Transcriptomics Analysis of the Chinese Pear Pathotype of Alternaria alternata Gives Insights into Novel Mechanisms of HSAF Antifungal Activities

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Collection of the Black Spot Disease Pathogen A. alternata Strains from Infected Pear Leaves in Southern China
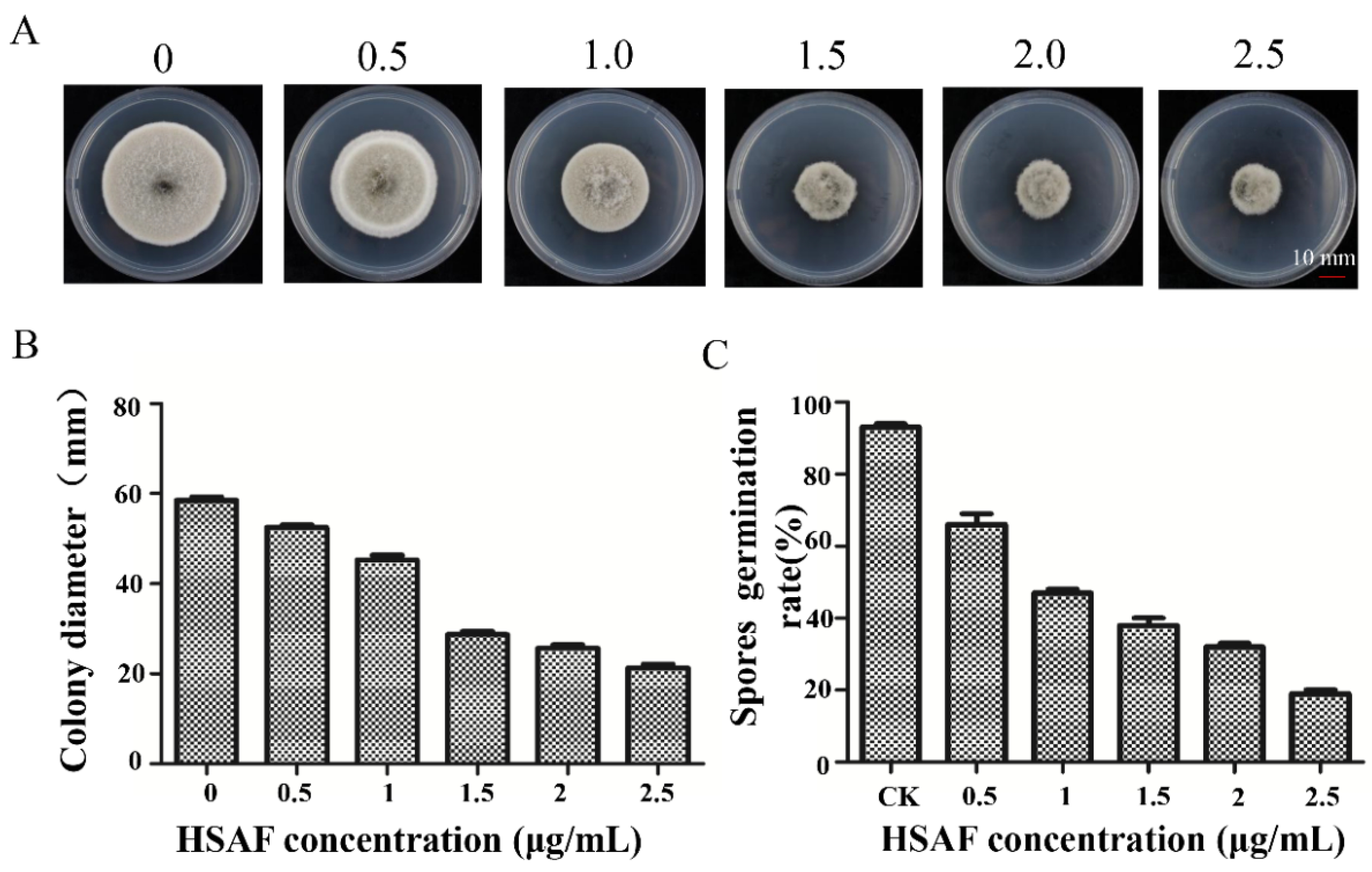
2.2. The Growth Inhibitory Effects of A. alternata by HSAF
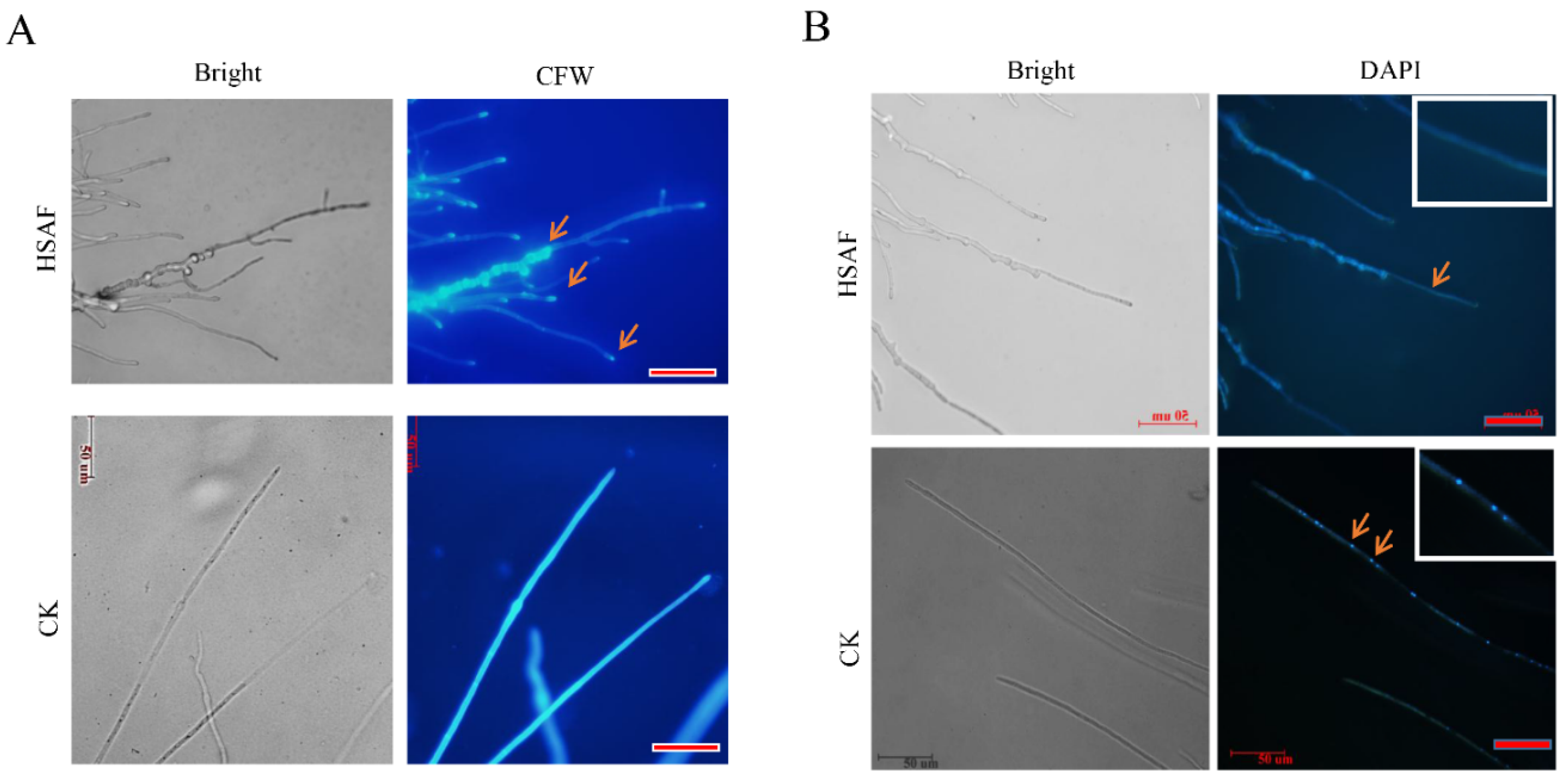
2.3. Cellular Modification of A. alternata against HSAF
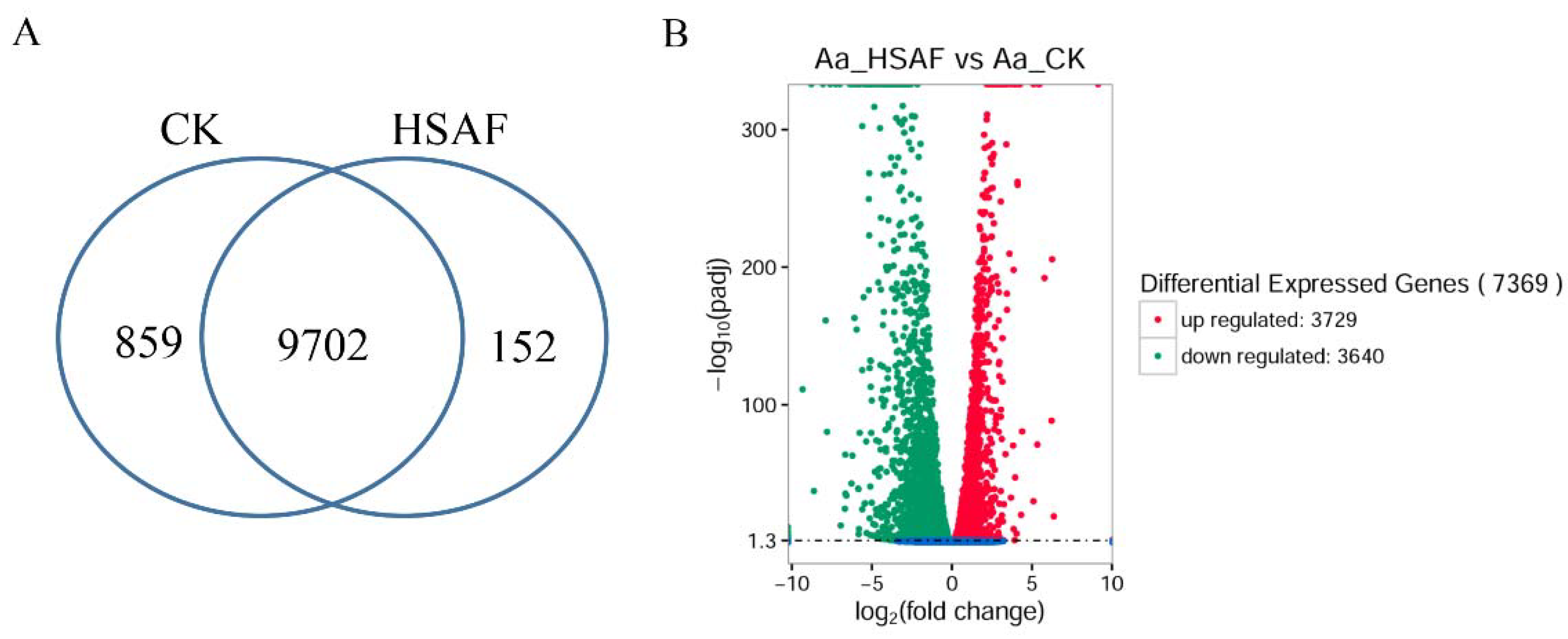
2.4. RNA Sequencing and Novel Genes Prediction
2.5. Differentially Expressed Genes Analysis and Function Annotation
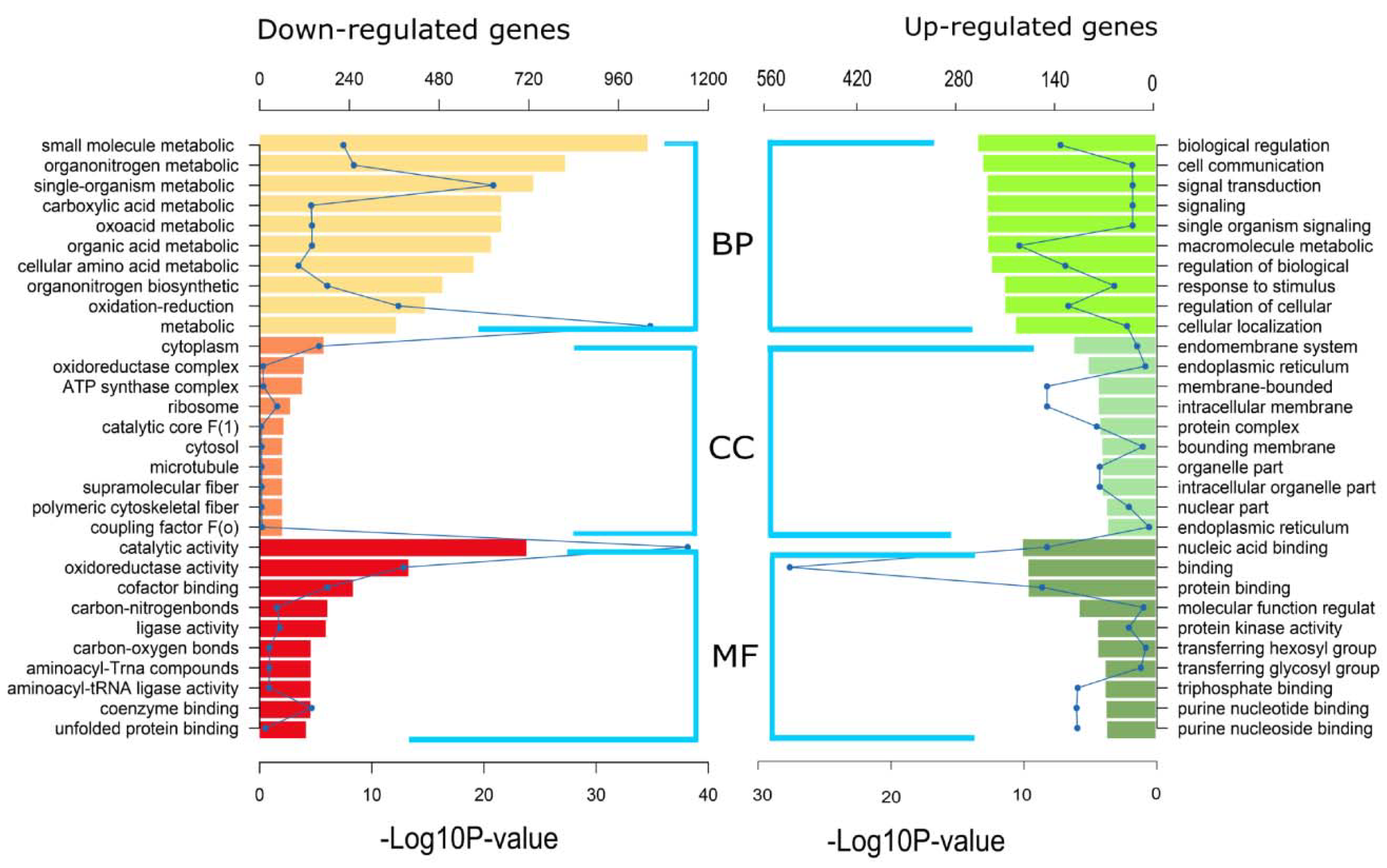
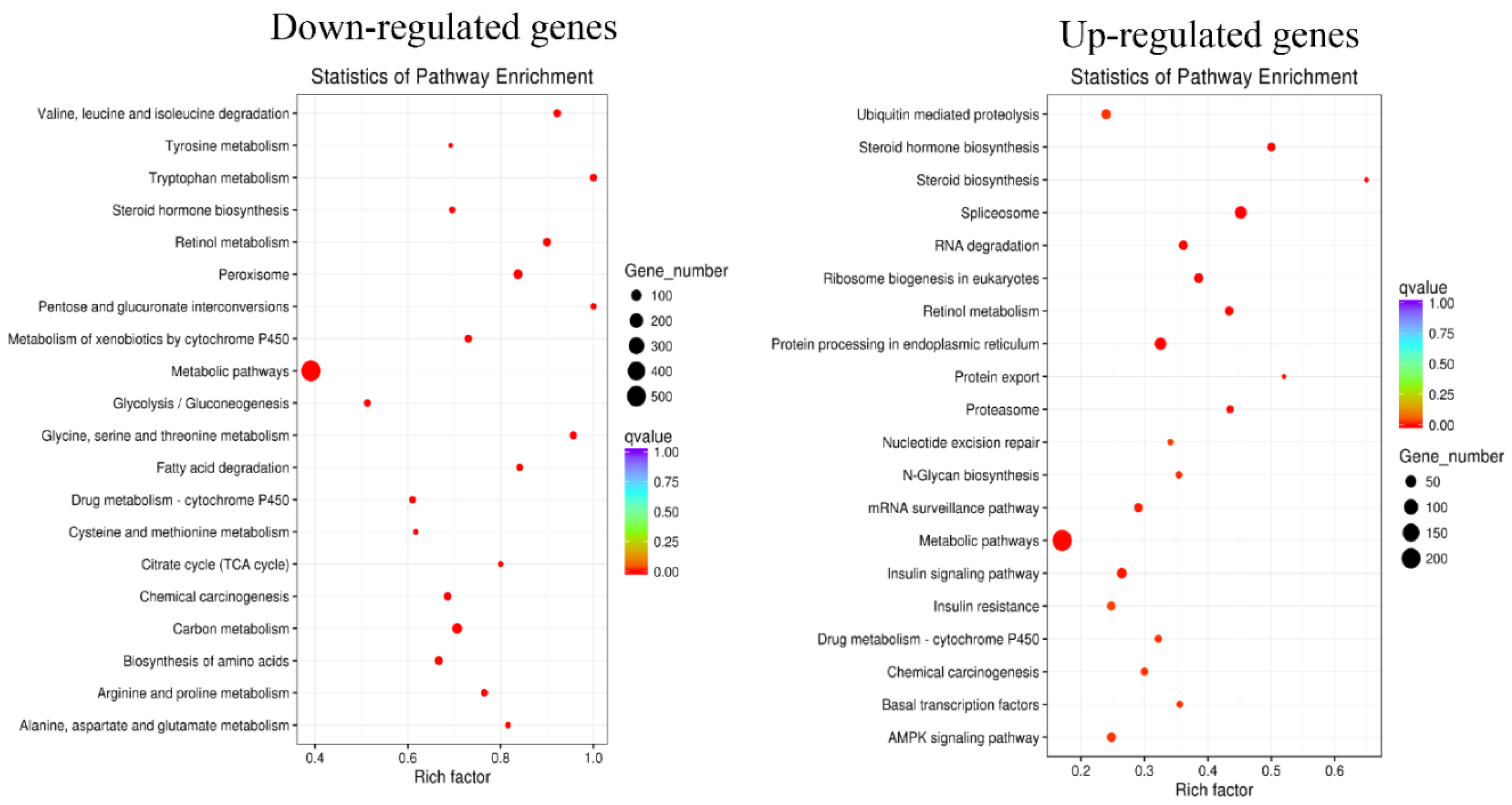
2.6. Gene Ontology (GO) and KEGG Pathway Enrichment of DEGs
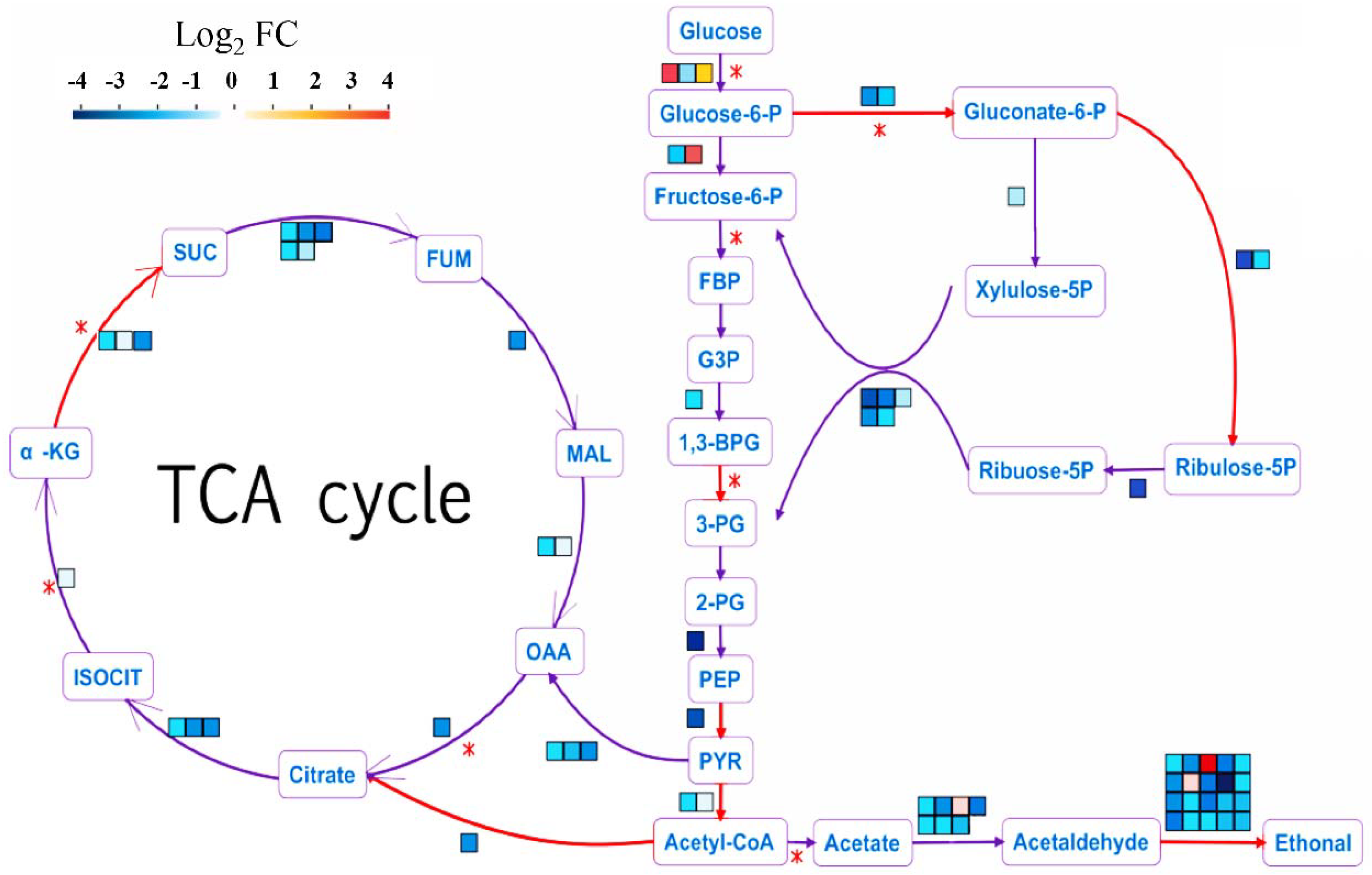
2.7. Genes Involved in Central Carbon Metabolism Responded to HSAF
2.8. Chitin Synthases Encoding Genes Were Predominately Up-Regulated upon HSAF Treatment
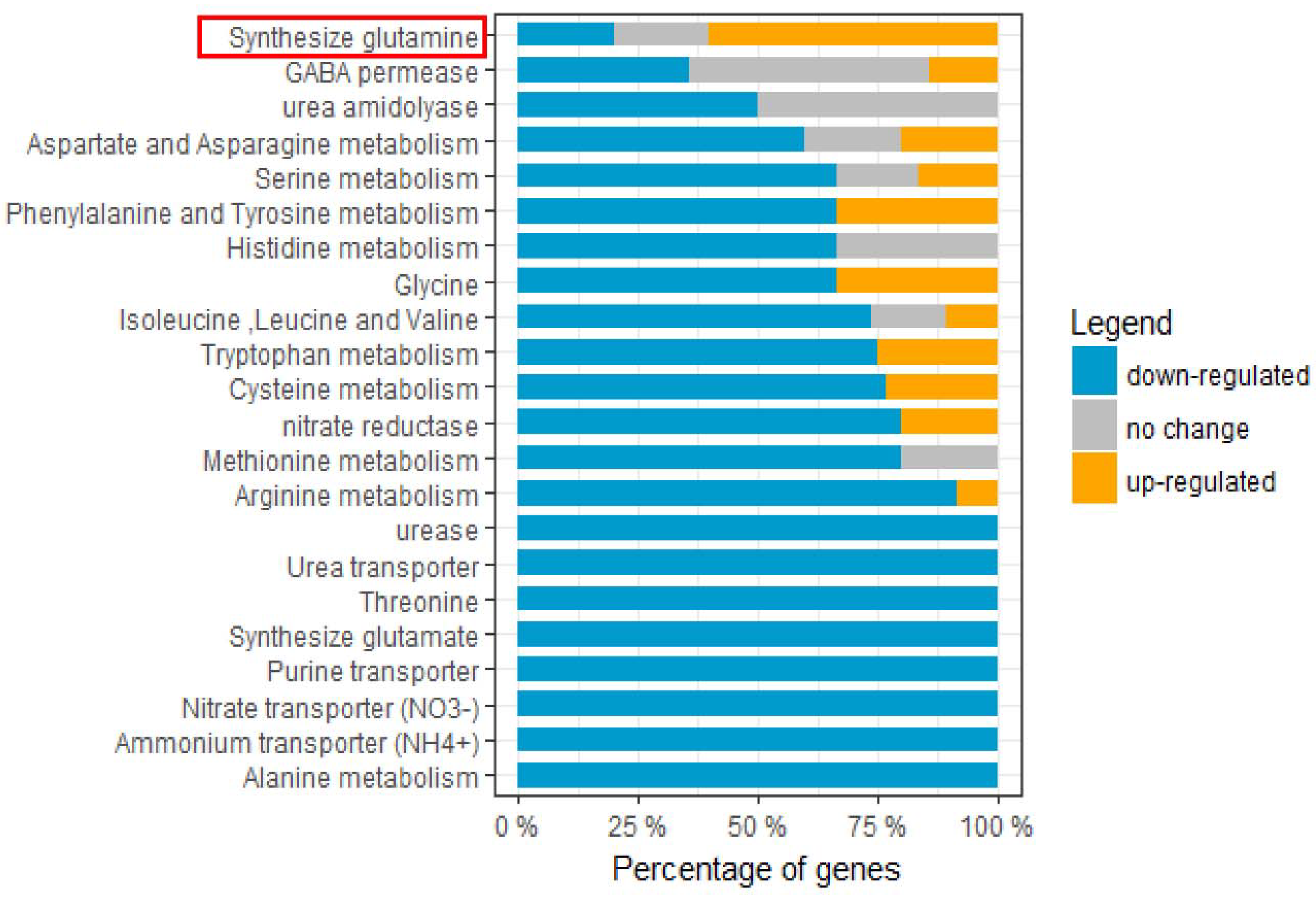
2.9. Nitrogen-Metabolism-Related Genes Were Predominately Down-Regulated upon HSAF Treatment
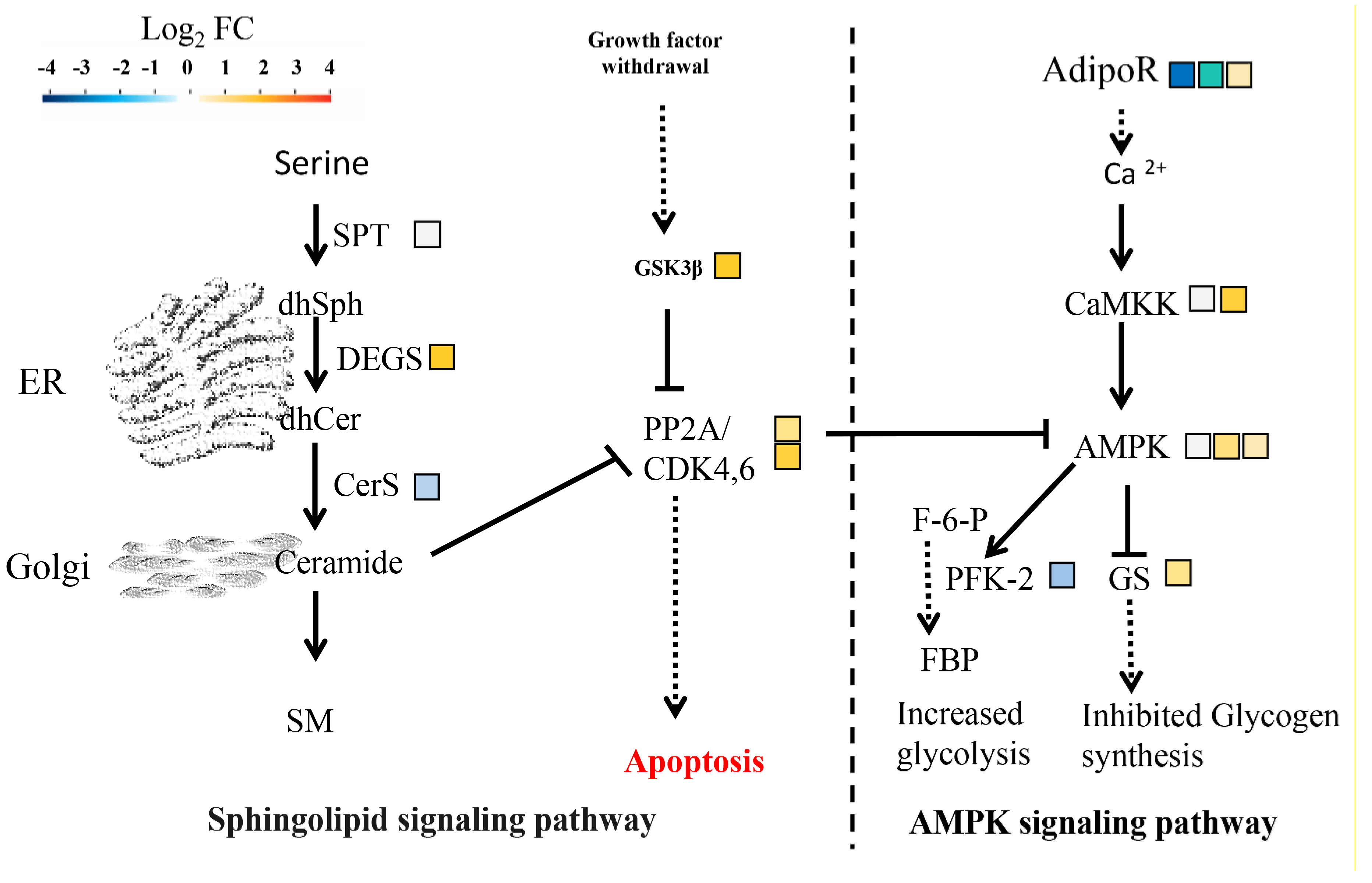
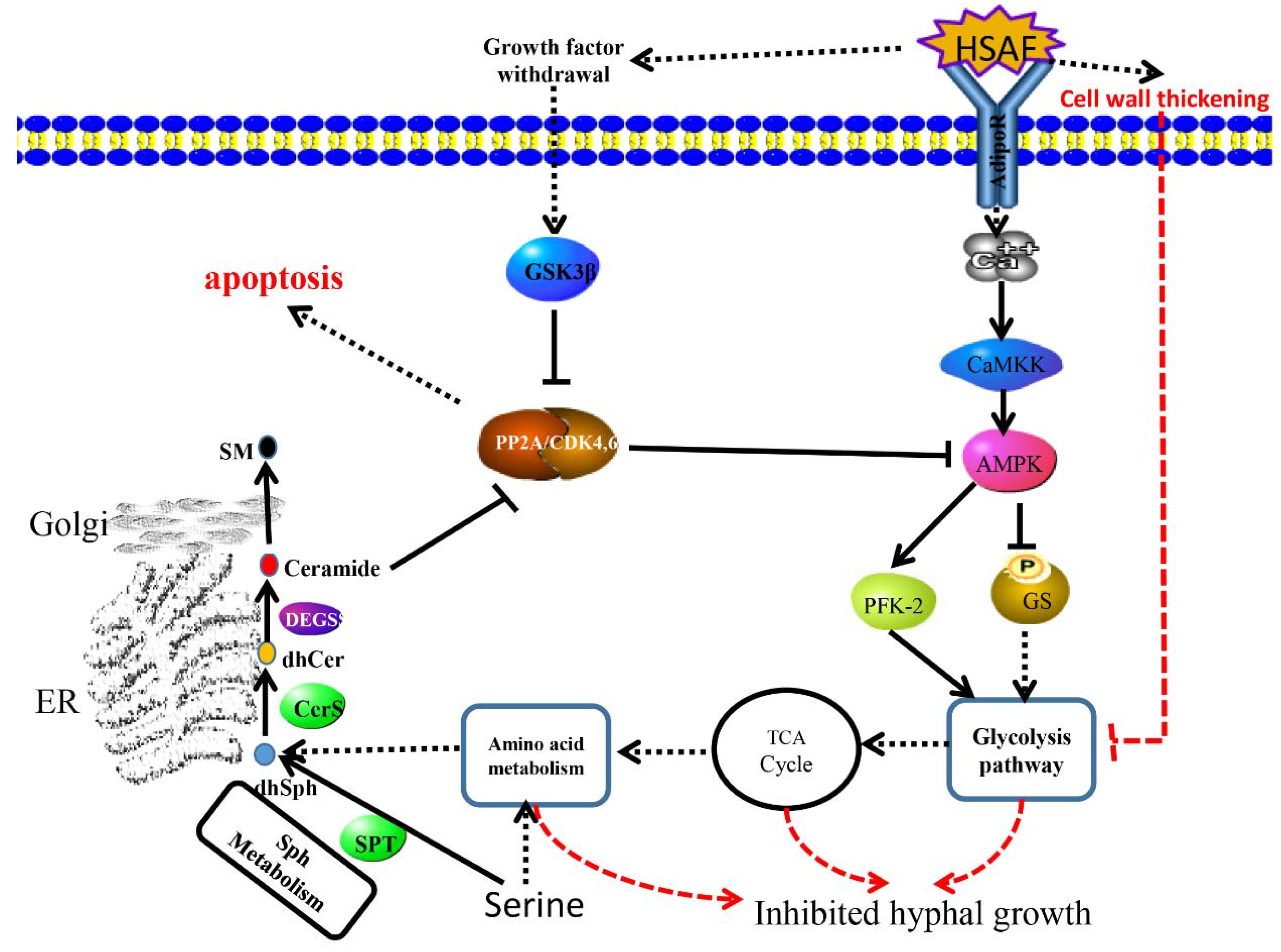
2.10. Signaling Pathway Responding to HSAF
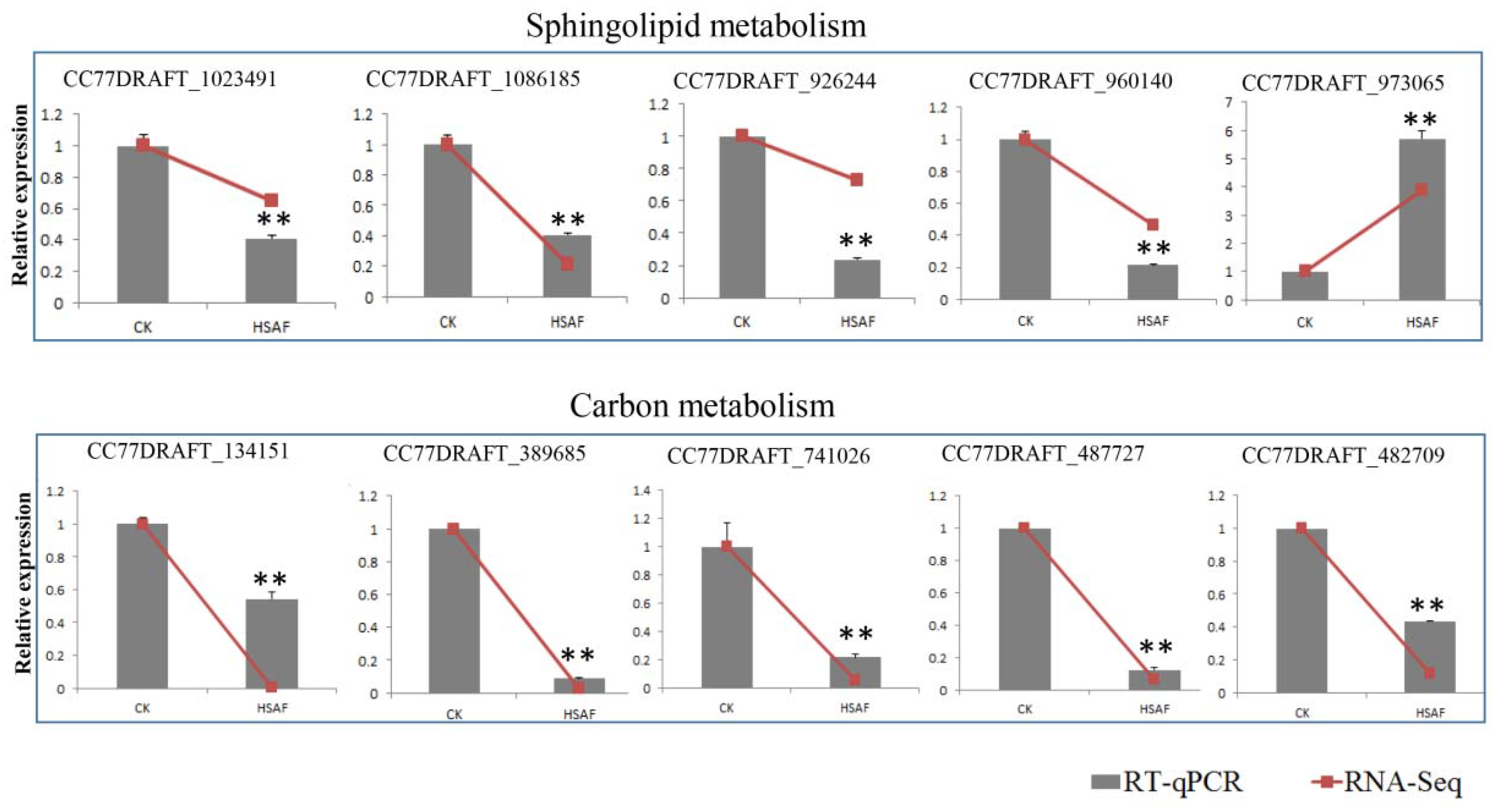
2.11. RT-qPCR Validation of DEGs
3. Discussions
4. Materials and Methods
4.1. HSAF Extraction and Quantification in L. enzymogenes OH11 Cultures
4.2. Measurement of Growth Inhibition of A. alternata by HSAF
4.3. Staining of Mycelia Using Calcofluor White and 4′,6-Diamidino-2-phenylindole
4.4. Sample Preparation for RNA Sequencing
4.5. RT-qPCR Validation
4.6. cDNA Library Preparation and Sequencing
4.7. Bioinformatics Analysis
4.8. Analysis for DEGs
4.9. GO and KEGG Enrichment Analysis
4.10. Prediction of Nitrogen-Metabolism-Related Genes in A. alternata
4.11. Prediction of Chitin Synthase Involving in Cell Wall Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Sullivan, R.F.; Holtman, M.A.; Zylstra, G.J.; White, J.F.; Kobayashi, D.Y. Taxonomic positioning of two biological control agents for plant diseases as Lysobacter enzymogenes based on phylogenetic analysis of 16s rdna, fatty acid composition and phenotypic characteristics. J. Appl. Microbiol. 2003, 94, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Folman, L.B.; Postma, J.; van Veen, J.A. Characterisation of Lysobacter enzymogenes (Christensen and Cook 1978) strain 3.1T8, a powerful antagonist of fungal diseases of cucumber. Microbiol. Res. 2003, 158, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Giesler, L.J.; Yuen, G.Y. Evaluation of Stenotrophomonas maltophilia strain C3 for biocontrol of brown patch disease. Crop Prot. 1998, 17, 509–513. [Google Scholar] [CrossRef]
- Qian, G.L.; Hu, B.S.; Jiang, Y.H.; Liu, F.Q. Identification and characterization of Lysobacter enzymogenes as a biological control agent against some fungal pathogens. Agric. Sci. China 2009, 8, 68–75. [Google Scholar] [CrossRef]
- Chen, J.; Moore, W.H.; Yuen, G.Y.; Kobayashi, D.; Caswell-Chen, E.P. Influence of Lysobacter enzymogenes strain C3 on nematodes. J. Nematol. 2006, 38, 233–239. [Google Scholar] [PubMed]
- Kobayashi, D.Y.; Yuen, G.Y. Molecular mechanisms of interactions between Lysobacter enzymogenes and plant pathogens. Phytopathology 2006, 96, S154. [Google Scholar]
- Yuen, G.; Li, S.; Harris, S.; Yu, F.; Du, L. An antibiotic produced by the biocontrol agent Lysobacter enzymogenes C3 inhibits fungal growth by blocking ceramide synthesis. Phytopathology 2005, 95, S116. [Google Scholar]
- Yuen, G.Y.; Kobayashi, D.K.; Caswell-Chen, E.P. Ecology and biological control of plant pathogens by Lysobacter enzymogenes. Phytopathology 2006, 96, S154. [Google Scholar]
- Zhao, Y.; Qian, G.; Chen, Y.; Du, L.; Liu, F. Transcriptional and antagonistic responses of biocontrol strain Lysobacter enzymogenes OH11 to the plant pathogenic oomycete pythium aphanidermatum. Front. Microbiol. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lou, L.L.; Qian, G.L.; Xie, Y.X.; Hang, J.L.; Chen, H.T.; Zaleta-Riyera, K.; Li, Y.Y.; Shen, Y.M.; Dussault, P.H.; Liu, F.Q.; et al. Biosynthesis of hsaf, a tetramic acid-containing macrolactam from Lysobacter enzymogenes. J. Am. Chem. Soc. 2011, 133, 643–645. [Google Scholar] [CrossRef] [PubMed]
- Jochum, C.C.; Osborne, L.E.; Yuen, G.Y. Fusarium head blight biological control with Lysobacter enzymogenes. Biol. Control 2006, 39, 336–344. [Google Scholar] [CrossRef]
- Mathioni, S.M.; Patel, N.; Riddick, B.; Sweigard, J.A.; Czymmek, K.J.; Caplan, J.L.; Kunjeti, S.G.; Kunjeti, S.; Raman, V.; Hillman, B.I.; et al. Transcriptomics of the rice blast fungus magnaporthe oryzae in response to the bacterial antagonist Lysobacter enzymogenes reveals candidate fungal defense response genes. PLoS ONE 2013, 8, e76487. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Calvo, A.M.; Yuen, G.Y.; Du, L.; Harris, S.D. Induction of cell wall thickening by the antifungal compound dihydromaltophilin disrupts fungal growth and is mediated by sphingolipid biosynthesis. J. Eukaryot. Microbiol. 2009, 56, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.J.; Li, Z.Y.; Li, Y.Y.; Lu, C.H.; Wang, H.X.; Shen, Y.M.; Du, L.C. HSAF-induced antifungal effects in Candida albicans through ROS-mediated apoptosis. RSC Adv. 2016, 6, 30895–30904. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.Y.; Wang, R.P.; Zhao, Y.Y.; Fu, Z.Q.; Qian, G.L.; Liu, F.Q. LesR is a novel upstream regulator that controls downstream Clp expression to modulate antibiotic HSAF biosynthesis and cell aggregation in Lysobacter enzymogenes OH11. Microb. Cell Fact. 2017, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.P.; Xu, H.Y.; Du, L.C.; Chou, S.H.; Liu, H.X.; Liu, Y.Z.; Liu, F.Q.; Qian, G.L. A TonB-dependent receptor regulates antifungal HSAF biosynthesis in Lysobacter. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Olson, A.S.; Chen, H.T.; Du, L.C.; Dussault, P.H. Synthesis of a 2,4,6,8,10-dodecapentanoic acid thioester as a substrate for biosynthesis of heat stable antifungal factor (HSAF). RSC Adv. 2015, 5, 11644–11648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.Y.; Huffman, J.; Li, Y.; Du, L.C.; Shen, Y.M. 3-Hydroxylation of the polycyclic tetramate macrolactam in the biosynthesis of antifungal hsaf from Lysobacter enzymogenes C3. MedChemComm 2012, 3, 982–986. [Google Scholar] [CrossRef]
- Han, Y.; Wang, Y.; Tombosa, S.; Wright, S.; Huffman, J.; Yuen, G.; Qian, G.L.; Liu, F.Q.; Shen, Y.M.; Du, L.C. Identification of a small molecule signaling factor that regulates the biosynthesis of the antifungal polycyclic tetramate macrolactam HSAF in Lysobacter enzymogenes. Appl. Microbiol. Biotechnol. 2015, 99, 801–811. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, M.; Nakano, T.; Akamatsu, H.; Kodama, M.; Otani, H.; Osaki-Oka, K. Alternaria alternata apple pathotype (A. mali) causes black spot of European pear. Eur. J. Plant Pathol. 2016, 145, 787–795. [Google Scholar] [CrossRef]
- Duan, S.; Ma, X.; Chen, W.; Wan, W.; He, Y.; Ma, X.; Ma, Y.; Long, N.; Tan, Y.; Wang, Y.; et al. Transcriptomic profile of tobacco in response to Alternaria longipes and Alternaria alternata infections. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Kawakita, K.; Tsuge, T.; Doke, N. Stimulation of phospholipase-A2 activity in strawberry cells treated with AF-toxin-I produced by Alternaria alternata strawberry pathotype. Physiol. Mol. Plant Pathol. 1992, 41, 283–294. [Google Scholar] [CrossRef]
- Tsuge, T.; Kobayashi, H.; Nishimura, S. Organization of ribosomal RNA genes in Alternaria alternata japanese pear pathotype, a host-selective AK-toxin-producing fungus. Curr. Genet. 1989, 16, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Armitage, A.D.; Barbara, D.J.; Harrison, R.J.; Lane, C.R.; Sreenivasaprasad, S.; Woodhall, J.W.; Clarkson, J.P. Discrete lineages within Alternaria alternata species group: Identification using new highly variable loci and support from morphological characters. Fungal Biol. 2015, 119, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Zeiner, C.A.; Purvine, S.O.; Zink, E.M.; Pasa-Tolic, L.; Chaput, D.L.; Haridas, S.; Wu, S.; LaButti, K.; Grigoriev, I.V.; Henrissat, B.; et al. Comparative analysis of secretome profiles of manganese(II)-oxidizing ascomycete fungi. PLoS ONE 2016, 11, e0157844. [Google Scholar] [CrossRef] [PubMed]
- Li, K.B. Clustalw-mpi: Clustalw analysis using distributed and parallel computing. Bioinformatics 2003, 19, 1585–1586. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. Mega5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- McCann, M.P.; Snetselaar, K.M. A genome-based analysis of amino acid metabolism in the biotrophic plant pathogen Ustilago maydis. Fungal Genet. Biol. 2008, 45 (Suppl. 1), S77–S87. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Du, L.; Yuen, G.; Harris, S.D. Distinct ceramide synthases regulate polarized growth in the filamentous fungus Aspergillus nidulans. Mol. Biol. Cell 2006, 17, 1218–1227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Shen, Y. Antifungal compound honokiol triggers oxidative stress responsive signalling pathway and modulates central carbon metabolism. Mycology 2016, 7, 1–10. [Google Scholar] [CrossRef]
- Zhao, Y.; Paderu, P.; Park, S.; Dukhan, A.; Senter, M.; Perlin, D.S. Expression turnover profiling to monitor the antifungal activities of amphotericin B, voriconazole, and micafungin against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2012, 56, 2770–2772. [Google Scholar] [CrossRef] [PubMed]
- Marioni, J.C.; Mason, C.E.; Mane, S.M.; Stephens, M.; Gilad, Y. Rna-seq: An assessment of technical reproducibility and comparison with gene expression arrays. Genome Res. 2008, 18, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Sun, X.; Yu, D.; Xu, J.; Chung, K.; Li, H. Genomic and transcriptomic analyses of the tangerine pathotype of Alternaria alternata in response to oxidative stress. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Muller, C.; Hjort, C.M.; Hansen, K.; Nielsen, J. Altering the expression of two chitin synthase genes differentially affects the growth and morphology of Aspergillus oryzae. Microbiology 2002, 148, 4025–4033. [Google Scholar] [CrossRef] [PubMed]
- Feofilova, E.P. The fungal cell wall: Modern concepts of its composition and biological function. Mikrobiologiia 2010, 79, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Horiuchi, H.; Fujiwara, M.; Yamashita, S.; Ohta, A.; Takagi, M. Proliferation of intrahyphal hyphae caused by disruption of csmA, which encodes a class V chitin synthase with a myosin motor-like domain in Aspergillus nidulans. J. Bacteriol. 1999, 181, 3721–3729. [Google Scholar] [PubMed]
- Liu, J.H.; Tang, X.; Wang, H.Y.; Balasubramanian, M. Bgs2p, a 1,3-beta-glucan synthase subunit, is essential for maturation of ascospore wall in Schizosaccharomyces pombe. FEBS Lett. 2000, 478, 105–108. [Google Scholar] [CrossRef]
- Lagorce, A.; Le Berre-Anton, V.; Aguilar-Uscanga, B.; Martin-Yken, H.; Dagkessamanskaia, A.; Francois, J. Involvement of GFA1, which encodes glutamine-fructose-6-phosphate amidotransferase, in the activation of the chitin synthesis pathway in response to cell-wall defects in Saccharomyces cerevisiae. FEBS J. 2002, 269, 1697–1707, Erratum in 2006, 273, 3869. [Google Scholar]
- Garcia, A.B.; Vinuela-Prieto, J.M.; Lopez-Gonzalez, L.; Candel, F.J. Correlation between resistance mechanisms in Staphylococcus aureus and cell wall and septum thickening. Infect. Drug Resist. 2017, 10, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Song, D.; Sun, J.; Li, L. Populus endo-beta-mannanase ptrman6 plays a role in coordinating cell wall remodeling with suppression of secondary wall thickening through generation of oligosaccharide signals. Plant J. Cell Mol. Biol. 2013, 74, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Hyo, Y.; Yamada, S.; Fukutsuji, K.; Harada, T. Thickening of the cell wall in macrolide-resistant Staphylococcus aureus. Med. Mol. Morphol. 2013, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Zhang, X.; Mafurah, J.J.; Zhang, M.; Qian, G.; Wang, R.; Safdar, A.; Yang, X.; Liu, F.; Dou, D. The transcription factor vpcrz1 is required for fruiting body formation and pathogenicity in Valsa pyri. Microb. Pathog. 2016, 95, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Zhang, Y.L.; Fan, S.R.; Nobile, C.J.; Guan, G.B.; Huang, G.H. Integration of the tricarboxylic acid (TCA) cycle with camp signaling and Sfl2 pathways in the regulation of CO2 sensing and hyphal development in Candida albicans. PLos Genet. 2017, 13, e1006949. [Google Scholar] [CrossRef] [PubMed]
- Paumen, M.B.; Ishida, Y.; Muramatsu, M.; Yamamoto, M.; Honjo, T. Inhibition of carnitine palmitoyltransferase I augments sphingolipid synthesis and palmitate-induced apoptosis. J. Biol. Chem. 1997, 272, 3324–3329. [Google Scholar] [CrossRef] [PubMed]
- Krown, K.A.; Page, M.T.; Nguyen, C.; Zechner, D.; Gutierrez, V.; Comstock, K.L.; Glembotski, C.C.; Quintana, P.J.; Sabbadini, R.A. Tumor necrosis factor alpha-induced apoptosis in cardiac myocytes. Involvement of the sphingolipid signaling cascade in cardiac cell death. J. Clin. Investig. 1996, 98, 2854–2865. [Google Scholar] [CrossRef] [PubMed]
- Young, M.M.; Kester, M.; Wang, H.G. Sphingolipids: Regulators of crosstalk between apoptosis and autophagy. J. Lipid Res. 2013, 54, 5–19. [Google Scholar] [CrossRef] [PubMed]
- Yamaji-Hasegawa, A.; Takahashi, A.; Tetsuka, Y.; Senoh, Y.; Kobayashi, T. Fungal metabolite sulfamisterin suppresses sphingolipid synthesis through inhibition of serine palmitoyltransferase. Biochemistry 2005, 44, 268–277. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.; Gustafsson, H.T.; O’Sullivan, C.; Bisceglia, G.; Huang, X.H.; Klose, C.; Schevchenko, A.; Dickson, R.C.; Cavaliere, P.; Dephoure, N.; et al. Serine-dependent sphingolipid synthesis is a metabolic liability of aneuploid cells. Cell Rep. 2017, 21, 3807–3818. [Google Scholar] [CrossRef] [PubMed]
- Freitas, F.Z.; de Paula, R.M.; Barbosa, L.C.; Terenzi, H.F.; Bertolini, M.C. Camp signaling pathway controls glycogen metabolism in neurospora crassa by regulating the glycogen synthase gene expression and phosphorylation. Fungal Genet. Biol. 2010, 47, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Altiok, S.; Xu, M.; Spiegelman, B.M. Ppargamma induces cell cycle withdrawal: Inhibition of E2F/DP DNA-binding activity via down-regulation of PP2A. Genes Dev. 1997, 11, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Ruvolo, P.P.; Deng, X.; Ito, T.; Carr, B.K.; May, W.S. Ceramide induces Bcl2 dephosphorylation via a mechanism involving mitochondrial PP2A. J. Biol. Chem. 1999, 274, 20296–20300. [Google Scholar] [CrossRef] [PubMed]
- Kurimchak, A.; Grana, X. PP2A holoenzymes negatively and positively regulate cell cycle progression by dephosphorylating pocket proteins and multiple CDK substrates. Gene 2012, 499, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cammisotto, P.G.; Londono, I.; Gingras, D.; Bendayan, M. Control of glycogen synthase through ADIPOR1-AMPK pathway in renal distal tubules of normal and diabetic rats. Am. J. Phys.-Renal Physiol. 2008, 294, F881–F889. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Youn, B.; Zheng, X.L.; Wu, D.; Xu, A.; Sweeney, G. Globular adiponectin, acting via AdipoR1/APPL1, protects H9c2 cells from hypoxia/reoxygenation-induced apoptosis. PLoS ONE 2011, 6, e19143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.; Choi, S.M.; Whitcomb, D.J.; Kim, B.C. Adiponectin controls the apoptosis and the expression of tight junction proteins in brain endothelial cells through AdipoR1 under beta amyloid toxicity. Cell Death Dis. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.Q.; Yang, Y.J.; Li, Q.; Huang, P.S.; Li, X.D.; Jin, C.; Qi, K.; Jiang, L.P.; Chen, G.H. Globular adiponectin inhibits the apoptosis of mesenchymal stem cells induced by hypoxia and serum deprivation via the AdipoR1-mediated pathway. Cell. Physiol. Biochem. 2016, 38, 909–925. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Z.; Liang, B.; Lau, W.B.; Wang, Y.; Zhao, J.; Li, R.; Wang, X.; Yuan, Y.; Lopez, B.L.; Christopher, T.A.; et al. High glucose/high lipids impair vascular adiponectin function via inhibition of caveolin-1/AdipoR1 signalsome formation. Free Radic. Biol. Med. 2015, 89, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, M.; Kitayama, J.; Yamauchi, T.; Kadowaki, T.; Maki, T.; Miyato, H.; Yamashita, H.; Nagawa, H. Adiponectin inhibits the growth and peritoneal metastasis of gastric cancer through its specific membrane receptors AdipoR1 and AdipoR2. Cancer Sci. 2007, 98, 1120–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, G.; Wang, Y.; Liu, Y.; Xu, F.; He, Y.W.; Du, L.; Venturi, V.; Fan, J.; Hu, B.; Liu, F. Lysobacter enzymogenes uses two distinct cell-cell signaling systems for differential regulation of secondary-metabolite biosynthesis and colony morphology. Appl. Environ. Microbiol. 2013, 79, 6604–6616. [Google Scholar] [CrossRef] [PubMed]
- Takaoka, S.; Kurata, M.; Harimoto, Y.; Hatta, R.; Yamamoto, M.; Akimitsu, K.; Tsuge, T. Complex regulation of secondary metabolism controlling pathogenicity in the phytopathogenic fungus Alternaria alternata. New Phytol. 2014, 202, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. Hmmer web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level—The DESeq package. EMBL 2013, 1–32. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-Seq: Accounting for selection bias. Genome Biol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schaffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped blast and psi-blast: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]











© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, F.; Li, B.; Ai, G.; Kange, A.M.; Zhao, Y.; Zhang, X.; Jia, Y.; Dou, D.; Liu, F.; Cao, H. Transcriptomics Analysis of the Chinese Pear Pathotype of Alternaria alternata Gives Insights into Novel Mechanisms of HSAF Antifungal Activities. Int. J. Mol. Sci. 2018, 19, 1841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071841
He F, Li B, Ai G, Kange AM, Zhao Y, Zhang X, Jia Y, Dou D, Liu F, Cao H. Transcriptomics Analysis of the Chinese Pear Pathotype of Alternaria alternata Gives Insights into Novel Mechanisms of HSAF Antifungal Activities. International Journal of Molecular Sciences. 2018; 19(7):1841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071841
Chicago/Turabian StyleHe, Feng, Bingxin Li, Gan Ai, Alex Machio Kange, Yancun Zhao, Xiong Zhang, Yifan Jia, Daolong Dou, Fengquan Liu, and Haiqun Cao. 2018. "Transcriptomics Analysis of the Chinese Pear Pathotype of Alternaria alternata Gives Insights into Novel Mechanisms of HSAF Antifungal Activities" International Journal of Molecular Sciences 19, no. 7: 1841. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19071841