Silibinin Ameliorates O-GlcNAcylation and Inflammation in a Mouse Model of Nonalcoholic Steatohepatitis

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Two-Dimensional Gel Electrophoresis
2.3. In-Gel Digestion and MALDI-TOF Analysis
2.4. Immunoblotting
2.5. Immunohistochemistry
2.6. Immunoprecipitation
2.7. Quantitative Reverse-Transcription Polymerase Chain Reaction
2.8. Statistical Analysis
3. Results
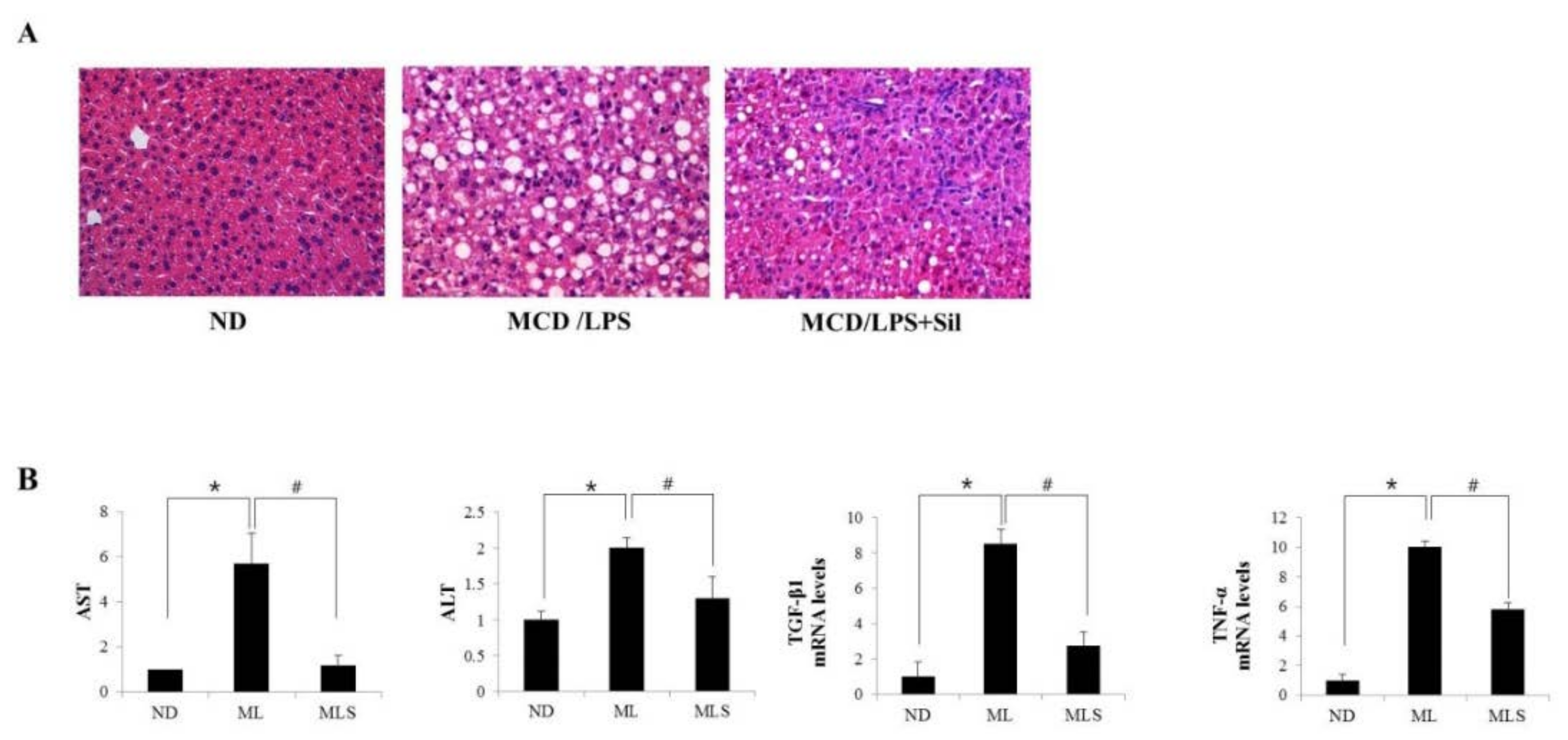
3.1. Effect of Silibinin on MCD Diet-Induced Steatohepatitis
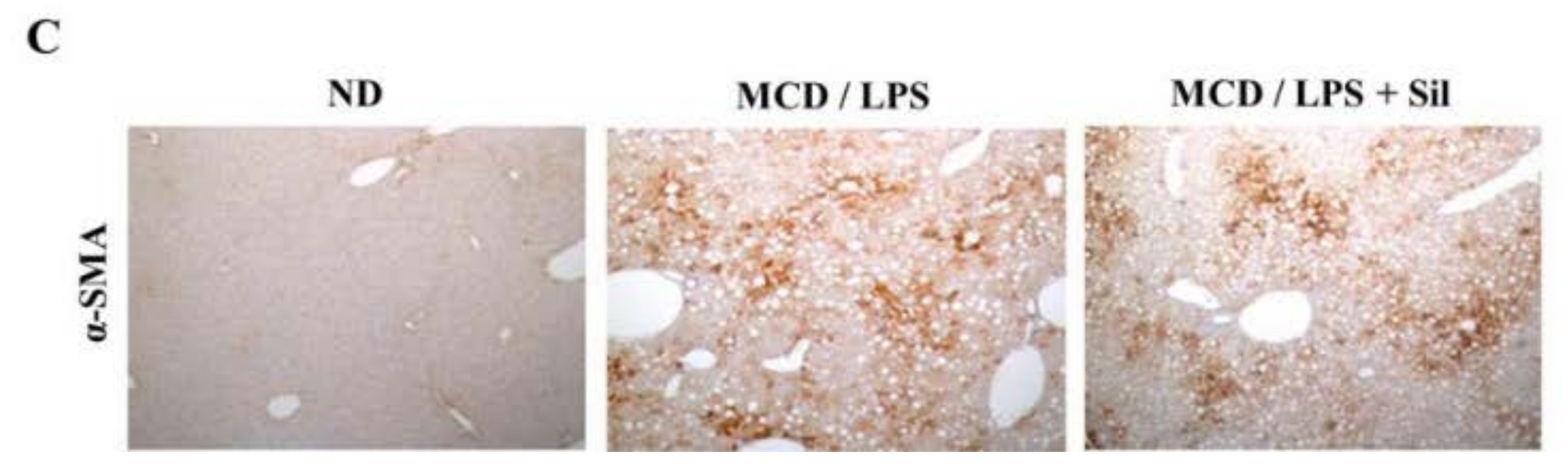
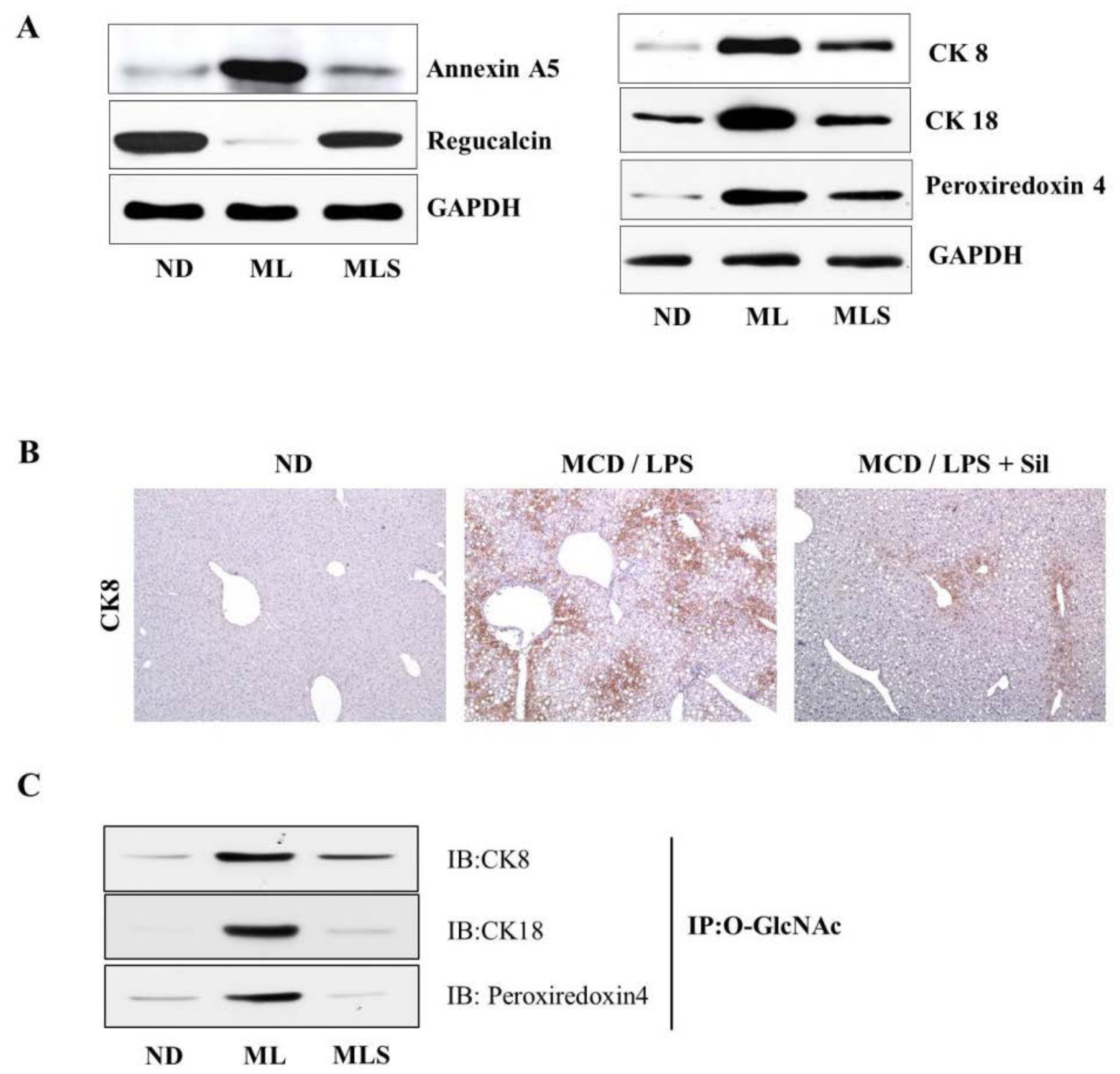
3.2. DE Analysis for Differentially Expressed Proteins
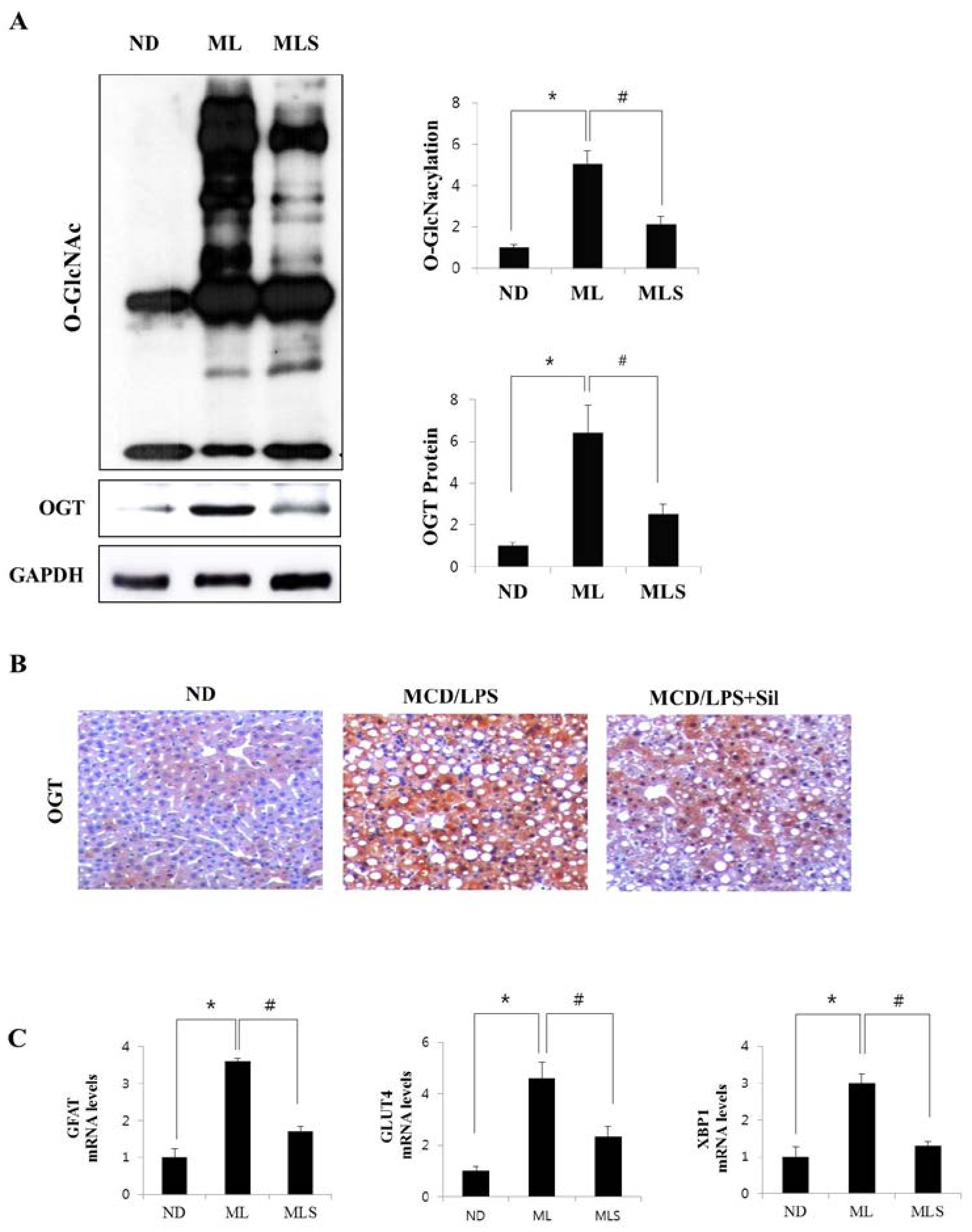
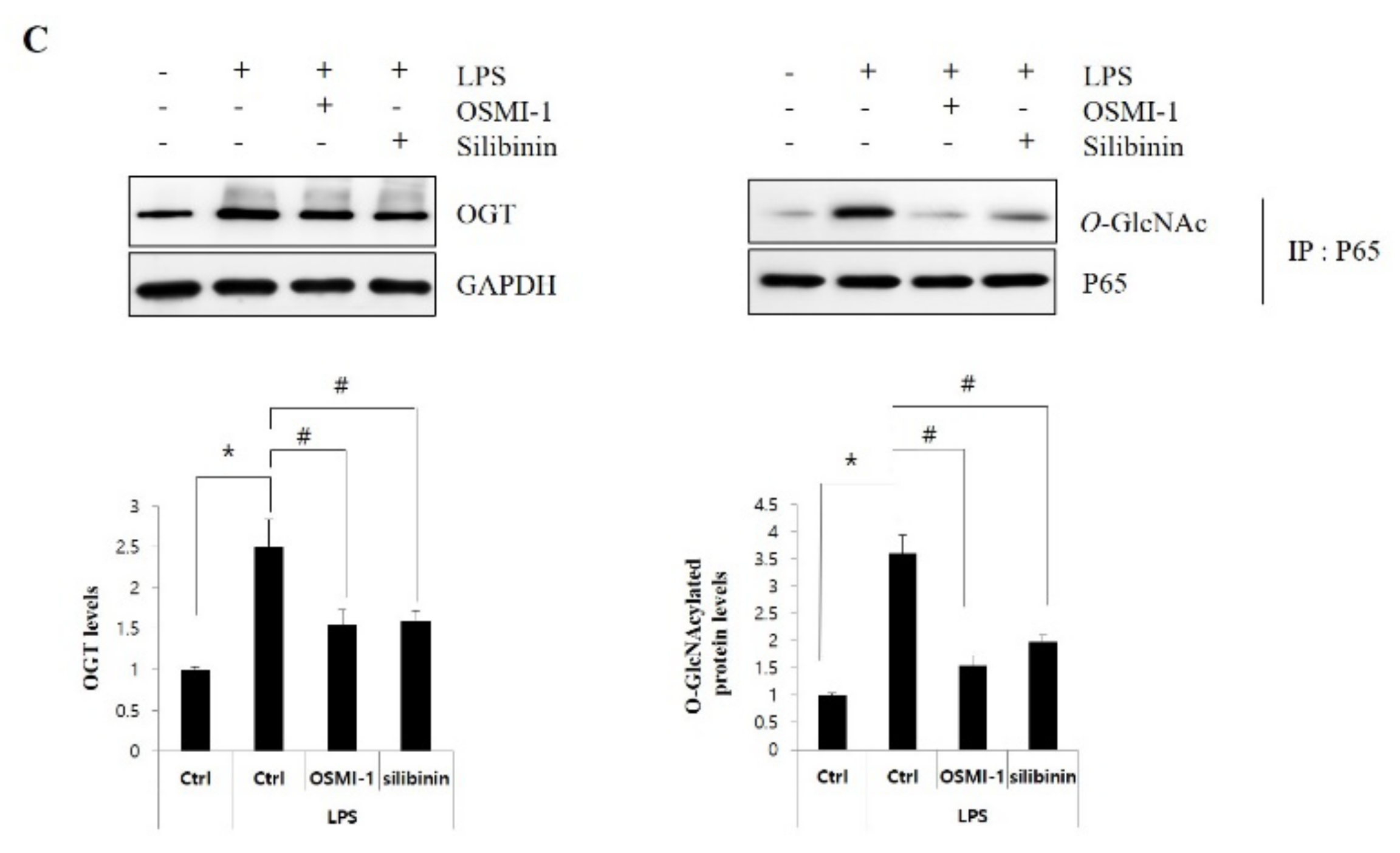
3.3. Suppression of O-GlcNAcylation by Silibinin in Steatohepatitis
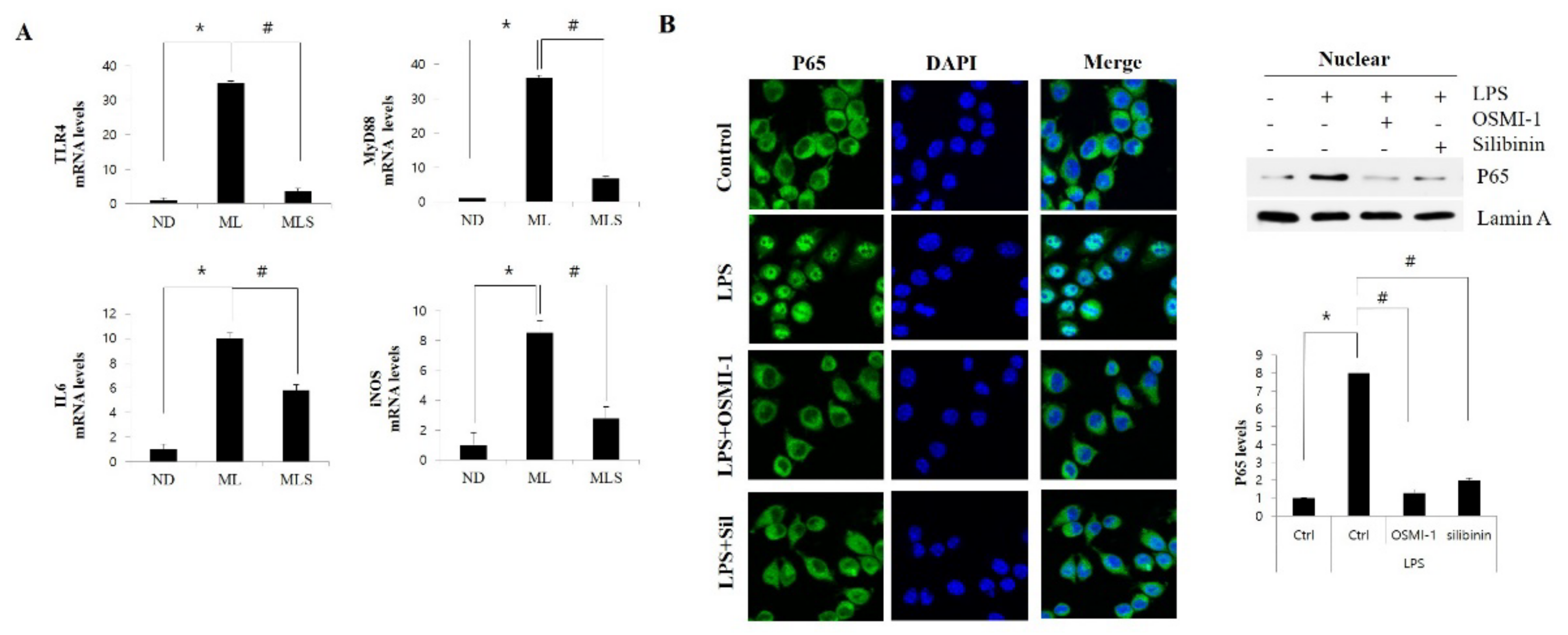
3.4. Silibinin Ameliorates Inflammation by Inhibiting NF-κB Signaling and Glcnacylation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Wong, V.W.; Chan, W.K.; Chitturi, S.; Chawla, Y.; Dan, Y.Y.; Duseja, A.; Fan, J.; Goh, K.L.; Hamaguchi, M.; Hashimoto, E.; et al. Asia-Pacific Working Party on Non-alcoholic Fatty Liver Disease guidelines 2017—Part 1: Definition, risk factors and assessment. J. Gastroenterol. Hepatol. 2018, 33, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Chitturi, S.; Wong, V.W.; Chan, W.K.; Wong, G.L.; Wong, S.K.; Sollano, J.; Ni, Y.H.; Liu, C.J.; Lin, Y.C.; Lesmana, L.A.; et al. The Asia-Pacific Working Party on Non-alcoholic Fatty Liver Disease guidelines 2017—Part 2: Management and special groups. J. Gastroenterol. Hepatol. 2018, 33, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Miura, R. Acyl-CoA dehydrogenases and acyl-CoA oxidases. Structural basis for mechanistic similarities and differences. Eur. J. Biochem. 2004, 271, 483–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pessayre, D.; Fromenty, B. NASH: A mitochondrial disease. J. Hepatol. 2005, 42, 928–940. [Google Scholar] [CrossRef] [PubMed]
- Szabo, G.; Dolganiuc, A.; Mandrekar, P. Pattern recognition receptors: A contemporary view on liver diseases. Hepatology 2006, 44, 287–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladas, E.J.; Cheng, B.; Hughes, D.; Kroll, D.J.; Graf, T.N.; Oberlies, N.H.; Rheingold, S.; Sacks, N.; Kelly, K.M. Milk Thistle is associated with reductions in liver function tests (LFTs) in children undergoing therapy for acute lymphoblastic leukemia (ALL). Blood 2006, 108, 533a–534a. [Google Scholar]
- Polyak, S.J.; Morishima, C.; Lohmann, V.; Pal, S.; Lee, D.Y.; Liu, Y.; Graf, T.N.; Oberlies, N.H. Identification of hepatoprotective flavonolignans from silymarin. Proc. Natl. Acad. Sci. USA 2010, 107, 5995–5999. [Google Scholar] [CrossRef] [PubMed]
- Saliou, C.; Valacchi, G.; Rimbach, G. Assessing bioflavonoids as regulators of NF-kappa B activity and inflammatory gene expression in mammalian cells. Methods Enzymol. 2001, 335, 380–387. [Google Scholar] [PubMed]
- Abdal Dayem, A.; Choi, H.Y.; Yang, G.M.; Kim, K.; Saha, S.K.; Cho, S.G. The Anti-Cancer Effect of Polyphenols against Breast Cancer and Cancer Stem Cells: Molecular Mechanisms. Nutrients 2016, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Buse, M.G.; Robinson, K.A.; Marshall, B.A.; Hresko, R.C.; Mueckler, M.M. Enhanced O-GlcNAc protein modification is associated with insulin resistance in GLUT1-overexpressing muscles. Am. J. Physiol.-Endocrinol. Metab. 2002, 283, E241–E250. [Google Scholar] [CrossRef] [PubMed]
- Peterson, S.B.; Hart, G.W. New insights: A role for O-GlcNAcylation in diabetic complications. Crit. Rev. Biochem. Mol. Biol. 2016, 51, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Slawson, C.; Copeland, R.J.; Hart, G.W. O-GIcNAc signaling: A metabolic link between diabetes and cancer? Trends Biochem. Sci. 2010, 35, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Zeidan, Q.; Hart, G.W. The intersections between O-GlcNAcylation and phosphorylation: Implications for multiple signaling pathways. J. Cell Sci. 2010, 123, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hart, G.W. Dynamic O-linked glycosylation of nuclear and cytoskeletal proteins. Annu. Rev. Biochem. 1997, 66, 315–335. [Google Scholar] [CrossRef] [PubMed]
- Van Herck, M.A.; Vonghia, L.; Francque, S.M. Animal Models of Nonalcoholic Fatty Liver Disease—A Starter’s Guide. Nutrients 2017, 9, 1072. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radical Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Ryu, H.S.; Kim, H.T.; Lee, S.J.; Choi, U.K.; Park, Y.B.; Huh, T.L.; Choi, M.S.; Kang, T.C.; Choi, S.Y.; et al. Proteomic analysis of human macrophages exposed to hypochlorite-oxidized low-density lipoprotein. Biochim. Biophys. Acta 2009, 1794, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.H.; Cho, H.J.; Lee, I.S.; Kim, M.; Lee, I.K.; Chang, Y.C. Comparative proteome analysis of TGF-beta1-induced fibrosis processes in normal rat kidney interstitial fibroblast cells in response to ascofuranone. Proteomics 2009, 9, 4445–4456. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Kang, J.H.; Choi, S.Y.; Suk, K.T.; Kim, D.J.; Kwon, O.S. PKC delta as a Regulator for TGF beta 1-Induced alpha-SMA Production in a Murine Nonalcoholic Steatohepatitis Model. PLoS ONE 2013, 8, e55979. [Google Scholar]
- Rinella, M.E.; Green, R.M. The methionine-choline deficient dietary model of steatohepatitis does not exhibit insulin resistance. J. Hepatol. 2004, 40, 47–51. [Google Scholar] [CrossRef] [PubMed]
- De Souza, C.O.; Peracoli, M.T.S.; Weel, I.C.; Bannwart, C.F.; Romao, M.; Nakaira-Takahagi, E.; de Medeiros, L.T.L.; da Silva, M.G.; Peracoli, J.C. Hepatoprotective and anti-inflammatory effects of silibinin on experimental preeclampsia induced by 1-NAME in rats. Life Sci. 2012, 91, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, C.; Wadhwa, R.; Deep, G.; Biedermann, D.; Gazak, R.; Kren, V.; Agarwal, R. Anti-Cancer Efficacy of Silybin Derivatives—A Structure-Activity Relationship. PLoS ONE 2013, 8, e60074. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA is induced by ATF6 and spliced by IRE1 in response to ER stress to produce a highly active transcription factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef]
- Chaiyawat, P.; Netsirisawan, P.; Svasti, J.; Champattanachai, V. Aberrant O-GlcNAcylated Proteins: New Perspectives in Breast and Colorectal Cancer. Front. Endocrinol. (Lausanne) 2014, 5, 193. [Google Scholar] [CrossRef] [PubMed]
- Baldini, S.F.; Lefebvre, T. O-GlcNAcylation and the Metabolic Shift in High-Proliferating Cells: All the Evidence Suggests that Sugars Dictate the Flux of Lipid Biogenesis in Tumor Processes. Front. Oncol. 2016, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Baudoin, L.; Issad, T. O-GlcNAcylation and inflammation: A vast territory to explore. Front. Endocrinol. 2015, 5, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Hart, G.W. Protein O-GlcNAcylation in diabetes and diabetic complications. Expert Rev. Proteom. 2013, 10, 365–380. [Google Scholar] [CrossRef] [PubMed]
- Visinoni, S.; Khalid, N.F.I.; Joannides, C.N.; Shulkes, A.; Yim, M.; Whitehead, J.; Tiganis, T.; Lamont, B.J.; Favaloro, J.M.; Proietto, J.; et al. The Role of Liver Fructose-1,6-Bisphosphatase in Regulating Appetite and Adiposity. Diabetes 2012, 61, 1122–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Makino, T.; Yamasaki, M.; Takeno, A.; Shirakawa, M.; Miyata, H.; Takiguchi, S.; Nakajima, K.; Fujiwara, Y.; Nishida, T.; Matsuura, N.; et al. Cytokeratins 18 and 8 are poor prognostic markers in patients with squamous cell carcinoma of the oesophagus. Br. J. Cancer 2009, 101, 1298–306. [Google Scholar] [CrossRef] [PubMed]
- Schussler, M.H.; Skoudy, A.; Ramaekers, F.; Real, F.X. Intermediate Filaments as Differentiation Markers of Normal Pancreas and Pancreas Cancer. Am. J. Pathol. 1992, 140, 559–568. [Google Scholar] [PubMed]
- Chou, C.F.; Smith, A.J.; Omary, M.B. Characterization and Dynamics of O-Linked Glycosylation of Human Cytokeratin-8 and Cytokeratin-18. J. Biol. Chem. 1992, 267, 3901–3906. [Google Scholar] [PubMed]
- Ku, N.O.; Omary, M.B. Identification and Mutational Analysis of the Glycosylation Sites of Human Keratin-18. J. Biol. Chem. 1995, 270, 11820–11827. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, B.; Vaidya, M.M.; Kalraiya, R.D. O-GlcNAcylation Determines the Solubility, Filament Organization, and Stability of Keratins 8 and 18. J. Biol. Chem. 2010, 285, 34062–34071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okado-Matsumoto, A.; Matsumoto, A.; Fujii, J.; Taniguchi, N. Peroxiredoxin IV is a secretable protein with heparin-binding properties under reduced conditions. FASEB J. 2000, 14, 493–501. [Google Scholar] [CrossRef]
- Fujii, J.; Tsunoda, S. Redox regulation of fertilisation and the spermatogenic process. Asian J. Androl. 2011, 13, 420–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuchi, Y.; Okada, F.; Tsunoda, S.; Kibe, N.; Shirasawa, N.; Ikawa, M.; Okabe, M.; Ikeda, Y.; Fujii, J. Peroxiredoxin 4 knockout results in elevated spermatogenic cell death via oxidative stress. Biochem. J. 2009, 419, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Galligan, J.J.; Petersen, D.R. The human protein disulfide isomerase gene family. Hum. Genom. 2012, 6, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.Q.; Yang, J.T.; Guo, Y.B.; Ye, H.; Yu, C.H.; Xu, C.F.; Xu, L.; Wu, S.F.; Sun, W.; Wei, H.D.; et al. Functional Proteomic Analysis of Nonalcoholic Fatty Liver Disease in Rat Models: Enoyl-Coenzyme A Hydratase Down-Regulation Exacerbates Hepatic Steatosis. Hepatology 2010, 51, 1190–1199. [Google Scholar] [CrossRef] [PubMed]
- Greten, F.R.; Arkan, M.C.; Bollrath, J.; Hsu, L.C.; Goode, J.; Miething, C.; Goktuna, S.I.; Neuenhahn, M.; Fierer, J.; Paxian, S.; et al. NF-kappaB is a negative regulator of IL-1beta secretion as revealed by genetic and pharmacological inhibition of IKKbeta. Cell 2007, 130, 918–931. [Google Scholar] [CrossRef] [PubMed]
- Karin, M. NF-kappa B as a Critical Link between Inflammation and Cancer. Cold Spring Harb Perspect. Biol. 2009, 1, a000141. [Google Scholar] [CrossRef] [PubMed]
- Karin, M.; Lawrence, T.; Nizet, V. Innate immunity gone awry: Linking microbial infections to chronic inflammation and cancer. Cell 2006, 124, 823–835. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.D.; Nathan, C.; Hom, G.; Chartrain, N.; Fletcher, D.S.; Trumbauer, M.; Stevens, K.; Xie, Q.W.; Sokol, K.; Hutchinson, N.; et al. Altered responses to bacterial infection and endotoxic shock in mice lacking inducible nitric oxide synthase. Cell 1995, 81, 641–650. [Google Scholar] [CrossRef]
- James, L.R.; Tang, D.; Ingram, A.; Ly, H.; Thai, K.; Cai, L.; Scholey, J.W. Flux through the hexosamine pathway is a determinant of nuclear factor kappa B-dependent promoter activation. Diabetes 2002, 51, 1146–1156. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.H.; Park, S.Y.; Nam, H.W.; Kim, D.H.; Kang, J.G.; Kang, E.S.; Kim, Y.S.; Lee, H.C.; Kim, K.S.; Cho, J.W. NF kappa B activation is associated with its O-GlcNAcylation state under hyperglycemic conditions. Proc. Natl. Acad. Sci. USA 2008, 105, 17345–17350. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, P.; Clark, P.M.; Mason, D.E.; Peters, E.C.; Hsieh-Wilson, L.C.; Baltimore, D. Activation of the transcriptional function of the NF-kappaB protein c-Rel by O-GlcNAc glycosylation. Sci. Signal. 2013, 6, ra75. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No a | Identified Protein | Accession No. | MW (KDA) | PI | MOWSE SCORE | Coverage (%) | Fold Change (ND = 1) | |
|---|---|---|---|---|---|---|---|---|
| ML | MLS | |||||||
| Metabolism | ||||||||
| 1 | Ornithine aminotransferase, mitochondrial | P29578 | 48.4 | 6.2 | 1.43 × 106 | 33.7 | 0.5 | 0.8 |
| 2 | Indolethylamine N-methyltransferase | P40936 | 29.5 | 6.0 | 282 | 28.8 | 0.2 | 0.8 |
| 3 | Glutathione synthase | P51855 | 52.2 | 5.6 | 73,657 | 24.1 | 1.6 | 0.6 |
| 4 | Fructose-1,6-bisphosphatase | Q9QXD6 | 36.9 | 6.1 | 1.23 × 106 | 44.7 | 2 | 1.1 |
| 5 | Annexin A5 | P48036 | 35.8 | 4.8 | 1531 | 15.0 | 3 | 1.2 |
| 6 | Zinc finger protein 330 | Q922H9 | 35.6 | 5.8 | 50.9 | 17.1 | 0.3 | 1 |
| Structural | ||||||||
| 7 | Keratin, type II cytoskeletal 8 * | P11679 | 54.6 | 5.7 | 20,152 | 21.0 | 1.4 | 0.8 |
| 8 | Keratin, type II cytoskeletal 18 * | P05784 | 47.5 | 5.2 | 3.72 × 107 | 36.6 | 4 | 1.2 |
| 9 | Protein disulfide-isomerase A3 * | P22273 | 56.6 | 5.9 | 66,094 | 34.3 | 1.5 | 1 |
| 10 | Actin, cytoplasmic 1 * | P60710 | 41.7 | 5.3 | 5421 | 21.1 | 5 | 2.5 |
| 11 | Rho GDP-dissociation inhibitor 1 * | Q99PT1 | 23.4 | 5.1 | 1960 | 59.3 | 2.5 | 1.4 |
| 12 | Tropomyosin alpha-1 chain | P58771 | 32.7 | 4.7 | 32.5 | 9.9 | 3 | 1 |
| Genomic | ||||||||
| 13 | Transcription factor E2F6 | Q61502 | 36.6 | 4.9 | 110 | 11.0 | 0.4 | 0.7 |
| 14 | 40s ribosomal protein SA * | P14206 | 32.8 | 4.8 | 21,692 | 40.3 | 1.7 | 1 |
| 15 | Protein NDRG2 | Q9QYG0 | 40.8 | 5.2 | 31,743 | 29.4 | 0.2 | 0.9 |
| 16 | Proliferating cell nuclear antigen | P17918 | 28.8 | 4.7 | 100 | 11.5 | 1.5 | 0.9 |
| Oxidative stress | ||||||||
| 17 | Peroxiredoxin-4 * | Q08807 | 31.0 | 6.7 | 4.96 × 106 | 48.5 | 5 | 2 |
| 18 | Regucalcin | Q64374 | 33.4 | 5.2 | 480,847 | 42.5 | 0.3 | 1.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.J.; Nam, M.J.; Lee, D.E.; Park, J.-W.; Kang, B.S.; Lee, D.-S.; Lee, H.-S.; Kwon, O.-S. Silibinin Ameliorates O-GlcNAcylation and Inflammation in a Mouse Model of Nonalcoholic Steatohepatitis. Int. J. Mol. Sci. 2018, 19, 2165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082165
Lee SJ, Nam MJ, Lee DE, Park J-W, Kang BS, Lee D-S, Lee H-S, Kwon O-S. Silibinin Ameliorates O-GlcNAcylation and Inflammation in a Mouse Model of Nonalcoholic Steatohepatitis. International Journal of Molecular Sciences. 2018; 19(8):2165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082165
Chicago/Turabian StyleLee, Su Jin, Min Jung Nam, Da Eun Lee, Jeen-Woo Park, Beom Sik Kang, Dong-Seok Lee, Hyun-Shik Lee, and Oh-Shin Kwon. 2018. "Silibinin Ameliorates O-GlcNAcylation and Inflammation in a Mouse Model of Nonalcoholic Steatohepatitis" International Journal of Molecular Sciences 19, no. 8: 2165. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082165