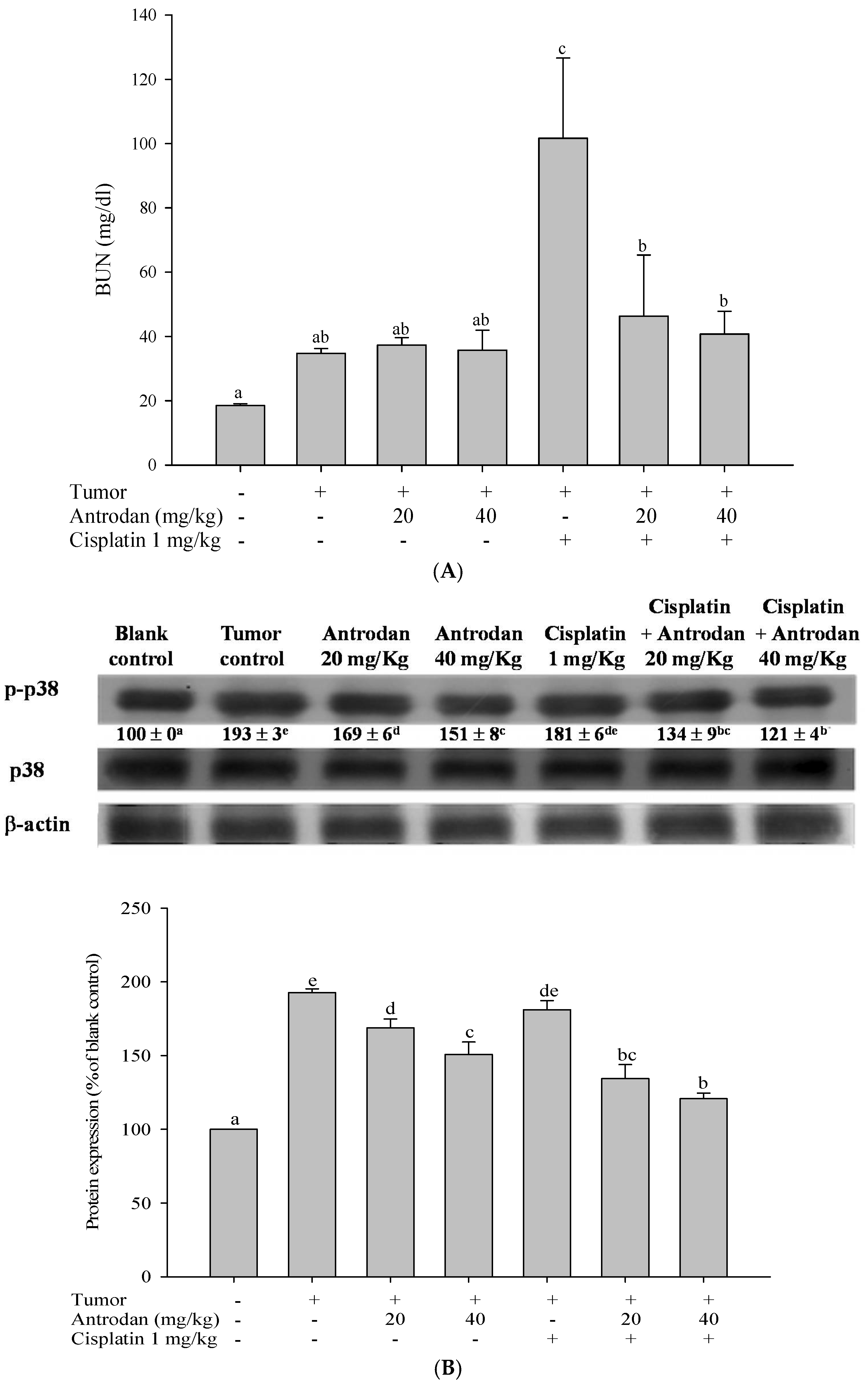
2.6. Antrodan Ameliorated Cisplatin-Induced Kidney Dysfunction in LLC-Bearing Mice
2.7. Treatments with Antrodan Only, and Its Combined with Cisplatin Reduced the Phosphorylation of Kidney p38 MAPK
3. Discussion
4. Materials and Methods
4.1. Chemicals and Antibodies
4.2. Cell Lines
4.3. Source of Antrodan
4.4. Tumor Xenografts and Antrodan/Cisplatin Treatments
4.5. Determination of the Levels of Plasma Cytokines—IL-6 and IFN-γ
4.6. Determination of MMP-2, MMP-9, and uPA Levels in Plasma through Zymography
4.7. Western Blotting
4.8. Histological Examination of Lung, Liver, and Kidney Tissues
4.9. Determination of Blood Urea Nitrogen (BUN) Levels
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgements
Conflicts of Interest
References
- Spano, D.; Heck, C.; De Antonellis, P.; Christofori, G.; Zollo, M. Molecular networks that regulate cancer metastasis. Semin. Cancer Biol. 2012, 22, 234–249. [Google Scholar] [CrossRef] [PubMed]
- Chambers, A.F.; Groom, A.C.; MacDonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Talmadge, J.E.; Fidler, I.J. AACR centennial series: The biology of cancer metastasis: Historical perspective. Cancer Res. 2010, 70, 5649–5669. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Woessner, J.F. Matrix metalloproteinases. J. Biol. Chem. 1999, 274, 21491–21494. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.; Li, M.; Luo, T.; Yin, Y.; Jiang, Y. Matrix metalloproteinases in tumorigenesis: An evolving paradigm. Cell Mol. Life Sci. 2011, 68, 3853–3868. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.O.; Park, S.J.; Yun, C.H.; Chung, A.S. Roles of matrix metalloproteinases in tumor metastasis and angiogenesis. J. Biochem. Mol. Biol. 2003, 36, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Choong, P.F.; Nadesapillai, A.P. Urokinase plasminogen activator system: A multifunctional role in tumor progression and metastasis. Clin. Orthop. Relat. Res. 2003, 415, S46–S58. [Google Scholar] [CrossRef]
- Ornstein, D.L.; Zacharski, L.R.; Memoli, V.A.; Kisiel, W.; Kudryk, B.J.; Hunt, J.; Rousseau, S.M.; Stump, D.C. Coexisting macrophage-associated fibrin formation and tumor cell urokinase in squamous cell and adenocarcinoma of the lung tissues. Cancer 1991, 68, 1061–1067. [Google Scholar] [CrossRef]
- Duffy, M.J. Urokinase plasminogen activator and its inhibitor, PAI-1, as prognostic markers in breast cancer: From pilot to level 1 evidence studies. Clin. Chem. 2002, 48, 1194–1197. [Google Scholar] [PubMed]
- Xiong, A.; Yang, Z.; Shen, Y.; Zhou, J.; Shen, Q. Transcription factor STAT3 as a novel molecular target for cancer prevention. Cancers 2014, 6, 926–957. [Google Scholar] [CrossRef] [PubMed]
- Tong, M.; Wang, J.; Jiang, N.; Pan, H.; Li, D. Correlation between p-STAT3 overexpression and prognosis in lung cancer: A systematic review and meta-analysis. PLoS ONE 2017, 12, e0182282. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Bournazou, E.; Sansone, P.; Berishaj, M.; Gao, S.P.; Daly, L.; Wels, J.; Theilen, T.; Granitto, S.; Zhang, X.; Cotari, J.; et al. The IL-6/JAK/Stat3 feed-forward loop drives tumorigenesis and metastasis. Neoplasia 2013, 15, 848–862. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L.; Lapadat, R. Mitogen-activated protein kinase pathways mediated by ERK, JNK, and p38 protein kinases. Science 2002, 298, 1911–1912. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.L.; Lippard, S.J. Sequence-dependent termination of in vitro DNA synthesis by cis- and trans-diamminedichloroplatinum (II). Proc. Natl. Acad. Sci. USA 1985, 82, 4616–4619. [Google Scholar] [CrossRef] [PubMed]
- Barabas, K.; Milner, R.; Lurie, D.; Adin, C. Cisplatin: A review of toxicities and therapeutic applications. Vet. Comp. Oncol. 2008, 6, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.P.; Tadagavadi, R.K.; Ramesh, G.; Reeves, W.B. Mechanisms of cisplatin nephrotoxicity. Toxins 2010, 2, 2490–2518. [Google Scholar] [CrossRef] [PubMed]
- Pilkington, K.; Leach, J.; Teng, L.; Storey, D.; Liu, J.P. Coriolus versicolor mushroom for colorectal cancer treatment. Cochrane Database Syst. Rev. 2016, CD012053. [Google Scholar] [CrossRef]
- Torkelson, C.J.; Sweet, E.; Martzen, M.R.; Sasagawa, M.; Wenner, C.A.; Gay, J.; Putiri, A.; Standish, L.J. Phase 1 clinical trial of Trametes versicolor in women with breast cancer. ISRN Oncol. 2012, 251632. [Google Scholar] [CrossRef] [PubMed]
- Ina, K.; Kataoka, T.; Ando, T. The use of lentinan for treating gastric cancer. Anticancer Agents Med. Chem. 2013, 13, 681–688. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.-H.; Peng, C.-C.; Ker, Y.-B.; Chen, C.-C.; Lee, A.; Chang, W.-L.; Chyau, C.-C.; Peng, R.Y. Physicochemical characteristics and anti-inflammatory activities of antrodan, a novel glycoprotein isolated from Antrodia cinnamomea mycelia. Molecules 2014, 19, 22–40. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-C.; Lin, Y.-T.; Chen, K.-C.; Chyau, C.-C.; Peng, R.Y. Antrodan, a β-glucan obtained from Antrodia cinnamomea mycelia, is beneficial to benign prostate hyperplasia. Food Funct. 2015, 6, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Ker, Y.-B.; Peng, C.-C.; Chang, W.-L.; Chyau, C.-C.; Peng, R. Hepatoprotective bioactivity of the glycoprotein, antrodan, isolated from Antrodia cinnamomea mycelia. PLoS ONE 2014, 9, e93191. [Google Scholar] [CrossRef] [PubMed]
- Fa, K.-N.; Yang, C.-M.; Chen, P.-C.; Lee, Y.-Y.; Chyau, C.-C.; Hu, M.-L. Anti-metastatic effects of antrodan, the Antrodia cinnamomea mycelia glycoprotein, in lung carcinoma cells. Int. J. Biol. Macromol. 2015, 74, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Mignatti, P.; Rifkin, D.B. Biology and biochemistry of proteinases in tumor invasion. Physiol. Rev. 1993, 73, 161–195. [Google Scholar] [CrossRef] [PubMed]
- Stamenkovic, I. Matrix metalloproteinases in tumor invasion and metastasis. Semin. Cancer Biol. 2000, 10, 415–433. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; New, L.; Pan, Z.; Han, J.; Nemerow, G.R. Urokinase plasminogen activator/urokinase-specific surface receptor expression and matrix invasion by breast cancer cells requires constitutive p38alpha mitogen-activated protein kinase activity. J. Biol. Chem. 2000, 275, 12266–12272. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.; Han, Y.; Hughart, N.; McCarra, J.; Alpini, G.; Meng, F. Interleukin-6 and its receptor, key players in hepatobiliary inflammation and cancer. Transl. Gastrointest. Cancer 2012, 1, 58–70. [Google Scholar] [PubMed]
- Ikeda, H.; Old, L.J.; Schreiber, R.D. The roles of IFNγ in protection against tumor development and cancer immunoediting. Cytokine Growth Factor Rev. 2002, 13, 95–109. [Google Scholar] [CrossRef]
- Reddy, K.B.; Nabha, S.M.; Atanaskova, N. Role of MAP kinase in tumor progression and invasion. Cancer Metast. Rev. 2003, 22, 395–403. [Google Scholar] [CrossRef]
- Roberts, P.J.; Der, C.J. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene 2007, 26, 3291–3310. [Google Scholar] [CrossRef] [PubMed]
- Zarubin, T.; Jiahuai, H.A.N. Activation and signaling of the p38 MAP kinase pathway. Cell Res. 2005, 15, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Koul, S.; Petersen, J.; Khandrika, L.; Hwa, J.S.; Meacham, R.B.; Wilson, S.; Koul, H.K. p38 Mitogen-Activated Protein Kinase–Driven MAPKAPK2 Regulates Invasion of Bladder Cancer by Modulation of MMP-2 and MMP-9 Activity. Cancer Res. 2010, 70, 832–841. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, P.A.; Viswanatha, G.L.; Radheshyam, N.; Mukund, H.; Belliyappa, M.S. Science behind cisplatin-induced nephrotoxicity in humans: A clinical study. Asian Pac. J. Trop. Biomed. 2012, 2, 640–644. [Google Scholar] [CrossRef]
- Tisdale, M.J. Cachexia in cancer patients. Nat. Rev. Cancer 2002, 2, 862–871. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, G.; Reeves, W.B. p38 MAP kinase inhibition ameliorates cisplatin nephrotoxicity in mice. Am. J. Physiol. Renal Physiol. 2005, 289, F166–F174. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Stetler-Stevenson, W.G. Quantitative zymography: Detection of picogram quantities of gelatinases. Anal. Biochem. 1994, 218, 325–329. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Body Weight (g) | Relative Body Weight (g) 2 | Primary Tumor Weight (g) | Relative Weight of Organs 3 | ||
|---|---|---|---|---|---|---|---|
| Initial | Final | Liver | Kidney | ||||
| Blank | 6 | 24.3 ± 1.3 a | 27.6 ± 1.5 a | 27.6 ± 1.5 a | — | 3.7 ± 0.5 b,c | 1.2 ± 0.2 a |
| Tumor control | 8 | 24.4 ± 2.1 a | 30.8 ± 3.0 a | 20.4 ± 3.6 b,c | 10.4 ± 2.5 a | 4.5 ± 0.9 a,b | 1.0 ± 0.3 a |
| Antrodan 20 mg/kg | 6 | 24.9 ± 1.9 a | 30.3 ± 3.1 a | 19.4 ± 2.4 c | 10.9 ± 2.4 a | 4.0 ± 0.1 b | 1.0 ± 0.1 a |
| Antrodan 40 mg/kg | 6 | 24.2 ± 1.8 a | 30.1 ± 4.1 a | 19.2 ± 1.0 c | 10.1 ± 3.1 a,b | 4.3 ± 0.7 a,b | 1.1 ± 0.3 a |
| Cisplatin 1 mg/kg | 6 | 23.6 ± 1.1 a | 30.2 ± 1.4 a | 23.0 ± 2.0 b | 6.8 ± 1.7 c | 4.5 ± 0.9 a,b | 1.1 ± 0.2 a |
| Antrodan 20 mg/kg + Cisplatin | 5 | 23.1 ± 1.5 a | 27.4 ± 1.4 a | 20.4 ± 1.5 b,c | 6.9 ± 0.6 c | 5.0 ± 0.3 a | 1.2 ± 0.1 a |
| Antrodan 40 mg/kg + Cisplatin | 6 | 23.4 ± 1.3 a | 29.1 ± 3.8 a | 21.4 ± 3.6 b,c | 7.7 ± 0.9 b,c | 4.3 ± 0.6 a,b | 1.0 ± 0.1 a |
| Group | n | IL-6 (pg/mL) | IFN-γ (pg/mL) |
|---|---|---|---|
| Blank | 6 | 135 ± 30 a | 534 ± 221 a |
| Tumor control | 8 | 293 ± 91 b | 2410 ± 450 b |
| Antrodan 20 mg/kg | 6 | 179 ± 28 a | 2562 ± 569 b |
| Antrodan 40 mg/kg | 6 | 169 ± 38 a | 4872 ± 482 c |
| Cisplatin 1 mg/kg | 6 | 275 ± 84 b | 2510 ± 138 b |
| Antrodan 20 mg/kg + Cisplatin | 5 | 172 ± 18 a | 2760 ± 547 b |
| Antrodan 40 mg/kg + Cisplatin | 6 | 173 ± 36 a | 3013 ± 375 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, P.-C.; Chen, C.-C.; Ker, Y.-B.; Chang, C.-H.; Chyau, C.-C.; Hu, M.-L. Anti-Metastatic Effects of Antrodan with and without Cisplatin on Lewis Lung Carcinomas in a Mouse Xenograft Model. Int. J. Mol. Sci. 2018, 19, 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061565
Chen P-C, Chen C-C, Ker Y-B, Chang C-H, Chyau C-C, Hu M-L. Anti-Metastatic Effects of Antrodan with and without Cisplatin on Lewis Lung Carcinomas in a Mouse Xenograft Model. International Journal of Molecular Sciences. 2018; 19(6):1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061565
Chicago/Turabian StyleChen, Pei-Chun, Chin-Chu Chen, Yaw-Bee Ker, Chi-Huang Chang, Charng-Cherng Chyau, and Miao-Lin Hu. 2018. "Anti-Metastatic Effects of Antrodan with and without Cisplatin on Lewis Lung Carcinomas in a Mouse Xenograft Model" International Journal of Molecular Sciences 19, no. 6: 1565. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19061565