Vitamin D’s Effect on the Proliferation and Inflammation of Human Intervertebral Disc Cells in Relation to the Functional Vitamin D Receptor Gene FokI Polymorphism

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
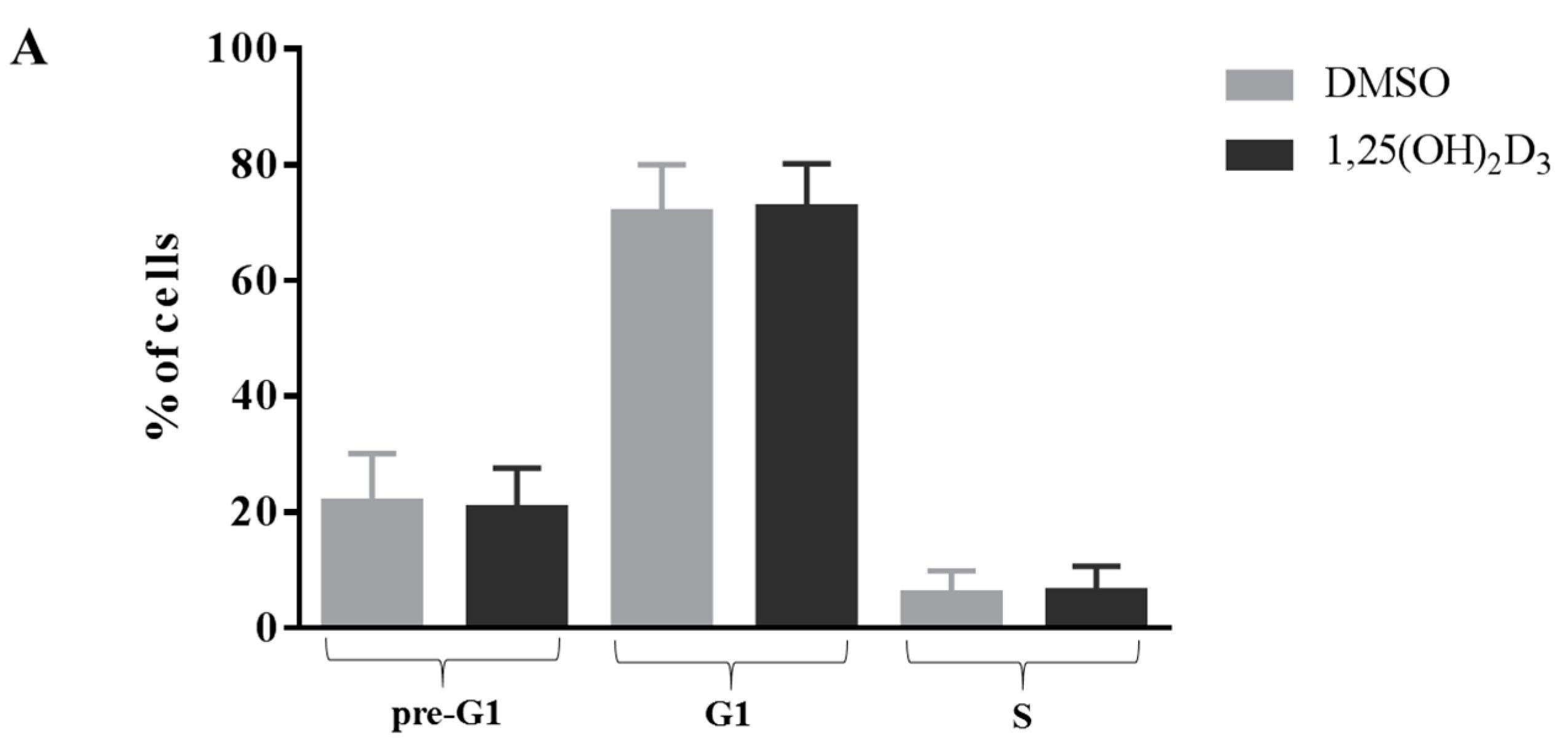
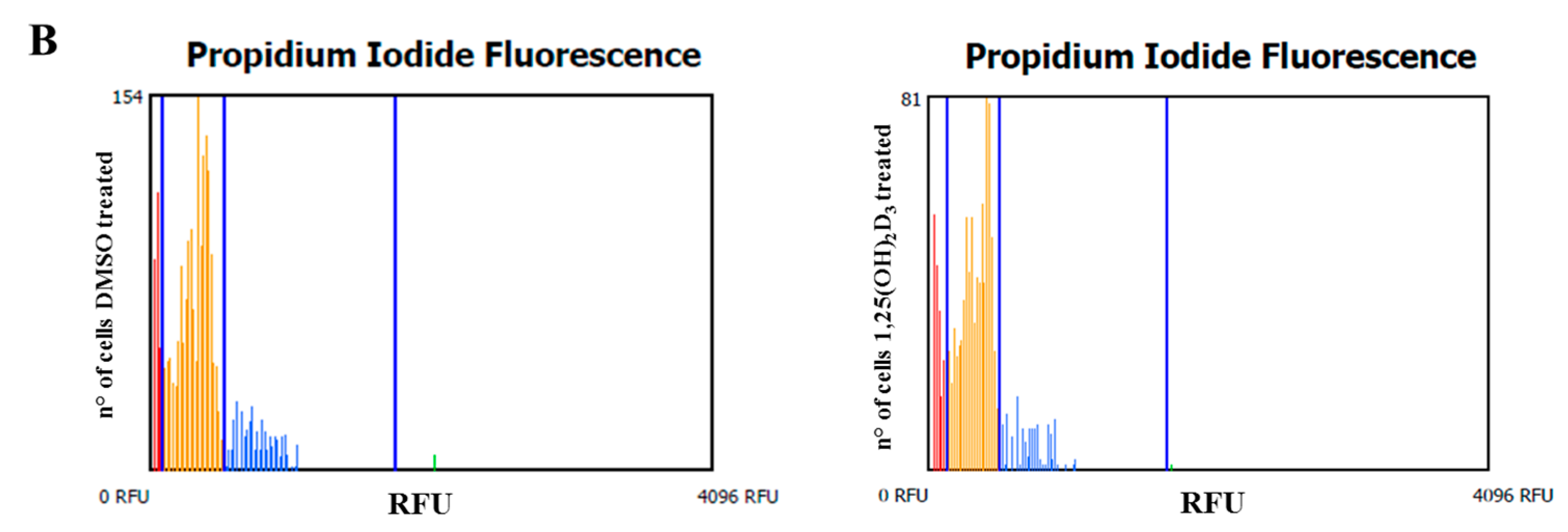
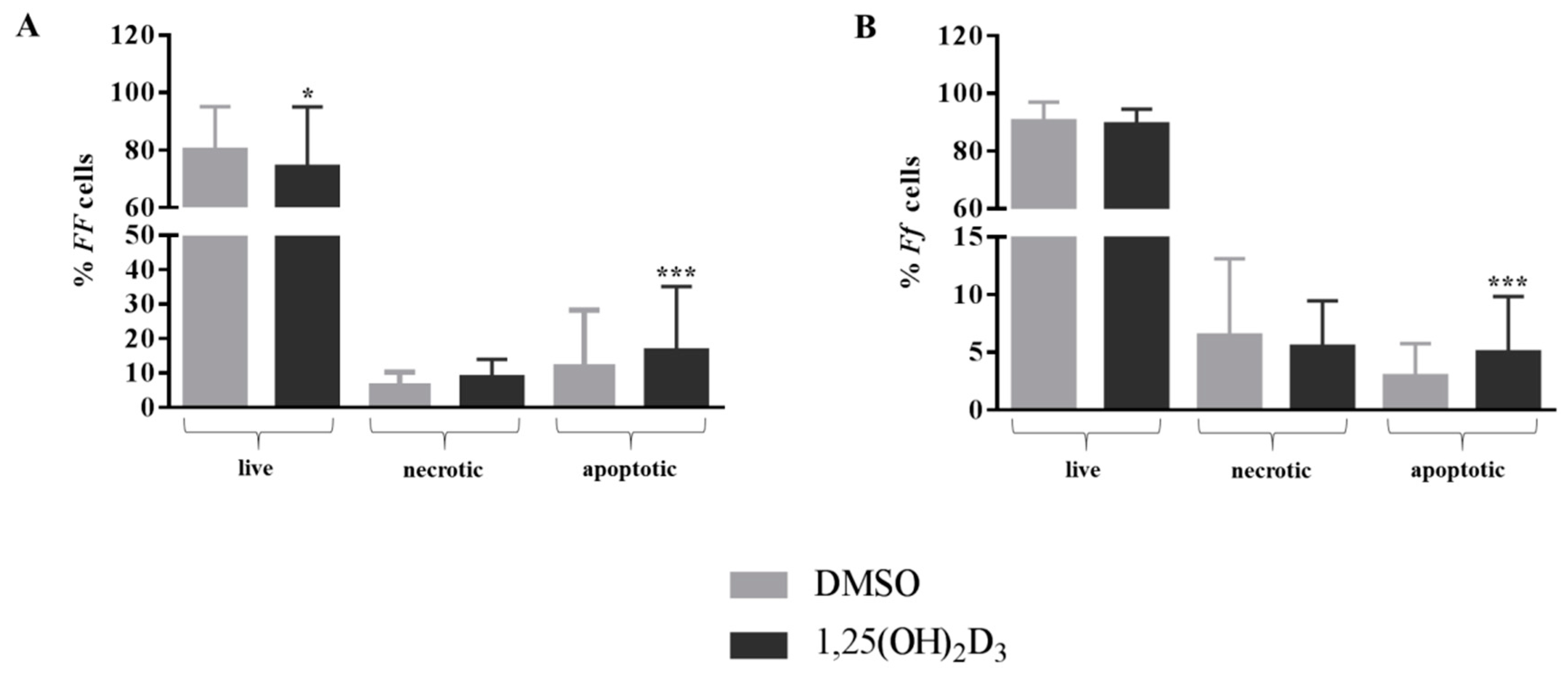
2.1. Anti-Proliferative Effect of Vitamin D Is Related to Induction of Apoptosis
2.2. Effect of IL-1β on Disc Cells Cultured in Absence of Vitamin D
2.3. Vitamin D Supplementation Did Not Influence the Release of IL-1β in Disc Cells
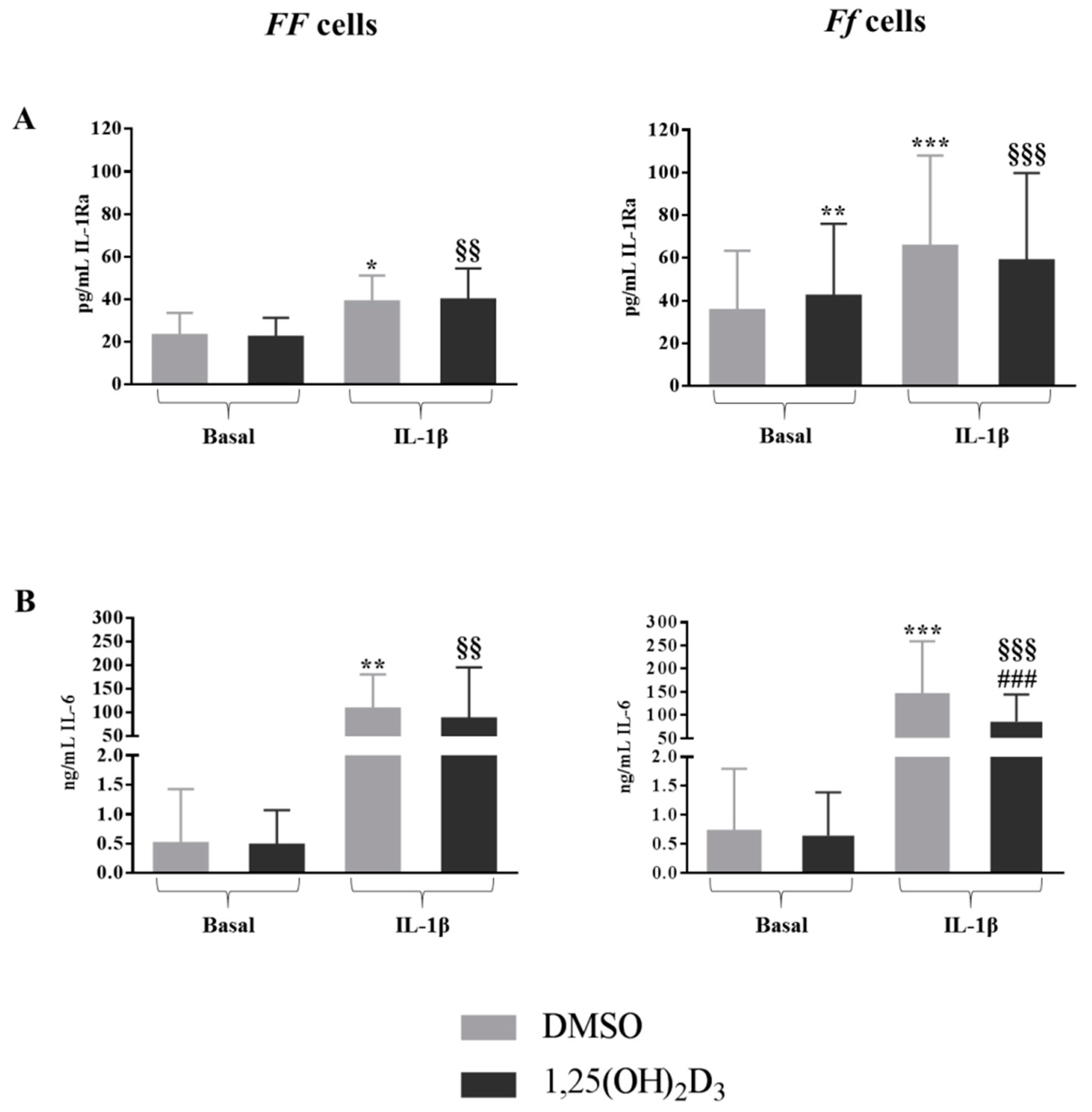
2.4. Vitamin D Upregulated IL-1Ra Release in Basal Condition and Downregulated IL-6 Release in Inflamed Condition in Cells Bearing the FokI VDR Ff Genotype
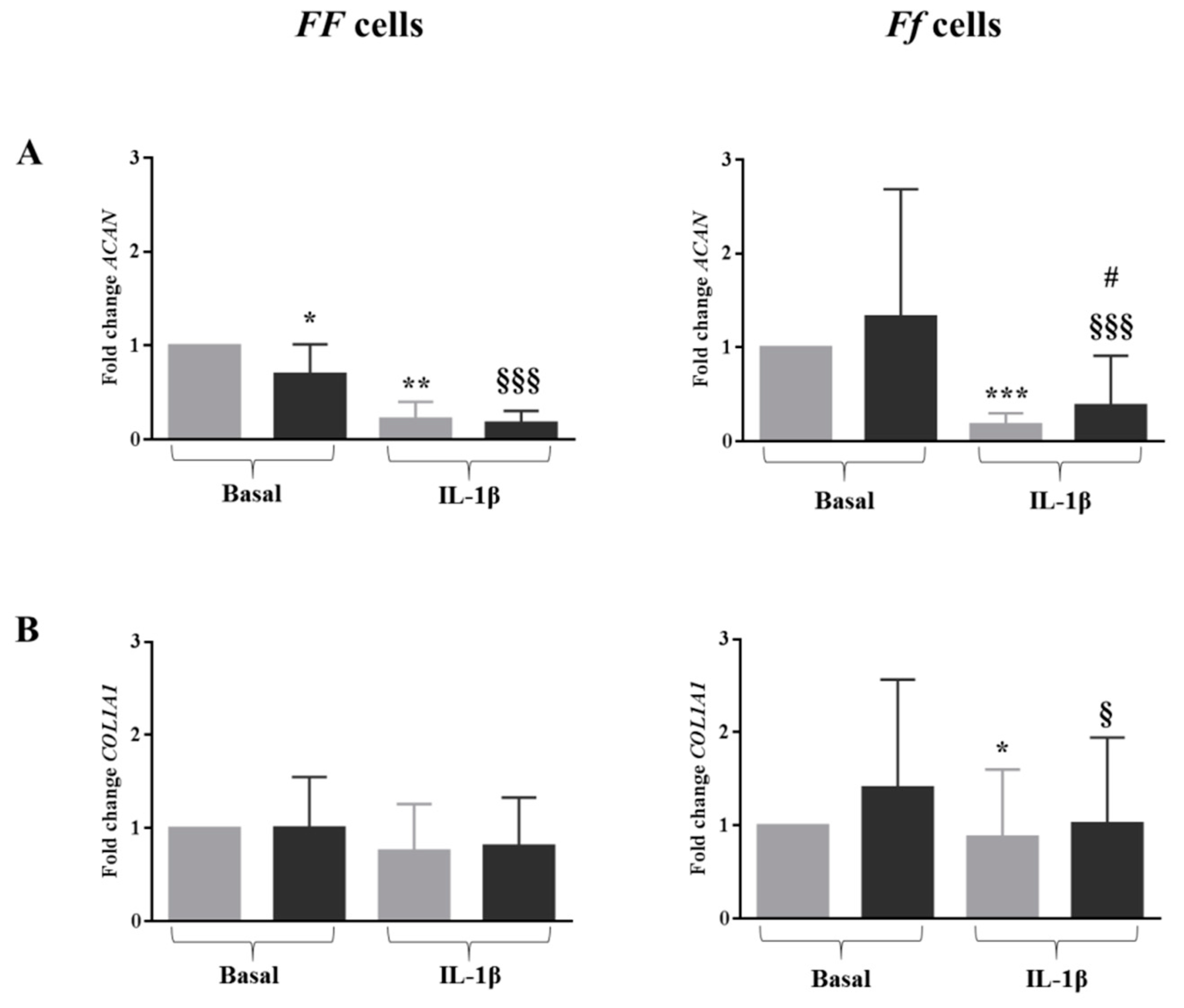
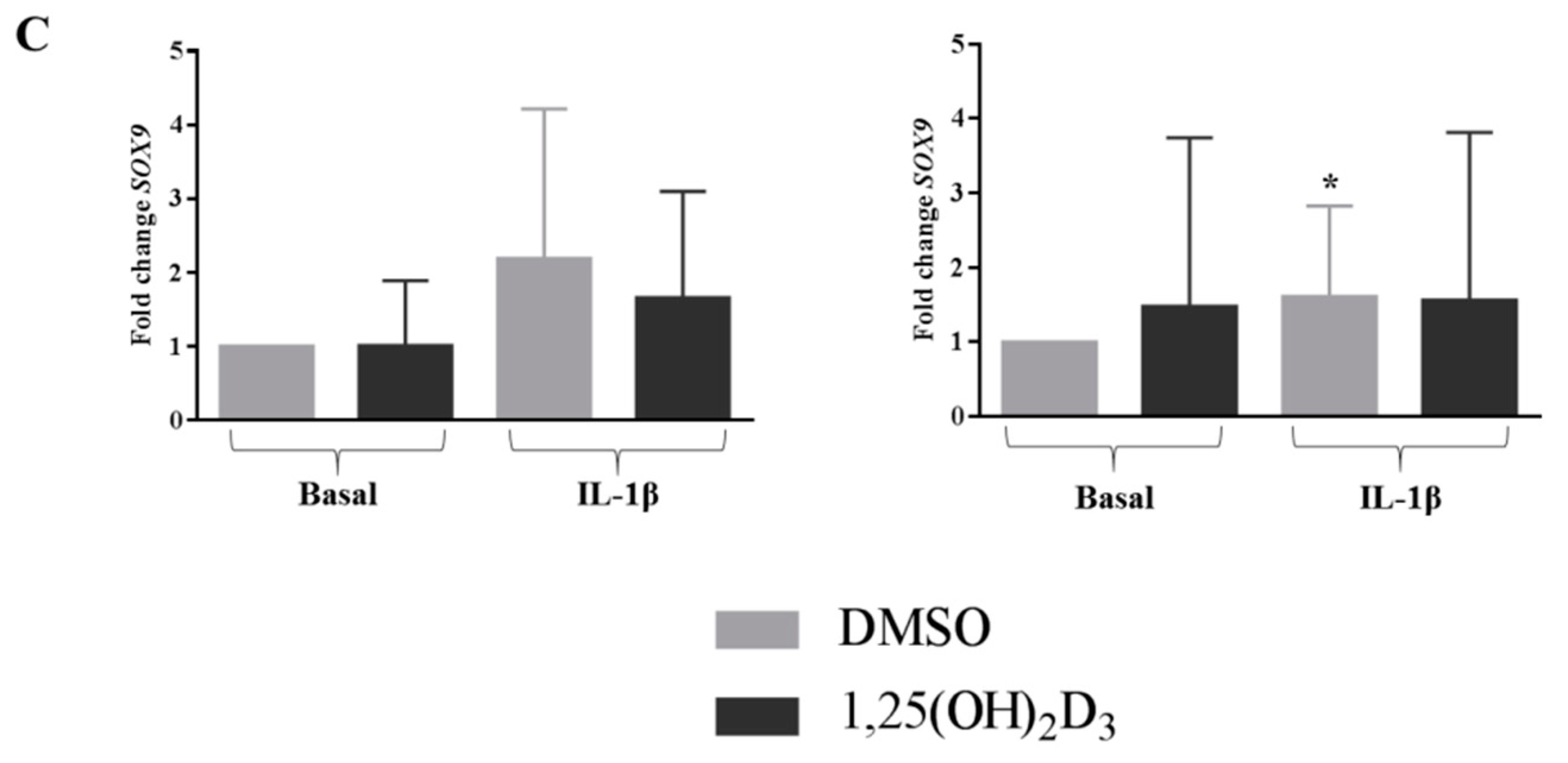
2.5. In Inflamed Condition Vitamin D Upregulated the Expression of ACAN in Ff Disc Cells, But Did Not Have an Effect on SOX9 and COL1A1 Expression
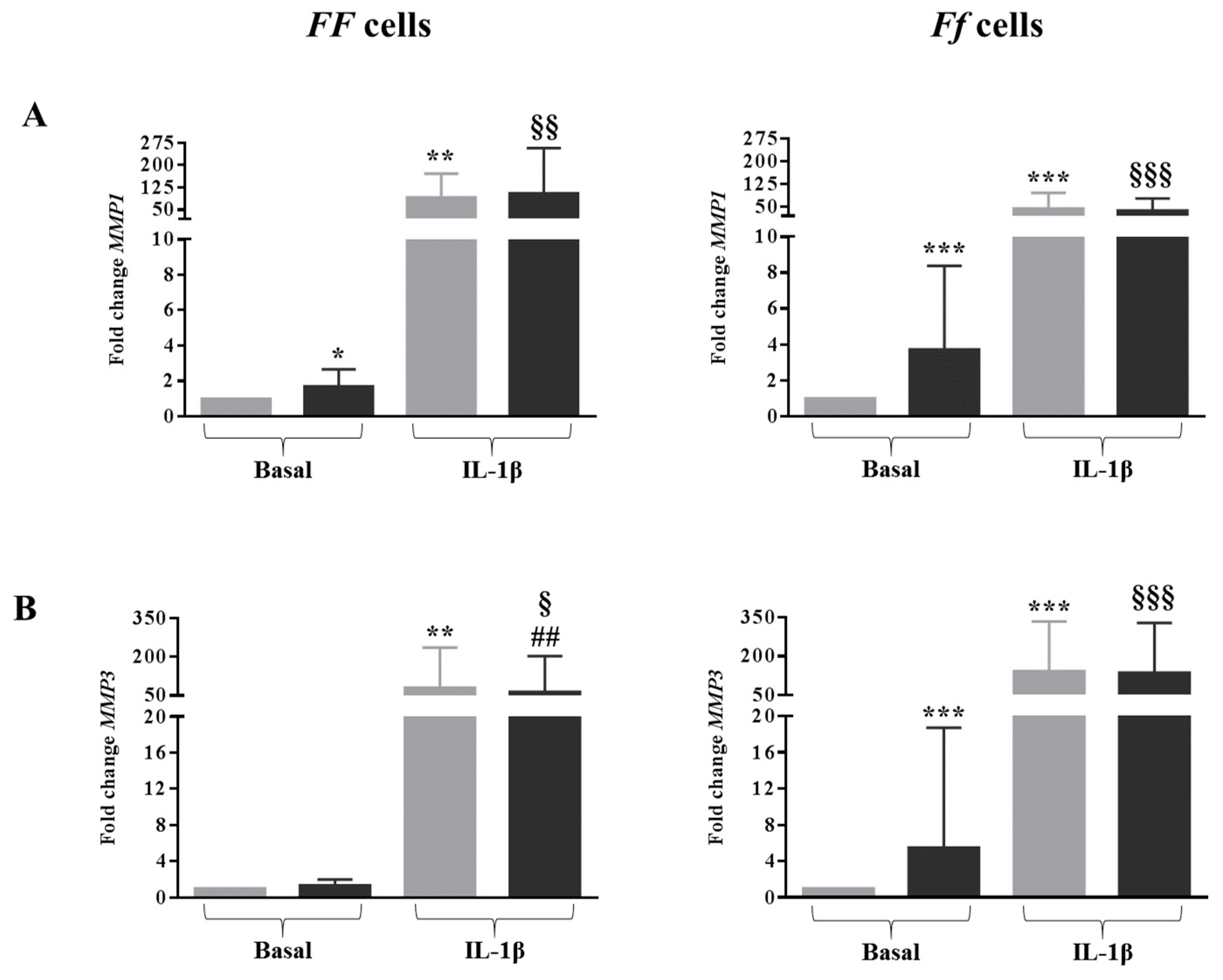
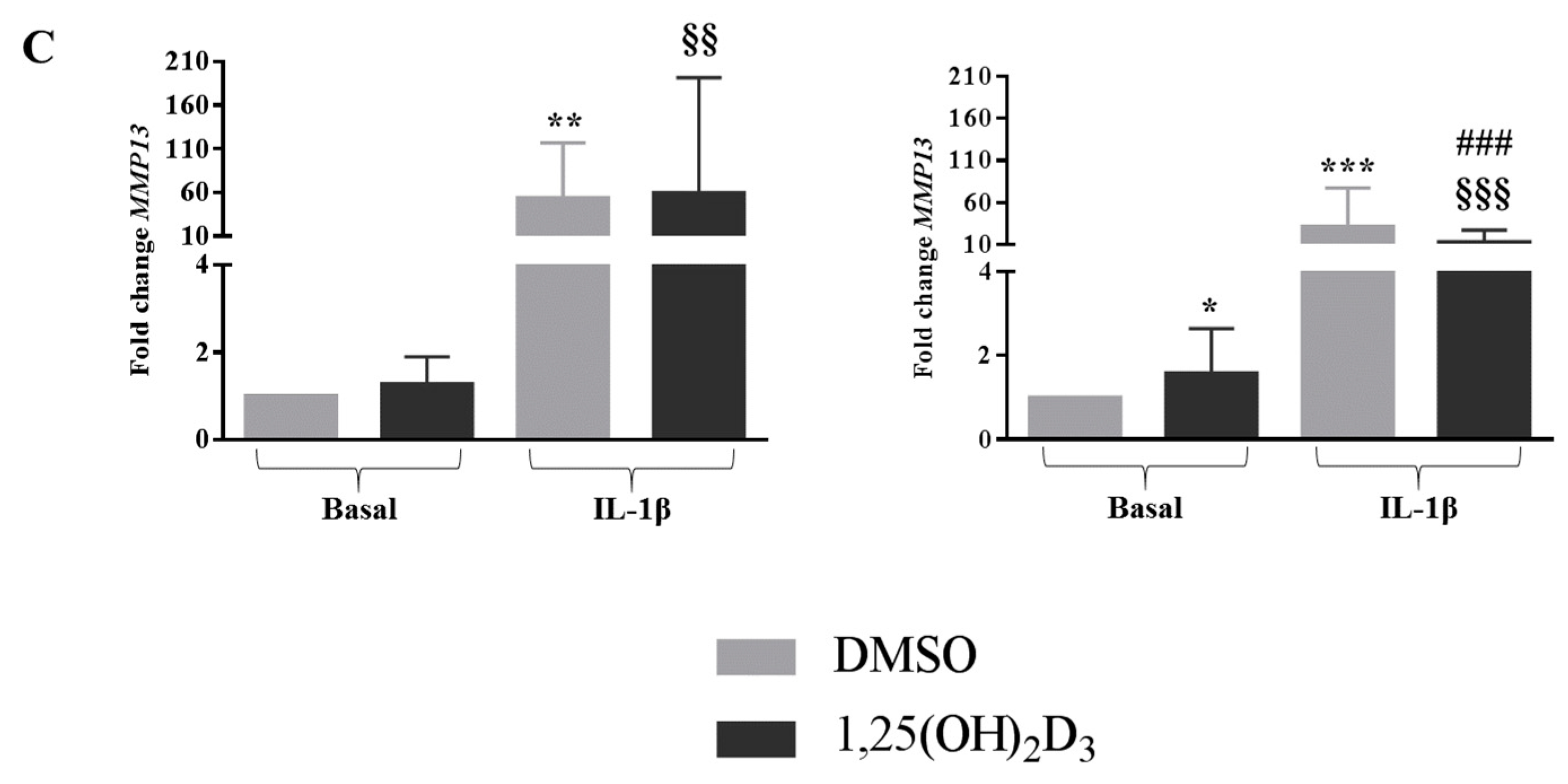
2.6. Vitamin D Was Able to Affect the Expression of MMPs in Cells Bearing the Ff Genotype in Both Basal and Inflamed Conditions
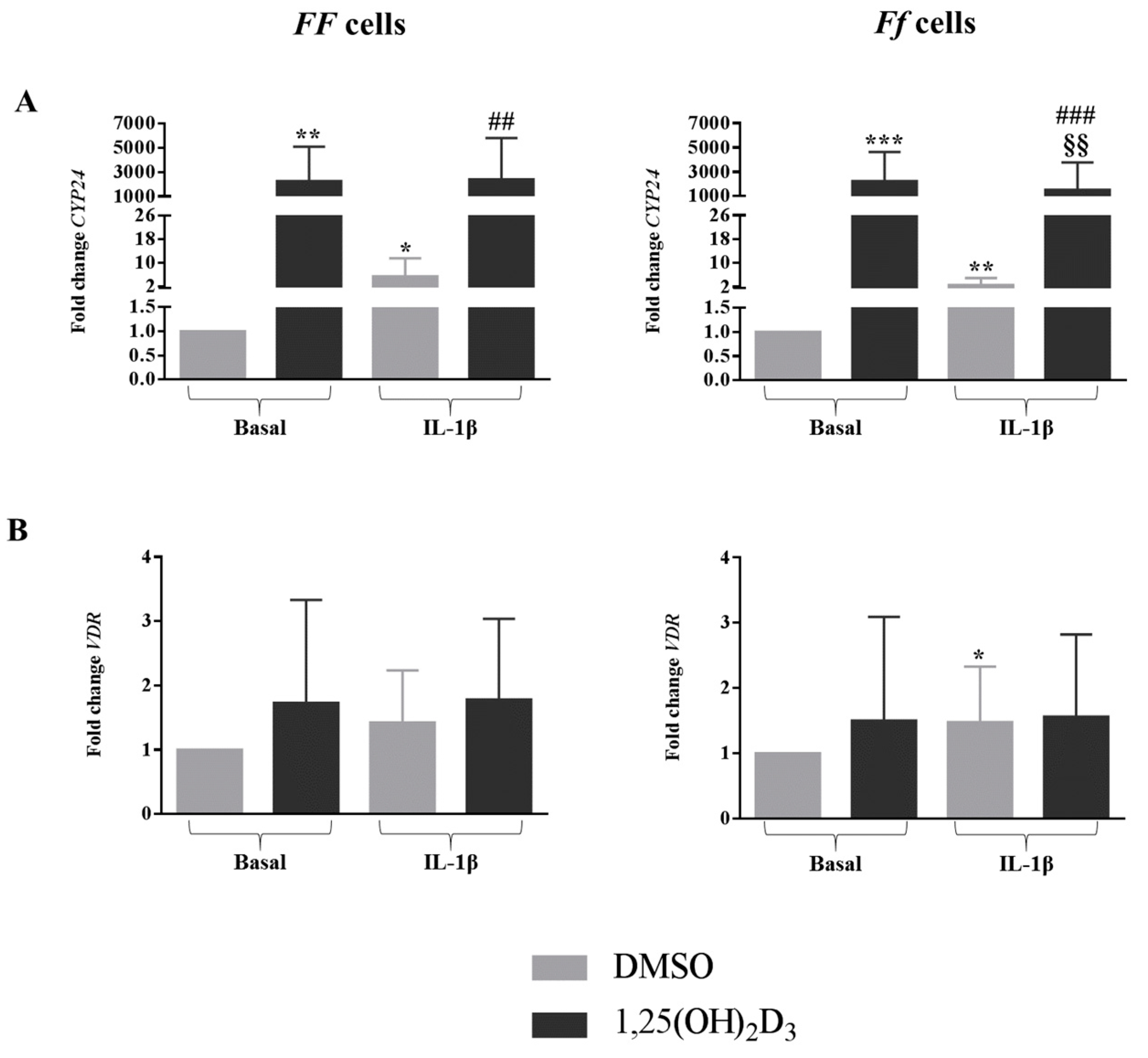
2.7. Vitamin D Upregulated the Vitamin D-Dependent Signaling Pathway, but This Was Downregulated in Inflamed Cells Bearing the Ff Genotype
3. Discussion
4. Materials and Methods
4.1. Study Population and Tissue Samples Collected
4.2. Isolation and Expansion of Disc Cells
4.3. Determination of FokI VDR Genotypes
4.4. Calcitriol (1,25(OH)2D3) and IL-1β Treatment Protocols
4.5. Cell Cycle Analysis
4.6. Evaluation of Apoptosis
4.7. Gene Expression Analysis
4.8. Determination of Cytokines
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| VDR | Vitamin D receptor |
| IL-1Ra | Interleukin-1 receptor antagonist |
| IL-6 | Interleukin 6 |
| NP | Nucleus pulposus |
| AF | Annulus fibrosus |
| CEP | Osteo-cartilaginous endplate |
| IL-1β | Interleukin 1β |
| ACAN | Aggrecan |
| COL1A1 | Collagen Type I Alpha 1 Chain |
| SOX9 | SRY-Box 9 |
| MMP | Matrix metalloprotease |
| CYP24 | Cytochrome P450 Family 24 Subfamily A Member 1 |
| DMSO | Dimethyl sulfoxide |
| TBP | TATA-binding protein |
References
- Gruber, H.E.; Hoelscher, G.; Ingram, J.A.; Chow, Y.; Loeffler, B.; Hanley, E.N., Jr. 1,25(OH)2-vitamin D3 inhibits proliferation and decreases production of monocyte chemoattractant protein-1, thrombopoietin, VEGF, and angiogenin by human annulus cells in vitro. Spine 2008, 33, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Colombini, A.; Lanteri, P.; Lombardi, G.; Grasso, D.; Recordati, C.; Lovi, A.; Banfi, G.; Bassani, R.; Brayda-Bruno, M. Metabolic effects of vitamin D active metabolites in monolayer and micromass cultures of nucleus pulposus and annulus fibrosus cells isolated from human intervertebral disc. Int. J. Biochem. Cell Boil. 2012, 44, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Colombini, A.; Cauci, S.; Lombardi, G.; Lanteri, P.; Croiset, S.; Brayda-Bruno, M.; Banfi, G. Relationship between vitamin D receptor gene (VDR) polymorphisms, vitamin D status, osteoarthritis and intervertebral disc degeneration. J. Steroid Biochem. Mol. Boil. 2013, 138, 24–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhao, S.; Niu, F.; Bi, G.B. Association between vitamin D receptor gene polymorphisms and intervertebral disc degeneration: A meta-analysis. J. Orthop. Sci. 2017, 22, 184–189. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Qin, Z.L.; Zong, S.H.; He, M.L.; Zhan, X.L.; Xiao, Z.M.; Wei, Q.J. Vitamin D receptor gene polymorphisms and lumbar disc degeneration: A systematic review and meta-analysis. Eur. Spine J. 2017, 26, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, M.; Shao, J.; Bai, Y.; Li, M. Association between VDR FokI Polymorphism and Intervertebral Disk Degeneration. Genom. Proteom. Bioinform. 2015, 13, 371–376. [Google Scholar] [CrossRef] [PubMed]
- Colombini, A.; Brayda-Bruno, M.; Lombardi, G.; Croiset, S.J.; Vrech, V.; Maione, V.; Banfi, G.; Cauci, S. FokI polymorphism in the vitamin D receptor gene (VDR) and its association with lumbar spine pathologies in the Italian population: A case-control study. PLoS ONE 2014, 9, e97027. [Google Scholar] [CrossRef] [PubMed]
- Colombini, A.; Brayda-Bruno, M.; Ferino, L.; Lombardi, G.; Maione, V.; Banfi, G.; Cauci, S. Gender differences in the VDR-FokI polymorphism and conventional non-genetic risk factors in association with lumbar spine pathologies in an Italian case-control study. Int. J. Mol. Sci. 2015, 16, 3722–3739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombini, A.; Brayda-Bruno, M.; Lombardi, G.; Croiset, S.J.; Ceriani, C.; Buligan, C.; Barbina, M.; Banfi, G.; Cauci, S. BsmI, ApaI and TaqI Polymorphisms in the Vitamin D Receptor Gene (VDR) and Association with Lumbar Spine Pathologies: An Italian Case-Control Study. PLoS ONE 2016, 11, e0155004. [Google Scholar] [CrossRef] [PubMed]
- Mashayekhi, S.; Saberi, A.; Salehi, Z.; Biazar, G.; Mehrdel, R. VDR and GC gene polymorphisms modulate the risk of lumbar disc degeneration in Iran. Clin. Neurol. Neurosurg. 2018, 165, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Brayda-Bruno, M.; Vigano, M.; Cauci, S.; Vitale, J.A.; de Girolamo, L.; De Luca, P.; Lombardi, G.; Banfi, G.; Colombini, A. Plasma vitamin D and osteo-cartilaginous markers in Italian males affected by intervertebral disc degeneration: Focus on seasonal and pathological trend of type II collagen degradation. Clin. Chim. Acta Int. J. Clin. Chem. 2017, 471, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Cauci, S.; Vigano, M.; de Girolamo, L.; De Luca, P.; Perucca Orfei, C.; Banfi, G.; Lombardi, G.; Brayda-Bruno, M.; Colombini, A. High Levels of Circulating Type II Collagen Degradation Marker (CTx-II) Are Associated with Specific VDR Polymorphisms in Patients with Adult Vertebral Osteochondrosis. Int. J. Mol. Sci. 2017, 18, 2073. [Google Scholar] [CrossRef] [PubMed]
- Uitterlinden, A.G.; Fang, Y.; Van Meurs, J.B.; Pols, H.A.; Van Leeuwen, J.P. Genetics and biology of vitamin D receptor polymorphisms. Gene 2004, 338, 143–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costanzo, P.; Santini, A.; Fattore, L.; Novellino, E.; Ritieni, A. Toxicity of aflatoxin B1 towards the vitamin D receptor (VDR). Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2015, 76, 77–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurutka, P.W.; Whitfield, G.K.; Hsieh, J.C.; Thompson, P.D.; Haussler, C.A.; Haussler, M.R. Molecular nature of the vitamin D receptor and its role in regulation of gene expression. Rev. Endocr. Metab. Disord. 2001, 2, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Guillot, X.; Semerano, L.; Saidenberg-Kermanac’h, N.; Falgarone, G.; Boissier, M.C. Vitamin D and inflammation. Joint Bone Spine 2010, 77, 552–557. [Google Scholar] [CrossRef] [PubMed]
- Molinos, M.; Almeida, C.R.; Caldeira, J.; Cunha, C.; Goncalves, R.M.; Barbosa, M.A. Inflammation in intervertebral disc degeneration and regeneration. J. R. Soc. Interface 2015, 12, 20150429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombini, A.; Lombardi, G.; Corsi, M.M.; Banfi, G. Pathophysiology of the human intervertebral disc. Int. J. Biochem. Cell Boil. 2008, 40, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Le Maitre, C.L.; Freemont, A.J.; Hoyland, J.A. The role of interleukin-1 in the pathogenesis of human intervertebral disc degeneration. Arth. Res. Ther. 2005, 7, R732–R745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Maitre, C.L.; Pockert, A.; Buttle, D.J.; Freemont, A.J.; Hoyland, J.A. Matrix synthesis and degradation in human intervertebral disc degeneration. Biochem. Soc. Trans. 2007, 35 Pt 4, 652–655. [Google Scholar] [CrossRef] [PubMed]
- Johnson, Z.I.; Schoepflin, Z.R.; Choi, H.; Shapiro, I.M.; Risbud, M.V. Disc in flames: Roles of TNF-alpha and IL-1beta in intervertebral disc degeneration. Eur. Cells Mater. 2015, 30, 104–116, discussion 116–117. [Google Scholar] [CrossRef]
- McFadden, K.D.; Taylor, J.R. End-plate lesions of the lumbar spine. Spine 1989, 14, 867–869. [Google Scholar] [CrossRef] [PubMed]
- Aufdermaur, M. Juvenile kyphosis (Scheuermann’s disease): Radiography, histology, and pathogenesis. Clin. Orthop. Relat. Res. 1981, 166–174. [Google Scholar]
- Kyere, K.A.; Than, K.D.; Wang, A.C.; Rahman, S.U.; Valdivia-Valdivia, J.M.; La Marca, F.; Park, P. Schmorl’s nodes. Eur. Spine J. 2012, 21, 2115–2121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, J.P.; Smith, S.; Fairbank, J.C. Nutrition of the intervertebral disc. Spine 2004, 29, 2700–2709. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Li, F.C.; Huang, Y.J.; Teng, C.; Chen, W.S. Possible key role of immune system in Schmorl’s nodes. Med. Hypotheses 2010, 74, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Goltzman, D. Functions of vitamin D in bone. Histochem. Cell Biol. 2018, 149, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Lopa, S.; Ceriani, C.; Cecchinato, R.; Zagra, L.; Moretti, M.; Colombini, A. Stability of housekeeping genes in human intervertebral disc, endplate and articular cartilage cells in multiple conditions for reliable transcriptional analysis. Eur. Cells Mater. 2016, 31, 395–406. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, H.; Suh, M.J.; Shin, J.H.; Hwang, M.H.; Lee, H.M. Effect of biphasic electrical current stimulation on IL-1beta-stimulated annulus fibrosus cells using in vitro microcurrent generating chamber system. Spine 2013, 38, E1368–E1376. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Age | Spine Disorder | Disc Level | FokI VDR Genotype |
|---|---|---|---|---|
| F | 27 | Spondylolisthesis | L5-S1 | FF |
| F | 39 | Discopathy | L4-L5 | FF |
| F | 46 | Degenerative discopathy | L5-S1 | Ff |
| F | 48 | Spondylolisthesis | L5-S1 | FF |
| F | 48 | Degenerative discopathy | L5-S1 | Ff |
| F | 50 | Discopathy | L5-S1 | Ff |
| F | 50 | Discopathy | L5-S1 | Ff |
| M | 52 | Discopathy | L5-S1 | Ff |
| F | 55 | Discopathy | L5-S1 | Ff |
| M | 57 | Discopathy | L5-S1 | Ff |
| F | 65 | Discopathy | L5-S1 | Ff |
| F | 66 | Degenerative discopathy | L5-S1 | FF |
| F | 66 | Discopathy | L5-S1 | Ff |
| F | 74 | Discopathy | L5-S1 | Ff |
| F | 76 | Discopathy | L5-S1 | Ff |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Luca, P.; De Girolamo, L.; Perucca Orfei, C.; Viganò, M.; Cecchinato, R.; Brayda-Bruno, M.; Colombini, A. Vitamin D’s Effect on the Proliferation and Inflammation of Human Intervertebral Disc Cells in Relation to the Functional Vitamin D Receptor Gene FokI Polymorphism. Int. J. Mol. Sci. 2018, 19, 2002. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19072002
De Luca P, De Girolamo L, Perucca Orfei C, Viganò M, Cecchinato R, Brayda-Bruno M, Colombini A. Vitamin D’s Effect on the Proliferation and Inflammation of Human Intervertebral Disc Cells in Relation to the Functional Vitamin D Receptor Gene FokI Polymorphism. International Journal of Molecular Sciences. 2018; 19(7):2002. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19072002
Chicago/Turabian StyleDe Luca, Paola, Laura De Girolamo, Carlotta Perucca Orfei, Marco Viganò, Riccardo Cecchinato, Marco Brayda-Bruno, and Alessandra Colombini. 2018. "Vitamin D’s Effect on the Proliferation and Inflammation of Human Intervertebral Disc Cells in Relation to the Functional Vitamin D Receptor Gene FokI Polymorphism" International Journal of Molecular Sciences 19, no. 7: 2002. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19072002