From the Clinical Problem to the Basic Research—Co-Culture Models of Osteoblasts and Osteoclasts

 , ,
, ,
Abstract
:1. Introduction
- Which cell type(s) should be used?
- Single mono-culture model or a co-culture model?
- Which cell culture type should be used?
- o
- two dimensional (2D) or three dimensional (3D)?
- o
- static or dynamic?
- In case of 3D cultures, which matrix should be used?
- What are the molecular and functional markers to validate possible changes in the applied co-culture models?
- How can co-culture models be normalized to study cell specific changes within the co-culture models?
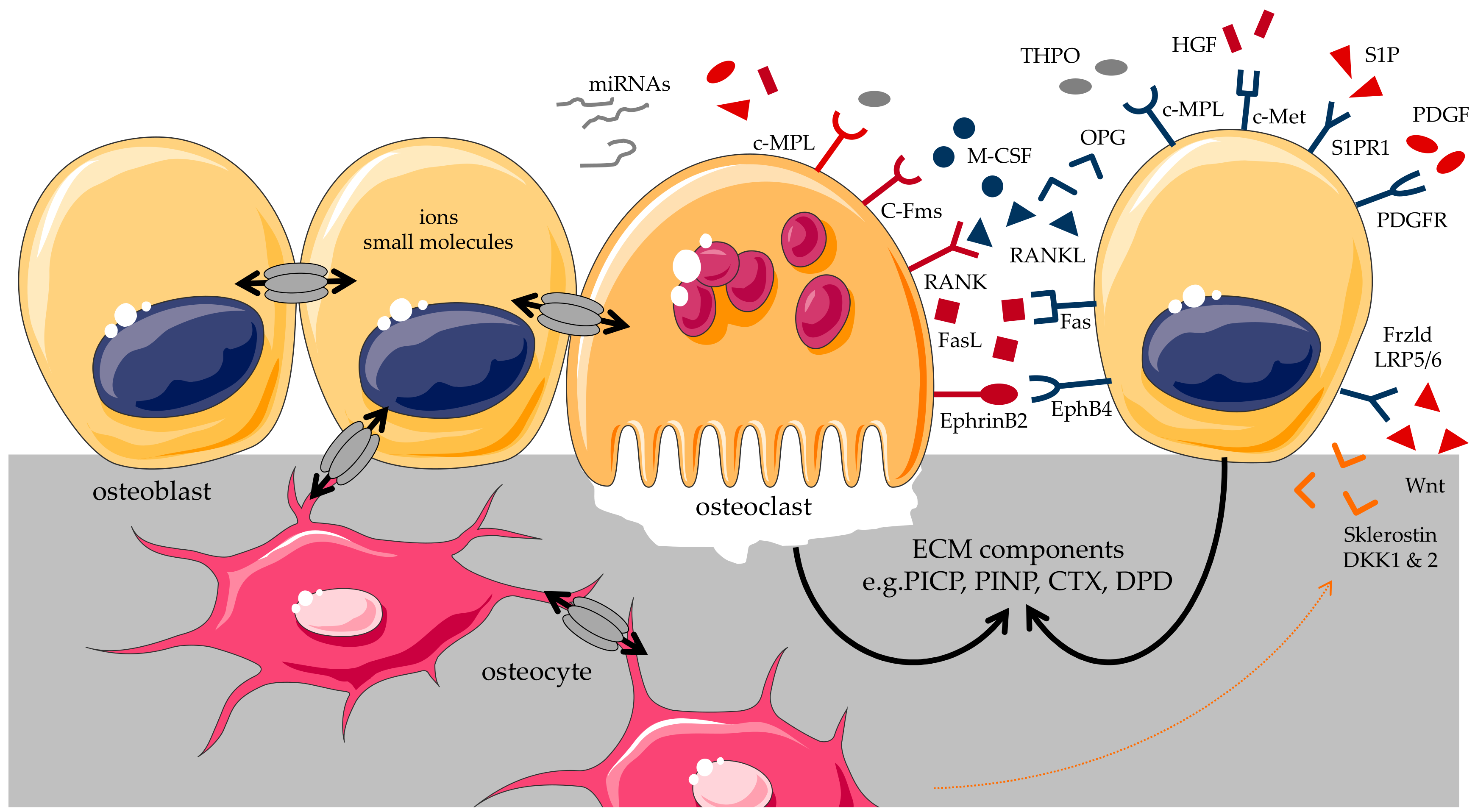
2. Osteoblast and Osteoclast Interaction
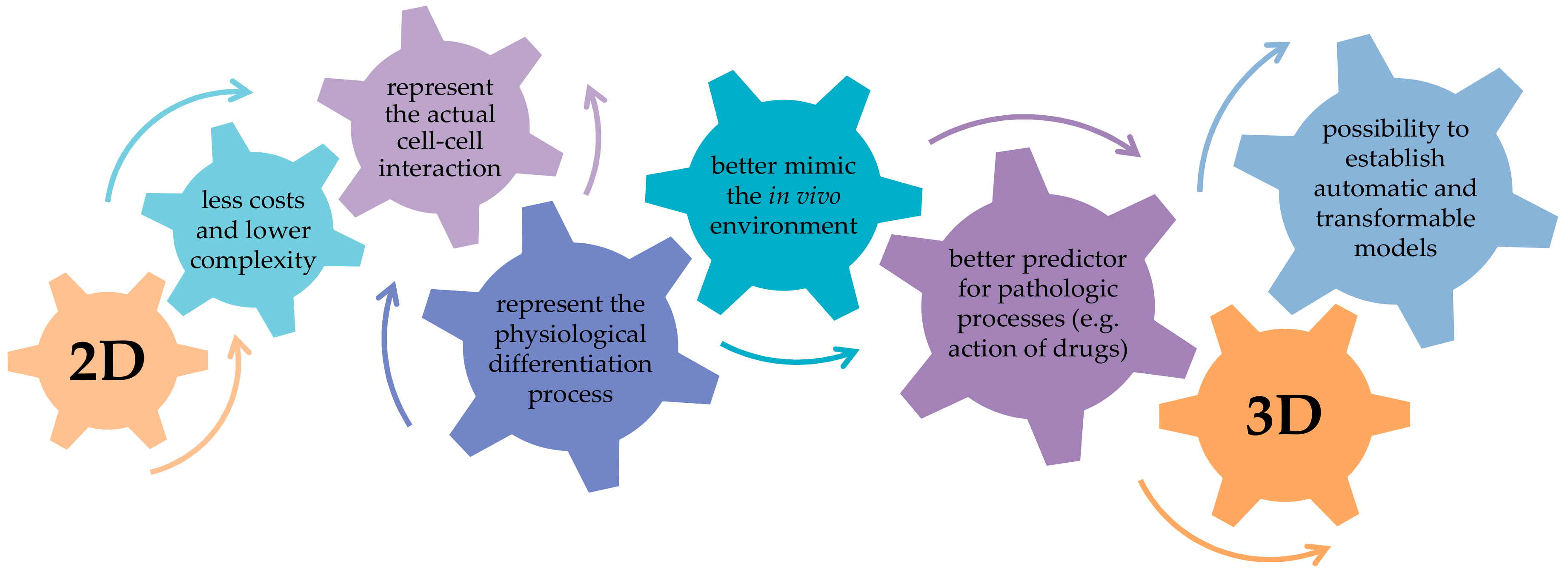
3. Co-Culture Models
4. Co-Cultures of Osteoblasts and Osteoclasts
4.1. Selection of Culture Models
4.2. Requirements for the 3D Matrix Used for Culturing Bone Cells
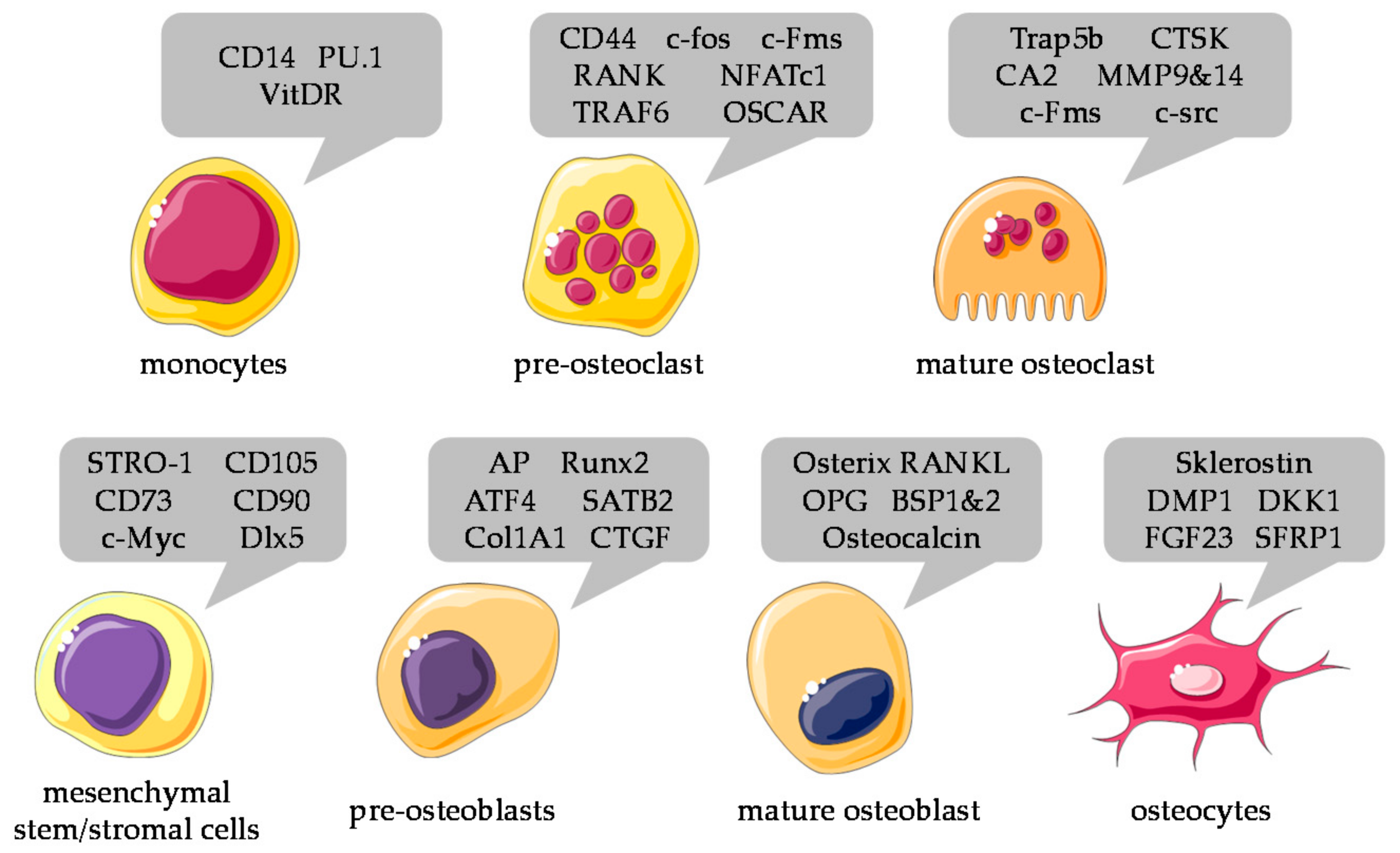
4.3. Selection of Cells
4.4. Selection of Medium
5. Characterization and Normalization of Different Cell Types in Co-Culture Systems
6. Recent Developments and Future Challenges
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| Ad-MSCs | MSCs derived from adipose tissue |
| AP | alkaline phosphatase |
| ATF4 | activating transcription factor 4 |
| BCL-2 | B-cell lymphoma 2 |
| BMP | bone morphogenetic proteins |
| B-MSCs | MSCs derived from bone marrow |
| BSP | bone sialoprotein |
| CAII | carbonic anhydrase II |
| CALCR | calcitonin receptor |
| cAMP | cyclic adenosine monophosphate |
| CD | cluster of differentiation |
| COL1A1 | collagen type I α 1 |
| CT | computed tomography |
| CTGF | connective tissue growth factor |
| CTSK | cathepsin K |
| CTX | collagen type 1 C-telopeptide |
| DKK1&2 | dickkopf 1 & 2 |
| DMP1 | dentin matrix acidic phosphoprotein 1 |
| DOK3 | downstream of kinase 3 |
| DPD | deoxypyridinoline |
| ECM | extracellular matrix |
| FGF-23 | fibroblast growth factor 23 |
| HGF | hepatic growth factor |
| hiPSCs | human induced pluripotent stem cells |
| IGF | insulin-like growth factor |
| IL | interleukin |
| IP3 | inositol trisphosphate |
| JAK | Janus kinase |
| LDH | lactate dehydrogenase |
| MATF | melanogenesis associated transcription factor |
| M-CSF | macrophage colony stimulating factor |
| MMP | matrix metalloproteinase |
| MSCs | mesenchymal stem cells |
| MSDK | melatonin, strontium, vitamin D3 and vitamin K2 |
| NFATC1 | nuclear factor of activated T-cells, cytoplasmic 1 |
| NSAID | nonsteroidal anti-inflammatory drug |
| OC | osteocalcin |
| OPG | osteoprotegerin |
| OSCAR | osteoclast-associated receptor |
| PBMCs | peripheral blood monocytes |
| PDGF | platelet derived growth factor |
| PET-CT | positron emission tomography-computed tomography |
| PICP | procollagen type I carboxy-terminal propeptide |
| PINP | procollagen type I N-terminal propeptide |
| RANK | receptor activator of nuclear factor-kb |
| RANKL | receptor activator of nuclear factor-kb ligand |
| RUNX2 | runt-related transcription factor 2 |
| S1P | sphingosine-1-phosphate |
| SATB2 | special AT-rich sequence-binding protein 2 |
| SEM | scanning electron microscopy |
| SFRP1 | secreted frizzled related protein 1 |
| SOST | gene name for sclerostin |
| Sphk1 | sphingosine kinase 1 |
| SRB | sulforhodamine B |
| TGF-β | transforming growth factor beta |
| THPO | thrombopoietin |
| TRACER | tissue roll for the analysis of cellular environment and response |
| TRAF6 | TNF receptor associated factor 6 |
| TRAP | tartrate-resistant acid phosphatase 5b |
| VitDR | vitamin D receptor |
References
- Hernandez, R.K.; Do, T.P.; Critchlow, C.W.; Dent, R.E.; Jick, S.S. Patient-related risk factors for fracture-healing complications in the United Kingdom General Practice Research Database. Acta Orthop. 2012, 83, 653–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gortler, H.; Rusyn, J.; Godbout, C.; Chahal, J.; Schemitsch, E.H.; Nauth, A. Diabetes and Healing Outcomes in Lower Extremity Fractures: A Systematic Review. Injury 2018, 49, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Jeffcoach, D.R.; Sams, V.G.; Lawson, C.M.; Enderson, B.L.; Smith, S.T.; Kline, H.; Barlow, P.B.; Wylie, D.R.; Krumenacker, L.A.; McMillen, J.C.; et al. Nonsteroidal anti-inflammatory drugs’ impact on nonunion and infection rates in long-bone fractures. J. Trauma. Acute Care Surg. 2014, 76, 779–783. [Google Scholar] [CrossRef] [PubMed]
- Ill, P.O.; Alexandre, C. Tobacco as risk factor of osteoporosis, myth or reality? Rev. Rhum. Ed. Fr. 1993, 60, 280–286. [Google Scholar] [PubMed]
- Pearson, R.G.; Clement, R.G.; Edwards, K.L.; Scammell, B.E. Do smokers have greater risk of delayed and non-union after fracture, osteotomy and arthrodesis? A systematic review with meta-analysis. BMJ Open 2016, 6, e10303. [Google Scholar] [CrossRef] [PubMed]
- Scolaro, J.A.; Schenker, M.L.; Yannascoli, S.; Baldwin, K.; Mehta, S.; Ahn, J. Cigarette smoking increases complications following fracture: A systematic review. J. Bone Jt. Surg. Am. 2014, 96, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Ihle, C.; Freude, T.; Bahrs, C.; Zehendner, E.; Braunsberger, J.; Biesalski, H.K.; Lambert, C.; Stockle, U.; Wintermeyer, E.; Grunwald, J.; et al. Malnutrition—An underestimated factor in the inpatient treatment of traumatology and orthopedic patients: A prospective evaluation of 1055 patients. Injury 2017, 48, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Pirlich, M.; Schutz, T.; Norman, K.; Gastell, S.; Lubke, H.J.; Bischoff, S.C.; Bolder, U.; Frieling, T.; Guldenzoph, H.; Hahn, K.; et al. The German hospital malnutrition study. Clin. Nutr. 2006, 25, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Lambert, C.; Nussler, A.; Biesalski, H.K.; Freude, T.; Bahrs, C.; Ochs, G.; Flesch, I.; Stockle, U.; Ihle, C. Age-dependent risk factors for malnutrition in traumatology and orthopedic patients. Nutrition 2017, 37, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Freude, T.; Ihle, C.; Mayer, L.; Braun, B.; Graeser, J.; Flesch, I.; Stockle, U.; Nussler, A.K.; Pscherer, S. Factors circulating in the blood of type 2 diabetes mellitus patients affect osteoblast maturation—Description of a novel in vitro model. Exp. Cell Res. 2015, 332, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Freude, T.; Braun, K.F.; Haug, A.; Pscherer, S.; Stockle, U.; Nussler, A.K.; Ehnert, S. Hyperinsulinemia reduces osteoblast activity in vitro via upregulation of TGF-β. J. Mol. Med. 2012, 90, 1257–1266. [Google Scholar] [CrossRef] [PubMed]
- Marsell, R.; Einhorn, T.A. The biology of fracture healing. Injury 2011, 42, 551–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghayor, C.; Weber, F.E. Epigenetic Regulation of Bone Remodeling and Its Impacts in Osteoporosis. Int. J. Mol. Sci. 2016, 17, 1446. [Google Scholar] [CrossRef] [PubMed]
- Langdahl, B.; Ferrari, S.; Dempster, D.W. Bone modeling and remodeling: Potential as therapeutic targets for the treatment of osteoporosis. Ther. Adv. Musculoskelet. Dis. 2016, 8, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Kikuta, J.; Ishii, M. Bone Imaging: Osteoclast and Osteoblast Dynamics. Methods Mol. Biol. 2018, 1763, 1–9. [Google Scholar] [PubMed]
- Crockett, J.C.; Rogers, M.J.; Coxon, F.P.; Hocking, L.J.; Helfrich, M.H. Bone remodelling at a glance. J. Cell Sci. 2011, 124, 991–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eriksen, E.F. Cellular mechanisms of bone remodeling. Rev. Endocr. Metab. Disord. 2010, 11, 219–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capulli, M.; Paone, R.; Rucci, N. Osteoblast and osteocyte: Games without frontiers. Arch. Biochem. Biophys. 2014, 561, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.J.; Sims, N.A. Osteoclast-derived activity in the coupling of bone formation to resorption. Trends Mol. Med. 2005, 11, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Pfeilschifter, J.; Mundy, G.R. Modulation of type β transforming growth factor activity in bone cultures by osteotropic hormones. Proc. Natl. Acad. Sci. USA 1987, 84, 2024–2028. [Google Scholar] [CrossRef] [PubMed]
- Mese, G.; Richard, G.; White, T.W. Gap junctions: Basic structure and function. J. Investig. Dermatol. 2007, 127, 2516–2524. [Google Scholar] [CrossRef] [PubMed]
- Laird, D.W. Closing the gap on autosomal dominant connexin-26 and connexin-43 mutants linked to human disease. J. Biol. Chem. 2008, 283, 2997–3001. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, K.; Irie, N. Osteoclast-osteoblast communication. Arch. Biochem. Biophys. 2008, 473, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Fouque-Aubert, A.; Chapurlat, R. Influence of RANKL inhibition on immune system in the treatment of bone diseases. Jt. Bone Spine 2008, 75, 5–10. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Kim, H.J.; Chang, E.J.; Huang, H.; Banno, Y.; Kim, H.H. Sphingosine 1-phosphate as a regulator of osteoclast differentiation and osteoclast-osteoblast coupling. EMBO J. 2006, 25, 5840–5851. [Google Scholar] [CrossRef] [PubMed]
- Graves, D.T.; Jiang, Y.; Valente, A.J. The expression of monocyte chemoattractant protein-1 and other chemokines by osteoblasts. Front. Biosci. 1999, 4, D571–D580. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Baylink, D.J. Bone growth factors. Clin. Orthop. Relat. Res. 1991, 30–48. [Google Scholar] [CrossRef] [PubMed]
- Lacey, D.L.; Timms, E.; Tan, H.L.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Elliott, G.; Scully, S.; et al. Osteoprotegerin ligand is a cytokine that regulates osteoclast differentiation and activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef]
- Shakibaei, M.; Shayan, P.; Busch, F.; Aldinger, C.; Buhrmann, C.; Lueders, C.; Mobasheri, A. Resveratrol mediated modulation of Sirt-1/Runx2 promotes osteogenic differentiation of mesenchymal stem cells: Potential role of Runx2 deacetylation. PLoS ONE 2012, 7, e35712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simonet, W.S.; Lacey, D.L.; Dunstan, C.R.; Kelley, M.; Chang, M.S.; Luthy, R.; Nguyen, H.Q.; Wooden, S.; Bennett, L.; Boone, T.; et al. Osteoprotegerin: A novel secreted protein involved in the regulation of bone density. Cell 1997, 89, 309–319. [Google Scholar] [CrossRef]
- Kitazawa, R.; Haraguchi, R.; Fukushima, M.; Kitazawa, S. Pathologic conditions of hard tissue: Role of osteoclasts in osteolytic lesion. Histochem. Cell Biol. 2018, 149, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, Z.; Duan, N.; Zhu, G.; Schwarz, E.M.; Xie, C. Osteoblast-osteoclast interactions. Connect. Tissue Res. 2018, 59, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Derer, A.; Andes, F.T.; Lezuo, P.; Bozec, A.; Schett, G.; Herrmann, M.; Harre, U. Galectin-3 as a novel regulator of osteoblast-osteoclast interaction and bone homeostasis. Bone 2017, 105, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Xing, J.; Long, C.L.; Peng, Q.; Humphrey, M.B. DOK3 Modulates Bone Remodeling by Negatively Regulating Osteoclastogenesis and Positively Regulating Osteoblastogenesis. J. Bone Miner Res. 2017, 32, 2207–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meijome, T.E.; Baughman, J.T.; Hooker, R.A.; Cheng, Y.H.; Ciovacco, W.A.; Balamohan, S.M.; Srinivasan, T.L.; Chitteti, B.R.; Eleniste, P.P.; Horowitz, M.C.; et al. C-Mpl Is Expressed on Osteoblasts and Osteoclasts and Is Important in Regulating Skeletal Homeostasis. J. Cell Biochem. 2016, 117, 959–969. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.K.; Zhang, M.C. IL-23 promotes osteoclastogenesis in osteoblast-osteoclast co-culture system. Genet. Mol. Res. 2014, 13, 4673–4679. [Google Scholar] [CrossRef] [PubMed]
- Pederson, L.; Ruan, M.; Westendorf, J.J.; Khosla, S.; Oursler, M.J. Regulation of bone formation by osteoclasts involves Wnt/BMP signaling and the chemokine sphingosine-1-phosphate. Proc. Natl. Acad. Sci. USA 2008, 105, 20764–20769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, P.; Lv, H.; Li, Y.; Deng, Y.; Zhang, L.; Tang, P. Exosome-Mediated Genetic Information Transfer, a Missing Piece of Osteoblast-Osteoclast Communication Puzzle. Front. Endocrinol. (Lausanne) 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, C.J.; Beaupre, G.S.; Carter, D.R. A theoretical analysis of the relative influences of peak BMD, age-related bone loss and menopause on the development of osteoporosis. Osteoporos. Int. 2003, 14, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Roeder, I.; Loeffler, M.; Glauche, I. Towards a quantitative understanding of stem cell-niche interaction: Experiments, models, and technologies. Blood Cells Mol. Dis. 2011, 46, 308–317. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.S.; Beers, W.H.; Gilula, N.B. Transmission of hormonal stimulation by cell-to-cell communication. Nature 1978, 272, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Botelho, C.M.; Brooks, R.A.; Best, S.M.; Lopes, M.A.; Santos, J.D.; Rushton, N.; Bonfield, W. Human osteoblast response to silicon-substituted hydroxyapatite. J. Biomed. Mater. Res. A 2006, 79, 723–730. [Google Scholar] [CrossRef] [PubMed]
- Detsch, R.; Mayr, H.; Ziegler, G. Formation of osteoclast-like cells on HA and TCP ceramics. Acta Biomater. 2008, 4, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, C.; Heinemann, S.; Bernhardt, A.; Lode, A.; Worch, H.; Hanke, T. In vitro osteoclastogenesis on textile chitosan scaffold. Eur. Cell Mater. 2010, 19, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Narducci, P.; Nicolin, V. Differentiation of activated monocytes into osteoclast-like cells on a hydroxyapatite substrate: An in vitro study. Ann. Anat. 2009, 191, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Lindstrom, S.; Andersson-Svahn, H. Single-cell culture in microwells. Methods Mol. Biol. 2012, 853, 41–52. [Google Scholar] [PubMed]
- Jones, G.L.; Motta, A.; Marshall, M.J.; El, H.A.; Cartmell, S.H. Osteoblast: Osteoclast co-cultures on silk fibroin, chitosan and PLLA films. Biomaterials 2009, 30, 5376–5384. [Google Scholar] [CrossRef] [PubMed]
- Orlandini, S.Z.; Formigli, L.; Benvenuti, S.; Lasagni, L.; Franchi, A.; Masi, L.; Bernabei, P.A.; Santini, V.; Brandi, M.L. Functional and structural interactions between osteoblastic and preosteoclastic cells in vitro. Cell Tissue Res. 1995, 281, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Bogdanowicz, D.R.; Lu, H.H. Multifunction co-culture model for evaluating cell-cell interactions. Methods Mol. Biol. 2014, 1202, 29–36. [Google Scholar] [PubMed]
- Boanini, E.; Torricelli, P.; Gazzano, M.; Fini, M.; Bigi, A. The effect of zoledronate-hydroxyapatite nanocomposites on osteoclasts and osteoblast-like cells in vitro. Biomaterials 2012, 33, 722–730. [Google Scholar] [CrossRef] [PubMed]
- Sreekumar, V.; Aspera-Werz, R.H.; Tendulkar, G.; Reumann, M.K.; Freude, T.; Breitkopf-Heinlein, K.; Dooley, S.; Pscherer, S.; Ochs, B.G.; Flesch, I.; et al. BMP9 a possible alternative drug for the recently withdrawn BMP7? New perspectives for (re-)implementation by personalized medicine. Arch. Toxicol. 2017, 91, 1353–1366. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Aspera-Werz, R.H.; Freude, T.; Reumann, M.K.; Ochs, B.G.; Bahrs, C.; Schroter, S.; Wintermeyer, E.; Nussler, A.K.; Pscherer, S. Distinct Gene Expression Patterns Defining Human Osteoblasts’ Response to BMP2 Treatment: Is the Therapeutic Success All a Matter of Timing? Eur. Surg. Res. 2016, 57, 197–210. [Google Scholar] [CrossRef] [PubMed]
- Cenni, E.; Perut, F.; Baldini, N. In vitro models for the evaluation of angiogenic potential in bone engineering. Acta Pharmacol. Sin. 2011, 32, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Mo, G.Y.; Zhang, S.C.; Li, Y.X.; Guo, H.Z.; Guo, D.Q.; Li, D.X.; Tang, Y.C.; Mo, L.; Luo, P.J.; Ma, Y.H. Establish mouse osteoblast -osteoclast cell co-culture system in a Transwell chamber. Zhongguo Gu Shang 2018, 31, 241–247. [Google Scholar] [PubMed]
- Kim, B.S.; Lee, J.S.; Gao, G.; Cho, D.W. Direct 3D cell-printing of human skin with functional transwell system. Biofabrication 2017, 9, 25034. [Google Scholar] [CrossRef] [PubMed]
- Thayanithy, V.; O’Hare, P.; Wong, P.; Zhao, X.; Steer, C.J.; Subramanian, S.; Lou, E. A transwell assay that excludes exosomes for assessment of tunneling nanotube-mediated intercellular communication. Cell Commun. Signal 2017, 15, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Barrios, M.P.; Alani, R.M.; Cabodi, M.; Wong, J.Y. A microfluidic Transwell to study chemotaxis. Exp. Cell Res. 2016, 342, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, M.P.; Young, H.; Hurlstone, A.; Wellbrock, C. Differentiation of THP1 Cells into Macrophages for Transwell Co-culture Assay with Melanoma Cells. Bio-Protocol 2015, 5, e1638. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; van Griensven, M.; Unger, M.; Scheffler, H.; Falldorf, K.; Fentz, A.K.; Seeliger, C.; Schroter, S.; Nussler, A.K.; Balmayor, E.R. Co-Culture with Human Osteoblasts and Exposure to Extremely Low Frequency Pulsed Electromagnetic Fields Improve Osteogenic Differentiation of Human Adipose-Derived Mesenchymal Stem Cells. Int. J. Mol. Sci. 2018, 19, 994. [Google Scholar] [CrossRef] [PubMed]
- Malekshah, A.K.; Moghaddam, A.E.; Daraka, S.M. Comparison of conditioned medium and direct co-culture of human granulosa cells on mouse embryo development. Indian J. Exp. Biol. 2006, 44, 189–192. [Google Scholar] [PubMed]
- Hess, M.W.; Pfaller, K.; Ebner, H.L.; Beer, B.; Hekl, D.; Seppi, T. 3D versus 2D cell culture implications for electron microscopy. Methods Cell Biol. 2010, 96, 649–670. [Google Scholar] [PubMed]
- Boiron, M.; Guillemain, B.; Bernard, C.; Peries, J.; Chuat, J.C. Presence in murine sarcoma virus stocks of a 3d component which alone initiates cellular conversion. Nature 1968, 219, 748–749. [Google Scholar] [CrossRef] [PubMed]
- Schyschka, L.; Sanchez, J.J.; Wang, Z.; Burkhardt, B.; Muller-Vieira, U.; Zeilinger, K.; Bachmann, A.; Nadalin, S.; Damm, G.; Nussler, A.K. Hepatic 3D cultures but not 2D cultures preserve specific transporter activity for acetaminophen-induced hepatotoxicity. Arch. Toxicol. 2013, 87, 1581–1593. [Google Scholar] [CrossRef] [PubMed]
- Duval, K.; Grover, H.; Han, L.H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2D vs. 3D Cell Culture. Physiology (Bethesda) 2017, 32, 266–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, K.; Lin-Gibson, S.; Wallace, W.E.; Parekh, S.H.; Lee, Y.J.; Cicerone, M.T.; Young, M.F.; Simon, C.J. The effect of 3D hydrogel scaffold modulus on osteoblast differentiation and mineralization revealed by combinatorial screening. Biomaterials 2010, 31, 5051–5062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benson, K.; Galla, H.J.; Kehr, N.S. Cell adhesion behavior in 3D hydrogel scaffolds functionalized with d- or l-aminoacids. Macromol. Biosci. 2014, 14, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Sung, W.C.; Chen, H.H.; Makamba, H.; Chen, S.H. Functionalized 3D-hydrogel plugs covalently patterned inside hydrophilic poly(dimethylsiloxane) microchannels for flow-through immunoassays. Anal. Chem. 2009, 81, 7967–7973. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; Boone, J.; Wie, J.; Yip, K.P.; Zhang, J.; Wang, L.; Liu, R. Application of Hanging Drop Technique for Kidney Tissue Culture. Kidney Blood Press. Res. 2017, 42, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Amirpour, N.; Razavi, S.; Esfandiari, E.; Hashemibeni, B.; Kazemi, M.; Salehi, H. Hanging drop culture enhances differentiation of human adipose-derived stem cells into anterior neuroectodermal cells using small molecules. Int. J. Dev. Neurosci. 2017, 59, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Ditadi, A.; Sturgeon, C.M.; Keller, G. A view of human haematopoietic development from the Petri dish. Nat. Rev. Mol. Cell Biol. 2017, 18, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Loh, Q.L.; Choong, C. Three-dimensional scaffolds for tissue engineering applications: Role of porosity and pore size. Tissue Eng. Part B Rev. 2013, 19, 485–502. [Google Scholar] [CrossRef] [PubMed]
- Holzapfel, B.M.; Rudert, M.; Hutmacher, D.W. Scaffold-based Bone Tissue Engineering. Orthopade 2017, 46, 701–710. [Google Scholar] [CrossRef] [PubMed]
- Gomez, S.; Vlad, M.D.; Lopez, J.; Fernandez, E. Design and properties of 3D scaffolds for bone tissue engineering. Acta Biomater. 2016, 42, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, M.; Steimberg, N.; Boniotti, J.; Berenzi, A.; Belloni, D.; Mazzoleni, G.; Ferrero, E. 3D-Dynamic Culture Models of Multiple Myeloma. Methods Mol. Biol. 2017, 1612, 177–190. [Google Scholar] [PubMed]
- Tang, Y.; Xu, Y.; Xiao, Z.; Zhao, Y.; Li, J.; Han, S.; Chen, L.; Dai, B.; Wang, L.; Chen, B.; et al. The combination of three-dimensional and rotary cell culture system promotes the proliferation and maintains the differentiation potential of rat BMSCs. Sci. Rep. 2017, 7, 192. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Ono, K.; Hata, S.; Suzuki, T.; Kumamoto, H.; Sasano, H. The advantages of co-culture over mono cell culture in simulating in vivo environment. J. Steroid. Biochem. Mol. Biol. 2012, 131, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Young, M.; Rodenhizer, D.; Dean, T.; D’Arcangelo, E.; Xu, B.; Ailles, L.; McGuigan, A.P. A TRACER 3D Co-Culture tumour model for head and neck cancer. Biomaterials 2018, 164, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Foster, N.C.; Henstock, J.R.; Reinwald, Y.; El, H.A. Dynamic 3D culture: Models of chondrogenesis and endochondrall ossification. Birth Defects Res. C Embryo Today 2015, 105, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Animal models for osteoporosis. Eur. J. Pharmacol. 2015, 759, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochrath, K.; Ehnert, S.; Ackert-Bicknell, C.L.; Lau, Y.; Schmid, A.; Krawczyk, M.; Hengstler, J.G.; Dunn, J.; Hiththetiya, K.; Rathkolb, B.; et al. Modeling hepatic osteodystrophy in Abcb4 deficient mice. Bone 2013, 55, 501–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nussler, A.K.; Wildemann, B.; Freude, T.; Litzka, C.; Soldo, P.; Friess, H.; Hammad, S.; Hengstler, J.G.; Braun, K.F.; Trak-Smayra, V.; et al. Chronic CCl4 intoxication causes liver and bone damage similar to the human pathology of hepatic osteodystrophy: A mouse model to analyse the liver-bone axis. Arch. Toxicol. 2014, 88, 997–1006. [Google Scholar] [CrossRef] [PubMed]
- Goers, L.; Freemont, P.; Polizzi, K.M. Co-culture systems and technologies: Taking synthetic biology to the next level. J. R. Soc. Interface 2014, 11, 20140065. [Google Scholar] [CrossRef] [PubMed]
- Zehnder, T.; Boccaccini, A.R.; Detsch, R. Biofabrication of a co-culture system in an osteoid-like hydrogel matrix. Biofabrication 2017, 9, 25016. [Google Scholar] [CrossRef] [PubMed]
- Dahlin, R.L.; Gershovich, J.G.; Kasper, F.K.; Mikos, A.G. Flow perfusion co-culture of human mesenchymal stem cells and endothelial cells on biodegradable polymer scaffolds. Ann. Biomed. Eng. 2014, 42, 1381–1390. [Google Scholar] [CrossRef] [PubMed]
- Bouet, G.; Cruel, M.; Laurent, C.; Vico, L.; Malaval, L.; Marchat, D. Validation of an in vitro 3D bone culture model with perfused and mechanically stressed ceramic scaffold. Eur. Cell Mater. 2015, 29, 250–266. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Eckert, H.; Hess, R.; Gelinsky, M.; Rancourt, D.; Krawetz, R.; Cuniberti, G.; Scharnweber, D. Developing a Customized Perfusion Bioreactor Prototype with Controlled Positional Variability in Oxygen Partial Pressure for Bone and Cartilage Tissue Engineering. Tissue Eng. Part C Methods 2017, 23, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Bongio, M.; Lopa, S.; Gilardi, M.; Bersini, S.; Moretti, M. A 3D vascularized bone remodeling model combining osteoblasts and osteoclasts in a CaP nanoparticle-enriched matrix. Nanomedicine (Lond.) 2016, 11, 1073–1091. [Google Scholar] [CrossRef] [PubMed]
- Kleinhans, C.; Schmid, F.F.; Schmid, F.V.; Kluger, P.J. Comparison of osteoclastogenesis and resorption activity of human osteoclasts on tissue culture polystyrene and on natural extracellular bone matrix in 2D and 3D. J. Biotechnol. 2015, 205, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, V.; Vogler, E.A.; Mastro, A.M. Three-Dimensional in Vitro Model to Study Osteobiology and Osteopathology. J. Cell Biochem. 2015, 116, 2715–2723. [Google Scholar] [CrossRef] [PubMed]
- Shor, L.; Guceri, S.; Wen, X.; Gandhi, M.; Sun, W. Fabrication of three-dimensional polycaprolactone/hydroxyapatite tissue scaffolds and osteoblast-scaffold interactions in vitro. Biomaterials 2007, 28, 5291–5297. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Dong, C.; Yang, L.; Lv, Y. 3D Scaffolds with Different Stiffness but the Same Microstructure for Bone Tissue Engineering. ACS Appl. Mater. Interfaces 2015, 7, 15790–15802. [Google Scholar] [CrossRef] [PubMed]
- Tsuruga, E.; Takita, H.; Itoh, H.; Wakisaka, Y.; Kuboki, Y. Pore size of porous hydroxyapatite as the cell-substratum controls BMP-induced osteogenesis. J. Biochem. 1997, 121, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Kuhne, J.H.; Bartl, R.; Frisch, B.; Hammer, C.; Jansson, V.; Zimmer, M. Bone formation in coralline hydroxyapatite. Effects of pore size studied in rabbits. Acta Orthop. Scand. 1994, 65, 246–252. [Google Scholar] [CrossRef] [PubMed]
- Mastrogiacomo, M.; Scaglione, S.; Martinetti, R.; Dolcini, L.; Beltrame, F.; Cancedda, R.; Quarto, R. Role of scaffold internal structure on in vivo bone formation in macroporous calcium phosphate bioceramics. Biomaterials 2006, 27, 3230–3237. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Yeatts, A.; Dean, D.; Fisher, J.P. Stereolithographic bone scaffold design parameters: Osteogenic differentiation and signal expression. Tissue Eng. Part B Rev. 2010, 16, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Huebsch, N.; Arany, P.R.; Mao, A.S.; Shvartsman, D.; Ali, O.A.; Bencherif, S.A.; Rivera-Feliciano, J.; Mooney, D.J. Harnessing traction-mediated manipulation of the cell/matrix interface to control stem-cell fate. Nat. Mater. 2010, 9, 518–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engler, A.J.; Sen, S.; Sweeney, H.L.; Discher, D.E. Matrix elasticity directs stem cell lineage specification. Cell 2006, 126, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Bellis, S.L. Advantages of RGD peptides for directing cell association with biomaterials. Biomaterials 2011, 32, 4205–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greiner, S.; Kadow-Romacker, A.; Schmidmaier, G.; Wildemann, B. Cocultures of osteoblasts and osteoclasts are influenced by local application of zoledronic acid incorporated in a poly(d,l-lactide) implant coating. J. Biomed. Mater. Res. A 2009, 91, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Paul, B.; Gelinsky, M. Biphasic Scaffolds from Marine Collagens for Regeneration of Osteochondral Defects. Mar. Drugs 2018, 16, 91. [Google Scholar] [CrossRef] [PubMed]
- Lode, A.; Meyer, M.; Bruggemeier, S.; Paul, B.; Baltzer, H.; Schropfer, M.; Winkelmann, C.; Sonntag, F.; Gelinsky, M. Additive manufacturing of collagen scaffolds by three-dimensional plotting of highly viscous dispersions. Biofabrication 2016, 8, 15015. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.S.; Sundaresan, A.; Vanderburg, C.R.; Banigan, M.G.; Pellis, N.R. A three-dimensional tissue culture model of bone formation utilizing rotational co-culture of human adult osteoblasts and osteoclasts. Acta Biomater. 2013, 9, 7908–7916. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Thieme, S.; Domaschke, H.; Springer, A.; Rosen-Wolff, A.; Gelinsky, M. Crosstalk of osteoblast and osteoclast precursors on mineralized collagen--towards an in vitro model for bone remodeling. J. Biomed. Mater. Res. A 2010, 95, 848–856. [Google Scholar] [CrossRef] [PubMed]
- Heinemann, C.; Heinemann, S.; Worch, H.; Hanke, T. Development of an osteoblast/osteoclast co-culture derived by human bone marrow stromal cells and human monocytes for biomaterials testing. Eur. Cell Mater. 2011, 21, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Lutter, A.H.; Hempel, U.; Wolf-Brandstetter, C.; Garbe, A.I.; Goettsch, C.; Hofbauer, L.C.; Jessberger, R.; Dieter, P. A novel resorption assay for osteoclast functionality based on an osteoblast-derived native extracellular matrix. J. Cell Biochem. 2010, 109, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Kadow-Romacker, A.; Hoffmann, J.E.; Duda, G.; Wildemann, B.; Schmidmaier, G. Effect of mechanical stimulation on osteoblast- and osteoclast-like cells in vitro. Cells Tissues Organs 2009, 190, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Kadow-Romacker, A.; Duda, G.N.; Bormann, N.; Schmidmaier, G.; Wildemann, B. Slight changes in the mechanical stimulation affects osteoblast- and osteoclast-like cells in co-culture. Transfus. Med. Hemother. 2013, 40, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Spano, A.; Barni, S.; Sciola, L. PMA withdrawal in PMA-treated monocytic THP-1 cells and subsequent retinoic acid stimulation, modulate induction of apoptosis and appearance of dendritic cells. Cell Prolif. 2013, 46, 328–347. [Google Scholar] [CrossRef] [PubMed]
- Schulze, S.; Wehrum, D.; Dieter, P.; Hempel, U. A supplement-free osteoclast-osteoblast co-culture for pre-clinical application. J. Cell. Physiol. 2018, 233, 4391–4400. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Birmingham, E.; Niebur, G.L.; McHugh, P.E.; Shaw, G.; Barry, F.P.; McNamara, L.M. Osteogenic differentiation of mesenchymal stem cells is regulated by osteocyte and osteoblast cells in a simplified bone niche. Eur. Cell Mater. 2012, 23, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Musina, R.A.; Bekchanova, E.S.; Belyavskii, A.V.; Sukhikh, G.T. Differentiation potential of mesenchymal stem cells of different origin. Bull. Exp. Biol. Med. 2006, 141, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Jeon, O.H.; Panicker, L.M.; Lu, Q.; Chae, J.J.; Feldman, R.A.; Elisseeff, J.H. Human iPSC-derived osteoblasts and osteoclasts together promote bone regeneration in 3D biomaterials. Sci. Rep. 2016, 6, 26761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, X. Induced Pluripotent Stem Cells as a new Strategy for Osteogenesis and Bone Regeneration. Stem Cell Rev. 2015, 11, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Chen, W.; Liu, J.; Weir, M.D.; Cheng, L.; Xu, H.H. Human induced pluripotent stem cell-derived mesenchymal stem cell seeding on calcium phosphate scaffold for bone regeneration. Tissue Eng. Part A 2014, 20, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Chen, W.; Zhao, Z.; Xu, H.H. Reprogramming of mesenchymal stem cells derived from iPSCs seeded on biofunctionalized calcium phosphate scaffold for bone engineering. Biomaterials 2013, 34, 7862–7872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cell Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Miki, Y.; Suzuki, T.; Tazawa, C.; Yamaguchi, Y.; Kitada, K.; Honma, S.; Moriya, T.; Hirakawa, H.; Evans, D.B.; Hayashi, S.; et al. Aromatase localization in human breast cancer tissues: Possible interactions between intratumoral stromal and parenchymal cells. Cancer Res. 2007, 67, 3945–3954. [Google Scholar] [CrossRef] [PubMed]
- Burdall, S.E.; Hanby, A.M.; Lansdown, M.R.; Speirs, V. Breast cancer cell lines: Friend or foe? Breast Cancer Res. 2003, 5, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Mohseny, A.B.; Machado, I.; Cai, Y.; Schaefer, K.L.; Serra, M.; Hogendoorn, P.C.; Llombart-Bosch, A.; Cleton-Jansen, A.M. Functional characterization of osteosarcoma cell lines provides representative models to study the human disease. Lab. Investig. 2011, 91, 1195–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.H.; Si, Y.; Xu, G.; Chen, X.M.; Xiong, H.; Lai, L.; Zheng, Y.Q.; Zhang, Z.G. High-dose PMA with RANKL and MCSF induces THP1 cell differentiation into human functional osteoclasts in vitro. Mol. Med. Rep. 2017, 16, 8380–8384. [Google Scholar] [CrossRef] [PubMed]
- Lauvrak, S.U.; Munthe, E.; Kresse, S.H.; Stratford, E.W.; Namlos, H.M.; Meza-Zepeda, L.A.; Myklebost, O. Functional characterisation of osteosarcoma cell lines and identification of mRNAs and miRNAs associated with aggressive cancer phenotypes. Br. J. Cancer 2013, 109, 2228–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottaviano, L.; Schaefer, K.L.; Gajewski, M.; Huckenbeck, W.; Baldus, S.; Rogel, U.; Mackintosh, C.; de Alava, E.; Myklebost, O.; Kresse, S.H.; et al. Molecular characterization of commonly used cell lines for bone tumor research: A trans-European EuroBoNet effort. Genes Chromosomes. Cancer 2010, 49, 40–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pantalone, A.; Antonucci, I.; Guelfi, M.; Pantalone, P.; Usuelli, F.G.; Stuppia, L.; Salini, V. Amniotic fluid stem cells: An ideal resource for therapeutic application in bone tissue engineering. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2884–2890. [Google Scholar] [PubMed]
- Yoshimura, Y.; Kikuiri, T.; Hasegawa, T.; Matsuno, M.; Minamikawa, H.; Deyama, Y.; Suzuki, K. How much medium do you use for cell culture? Medium volume influences mineralization and osteoclastogenesis in vitro. Mol. Med. Rep. 2017, 16, 429–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tendulkar, G.; Grau, P.; Ziegler, P.; Buck, A.S.; Buck, A.J.; Badke, A.; Kaps, H.P.; Ehnert, S.; Nussler, A.K. Imaging Cell Viability on Non-transparent Scaffolds—Using the Example of a Novel Knitted Titanium Implant. J. Vis. Exp. 2016, 115. [Google Scholar] [CrossRef]
- Jolly, J.J.; Chin, K.Y.; Farhana, M.; Alias, E.; Chua, K.H.; Hasan, W.; Ima-Nirwana, S. Optimization of the Static Human Osteoblast/Osteoclast Co-culture System. Iran. J. Med. Sci. 2018, 43, 208–213. [Google Scholar] [PubMed]
- Bartelt, A.; Behler-Janbeck, F.; Beil, F.T.; Koehne, T.; Muller, B.; Schmidt, T.; Heine, M.; Ochs, L.; Yilmaz, T.; Dietrich, M.; et al. Lrp1 in osteoblasts controls osteoclast activity and protects against osteoporosis by limiting PDGF-RANKL signaling. Bone Res. 2018, 6, 4. [Google Scholar] [CrossRef] [PubMed]
- Prento, P. Van Gieson’s picrofuchsin. The staining mechanisms for collagen and cytoplasm, and an examination of the dye diffusion rate model of differential staining. Histochemistry 1993, 99, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Yaccoby, S.; Wezeman, M.J.; Zangari, M.; Walker, R.; Cottler-Fox, M.; Gaddy, D.; Ling, W.; Saha, R.; Barlogie, B.; Tricot, G.; et al. Inhibitory effects of osteoblasts and increased bone formation on myeloma in novel culture systems and a myelomatous mouse model. Haematologica 2006, 91, 192–199. [Google Scholar] [PubMed]
- Rungby, J.; Kassem, M.; Eriksen, E.F.; Danscher, G. The von Kossa reaction for calcium deposits: Silver lactate staining increases sensitivity and reduces background. Histochem. J. 1993, 25, 446–451. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikov, D. Alcian blue/alizarin red staining of cartilage and bone in mouse. Cold Spring Harb. Protoc. 2009, 2009, t5170. [Google Scholar] [CrossRef] [PubMed]
- Filgueira, L. Fluorescence-based staining for tartrate-resistant acidic phosphatase (TRAP) in osteoclasts combined with other fluorescent dyes and protocols. J. Histochem. Cytochem. 2004, 52, 411–414. [Google Scholar] [CrossRef] [PubMed]
- Edsall, S.C.; Franz-Odendaal, T.A. A quick whole-mount staining protocol for bone deposition and resorption. Zebrafish 2010, 7, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Vesprey, A.; Yang, W. Pit Assay to Measure the Bone Resorptive Activity of Bone Marrow-derived Osteoclasts. Bio-Protocol 2016, 6, e1836. [Google Scholar] [CrossRef] [PubMed]
- Domon, T.; Yamazaki, Y.; Fukui, A.; Ohnishi, Y.; Takahashi, S.; Yamamoto, T.; Wakita, M. Ultrastructural study of cell-cell interaction between osteoclasts and osteoblasts/stroma cells in vitro. Ann. Anat. 2002, 184, 221–227. [Google Scholar] [CrossRef]
- Ayobian-Markazi, N.; Fourootan, T.; Kharazifar, M.J. Comparison of cell viability and morphology of a human osteoblast-like cell line (SaOS-2) seeded on various bone substitute materials: An in vitro study. Dent. Res. J. (Isfahan) 2012, 9, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Orellana, E.A.; Kasinski, A.L. Sulforhodamine B (SRB) Assay in Cell Culture to Investigate Cell Proliferation. Bio-Protocol 2016, 6, e1984. [Google Scholar] [CrossRef] [PubMed]
- Pacios, S.; Xiao, W.; Mattos, M.; Lim, J.; Tarapore, R.S.; Alsadun, S.; Yu, B.; Wang, C.Y.; Graves, D.T. Osteoblast Lineage Cells Play an Essential Role in Periodontal Bone Loss Through Activation of Nuclear Factor-Kappa B. Sci. Rep. 2015, 5, 16694. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.P.; Jansen, H.; Doht, S.; Filgueira, L.; Zellweger, R. Immunohistochemical and molecular characterization of the human periosteum. Sci. World J. 2013, 2013, 341078. [Google Scholar] [CrossRef] [PubMed]
- Kuru, L.; Griffiths, G.S.; Petrie, A.; Olsen, I. Alkaline phosphatase activity is upregulated in regenerating human periodontal cells. J. Periodontal Res. 1999, 34, 123–127. [Google Scholar] [CrossRef] [PubMed]
- Ballanti, P.; Minisola, S.; Pacitti, M.T.; Scarnecchia, L.; Rosso, R.; Mazzuoli, G.F.; Bonucci, E. Tartrate-resistant acid phosphate activity as osteoclastic marker: Sensitivity of cytochemical assessment and serum assay in comparison with standardized osteoclast histomorphometry. Osteoporos. Int. 1997, 7, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Reithmeier, A.; Lundback, T.; Haraldsson, M.; Frank, M.; Ek-Rylander, B.; Nyholm, P.G.; Gustavsson, A.L.; Andersson, G. Identification of inhibitors of Tartrate-resistant acid phosphatase (TRAP/ACP5) activity by small-molecule screening. Chem. Biol. Drug Des. 2018, 92, 1255–1271. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, A.; Koperski, K.; Schumacher, M.; Gelinsky, M. Relevance of osteoclast-specific enzyme activities in cell-based in vitro resorption assays. Eur. Cell Mater. 2017, 33, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Yang, X.; Chen, Y.; Zhai, J.; Liang, X. Effect of titanium particles on osteoclast activity in vitro. Mol. Med. Rep. 2010, 3, 1065–1069. [Google Scholar] [PubMed] [Green Version]
- Fu, Y.X.; Gu, J.H.; Zhang, Y.R.; Tong, X.S.; Zhao, H.Y.; Yuan, Y.; Liu, X.Z.; Bian, J.C.; Liu, Z.P. Osteoprotegerin influences the bone resorption activity of osteoclasts. Int. J. Mol. Med. 2013, 31, 1411–1417. [Google Scholar] [CrossRef] [PubMed]
- de Vrieze, E.; Sharif, F.; Metz, J.R.; Flik, G.; Richardson, M.K. Matrix metalloproteinases in osteoclasts of ontogenetic and regenerating zebrafish scales. Bone 2011, 48, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Ohshiba, T.; Miyaura, C.; Inada, M.; Ito, A. Role of RANKL-induced osteoclast formation and MMP-dependent matrix degradation in bone destruction by breast cancer metastasis. Br. J. Cancer 2003, 88, 1318–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Lin, Y.; Xiao, L.; Feng, E.; Wang, W.; Lin, L. The effects of icariine concentration on osteoclasts bone resorption induced by titanium particles in vitro. Regen. Biomater. 2015, 2, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillian, M.K.; Li, L.; Parker, J.B.; Patel, L.; Zhong, Z.; Gunnett, J.W.; Powers, W.J.; Johnson, M.D. An improved resazurin-based cytotoxicity assay for hepatic cells. Cell Biol. Toxicol. 2002, 18, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Legrand, C.; Bour, J.M.; Jacob, C.; Capiaumont, J.; Martial, A.; Marc, A.; Wudtke, M.; Kretzmer, G.; Demangel, C.; Duval, D.; et al. Lactate dehydrogenase (LDH) activity of the cultured eukaryotic cells as marker of the number of dead cells in the medium [corrected]. J. Biotechnol. 1992, 25, 231–243. [Google Scholar] [CrossRef]
- Solberg, L.B.; Stang, E.; Brorson, S.H.; Andersson, G.; Reinholt, F.P. Tartrate-resistant acid phosphatase (TRAP) co-localizes with receptor activator of NF-KB ligand (RANKL) and osteoprotegerin (OPG) in lysosomal-associated membrane protein 1 (LAMP1)-positive vesicles in rat osteoblasts and osteocytes. Histochem. Cell Biol. 2015, 143, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Ruggiu, A.; Cancedda, R. Bone mechanobiology, gravity and tissue engineering: Effects and insights. J. Tissue Eng. Regen. Med. 2015, 9, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- Metscher, B.D. MicroCT for comparative morphology: Simple staining methods allow high-contrast 3D imaging of diverse non-mineralized animal tissues. BMC Physiol. 2009, 9, 11. [Google Scholar] [CrossRef] [PubMed]
- Auzina, D.; Erts, R.; Lejniece, S. Prognostic value of the bone turnover markers in multiple myeloma. Exp. Oncol. 2017, 39, 53–56. [Google Scholar] [PubMed]
- Bauerle, T.; Komljenovic, D.; Semmler, W. Monitoring molecular, functional and morphologic aspects of bone metastases using non-invasive imaging. Curr. Pharm. Biotechnol. 2012, 13, 584–594. [Google Scholar] [CrossRef] [PubMed]
- Chappard, D.; Terranova, L.; Mallet, R.; Mercier, P. 3D Porous Architecture of Stacks of β-TCP Granules Compared with That of Trabecular Bone: A microCT, Vector Analysis, and Compression Study. Front. Endocrinol. (Lausanne) 2015, 6, 161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Winkler, R.; Dukic, M.; Zhao, J.; Plank, H.; Fantner, G.E. Probing the Morphology and Evolving Dynamics of 3D Printed Nanostructures Using High-Speed Atomic Force Microscopy. ACS Appl. Mater. Interfaces 2017, 9, 24456–24461. [Google Scholar] [CrossRef] [PubMed]
- Dubrovin, E.V.; Presnova, G.V.; Rubtsova, M.Y.; Egorov, A.M.; Grigorenko, V.G.; Yaminsky, I.V. The Use of Atomic Force Microscopy for 3D Analysis of Nucleic Acid Hybridization on Microarrays. Acta Naturae 2015, 7, 108–114. [Google Scholar] [PubMed]
- Franziska, V.; Schmid, C.K.F.F. Osteoclast Formation within a Human Co-Culture System on Bone Material as an In Vitro Model for Bone Remodeling Processes. J. Funct. Morphol. Kinesiol. 2018, 3, 17. [Google Scholar] [Green Version]
- Ichimaru, R.; Tominari, T.; Yoshinouchi, S.; Matsumoto, C.; Watanabe, K.; Hirata, M.; Numabe, Y.; Murphy, G.; Nagase, H.; Miyaura, C.; et al. Raloxifene reduces the risk of local alveolar bone destruction in a mouse model of periodontitis combined with systemic postmenopausal osteoporosis. Arch. Oral. Biol. 2018, 85, 98–103. [Google Scholar] [CrossRef] [PubMed]
- Sale, J.E.; Beaton, D.; Bogoch, E. Secondary prevention after an osteoporosis-related fracture: An overview. Clin. Geriatr. Med. 2014, 30, 317–332. [Google Scholar] [CrossRef] [PubMed]
- Mandatori, D.; Penolazzi, L.; Pipino, C.; Di Tomo, P.; Di Silvestre, S.; Di Pietro, N.; Trevisani, S.; Angelozzi, M.; Ucci, M.; Piva, R.; et al. Menaquinone-4 enhances osteogenic potential of human amniotic fluid mesenchymal stem cells cultured in 2D and 3D dynamic culture systems. J. Tissue Eng. Regen. Med. 2018, 12, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Maria, S.; Swanson, M.H.; Enderby, L.T.; D’Amico, F.; Enderby, B.; Samsonraj, R.M.; Dudakovic, A.; van Wijnen, A.J.; Witt-Enderby, P.A. Melatonin-micronutrients Osteopenia Treatment Study (MOTS): A translational study assessing melatonin, strontium (citrate), vitamin D3 and vitamin K2 (MK7) on bone density, bone marker turnover and health related quality of life in postmenopausal osteopenic women following a one-year double-blind RCT and on osteoblast-osteoclast co-cultures. Aging (Albany NY) 2017, 9, 256–285. [Google Scholar] [PubMed]
- Maria, S.; S amsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for MT2 melatonin receptors, MEK1/2, and MEK5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, M.N.; Shukla, P.; Kakaji, M.; Tyagi, A.M.; S rivastava, K.; Shukla, M.; Dixit, M.; Kureel, J.; Gupta, S.; Singh, D. IL-18BP is decreased in osteoporotic women: Prevents Inflammasome mediated IL-18 activation and reduces Th17 differentiation. Sci. Rep. 2016, 6, 33680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murakami, K.; Kobayashi, Y.; Uehara, S.; Suzuki, T.; Koide, M.; Yamashita, T.; Nakamura, M.; Takahashi, N.; Kato, H.; Udagawa, N.; et al. A Jak1/2 inhibitor, baricitinib, inhibits osteoclastogenesis by suppressing RANKL expression in osteoblasts in vitro. PLoS ONE 2017, 12, e181126. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; He, X.; Yang, Y.; Li, M.; Hao, D.; Jia, Z. The genus Epimedium: An ethnopharmacological and phytochemical review. J. Ethnopharmacol. 2011, 134, 519–541. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lee, L.; Gong, Y.; Shen, P.; Wong, S.P.; Wise, S.D.; Yong, E.L. Bioassays for estrogenic activity: Development and validation of estrogen receptor (ERα/ERβ) and breast cancer proliferation bioassays to measure serum estrogenic activity in clinical studies. Assay Drug Dev. Technol. 2009, 7, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Penolazzi, L.; Lolli, A.; Sardelli, L.; Angelozzi, M.; Lambertini, E.; Trombelli, L.; Ciarpella, F.; Vecchiatini, R.; Piva, R. Establishment of a 3D-dynamic osteoblasts-osteoclasts co-culture model to simulate the jawbone microenvironment in vitro. Life Sci. 2016, 152, 82–93. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Figure | Advantages | Disadvantages | Ref. | |
|---|---|---|---|---|
| conditioned medium |  |
|
| [51,52,53] |
| transwell co-culture |  |
|
| [54,55,56,57,58] |
| removable permeable divider |  |
|
| [49] |
| direct co-culture |  |
|
| [47,48,59,60] |
| Category | Description | Advantages | Disadvantages | Ref. | |
|---|---|---|---|---|---|
| hydrogels | natural hydrogel | water-swollen and cross-linked polymer network made from naturally occurring monomers |
|
| [64,65,66] |
| synthetic hydrogel | water-swollen and cross-linked polymer network from synthetic monomers |
|
| ||
| cell aggregation | hanging drop plates | special plates that allow consistent and controllable cell-aggregation into 3D spheroids |
|
| [68,69,70] |
| 3D petri dish | molds made of agarose that favor agglomeration of cells in a defined shape |
|
| ||
| scaffolds | natural scaffolds | 3D (polymer) matrix made of naturally occurring extracellular matrix |
|
| [71,72,73] |
| synthetic scaffolds | 3D (polymer) matrix made of different synthetic materials |
|
| ||
| dynamic models | often a 3D culture within a bioreactor to provide medium exchange and/or mechanical stimulation |
|
| [74,75] | |
| Cell Types | Advantages | Disadvantages | Ref. | |
|---|---|---|---|---|
| osteoblastic lineage | primary animal osteoblasts and MSCs |
|
| [118,119] |
| primary human osteoblasts and MSCs |
|
| [59,109,125] | |
| osteogenic cell lines |
|
| [120,121,123,124] | |
| osteoclastic lineage | human induced pluripotent stem cells (hiPSCs) |
|
| [114,115] |
| osteoclasts derived from animal monocytes |
|
| [118,119] | |
| osteoclasts derived from human monocytes |
|
| [59,109,125] | |
| osteoclasts derived from myeolytic (monocyte- or macrophage-like) cell lines |
|
| [120,121,123,124] | |
| Visual/Microscopic Methods | |||
|---|---|---|---|
| Methods | Use | Limitations | Ref. |
| van Gieson staining | histological method using picric acid and acid fuchsin to detect collagen |
| [128,129,130] |
| von Kossa staining | silver ions react with phosphates to demonstrate calcium phosphates |
| [59,114,131,132] |
| Alizarin Red staining | An anthraquinone dye to detect presence of calcium ions. Resolving the stain enables its quantification |
| [59,128,133] |
| TRAP staining | staining substrate (colorimetric or fluorogenic) is converted by TRAP to identify osteoclasts |
| [134,135] |
| AP staining | staining substrate (colorimetric or fluorogenic) is converted by AP to identify differentiating osteoblasts | [135] | |
| Pit assay | staining of resorption pits left by osteoclast on dentine chips. Shows bone resorption activity |
| [136] |
| SEM images | SEM can be used to analyze morphological characteristic of cells and 3D matrices |
| [137,138] |
| SRB staining | SRB binds to protonated amino- acids, which can be used to quantify total protein content and thus to determine the cell density |
| [59,139] |
| nuclear staining | A large variety of colorimetric (trypan blue, hematoxylin, etc.) and fluorescent (DAPI, Hoechst 33342, propidium iodide, SYTOX green, ethidium homodimer, etc.) |
| [140,141] |
| Functional Assays | |||
| Methods | Descriptions | Limitations | Ref. |
| AP activity | colorimetric or fluorogenic substrate is converted by osteoblastic AP |
| [138,142] |
| TRAP activity | colorimetric or fluorogenic substrate is converted by osteoclastic TRAP | [143,144] | |
| CTSK activity | colorimetric or fluorogenic substrate is converted by osteoclastic CTSK | [145] | |
| CAII activity | colorimetric or fluorogenic substrate is converted by osteoclastic CAII | [146,147] | |
| zymo- graphy | method to detect proteolytic enzymes, which get separated by non-denaturing gel electrophoresis |
| [148,149] |
| MTT/XTT assay | tetrazolium salts are reduced in mitochondria of viable cells (toxic), often used for normalization |
| [150] |
| Resazurin conversion | Resazurin (non-toxic) is reduced to the Resorufin in viable cells, which is often used for normalization | [151] | |
| LDH activity | LDH enzyme, located in the cytosol of most cells, leaks into the culture supernatant when cell membranes get damaged |
| [105,152] |
| Immunological Methods | |||
| Methods | Descriptions | Limitations | Ref. |
| flow cytometry | cells in a single cells suspension can be characterized based on their size, granularity and chosen stain (see figure on molecular markers) |
| [110] |
| Enzyme-Linked Immuno-Sorbent Assays | ELISA are used to quantify soluble target proteins, e.g., PICP; PINP; DPD; Pi; CTX; NTX; PYD; sclerostin; DKK1&2; MMPs; TIMPs; osteocalcin |
| [44,110] |
| immuno-stainings | antibodies are used to detect target proteins (see figure on molecular markers) in fixed cells; secondary antibodies for detection are either HRP- or fluorophore-labeled |
| [153] |
| Radiological Methods | |||
| Methods | Descriptions | Limitations | Ref. |
| X-ray images | 2D overview image visualizing the mineralized matrix, without destroying the original object |
| [154] |
| (micro-)CT images | visualization of mineralized matrix, without destroying the original object, in quantitative 3D images | [155,156,157] | |
| PET-CT images | combination of the (micro-)CT technique with specific markers | [156,157] | |
| Stiffness Measurements | |||
| Methods | Descriptions | Limitations | Ref. |
| compression tests | mechanical tests to determine the stiffness of a 3D matrix, which changes when cells produce/incorporate matrix during culture |
| [158] |
| atomic force microscopy | [159,160] | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, S.; Ehnert, S.; Rouß, M.; Häussling, V.; Aspera-Werz, R.H.; Chen, T.; Nussler, A.K. From the Clinical Problem to the Basic Research—Co-Culture Models of Osteoblasts and Osteoclasts. Int. J. Mol. Sci. 2018, 19, 2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082284
Zhu S, Ehnert S, Rouß M, Häussling V, Aspera-Werz RH, Chen T, Nussler AK. From the Clinical Problem to the Basic Research—Co-Culture Models of Osteoblasts and Osteoclasts. International Journal of Molecular Sciences. 2018; 19(8):2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082284
Chicago/Turabian StyleZhu, Sheng, Sabrina Ehnert, Marc Rouß, Victor Häussling, Romina H. Aspera-Werz, Tao Chen, and Andreas K. Nussler. 2018. "From the Clinical Problem to the Basic Research—Co-Culture Models of Osteoblasts and Osteoclasts" International Journal of Molecular Sciences 19, no. 8: 2284. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082284