Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
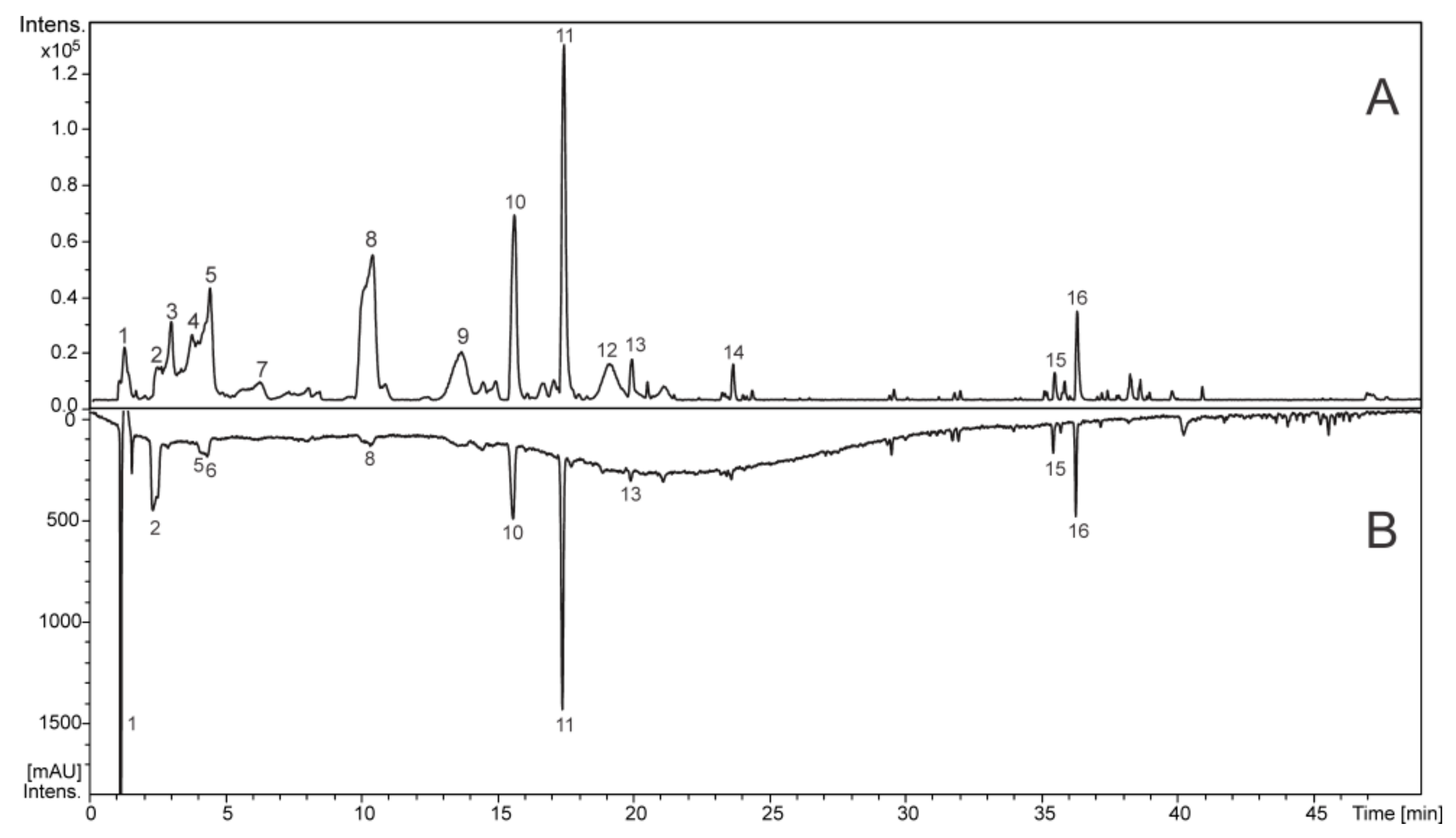
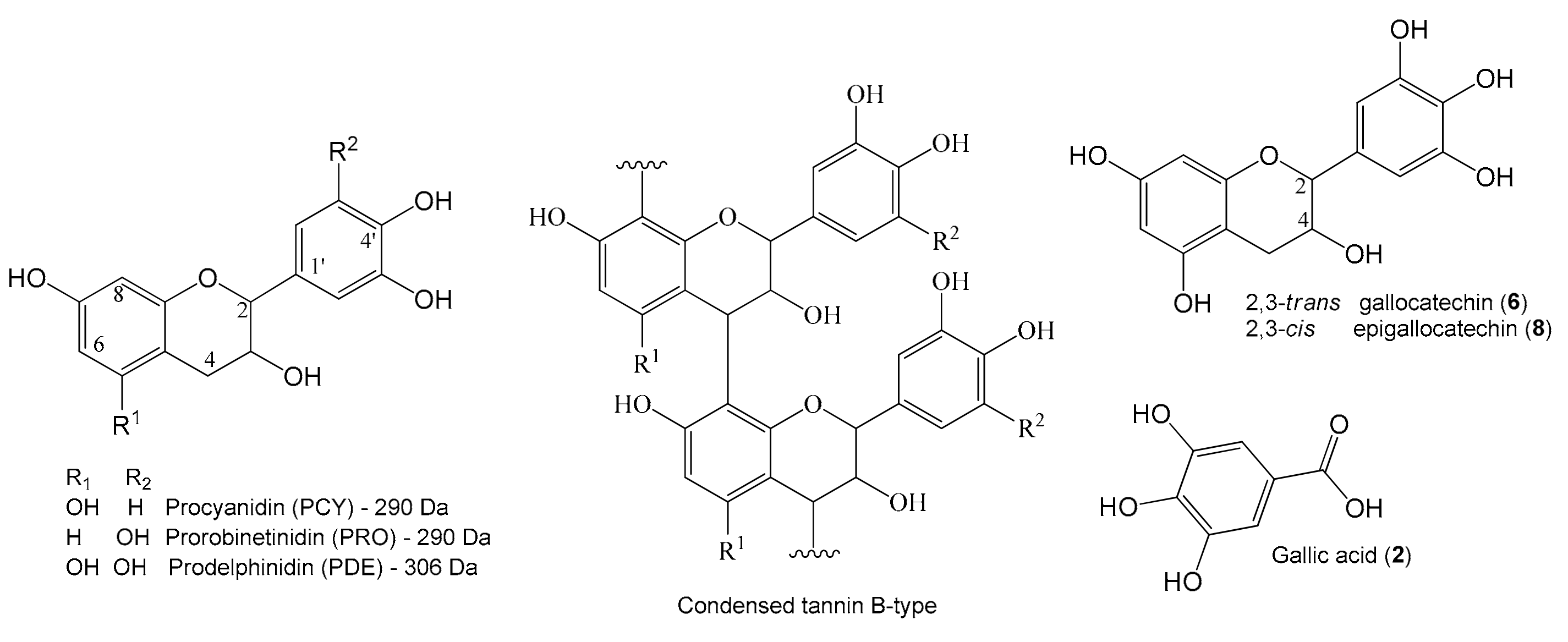
2.1. Identification of the Constituents from the SAAE by LC-DAD-MS/MS
2.2. Chemical Composition
2.3. Antioxidant Activity
2.3.1. ABTS and DPPH Free-Radical Scavenging
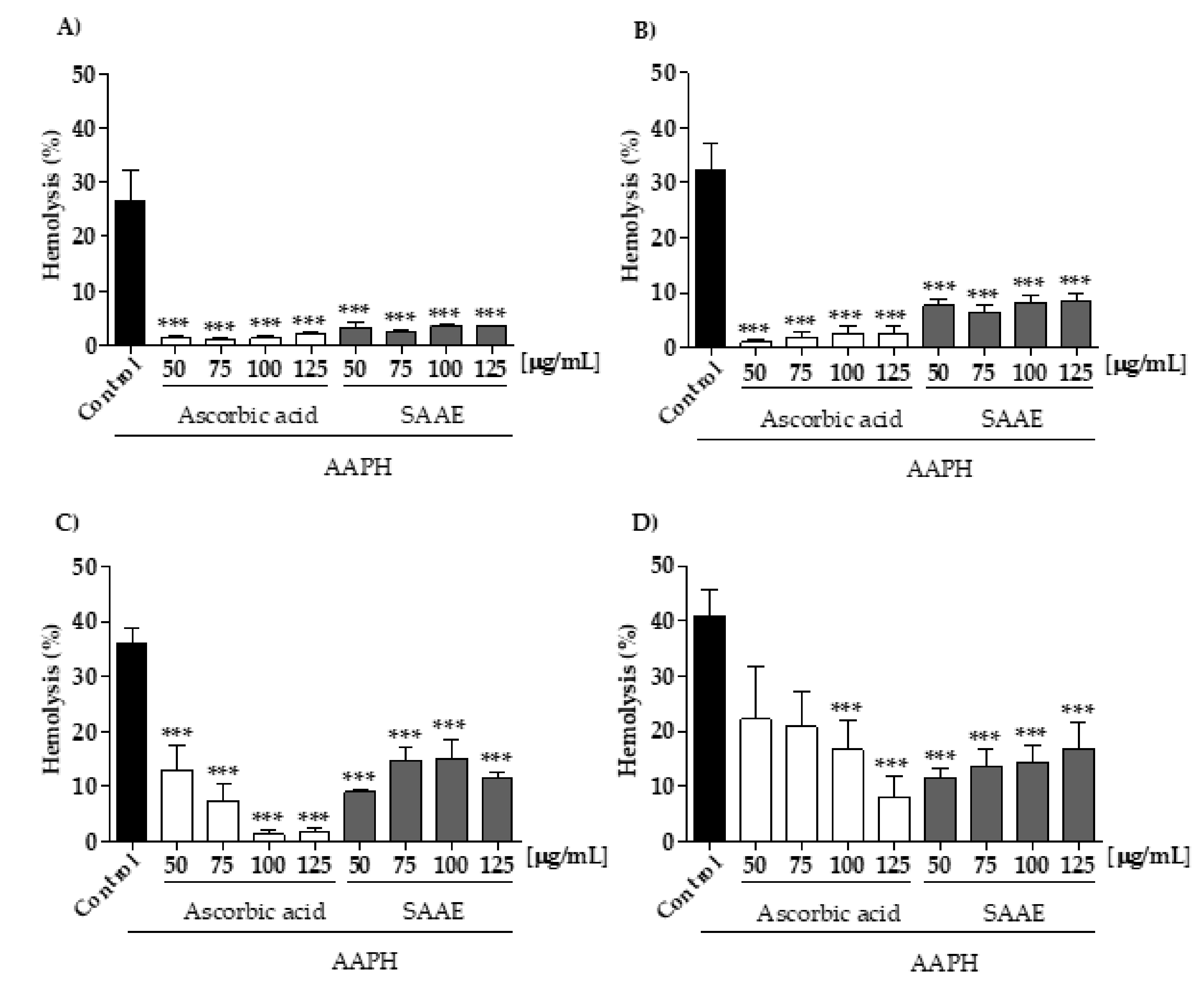
2.3.2. Protective Effect of the SAAE Against Oxidative Hemolysis
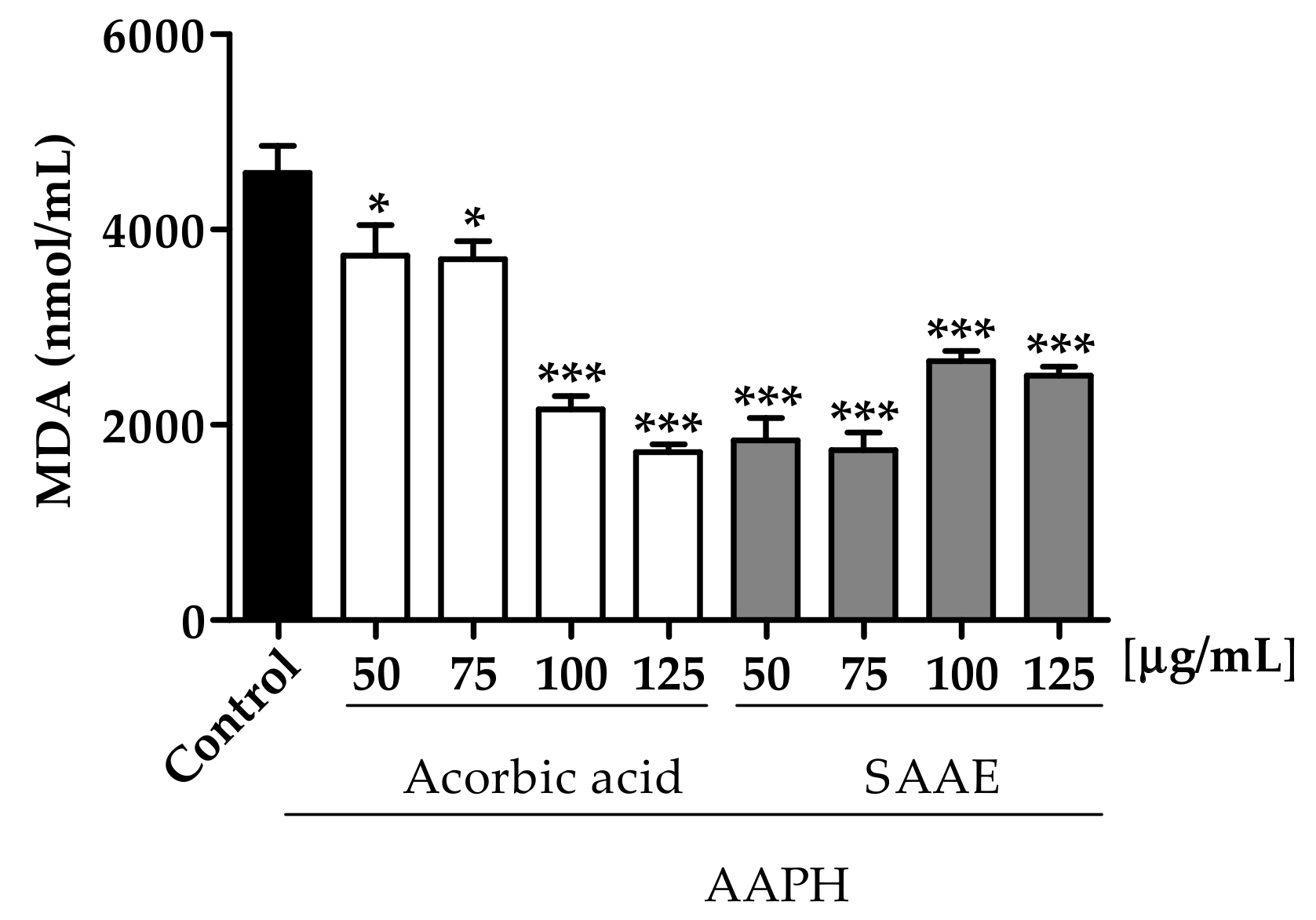
2.3.3. Malondialdehyde (MDA) Dosage
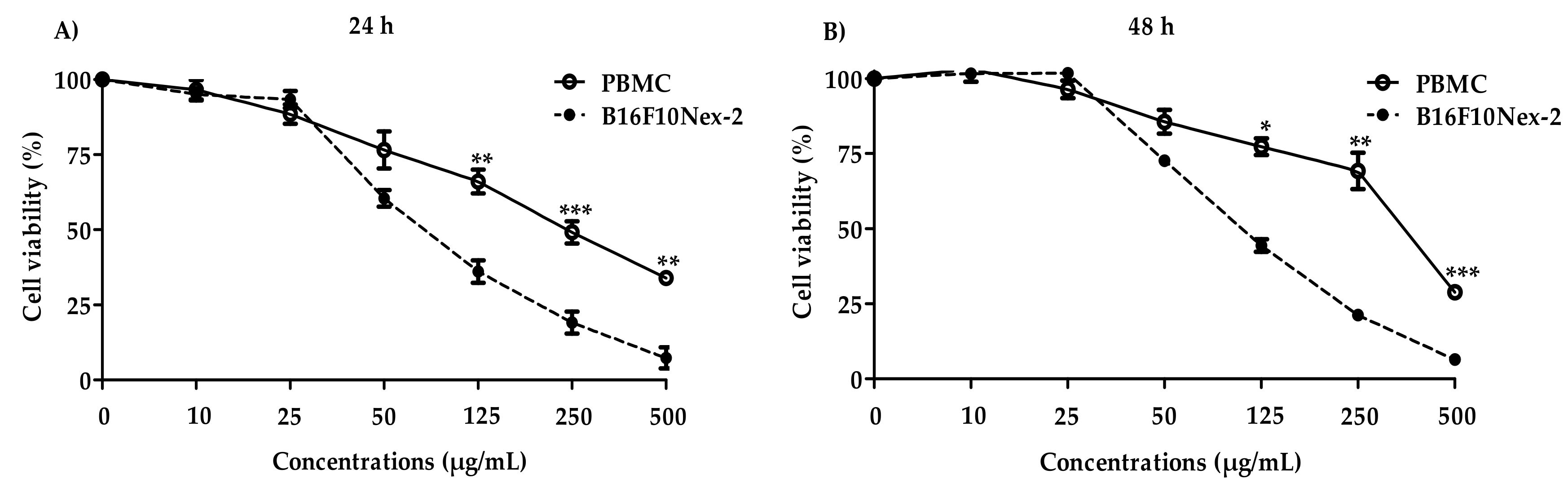
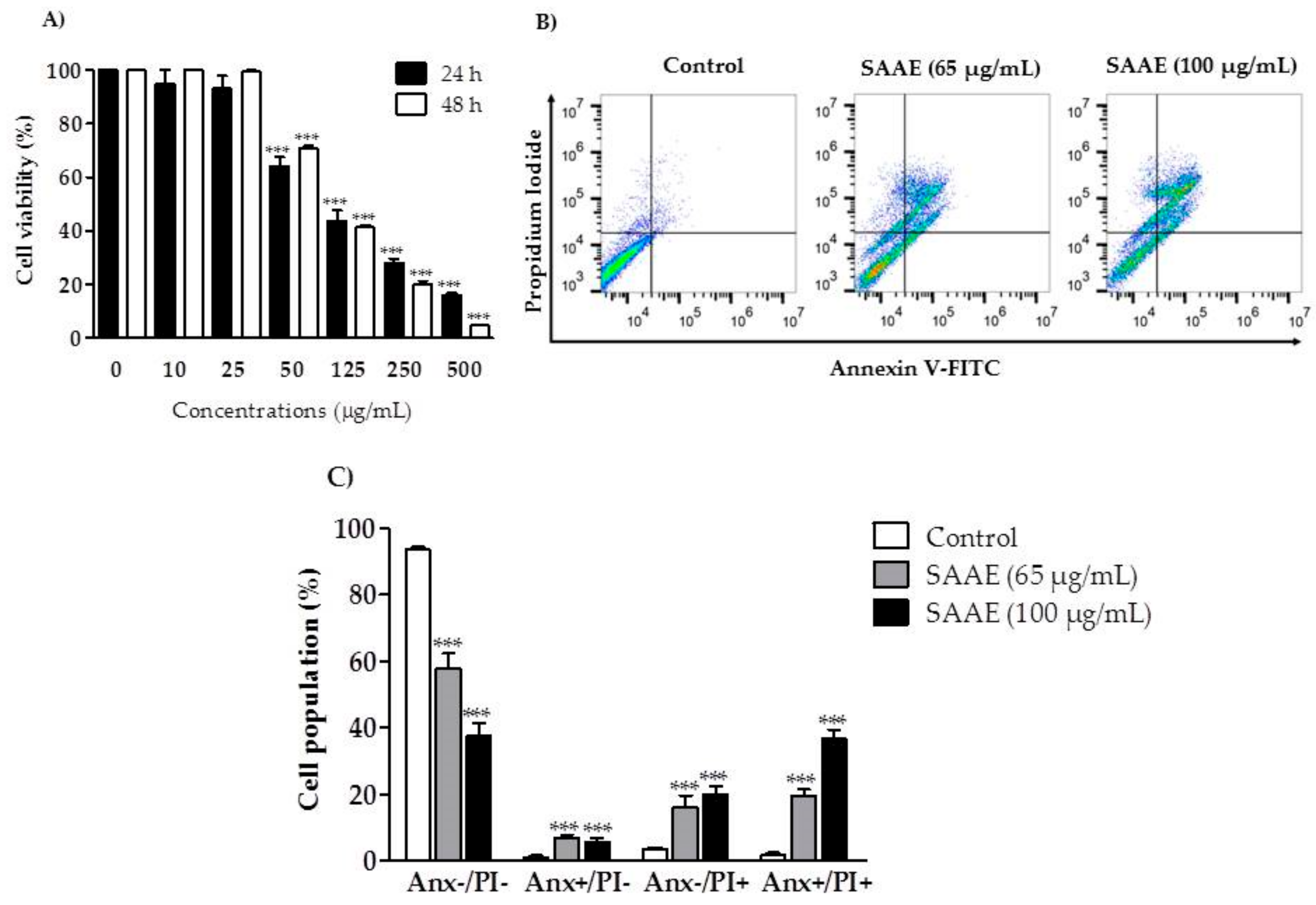
2.4. Cytotoxic Activity and Cell Death Profile
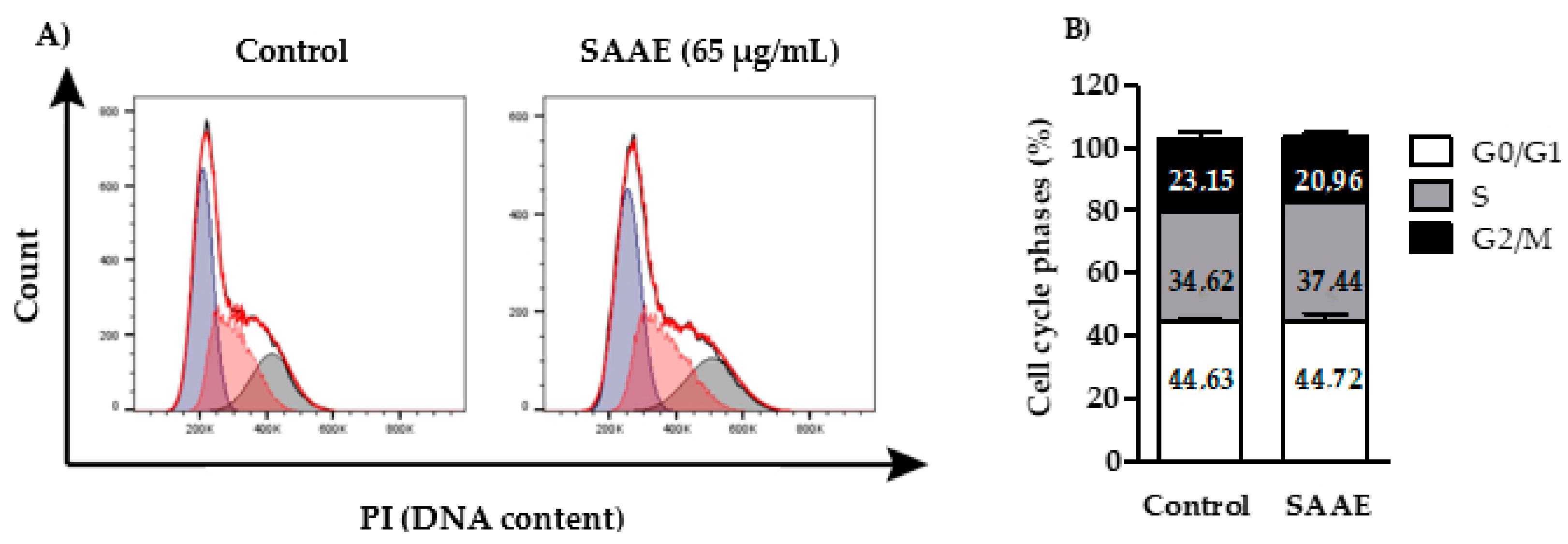
2.4.1. Cell Cycle Phases
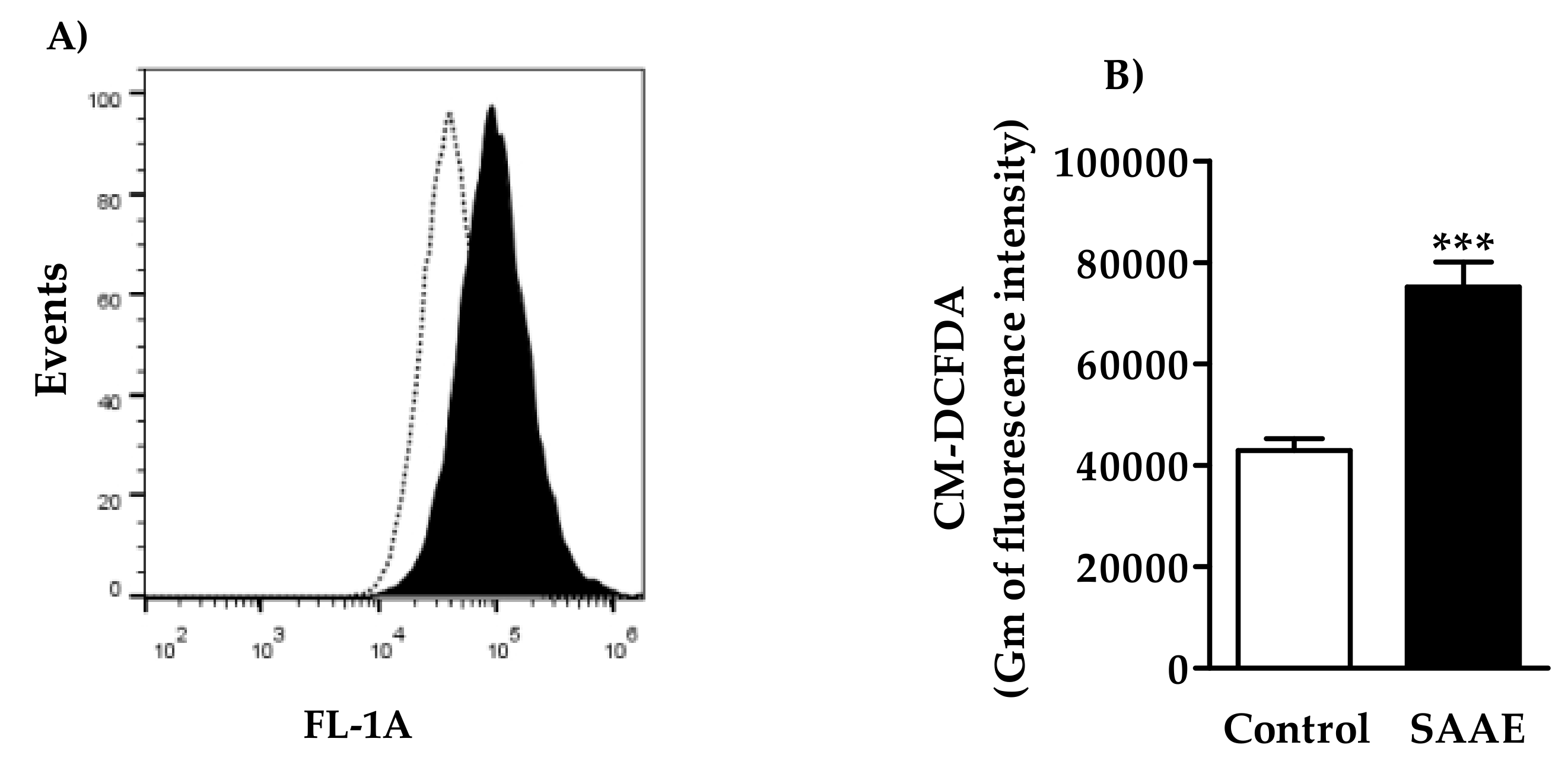
2.4.2. Reactive Oxygen Species (ROS) Levels
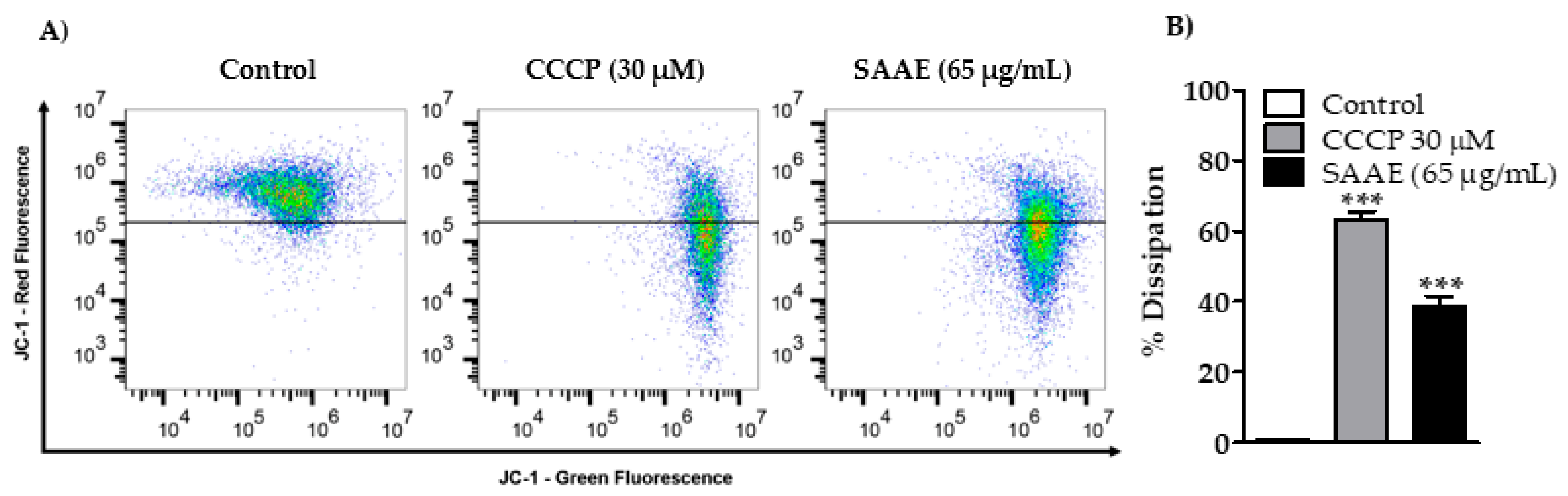
2.4.3. Mitochondrial Membrane Potential
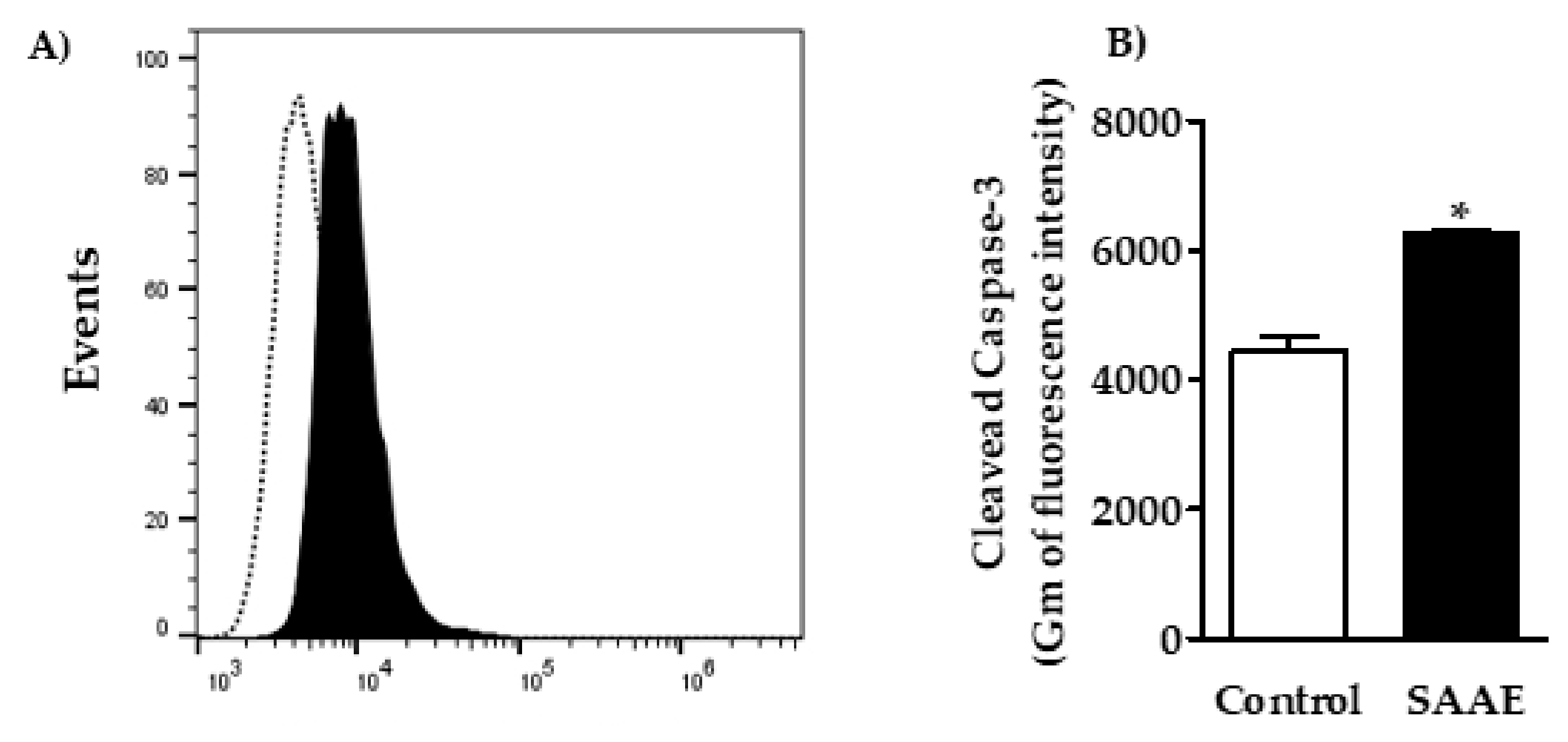
2.4.4. Caspase-3 Activation
3. Discussion
4. Materials and Methods
4.1. Plant Material Collection
4.2. Aqueous Extract Preparation
4.3. Chemical Analysis
4.3.1. Identification of Constituents by LC-DAD-MS
4.3.2. Determination of Phenolic Compounds and Total Flavonoids
Phenolic Compounds
Total Flavonoids
4.4. Antioxidant Activity
4.4.1. ABTS•+ Radical Discoloration Assay
4.4.2. DPPH Free Radical Capture Activity
4.4.3. Antioxidant Assay in Human Erythrocytes
Erythrocyte Suspension Preparation
Hemolytic Activity and Oxidative Hemolysis Inhibition
Malondialdehyde (MDA) Dosage
4.5. Cell Cultures
4.5.1. MTT Cell Viability Assay
4.5.2. Cell Death Profile
4.5.3. Cell Cycle Phases
4.5.4. Assessment of Reactive Oxygen Species (ROS) Levels
4.5.5. Assessment of the Mitochondrial Membrane Potential
4.5.6. Caspase-3 Activity
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AAPH | 2,2’-Azobis (2-methylpropionamidine) dihydrochloride |
| ABTS | 2,2’-Azino-bis(3-ethylbenzothiazoline-6-sulphonic acid) |
| CCCP | Carbonilcianeto-m-clorofenilhidrazona |
| CM-H2DCFDA | 2′,7′-dichlorodihydrofluorescein diacetate |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl |
| FBS | Fetal bovine serum |
| GAE | Gallic acid equivalente |
| HEPES | 4-(2-Hydroxyethyl) piperazine-1-ethanesulfonic acid |
| LICR | Ludwig Institute for Cancer Research |
| MDA | Malondialdehyde |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide |
| NaCl | Sodium chloride |
| PBMC | Human peripheral blood mononuclear cells |
| PI | Propidium iodide |
| QE | Quercetin equivalents |
| ROS | Reactive oxygen species |
| SAAE | Stryphnodendron adstringens aqueous extract |
| SEM | Standard error of the mean |
| TBA | Thiobarbituric acid |
References
- Santana, B.F.; Voeks, R.A.; Funch, L.S. Ethnomedicinal survey of a maroon community in Brazil’s Atlantic tropical forest. J. Ethnopharmacol. 2016, 181, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, U.P.; Monteiro, J.M.; Ramos, M.A.; de Amorim, E.L.C. Medicinal and magic plants from a public market in northeastern Brazil. J. Ethnopharmacol. 2007, 110, 76–91. [Google Scholar] [CrossRef] [PubMed]
- Hernandes, L.; Pereira, L.M.D.S.; Palazzo, F.; Mello, J.C.P.D. Wound-healing evaluation of ointment from Stryphnodendron adstringens (barbatimão) in rat skin. Braz. J. Pharm. Sci. 2010, 46, 431–436. [Google Scholar] [CrossRef]
- Lima, J.C.S.; Martins, D.T.O.; de Souza, P.T. Experimental evaluation of stem bark of Stryphnodendron adstringens (Mart.) Coville for antiinflammatory activity. Phytother Res. 1998, 12, 218–220. [Google Scholar] [CrossRef]
- Henriques, B.O.; Corrêa, O.; Azevedo, E.P.C.; Pádua, R.M.; de Oliveira, V.L.; Oliveira, T.H.C.; Boff, D.; Dias, A.C.F.; de Souza, D.G.; Amaral, F.A.; et al. In vitro TNF-inhibitory activity of brazilian plants and anti-inflammatory effect of Stryphnodendron adstringens in an acute arthritis model. Evid.-Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Morey, A.T.; Souza, F.C.; Santos, J.P.; Pereira, C.A.; Cardoso, J.D.; de Almeida, R.S.; Costa, M.A.; Mello, J.C.; Nakamura, C.V.; Pinge-Filho, P.; et al. Antifungal activity of condensed tannins from Stryphnodendron adstringens: Effect on candida tropicalis growth and adhesion properties. Curr. Pharm. Biotechnol. 2016, 17, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Luiz, R.L.; Vila, T.V.; Mello, J.C.; Nakamura, C.V.; Rozental, S.; Ishida, K. Proanthocyanidins polymeric tannin from Stryphnodendron adstringens are active against Candida albicans biofilms. BMC Complement. Altern. Med. 2015, 15, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mello, J.P.; Petereit, F.; Nahrstedt, A. Prorobinetinidins from Stryphnodendron adstringens. Phytochemistry 1996, 42, 857–862. [Google Scholar] [CrossRef]
- Mello, J.C.P.; Petereit, F.; Nahrstedt, A.A. Dimeric proanthocyanidin from Stryphnodendron adstringens. Phytochemistry 1999, 51, 1105–1107. [Google Scholar]
- Padumadasa, C.; Dharmadana, D.; Abeysekera, A.; Thammitiyagodage, M. In vitro antioxidant, anti-inflammatory and anticancer activities of ethyl acetate soluble proanthocyanidins of the inflorescence of Cocos nucifera L. BMC Complement. Altern. Med. 2016, 16, 345. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.H.; Liu, C.; Fan, R.; Zhu, L.F.; Yang, S.X.; Zhu, H.T.; Wang, D.; Yang, C.R.; Zhang, Y.J. Antioxidative Flavan-3-ol Dimers from the Leaves of Camellia fangchengensis. J. Agric. Food Chem. 2018, 66, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Casagrande, J.C.; Macorini, L.F.B.; Antunes, K.A.; dos Santos, U.P.; Campos, J.F.; Dias-Júnior, N.M.; Sangalli, A.; Cardoso, C.A.L.; Vieira, M.D.C.; Rabelo, L.A.; et al. Antioxidant and cytotoxic activity of hydroethanolic extract from Jacaranda decurrens leaves. PLoS ONE 2014, 9, e112748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos, J.F.; Castro, D.T.H.; Damião, M.J.; Torquato, H.F.V.; Paredes-Gamero, E.J.; Carollo, C.A.; Estevinho, L.M.; Souza, K.P.; Santos, E.L. The chemical profile of Senna velutina leaves and their antioxidant and cytotoxic effects. Oxid. Med. Cell Longev. 2016, 2016, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Espindola, P.T.; Rocha, P.S.; Carollo, C.A.; Schmitz, W.O.; Pereira, Z.V.; Vieira, M.C.; Santos, E.L.; Souza, K.P. Antioxidant and antihyperlipidemic effects of Campomanesia adamantium O. Berg root. Oxid. Med. Cell Longev. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.H.; Macorini, L.F.; Antunes, K.Á.; Espindola, P.T.; Alfredo, T.M.; da Rocha, P.S.; Pereira, Z.V.; Santos, E.L.; Souza, K.P. Antioxidant and hypolipidemic activity of the hydroethanolic extract of Curatella americana l. leaves. Oxid. Med. Cell Longev. 2016, 2016, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Santos, U.P.; Campos, J.F.; Torquato, H.F.; Paredes-Gamero, E.J.; Carollo, C.A.; Estevinho, L.M.; Souza, K.P.; Santos, E.L. Antioxidant, antimicrobial and cytotoxic properties as well as the phenolic content of the extract from Hancornia speciosa Gomes. PLoS ONE 2016, 11, e0167531. [Google Scholar] [CrossRef] [PubMed]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release. 2006, 113, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Acharya, A.; Das, I.; Chandhok, D.; Saha, T. Redox regulation in cancer: A double-edged sword with therapeutic potential. Oxid. Med. Cell Longev. 2010, 3, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Dror, S.; Sander, L.; Schwartz, H.; Sheinboim, D.; Barzilai, A.; Dishon, Y.; Apcher, S.; Golan, T.; Greenberger, S.; Barshack, I.; et al. Melanoma miRNA trafficking controls tumour primary niche formation. Nat. Cell Biol. 2016, 18, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Gorantla, V.C.; Kirkwood, J.M. State of melanoma: An historic overview of a field in transition. Hematol. Oncol. Clin. North Am. 2014, 28, 415–435. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Ultraviolet Radiation and the Intersun Programme. Skin Cancers. Available online: http://www.who.int/uv/faq/skincancer/en/index1.html (accessed on 15 July 2016).
- American Cancer Society (ACS). Melanoma Skin Cancer Detailed Guide. Available online: https://www.cancer.org/cancer/melanoma-skin-cancer/detection-diagnosis-staging/survival-rates-for-melanoma-skin-cancer-by-stage.html (accessed on 31 July 2017).
- Marelli, M.M.; Marzagalli, M.; Moretti, R.M.; Beretta, G.; Casati, L.; Comitato, R.; Gravina, G.L.; Festuccia, C.; Limonta, P. Vitamin E δ-tocotrienol triggers endoplasmic reticulum stress-mediated apoptosis in human melanoma cells. Sci Rep. 2016, 6, 30502. [Google Scholar] [CrossRef] [PubMed]
- Mattia, G.; Puglisi, R.; Ascione, B.; Malorni, W.; Carè, A.; Matarrese, P. Cell death-based treatments of melanoma:conventional treatments and new therapeutic strategies. Cell Death. Dis. 2018, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nocchi, S.R.; Companhoni, M.V.; de Mello, J.C.; Dias Filho, B.P.; Nakamura, C.V.; Carollo, C.A.; Silva, D.B.; Ueda-Nakamura, T. Antiviral Activity of Crude Hydroethanolic Extract from Schinus terebinthifolia against Herpes simplex Virus Type 1. Planta Med. 2017, 83, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Isler, A.C.; Lopes, G.C.; Cardoso, M.L.C.; Mello, J.C.P. Development and validation of a LC-method for the determination of phenols in a pharmaceutical formulation containing extracts from Stryphnodendron adstringens. Quím. Nova 2010, 33, 1126–1129. [Google Scholar] [CrossRef]
- Robeson, D.J.; Ingham, J.L.; Harborne, J.B. Identification of two chromone phytoalexins in the sweet pea, Lathyrus odoratus. Phtochemistry 1980, 19, 2171–2173. [Google Scholar] [CrossRef]
- Yang, S.J.; Ryu, J.H.; Jang, D.S.; Yang, L.; Han, H.K. A sensitive LC–MS/MS method for the quantitative determination of biflorin in rat plasma and its application to pharmacokinetic studies. J. Pharm. Biomed. Anal. 2015, 115, 272–276. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Li, X.; Shi, Q.; Feng, F. An effective integrated method for comprehensive identification of eighty-five compounds in Zhi-Zi-Da-Huang decoction by HPLC-DAD-ESI-MS (TOF) and HPLC-DAD-ESI-MS/MS (QqQ) without the help of reference standards. Anal. Methods 2014, 6, 4312–4327. [Google Scholar] [CrossRef]
- Pinto, S.C.; Bueno, F.G.; Panizzon, G.P.; Morais, G.; dos Santos, P.V.; Baesso, M.L.; Leite-Mello, E.V.; de Mello, J.C. Stryphnodendron adstringens: Clarifying wound healing in streptozotocin-induced diabetic rats. Planta Med. 2015, 81, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Z.; Meng, R.; Shi, C.; Guo, N. Antioxidative and anticancer properties of Licochalcone A from licorice. J. Ethnopharmacol. 2017, 198, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Allouche, Y.; Warleta, F.; Campos, M.; Sánchez-Quesada, C.; Uceda, M.; Beltrán, G.; Gaforio, J.J. Antioxidant, antiproliferative, and pro-apoptotic capacities of pentacyclic triterpenes found in the skin of olives on MCF-7 human breast cancer cells and their effects on DNA damage. J. Agric. Food Chem. 2010, 59, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Orlikova, B.; Tasdemir, D.; Golais, F.; Dicato, M.; Diederich, M. Dietary chalcones with chemopreventive and chemotherapeutic potential. Genes Nutr. 2011, 6, 125–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendoza-Wilson, A.M.; Castro-Arredondo, S.I.; Espinosa-Plascencia, A.; Robles-Burgueño, M.R.; Balandrán-Quintana, R.R.; Bermúdez-Almada, M.C. Chemical composition and antioxidant-prooxidant potential of a polyphenolic extract and a proanthocyanidin-rich fraction of apple skin. Heliyon 2016, 2, e00073. [Google Scholar] [CrossRef] [PubMed]
- Amoussa, A.M.; Bourjot, M.; Lagnika, L.; Vonthron-Sénécheau, C.; Sanni, A. Acthaside: A new chromone derivative from Acacia ataxacantha and its biological activities. BMC Complement. Altern. Med. 2016, 16, 506. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.F.D.; Campos, J.F.; Santos, C.M.D.; Balestieri, J.B.P.; Silva, D.B.; Carollo, C.A.; Souza, K.P.; Estevinho, L.M.; Santos, E.L. Chemical profile and antioxidant, anti-inflammatory, antimutagenic and antimicrobial activities of geopropolis from the stingless bee Melipona orbignyi. Int. J. Mol. Sci. 2017, 18, 953. [Google Scholar] [CrossRef] [PubMed]
- Kähkönen, M.P.; Hopia, A.I.; Vuorela, H.J.; Rauha, J.P.; Pihlaja, K.; Kujala, T.S.; Heinonen, M. Antioxidant activity of plant extracts containing phenolic compounds. J. Agric. Food Chem. 1999, 47, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef] [PubMed]
- Macáková, K.; Kolečkář, V.; Cahlíková, L.; Chlebek, J.; Hošťálková, A.; Kuča, K.; Jun, D.; Opletal, L. Chaper 6: Tannins and their Influence on Health. In Recent Advances in Medicinal Chemistry, 1st ed.; Choudhary, M.I., Perry, G., Rahman, A., Eds.; Elsevier, Bentham Science Publishers: San Antonio, TX, USA, 2014; Volume 1, pp. 159–208. [Google Scholar]
- López-Alarcón, C.; Denicola, A. Evaluating the antioxidant capacity of natural products: A review on chemical and cellular-based assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Suwalsky, M.; Colina, J.; Gallardo, M.J.; Jemiola-Rzeminska, M.; Strzalka, K.; Manrique-Moreno, M.; Sepúlveda, B. Antioxidant capacity of gallic acid in vitro assayed on human erythrocytes. J. Membr. Biol. 2016, 249, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Liu, J.; Guan, R.; Chen, J.; Yang, D.; Zhao, Z.; Wang, D. Chemical characterization of procyanidins from Spatholobus suberectus and their antioxidative and anticancer activities. J. Funct. Foods. 2015, 12, 468–477. [Google Scholar] [CrossRef]
- Zhu, Q.Y.; Schramm, D.D.; Gross, H.B.; Holt, R.R.; Kim, S.H.; Yamaguchi, T.; Kwik-Uribe, C.L.; Keen, C.L. Influence of cocoa flavanols and procyanidins on free-radical-induced human erythrocyte hemolysis. Clin. Dev. Immunol. 2005, 12, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Arwa, P.S.; Zeraik, M.L.; Ximenes, V.F.; da Fonseca, L.M.; Bolzani, V.S.; Siqueira Silva, D.H.S. Redox-active biflavonoids from Garcinia brasiliensis as inhibitors of neutrophil oxidative burst and human erythrocyte membrane damage. J. Ethnopharmacol. 2015, 174, 410–418. [Google Scholar] [CrossRef] [PubMed]
- Majchrzak, D.; Mitter, S.; Elmadfa, I. The effect of ascorbic acid on total antioxidant activity of black and green teas. Food Chem. 2004, 88, 447–451. [Google Scholar] [CrossRef]
- Macedo, J.A.; Battestin, V.; Ribeiro, M.L.; Macedo, G.A. Increasing the antioxidant power of tea extracts by biotransformation of polyphenols. Food Chem. 2011, 126, 491–497. [Google Scholar] [CrossRef]
- Chen, L.; Yang, X.; Jiao, H.; Zhao, B. Tea catechins protect against lead-induced cytotoxicity, lipid peroxidation, and membrane fluidity in HepG2 cells. Toxicol. Sci. 2002, 69, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.F.; Santos, U.P.; Macorini, L.F.B.; Melo, A.M.M.F.; Balestieri, J.B.P.; Paredes-Gamero, E.J.; Cardoso, C.A.L.; Souza, K.P.; Santos, E.L.D. Antimicrobial, antioxidant and cytotoxic activities of propolis from Melipona orbignyi (Hymenoptera, Apidae). Food Chem. Toxicol. 2014, 65, 374–380. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Fang, F.; Zhang, X.; Liu, B.; Luo, H.; Li, Z.; Zhang, X.; Zhang, Z.; Pang, X. B Type and complex A/B Type epicatechin trimers isolated from Litchi pericarp aqueous extract show high antioxidant and anticancer activity. Int. J. Mol. Sci. 2018, 19, 301. [Google Scholar] [CrossRef] [PubMed]
- Ravi Shankara, B.E.; Ramachandra, Y.L.; Rajan, S.S.; Ganapathy, P.S.; Yarla, N.S.; Richard, S.A.; Dhananjaya, B.L. Evaluating the anticancer potential of ethanolic gall extract of Terminalia chebula (Gaertn.) Retz. (Combretaceae). Pharmacognosy Res. 2016, 8, 209. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V.; Mbaveng, A.T.; Nono, E.C.; Simo, C.C.; Zeino, M.; Nkengfack, A.E.; Efferth, T. Cytotoxicity of seven naturally occurring phenolic compounds towards multi-factorial drug-resistant cancer cells. Phytomedicine 2016, 23, 856–863. [Google Scholar] [CrossRef] [PubMed]
- Brys, A.K.; Gowda, R.; Loriaux, D.B.; Robertson, G.P.; Mosca, P.J. Nanotechnology-based strategies for combating toxicity and resistance in melanoma therapy. Biotechnol. Adv. 2016, 34, 565–577. [Google Scholar] [CrossRef] [PubMed]
- Soengas, M.S.; Lowe, S.W. Apoptosis and melanoma chemoresistance. Oncogene 2003, 22, 3138–3151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Looi, C.Y.; Moharram, B.; Paydar, M.; Wong, Y.L.; Leong, K.H.; Mohamad, K.; Arya, A.; Wong, W.F.; Mustafa, M.R. Induction of apoptosis in melanoma A375 cells by a chloroform fraction of Centratherum anthelminticum (L.) seeds involves NF-kappaB, p53 and Bcl-2-controlled mitochondrial signaling pathways. BMC Complement. Altern. Med. 2013, 13, 166. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Cheng, Y.; Zhang, B.; Bian, H.J.; Bao, J.K. Polygonatum cyrtonema lectin induces apoptosis and autophagy in human melanoma A375 cells through a mitochondria-mediated ROS–p38–p53 pathway. Cancer Lett. 2009, 275, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Konorev, E.A.; Kotamraju, S.; Joseph, J.; Kalivendi, S.; Kalyanaraman, B. Doxorubicin induces apoptosis in normal and tumor cells via distinctly different mechanisms intermediacy of H2O2-and p53-dependent pathways. J. Biol. Chem. 2004, 279, 25535–25543. [Google Scholar] [CrossRef] [PubMed]
- Florea, A.M.; Büsselberg, D. Cisplatin as an anti-tumor drug: Cellular mechanisms of activity, drug resistance and induced side effects. Cancers 2011, 3, 1351–1371. [Google Scholar] [CrossRef] [PubMed]
- Selimovic, D.; Badura, H.E.; El-Khattouti, A.; Soell, M.; Porzig, B.B.; Spernger, A.; Ghanjati, F.; Santourlidis, S.; Haikel, Y.; Hassan, M. Vinblastine-induced apoptosis of melanoma cells is mediated by Ras homologous A protein (Rho A) via mitochondrial and non-mitochondrial-dependent mechanisms. Apoptosis 2013, 18, 980–997. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H.; Lee, S.H.; Meng, X.W.; Loegering, D.A.; Kottke, T.J.; Henzing, A.J.; Earnshaw, W.C.; Ruchaud, S.; Samejima, K.; Earnshaw, W.C. Apoptosis-associated caspase activation assays. Methods 2008, 44, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Bortner, C.D.; Cidlowski, J.A. Apoptotic volume decrease and the incredible shrinking cell. Cell Death. Differ. 2002, 9, 1307–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, B.; Dutta, D.; Mukherjee, S.; Das, S.; Maiti, N.C.; Das, P.; Chowdhury, C. Synthesis and biological evaluation of a novel betulinic acid derivative as an inducer of apoptosis in human colon carcinoma cells (HT-29). Eur. J. Med. Chem. 2015, 102, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Renschler, M.F. The emerging role of reactive oxygen species in cancer therapy. Eur. J. Cancer 2004, 40, 1934–1940. [Google Scholar] [CrossRef] [PubMed]
- Anantharaman, A.; Hemachandran, H.; Mohan, S.; Ayyathan, D.M.; Kumar, D.T.; Doss, G.P.C.; Siva, R. Induction of apoptosis by apocarotenoids in B16 melanoma cells through ROS-mediated mitochondrial-dependent pathway. J. Funct. Foods 2016, 20, 346–357. [Google Scholar] [CrossRef]
- Shi, X.J.; Yu, B.; Wang, J.W.; Qi, P.P.; Tang, K.; Huang, X.; Liu, H.M. Structurally novel steroidal spirooxindole by241 potently inhibits tumor growth mainly through ROS-mediated mechanisms. Sci Rep. 2016, 6, 31607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Lange, T.S.; Kim, K.K.; Brard, L.; Horan, T.; Moore, R.G.; Vorsa, N.; Singh, R.K. Purified cranberry proanthocyanidines (PAC-1A) cause pro-apoptotic signaling, ROS generation, cyclophosphamide retention and cytotoxicity in high-risk neuroblastoma cells. Int. J. Oncol. 2012, 40, 99–108. [Google Scholar] [PubMed]
- Zhang, Y.; Luo, M.; Zu, Y.; Fu, Y.; Gu, C.; Wang, W.; Yao, L.; Efferth, T. Dryofragin, a phloroglucinol derivative, induces apoptosis in human breast cancer MCF-7 cells through ROS-mediated mitochondrial pathway. Chem. Biol. Interact. 2012, 199, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.L.; Park, S.Y.; Kim, Y.H.; Park, G.; Lee, S.J. Halofuginone induces the apoptosis of breast cancer cells and inhibits migration via downregulation of matrix metalloproteinase-9. Int. J. Oncol. 2014, 44, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.K.; Supriady, H.; Goh, B.H.; Kadir, H.A. Elephantopus scaber induces apoptosis through ROS-dependent mitochondrial signaling pathway in HCT116 human colorectal carcinoma cells. J. Ethnopharmacol. 2015, 168, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Akhtar, J.; Sahabjada; Arshad, M. Evaluation of cytotoxic potential and apoptotic effect of a methanolic extract of Bauhinia racemosa Lam. against a human cancer cell line, HeLa. Eur. J. Integr. Med. 2016, 8, 513–518. [Google Scholar] [CrossRef]
- Qian, Y.; Guan, T.; Huang, M.; Cao, L.; Li, Y.; Cheng, H.; Jin, H.; Yu, D. Neuroprotection by the soy isoflavone, genistein, via inhibition of mitochondria-dependent apoptosis pathways and reactive oxygen induced-NF-κB activation in a cerebral ischemia mouse model. Neurochem. Int. 2012, 60, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Kuranaga, E. Beyond apoptosis: Caspase regulatory mechanisms and functions in vivo. Genes Cells 2012, 17, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.J.; Lee, J.H.; Moon, K.D.; Jeong, I.Y.; Ahn, D.U.; Lee, M.K.; Seo, K.I. Induction of apoptosis by isoegomaketone from Perilla frutescens L. in B16 melanoma cells is mediated through ROS generation and mitochondrial-dependent,-independent pathway. Food Chem. Toxicol. 2014, 65, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Ichim, G.; Tait, S.W. A fate worse than death: Apoptosis as an oncogenic process. Nat. Rev. Cancer. 2016, 18, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Liu, S.; Liu, Y.; Wang, F.; Ren, J.; Gu, J.; Zhou, K.; Shan, B. A novel cyclic pentapeptide, H-10, inhibits B16 cancer cell growth and induces cell apoptosis. Oncol. Lett. 2014, 8, 248–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, S.W.G.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Zhu, X.; Zhang, K.; Zhu, L.; Zhou, F. Investigation of gallic acid induced anticancer effect in human breast carcinoma MCF-7 cells. J. Biochem. Mol. Toxicol. 2014, 28, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Ma, L.; Weng, D.; Yao, J.; Liu, X.; Jin, F. Gallic acid induces apoptosis and enhances the anticancer effects of cisplatin in human small cell lung cancer H446 cell line via the ROS-dependent mitochondrial apoptotic pathway. Oncol. Rep. 2016, 35, 3075–3083. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.M.S.; Rocha, R.G.; Santos, H.O.; Guimarães, T.A.; Fraga, C.A.C.; Silveira, L.H.; Batista, P.R.; Oliveira, P.S.L.; Melo, G.A.; Santos, S.H.; et al. Gallic acid modulates phenotypic behavior and gene expression in oral squamous cell carcinoma cells by interfering with leptin pathway. Pathol. Res. Pract. 2018, 214, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Zhang, S.; Xie, Y.; Zhang, Z.; Zhao, W. Gallic acid as a selective anticancer agent that induces apoptosis in SMMC-7721 human hepatocellular carcinoma cells. Oncol. Lett. 2016, 11, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lin, J.J.; Yang, Z.Y.; Tsai, C.C.; Hsu, J.L.; Wu, Y.J. Proteomic study reveals a co-occurrence of gallic acid-induced apoptosis and glycolysis in B16F10 melanoma cells. J. Agric. Food Chem. 2014, 62, 11672–11680. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.; Lai, T.Y.; Yang, J.H.; Yang, J.S.; Ma, Y.S.; Weng, S.W.; Chen, Y.Y.; Lin, J.G.; Chung, J.G. Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int. J. Oncol. 2010, 37, 377–385. [Google Scholar] [PubMed]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. Rsc. Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Fu, Y.; Ye, X.; Lee, M.; Rankin, G.; Chen, Y.C. Prodelphinidins isolated from Chinese bayberry leaves induces apoptosis via the p53-dependent signaling pathways in OVCAR-3 human ovarian cancer cells. Oncol Lett. 2017, 13, 3210–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, W.; Toda, K.; Kawaguchi, K.; Kawahara, S.I.; Katoh, M.; Hattori, Y.; Fujii, H.; Makabe, H. Syntheses of prodelphinidin B3 and C2, and their antitumor activities through cell cycle arrest and caspase-3 activation. Tetrahedron 2013, 69, 3543–3550. [Google Scholar] [CrossRef] [Green Version]
- Engelbrecht, A.M.; Mattheyse, M.; Ellis, B.; Loos, B.; Thomas, M.; Smith, R.; Peters, S.; Smith, C.; Myburgh, K. Proanthocyanidin from grape seeds inactivates the PI3-kinase/PKB pathway and induces apoptosis in a colon cancer cell line. Cancer Lett. 2007, 258, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Deep, G.; Wempe, M.F.; Surek, J.; Kumar, A.; Agarwal, R.; Agarwal, C. Procyanidin B2 3,3″-di-O-gallate induces oxidative stress-mediated cell death in prostate cancer cells via inhibiting MAP kinase phosphatase activity and activating ERK1/2 and AMPK. Mol. Carcinog. 2017, 57, 57–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.P.; Lin, Y.H.; Chou, C.C.; Zhou, S.P.; Hsu, Y.C.; Liu, C.L.; Ku, F.M.; Chung, Y.C. Mechanisms of grape seed procyanidin-induced apoptosis in colorectal carcinoma cells. Anticancer Res. 2009, 29, 283–289. [Google Scholar] [PubMed]
- Reis, J.; Gaspar, A.; Milhazes, N.; Borges, F. Chromone as a Privileged Scaffold in Drug Discovery: Recent Advances. J. Med. Chem. 2017, 60, 7941–7957. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Kumar, S.; Chand, K.; Kathuria, A.; Gupta, A.; Jain, R. An Update on Natural Occurrence and Biological Activity of Chromones. Curr. Med. Chem. 2011, 18, 3825–3852. [Google Scholar] [CrossRef] [PubMed]
- Keri, R.S.; Budagumpi, S.; Pai, R.K.; Balakrishna, R.G. Chromones as a privileged scaffold in drug discovery: A review. Eur. J. Med. Chem. 2014, 6, 340–374. [Google Scholar] [CrossRef] [PubMed]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Liberio, S.A.; Pereira, A.L.; Dutra, R.P.; Reis, A.S.; Araújo, M.J.; Mattar, N.S.; Silva, L.A.; Ribeiro, M.N.; Nascimento, F.R.; Guerra, R.N.; Monteiro-Neto, V. Antimicrobial activity against oral pathogens and immunomodulatory effects and toxicity of geopropolis produced by the stingless bee Melipona fasciculata Smith. BMC Complement. Altern. Med. 2011, 11, 108. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Gupta, D.; Gupta, R.K. Bioprotective properties of Dragon’s blood resin: In vitro evaluation of antioxidant activity and antimicrobial activity. BMC Complement. Altern. Med. 2011, 13. [Google Scholar] [CrossRef] [PubMed]
- Campos, J.F.; dos Santos, U.P.; Rocha, P.S.; Damião, M.J.; Balestieri, J.B.P.; Cardoso, C.A.L.; Paredes-Gamero, E.J.; Estevinho, L.M.; Souza, K.P.; Santos, E.L. Antimicrobial, antioxidant, anti-inflammatory, and cytotoxic activities of propolis from the stingless bee Tetragonisca fiebrigi (Jataí). Evid.-Based Complement. Altern. Med. 2015, 296186. [Google Scholar] [CrossRef]
- Paredes-Gamero, E.J.; Martins, M.N.; Cappabianco, F.A.; Ide, J.S.; Miranda, A. Characterization of dual effects induced by antimicrobial peptides: Regulated cell death or membrane disruption. Biochim. Biophys. Acta 2012, 1820, 1062–1072. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moraes, V.W.R.; Caires, A.C.F.; Paredes-Gamero, E.J.; Rodrigues, T. Organopalladium compound 7b targets mitochondrial thiols and induces caspase-dependent apoptosis in human myeloid leukemia cells. Cell Death. Dis. 2013, 4, e658. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | RT (min) | Compound | UV (nm) | MF | Negative Mode (m/z) | Positive Mode (m/z) | ||
|---|---|---|---|---|---|---|---|---|
| MS [M − H]− (*) | MS/MS | MS [M + H]+ (*) | MS/MS | |||||
| 1 | 1.2 | PDE-PDE (B type) | 275 | C30H26O14 | 609.1250 (2.5) | 423, 305, 177 | 611.1411 (2.6) | 287, 263, 179 |
| PDE-PDE-PDE (B type) | C45H38O21 | 913.1803 (3.3) | 423, 305, 261, 243, 177 | 915.1955 (2.5) | - | |||
| di-hexoside | C12H22O11 | 341.1093 (1.0) | - | 365.1051 (0.9) | - | |||
| 2 | 2.4 | Gallic acid st | 270 | C7H6O5 | 169.0140 (1.7) | - | 171.0291 (1.9) | - |
| 3 | 2.9 | PDE-PDE (B type) | 275 | C30H26O14 | 609.1280 (4.9) | 423, 305, 177, 165 | 611.1392 (0.6) | 425, 299, 287, 275, 263, 179 |
| 4 | 3.6 | PDE-PDE (B type) | 276 | C30H26O14 | 609.1254 (0.8) | 423, 305, 177, 165 | 611.1395 (1.6) | 425, 299, 287, 263, 179 |
| 5 | 4.2 | PDE-PDE (B type) | 275 | C30H26O14 | 609.1259 (1.6) | 423, 305, 177, 165 | 611.1400 (0.7) | 425, 299, 287, 275, 263, 245, 179 |
| 6 | 4.3 | Gallocatechin st | 275 | C15H14O7 | 305.0673 (2.0) | 179 | 307.0812 (3.2) | 163, 159 |
| 7 | 6.1 | PDE-PDE (B type) | 275 | C30H26O14 | 609.1245 (0.8) | - | 611.1422 (4.3) | - |
| 8 | 10.3 | Epigallocatechin st | 274 | C15H14O7 | 305.0673 (1.9) | 167 | 307.0823 (3.4) | 195, 177, 163, 159 |
| 9 | 13.5 | PRO-PDE (B type) | 280 | C30H26O13 | 593.1312 (1.9) | 305, 177 | 595.1446 (0.1) | 427 |
| 10 | 15.5 | C-hexosyl O-pentosyl 5,7-dihydroxychromone | 257, 285, 327 sh | C21H26O13 | 485.1312 (2.4) | 353, 335, 245, 233, 215, 205 | 487.1460 (2.9) | 355, 337, 319, 289, 259, 235, 205 |
| 11 | 17.3 | C-hexosyl O-pentosyl 5,7-dihydroxychromone | 257, 295, 325 sh | C21H26O13 | 485.1317 (3.3) | 365, 353, 335, 263, 245, 263, 233, 215, 205 | 487.1452 (1.0) | 319, 301, 283, 259, 235, 205 |
| 12 | 19.0 | PCY-PDE (B type) | 278 | C30H26O13 | 593.1308 (1.3) | 305 | 595.1453 (1.1) | 427, 307, 289 |
| 13 | 19.8 | PDE-PDE benzoate (B type) | 278 | C37H30O16 | 729.1461 (3.5) | 423, 305, 287, 261, 177 | 731.1611 (0.6) | 425, 407, 299, 287, 275, 263 |
| 14 | 23.5 | NI | 280 | C26H32O12 | 535.1840 (3.6) | - | 537.1981 (2.7) | - |
| 15 | 35.4 | NI | 290, 330 | C32H36O16 | 675.1946 (2.2) | 245, 233, 215, 207 | 677.2088 (1.8) | 235, 191, 163 |
| 16 | 36.2 | NI | 300 | C33H38O17 | 705.2068 (4.5) | 573, 467, 365, 335, 317, 245, 237, 233, 215 | 707.2193 (1.6) | 325, 221, 191 |
| Methods | Ascorbic Acid | SAAE | ||||
|---|---|---|---|---|---|---|
| IC50 | Maximal | Inhibition | IC50 | Maximal | Inhibition | |
| µg/mL | % | µg/mL | µg/mL | % | µg/mL | |
| ABTS | 1.34 ± 0.01 | 99.67 ± 0.04 | 5 | 1.83 ± 0.15 | 99.68 ± 0.08 | 10 |
| DPPH | 2.65 ± 0.03 | 87.44 ± 2.13 | 10 | 3.81 ± 0.02 | 89.92 ± 1.36 | 50 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baldivia, D.D.S.; Leite, D.F.; Castro, D.T.H.d.; Campos, J.F.; Santos, U.P.d.; Paredes-Gamero, E.J.; Carollo, C.A.; Silva, D.B.; De Picoli Souza, K.; Dos Santos, E.L. Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. Int. J. Mol. Sci. 2018, 19, 2432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082432
Baldivia DDS, Leite DF, Castro DTHd, Campos JF, Santos UPd, Paredes-Gamero EJ, Carollo CA, Silva DB, De Picoli Souza K, Dos Santos EL. Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. International Journal of Molecular Sciences. 2018; 19(8):2432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082432
Chicago/Turabian StyleBaldivia, Débora Da Silva, Daniel Ferreira Leite, David Tsuyoshi Hiramatsu de Castro, Jaqueline Ferreira Campos, Uilson Pereira dos Santos, Edgar Julian Paredes-Gamero, Carlos Alexandre Carollo, Denise Brentan Silva, Kely De Picoli Souza, and Edson Lucas Dos Santos. 2018. "Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens" International Journal of Molecular Sciences 19, no. 8: 2432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19082432