The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease


1
Department of Biochemistry and Chemistry of Nutrition, Faculty of Veterinary Medicine, Cairo University, Giza 12211, Egypt
2
Department of Anatomy, Cell Biology and Physiological Sciences, Faculty of Medicine, American University of Beirut, Beirut, Lebanon
3
Department of Physiological Chemistry, University of Veterinary Medicine Hannover, Hannover 30559, Germany
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(9), 2560; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092560
Submission received: 6 August 2018
/
Revised: 23 August 2018
/
Accepted: 27 August 2018
/
Published: 29 August 2018
(This article belongs to the Special Issue Molecular Chaperones)
Abstract
:The mammalian HSP90 family of proteins is a cluster of highly conserved molecules that are involved in myriad cellular processes. Their distribution in various cellular compartments underlines their essential roles in cellular homeostasis. HSP90 and its co-chaperones orchestrate crucial physiological processes such as cell survival, cell cycle control, hormone signaling, and apoptosis. Conversely, HSP90, and its secreted forms, contribute to the development and progress of serious pathologies, including cancer and neurodegenerative diseases. Therefore, targeting HSP90 is an attractive strategy for the treatment of neoplasms and other diseases. This manuscript will review the general structure, regulation and function of HSP90 family and their potential role in pathophysiology.

1. Introduction
Serendipity played a decisive role in the discovery of what is now known as heat shock proteins (HSPs). In the laboratory of Italian scientist Ferruccio Ritossa [1], He examined flies, Drosophila melanogaster, that were left un-intentionally at an elevated temperature, and noticed a characteristic puffing in the chromosomes. He reasoned that this “puffing” phenomenon is due to the increased expression of specific proteins due to gene activation following heat stress. Hence, the name “heat shock proteins” came to light [2]. The discovery of heat shock proteins resulted in unraveling the heat shock response (also known as cellular stress response). This response is a common physiological process among almost all living organisms and includes alterations in the gene expression profile through increased HSP expression [3]. Overexpression of HSPs was found to promote cell survival and protect cellular proteins from the risk of damage or aggregation. In addition to heat shock, several factors increase the expression of HSPs including environmental and chemical factors, non-stress physiological causes and disease conditions [4].
HSPs are classified according their molecular mass, where HSP70 for instance meant subset of HSPs having molecular weight of 70-kDa. Due to the increasing numbers and confusing names of HSP members, which likely share a high degree of similarity in some cases, or differ greatly in other cases, there was a great need to classify these large overlapping proteins. Indeed, Kampinga and his colleagues [5] set a new letter/number based classification to simplify and unify the nomenclature of human HSPs. For instance, the previous names of HSP families—HSP70, HSP90, HSP110 and small HSPs (sHSPs)—which are still usable in the scientific community, were referred to as HSPA, HSPC, HSPH, and HSPB, respectively. Moreover, the members within each family were given numbers as HSPA1, HSPA2, HSPA6, HSPA8, etc.
The members belonging to HSP90 protein family are highly conserved ubiquitous molecules with an approximate molecular weight of 90-kDa. They are molecular chaperones promoting the folding of de novo synthesized or incorrectly folded proteins, thus counteracting their aggregation. They exist in all living kingdoms except for archaea [6] and represent about 1–2% of the total mammalian cellular proteins under non-stress conditions [7]. In mammals HSP90 chaperones include members that are arranged into various cellular compartments. Two forms, HSP90 alpha and HSP90 beta, are located in the cytoplasm, GRP94 (94-kDa glucose-regulated protein) exists in the ER, and TRAP-1 (tumor necrosis factor receptor-associated protein 1) is present in the mitochondria. HSP90 proteins take part in essential cellular processes and regulatory pathways like apoptosis, cell cycle control, cell viability, protein folding and degradation, and signalling events. In addition, they induce the adaptive immunity by activation of antigen presenting cells and dendritic cells. Their prominent role in proteostasis, cell differentiation, and development is well recognized. Importantly, overexpression of HSP90 has been linked to many pathological conditions, like several types of cancer, viral infection, inflammation, and neurodegenerative diseases, suggesting a potential role in the pathogenesis of many diseases [7,8,9].
2. Classification, Structure, and Molecular Characteristics
2.1. Classification
According to the new guidelines for HSP nomenclature, the human HSP90 family includes five members that are categorized under the superfamily name HSPC [5]. The recent names together with the alternative old names of HSP90 members are listed in Table 1.
HSP90 proteins can be classified according to their cellular localization, where cytoplasmic members include HSPC1 (HSP 90-alpha), HSPC2 (HSP 90-alpha A2), and HSPC3 (HSP 90-beta), ER resident member; HSPC4 or GRP94 (GP96), and mitochondrial localized member; and HSPC5 (TRAP1). Further classification of HSP90 is based on their expression pattern. For instance, Hsp90β is constitutively expressed, while Hsp90α is induced upon stress conditions [10,11].
2.2. General Structure of HSP90 Isoforms
The structural, biochemical, and molecular characteristics of HSP90 have been thoroughly reviewed [12,13,14]. We present in this manuscript a brief overview to provide the basic background for understanding HSP90 function under physiological and pathological conditions.
The overall molecular structure of HSP90 homologues comprises three main conserved domains; N-terminal domain (NTD), C-terminal domain (CTD), and middle domain (MD) [7,15,16,17,18]. In eukaryotes, there is a variable charger linker domain which links the NTD to the MD [19]. Each domain within the HSP90 structure performs a specific function. The NTD binds to the ATP, hence, called the nucleotide-binding site. The CTD is responsible for protein dimerization and contains special motifs MEEVD or KDEL depending on the HSP90 isoform and its cellular location either in the cytoplasm or the ER. Although it has a divergent sequence among many eukaryotic organisms, the charged linker domain was found to be essential in the chaperone function, interaction, and flexibility [19]. A schematic representation of the HSP90 topology is shown in Figure 1.
2.2.1. Molecular Characteristics of the Cytoplasmic Isoforms: Hsp90α and HSP90β
There are two main mammalian isoforms of HSP90 [17], presumably products of gene duplication [20], localized in the cytoplasm; the inducible form Hsp90α and the constitutive form HSP90β. Therefore, it is not surprising for the cytoplasmic HSP90 proteins to be highly homologous, with 85% sequence identity [21]. Although these cytoplasmic HSP90 isoforms have recently shown distinctive functions [17], most reviews do not distinguish them due to their remarkable structural and functional similarity. Therefore, the name HSP90 has been used for both HSP90 α and β unless indicated [18,22]. Studies dealing with the differentiation between cytoplasmic HSP90 isoforms, revealed that although both isoforms very often form dimers, HSP90α tends to dimerize frequently compared to HSP90β [17]. In addition, the α and β isoforms vary in specific regions along the protein sequence, suggesting diverse functions of the two isoforms.
Under physiological conditions, dimerization is essential for proper functioning of HSP90 [12]. Mammalian HSP90 is a phosphorylated dimer containing 2–3 covalently-bound phosphate molecules per monomer [18]. Moreover, the availability of other factors, such as divalent cations and nucleotides and elevated levels of HSP90 concentrations cause oligomerization of the molecule [18]. Structural studies, including sedimentation [23,24] and electron microscopy [25], revealed that dimeric forms of the protein have elongated structure [25]. As a hydrophobic chaperone, heat shock increases its hydrophobicity [26]. Another aspect is because HSP90 has an ATP binding site in its NTD, self-phosphorylation is likely to occur with profound conformational changes. Interestingly, although well known to have an ATP binding domain, HSP90 was found to have both ATPase and GTPase activity. In the presence of calcium, purified HSP90 was three-fold higher GTPase than ATPase [27].
The N-Terminal Domain (NTD)
Structurally, the N-terminal domain of human HSP90 appeared to be very similar to that of yeast HSPs [28]. It contains an ATP binding motif which shares high conservation with the GHKL (Gyrase, HSP90, histidine kinase, and MutL) superfamily and differs from the corresponding motifs of HSP70 or protein kinases [29]. The ATP-binding site is required for HSP90 ATPase activity necessary for the chaperone cycle and binding the HSP90 client proteins [29,30]. Of particular importance, NTD has been utilized as a major target for Hsp90 inhibitors such as geldanamycin (GA) and radicicol (RD). These are natural product inhibitors, which act by competing with ATP for binding the ATP-binding pocket in the NTD of HSP90.
The Charged Linker Region (CR)
Unlike prokaryotes, eukaryotic HSP90 has a short dynamic region, which links the NTD to the middle domain (MD). This domain is highly charged and has variable length and amino acid composition [31]. Since the eukaryotic HSP90 deals with a multitude of proteins, this charged linker region has been suggested to increase the flexibility and dynamicity to cope with the crowded environment of the eukaryotic cells [31,32]. There are contradicting reports about the role of the charged region in modulating HSP90 chaperone activity. While some studies suggest the charged region as a modulator of the chaperone activity [33], others neglect its potential influence on the HSP90 functions [34].
The Middle Domain (MD)
An important characteristic feature of the MD is that it modulates the HSP90 function by binding the γ-phosphate of ATP specified for the NTD thus modulating its ATPase activity [35]. Additionally, several studies demonstrated that this domain is implicated in binding co-chaperones like Aha1 and interacting with client proteins [36].
The C-Terminal Domain (CTD)
The CTD contains two key sites; one for calmodulin binding [37] and the other for HSP90 homodimerization [38]. In addition, it contains a nucleotide-binding site which opens following the occupancy of the N-terminal site and serves as allosteric regulator of the N-terminal ATPase activity [39]. Interestingly, the N- and C-terminal nucleotide binding sites exhibited differential specificities towards their ligands. The N-terminal binding site was found specific to adenosine nucleotides containing an intact adenine ring, like nicotinamide adenine dinucleotides and adenosine polyphosphate alarmones. On the other hand, The C-terminal binding site was much more unspecific by interacting with both purine and pyrimidine nucleotides. GTP and UTP nucleotides showed specificity towards the C-terminal binding site. Moreover, 2′,3′-O-(2,4,6-trinitrophenyl)-nucleotides (TNP-ATP, TNP-GTP) and pyrophosphate were able to bind the C-terminal binding site with no need for occupied N-terminal site [39]. Therefore, certain drugs like novobiocin (NB) [40] and epigallocatechin-3-gallate (EGCG) [41] could affect the function of HSP90 by targeting the CTD. It has been shown that CTD is included in the chaperone activity of HSP90 towards transcription factors and vesicular stomatitis virus G-protein [42]. Targeting the CTD by the AC-88 monoclonal anti-hsp90 antibody could hinder the interaction of HSP90 with many protein clients including steroid receptors and actin filaments [43]. Another characteristic feature of the HSP90 CTD is the existence of the MEEVD peptide sequence which binds the TPR-domain (tetratricopeptide-containing repeats) containing co-chaperones like HOP and immunophilins [44].
2.2.2. Molecular Characteristics of the ER Isoform; GRP94
GRP94 (GP96) is the most abundant glycoprotein in the ER hence known as endoplasmin [45,46]. It is highly conserved among mammalian species. Interestingly the human GRP94 was found to be 100% identical to that of camel species [47]. GRP94 shares 50% homology with the cytoplasmic HSP90 [20]. However, several biochemical similarities exist between GRP94 and its cytosolic homologues including binding and hydrolyzing ATP. However, it binds to the HSP90 inhibitors geldanamycin and radicicol in a slightly different direction [48]. Nevertheless, being the major calcium binding protein in the ER and having specific limited protein clients are properties that clearly distinguish GRP94 from the cytosolic homologues [49].
GRP94 exists in three main conformations; the extended or chair-like conformation, the less extended conformation and the closed or twisted V conformation. The extended conformation confers better surface accessibility for binding client proteins as well as nucleotides [48,50,51]. Indeed, like HSP 90, the GRP94 is very dynamic where the presence of excess nucleotides, client proteins, and co-chaperones shift the conformation to the closed state [49].
Like HSP90, GRP94 has three main structural domains: the NTD, MD, and CTD. Notably, the N-terminal amino acid sequence differs from that of cytosolic HSP90 in both length and sequence [44]. This variability suggests heterologous conformational changes of HSP90 and GRP94. As an ER resident chaperone, GRP94 contains an N-terminal 21 amino acid sequence required for ER targeting. Another important difference between the cytoplasmic and ER HSP90 homologues is that in GRP94, the lid region, a term used to describe amino acid residues 100–121 in the cytosolic HSP90 and serves to couple ATP binding to N-terminal association, contains five additional residues between the two helices [48,50].
Compared to the cytosolic HSP90, the charged linker region is shorter, rich in lysine residues, more acidic, and contains many calcium binding sites [15]. Additionally, the charged linker domain modulates the conformational changes upon ATP hydrolysis and its deletion dramatically affect the GRP94 function and interaction with clients [52]. GRP94 binds calcium in high capacity, however, low affinity as it was known to have 11 low-affinity calcium binding sites and four higher-affinity Ca binding sites. Calcium binding can affect the molecular conformation and the activity of GRP94 [53]. Occupation of the Ca2+-binding site, located in the charged linker domain of GRP94, by calcium augments the binding of client peptides with the peptide-binding site in the N-terminal domain of the protein, thus regulating the capability of GRP94 to bind peptides [53].
The middle and C-terminal domains of GRP94 resemble those in HSP90 except for few differences. The dimerization interface is stricter in GRP94 and the CTD of GRP94 contains an extra 55 amino acids with a KDEL retention signal rather than MEEVD peptide at its end [15].
2.2.3. Molecular Characteristics of the Mitochondrial Isoform; TRAP1
TRAP1 is the mitochondrial version of HSP90, which is localized primarily in the in the mitochondrial matrix and to a few extent in the inter-membrane space [54,55]. It shares high homology with the cytosolic counterpart reflecting 34% identity and 60% similarity [56] and have the same three domain architecture of HSP90: NTD, MD, and CTD. Similarities with other homologues exist where the NTD can bind ATP and it is vulnerable for inhibition by geldanamycin and radidcol. Notably, the binding affinity of TRAP1 to ATP is ten times higher than HSP90 [57,58]. Recently, X-ray diffraction studies showed that calcium can compensate for magnesium to support TRAP1 ATPase activity where the relative preference for magnesium or calcium is dependent upon the free ATP concentration [59]. In addition, calcium binding to TRAP1 induces cooperative ATP hydrolysis by the two HSP90 protomers, whereas magnesium binding causes non-cooperative ATP hydrolysis. Interestingly, heat shock can increase the expression of TRAP1 up to 200-fold [55]. This mitochondrial HSP90 has a 59 amino acid serving as a mitochondrial targeting sequence and is cleaved off upon import to the mitochondria [60]. Indeed, TRAP1 lacks both the C-terminal MEEVD motif and the charger linker domain connecting MD to the CTD which are present in its cytosolic homologue [61]. Moreover, certain co-chaperones, including p23 and Hop, are not required for its function [62].
3. Regulation of HSP90
There are different levels of regulation that control the function of HSP90; transcriptional regulation, posttranslational modification, and co-chaperones [12,63]. These factors can be summarized as follows.
3.1. Transcriptional Regulation
Generally, the expression of HSPs is controlled by a subset of specialized stress related transcription factors named heat shock factors (HSFs). Binding of activated HSFs to their specific place, known as heat shock element (HSE), in the HSP promoter region stimulates the action of RNA polymerase on the coding region of the HSP gene [18]. Recent models propose that binding of HSP90 and HSP70 to HSF1 cause its inactivation. When chaperone function needed, dissociation of HSF1 occurs and HSP transcription is induced [63,64].
3.1.1. Cytoplasmic HSP90
The gene encoding HSP90α is located on chromosome 14 while the gene encoding HSP90β is present in chromosome 6 [17]. Compared to the constitutively expressed HSP90β, HSP90α is highly induced by increased temperature. Several previous studies have demonstrated the promoter region and transcriptional regulation of HSP90 in yeast [18]. In human HSP90α, the promoter region contained a putative SP1 binding site and a serum-response element, together with multiple HSEs suggesting more complex regulation events [65].
3.1.2. GRP94
In humans, the gene encoding GRP94 is known as HSP90B1 and is located on chromosome 12. The regulation of GRP94 follows the same transcriptional regulation as in cytosolic HSP90. However, it is mainly induced by ER stress or overload rather than a simple heat shock. Like GRP78, the GRP94 promoter region has a CG/CAAT and a GC-rich sequence motif, which are essential for basal and induced expression of the genes [18]. Therefore, GRP94 regulation is associated with GRP78 regulation [15,49]. Decreased induction of GRP94 resulted in low induction of GRP78, while increased GRP78 induction caused enhanced GRP94 stimulation.
3.1.3. TRAP1
The gene encoding human TRAP1 is located on chromosome 16. Data are scarce about its transcriptional regulation. However, it appeared to follow the same general regulation of HSPs [60].
3.2. Regulation by Posttranslational Modification
Posttranslational modification (PTM) of HSP90 isoforms modulates their chaperone function in terms of accessibility of the binding sites [63]. The cytoplasmic isoforms; HSP90β and HSP90α undergo different PTMs including phosphorylation, acetylation, SUMOylation, methylation, ubiquitylation, and S-nitrosylation [12,66,67]. The endoplasmic reticulum form, GRP94, can be modified by glycosylation, phosphorylation and acetylation [49]. Additionally, the mitochondrial homologue TRAP1 is subject to phosphorylation [60,68]. However, other PTMs which affect cytosolic HSP90, like acetylation and nitrosylation, cannot be excluded for TRAP1 modification [60].
3.3. Co-Chaperones
Co-chaperones represent the most important layer of HSP90 regulation (Table 2). These molecules assist the chaperon proteins in performing their function. Several reports described many co-chaperones for cytosolic HSP90. A comprehensive up-to-date list of HSP90 co-chaperones can be obtained through (http://www.picard.ch/downloads/Hsp90interactors.pdf and [63,69]. Concrete data about GRP94 and TRAP1 co-chaperones are lacking because of their compartmentalization inside the ER and mitochondria [61,70]. However, recently MZB1 (pERp1) has been shown as a co-chaperone of GRP94 [71].
4. Client Proteins
4.1. HSP90
HSPs are biological catalysts. They mainly bind to their interacting partners through non-covalent binding where their substrates are commonly referred as clients [22]. As molecular chaperones, HSP90 isoforms have wide range of client proteins that are involved in numerous cellular pathways. Compared to other homologues, the cytoplasmic isoforms of HSP90 (Hsp90α and Hsp90β) interact with more clients than the ER resident GRP94 and mitochondrial localized TRAP1. For instance, the interaction list of HSP90 includes kinases such as Akt2, CDKs, PKC, many MAP kinases as well as transcription factors like steroid receptors, BCL-6, CAR, p53, Oct4. A valuable detailed information about the cytoplasmic HSP90 protein clients and co-chaperones is available on the web site sustained by Dr. Didier Picard at http://www.picard.ch/downloads/Hsp90interactors.pdf.
4.2. GRP94 and TRAP1
For these two HSP90 isoforms, the protein client list is relatively short but subject to increase. Sample clients are summarized in the following Table 3.
5. Mode of Action
5.1. ATPase Activity
The central dogma in the HSP90 mechanism of action is its ATPase activity and cycling between the closed and open states [94]. Although located in different compartments inside the cell, all HSP90 isoforms including cytoplasmic, ER, and mitochondrial homologues that act in a similar way in terms of conformational alterations following nucleotide binding [95]. As described earlier, HSP90 exists as a flexible homo-dimer whose monomers consist of three structural domains; NTD, MD, and CTD. Binding of ATP occurs in the ATP binding cleft in the NTD of HSP90 resulting in series of conformational events. An interesting event includes a short piece of the N-domain (ATP-lid), which trans-locates over the binding pocket and attaches to the corresponding N-domain of the other homo-dimer resulting ultimately in a twisted and compacted dimer [96]. As a result, the N- and M-domains get closer by 40 Å leading to finalizing the “split ATPase” site. After ATP hydrolysis, the N domains of the HSP90 homo-dimers dissociate with the release of ADP and Pi while the HSP90 returns to its original open conformation (Figure 2) [13]. Interestingly, ATP hydrolysis in humans occurs at a rate of 10–20-fold slower than in yeast [30,97], suggesting a complex conformational rearrangement of HSP90 in higher eukaryotes [95]. Critically, the well-known anticancer drugs geldanamycin and radicicol bind specifically to HSP90 in a fashion similar to how ATP does, thus serving as potent inhibitors of HSP90 ATPase activity [98,99].
5.2. Chaperone Cycle
The members of HSP90 family perform their folding function of client proteins through a complicated process known as the HSP90 chaperone cycle. Previous reviews demonstrated how HSP90 mediates the folding of its client proteins, however the complete understanding of the protein folding cycle is yet unreached [13,100,101]. In fact, the HSP90 chaperone machinery requires cooperation of different molecules including co-chaperones, partner proteins, and immunophilins, which act in precise and dynamic manner helping efficient protein folding by HSP90 (Figure 3).
6. Roles of HSP90 Isoforms in Cellular Physiology
6.1. Cytosolic HSP90
6.1.1. Chaperone Function
Several previous reports demonstrated the role of HSP90 in protein folding and preventing protein aggregation [108,109,110]. Although HSP90 requires ATP for its action, purified HSP90 could stabilize heat-denatured proteins, including citrate synthase, rhodanase, and protein kinase CK-II [111,112]. In addition, HSP90 prevented further aggregation of mild aggregates of CK-II resulting from low salt exposure [108,113]. Furthermore, experiments involving heat-denatured luciferase [114] and guanidinium.HCl-denatured β-galactosidase [114] showed that HSP90 confers folding competence to these molecules and facilitates their refolding by other chaperones [114]. In addition to its ATP-dependent activity, the ATP-devoid chaperone activity of HSP90 discriminates it from other HSPs like HSP60 and HSP70 [115] and provides complexity to its mode of action [13,116]. Noteworthy, HSP90 acts in cooperation with other chaperones, including HSP70 and HSP40, as well as co-chapeones, like TPR-containing co-chaperones, to refold many denatured proteins, such as steroid receptors and protein kinases [63].
6.1.2. Cell Signaling
HSP90 protein clients are associated with several signaling pathways like steroid hormone receptors and protein kinases (the link to the current client list is provided above). Similar to steroid receptors, HSP90 appeared to be essential in the maturation of the aryl-hydrocarbon (dioxin) receptor [117]. Other reports demonstrated that decreased expression of HSP90 deactivated retinoid receptors and hindered the development of high-affinity retinoid acid binding [118]. Likewise, HSP90 formed complexes with a large number of protein kinases where the first-described protein kinase in complex with HSP90 was the v-Src tyrosine kinase [119,120]. Binding of v-Src to HSP90 causes hypophosphorylation and abolishes protein kinase activity [18]. Raf kinase also forms a complex with HSP90 which facilitates its membrane association [121]. HSP90 has been shown to protect kinases from the action of phosphatases [122]. Interestingly, premature dissociation of HSP90 complexes, including Src- and Raf- hsp90 complexes, could be achieved by the hsp90-specific drugs geldanamycin and radicicol [123]. Dissociation of such kinases ultimately leads to enhanced proteasomal degradation [124]. Other kinases such as the tetratricopeptide domain-containing immunophilin (PP-5) and transcription factors have been reported to be a part of the HSP90 complex. Therefore, it is not surprising that HSP90 is designated as a key chaperone implicated in cell signaling.
6.1.3. Cytoskeleton
Due to its abundance and adhesion properties, cytosolic HSP90 was named as “molecular glue” [18]. HSP90 was found to crosslink actin filaments in vitro [25]. However, HSP90 was not co-localized with actin in human fibroblasts or human endometrial adenocarcinoma cells. Many early studies suggested a dynamic transient interaction of HSP90 with almost all cytoplasmic filamentous structures highlighting its role in maintaining the cyto-architecture [18]. Further studies showed that binding of HSP90 to actin filaments favors an ATP-lacking environment which mostly occurs in severe stress conditions [125,126]. Additionally, HSP90 appeared to modulate actin filament bundling behavior [127]. In addition to actin, HSP90 has been reported to interact with tubulin and protect it from heat denaturation [128], microtubules [129], and other non-microtubular and non-microfilamental structures of the cytoplasm. Furthermore, HSP90α has been shown to protect myosin from heat stress [130]. Therefore, HSP90 is involved in the arrangement and maintenance of almost the whole cytoskeleton.
6.1.4. Nuclear Functions
Under normal conditions, about 5–10% of total HSP90 exist in the nucleus of the cell. This nuclear localization increases upon heat stress [18,131]. HSP90 is associated with nucleoli and perichromatin ribonucleoprotein fibrils [132]. In fact, HSP90 contains a bipartite nuclear localization sequence in the middle, highly-charged region of the protein. Additionally, nuclear import of HSP90 is modulated by FKBP52, steroid receptors, and kinases [27,133]. Furthermore, the sequence of HSP90 contains other sequences homologous to the recognized traditional or alternative nuclear import and export signals [18]. Since it served to transport steroid receptors and kinases to the nucleus, it has been suggested that HSP90 binds NLS [134]. Importantly, HSP90 binds to DNA, RNA, and histones where it modulates the structure of DNA and contributes to RNA synthesis and processing [18]. In addition to histones, HSP90 interacts and forms complexes with many transcription factors, including zinc finger proteins, helix-loop-helix proteins, MyoD1, E12, hypoxia-inducible factor 1α, and heat-shock factor 1 [18].
6.1.5. Cell Cycle and Cellular Differentiation
HSP90 and the co-chaperone CDC37/p50 fold cell cycle-related protein kinases including cyclin-dependent kinase CDK4, cyclin-dependent kinase CDK11p110 [135] and the cyclin-dependent kinase regulator Wee1 [136]. Interestingly, the expression profile of HSP90 isoforms differs according to the stages of the cell cycle [137]. Transcription of HSP90α is stimulated at the G1/S transition of chicken hepatoma cells [138]. In fibroblasts, the ATP concentration has been shown to rise two-fold during mitosis [139]. Variations in ATP levels during cell cycle stages dramatically affect the functions of HSP90 isoforms. In addition to cell cycle changes, cell differentiation also caused alterations of HSP90, as well as GRP94 expression. Differentiation of embryonal carcinoma cells resulted in upregulation of HSP90β [140]. Interestingly, the expression level of HSP90α and HSP90β is reduced during osteoblast and HL-60 cell differentiation [141] suggesting that the expression pattern of HSP90 during differentiation is cell-type dependent [18]. Additionally, genetic mutation of HSP90β led to diminished hepatocyte differentiation from induced pluripotent stem cells [142]. HSP90 was found to stabilize the peroxisome proliferator-activated receptor-γ (PPARγ) thus, regulating adipose tissue differentiation and function [143]. Another interesting point, the expression profile of the cytoplasmic isoforms Hsp90α and HSP90β change during the differentiation of myoblasts into myotubes where HSP90β dominates and interacts with the muscle-specific HSP90 co-chaperone Aarsd1L [144].
6.1.6. Extracellular HSP90 or Secreted HSP90
Generally, initial reports concerning the idea of HSP secretion faced difficulties in convincing the scientific community about the fact that HSPs are released or secreted from intact cells [145]. Later, HSP70 was proven to be secreted from cultured rat embryo cells [146]. Further studies confirmed the secretion of many classes of HSPs, including small HSPs and HSP90, from different cells where they are suggested to perform specific immunological functions [147]. HSP90α has been secreted from cells following many stresses such as reactive oxygen species, heat, hypoxia, irradiation, and tissue injury release cytokines [15]. There has been a strong notion that normal cells do not secrete HSP90 provided being elicited by environmental influences [148]. Further studies suggested that the secretion of HSP90α occurs as a result of TGFα induction [149] and is regulated by the hypoxia-inducible factor-1alpha (HIF-1α) [150,151]. eHsp90α has been shown to enhance cell motility and support wound healing [148]. Interestingly, it was found that eHSP90α acts in a way different from its cytosolic homologues as the ATPase activity was not required for its function in cell motility [15]. For that, eHSP90 function was dependent on a fragment (F-5) including residues 236–350 and located between the linker region and the middle domain [15,152].
6.2. GRP94
6.2.1. Protein Folding
As a molecular chaperone, the main task of GRP94 is to promote the folding and assembly of secretory and membrane proteins [49]. An important characteristic that distinguishes GRP94 from other ER chaperones is its functional selectivity, which is reflected by its relatively short client protein list. In agreement with the selectivity hypothesis, GRP94 is not essential for folding or trafficking of various plasma membrane receptors and secretory molecules including transferrin receptor and MHC class I [49,90]. Moreover, in contrast to BiP, which has been shown to bind both V and C domains, GRP94 binds specifically to the light chain through Ig-fold variable domains. Furthermore, secretion of light chains was inhibited in GRP94 deficient B-cells [153]. Bile salt-dependent lipase (BSDL), also known as cholesterol esterase, represent another example of GRP94 client proteins forming complex to be secreted from pancreatic cells [154,155]. Known client proteins of GRP94 include Toll-like receptors and integrins, which require the ER chaperone for efficient trafficking and cell surface expression [156,157]. Interestingly, gp93 the drosophila homologue of GRP94 could help surface expression of many integrins and TLRs in mammalian cells lacking GRP94 [157]. Also, GRP94 interact with the platelet glycoprotein Ib-IX-V complex, a receptor for Willebrand factor whose absence results in defective platelets function. The interaction with GRP94 guarantees good surface expression and improves the ER-associated degradation (ERAD) of the complex [158]. Other clients include thyroglobulin, insulin-like growth factors, and others that have been listed in Table 3.
6.2.2. Calcium Homeostasis
GRP94 is a calcium binding protein which has low affinity, high capacity binding to calcium [46,159]. Together with other ER chaperones, it mediates calcium homeostasis in the ER and the cell. Although important, calcium binding by GRP94 was found not essential for life. Skeletal muscle cells which lacked GRP94 did not show clear defect in calcium homeostasis suggesting another compensating mechanism for calcium by calsequenstrin-1 [160].
6.2.3. ER Quality Control
To produce correctly folded secretory and membrane proteins, there is a network of chaperones inside the ER which control and maintain this process. GRP94 acts as a major checkpoint in this network. The function of GRP94 in the quality control of the ER can be summarized in terms of (1) chaperoning protein folding intermediates; (2) interaction with other components of the ER protein folding machinery; (3) calcium storage; and (4) helping targeting of mis-folded proteins to ERAD [161].
6.2.4. ER Stress
GRP94 and BiP (GRP78) are considered the hallmarks of the unfolded protein response (UPR). Both chaperones and other ER chaperons have analogous promoters, therefore, usually induced concomitantly under physiological as well as pathological conditions [49]. Overexpression of GRP94 have been reported in physiological events like differentiation of hepatic cells, acinar cells, and B cells [49,162]. Enhanced folding and promotion of IGF-1 and IGF-II by GRP94 leads to increased hormone output under stress and boosting IGF pathway. These consequences lead to improving the UPR, increasing the folding capacity of the ER, suppression of apoptosis and alleviating metabolic stresses [49].
6.2.5. Extracellular GRP94
Earlier investigations reported that GRP94 can be released outside the cells where its secretion was associated with severe tissue damage and necrotic cell death [163,164]. These studies suggested that necrotic, rather than apoptotic, cells produce extracellular forms of the ER chaperone [163]. GRP94 has been known to chaperone intracellular peptides as well as tumour-specific peptide and present them on the MHC class I molecules of antigen presenting cells (APCs) in a process called “cross-presentation” [165,166,167]. Moreover, although mainly localized in the ER, GRP94 was expressed on the cell surface of tumour cells [168]. Evdokimovskaya and his colleagues reported the release of GRP94 from BHK cells under normal culture conditions [165]. Moreover, we reported the secretion of GRP94 from other cell lines including COS-1 and CHO-K1 cells as well [47]. In fact, other ER chaperones like GRP78 and calreteculin have been reported to be released to the extracellular space where they are suggested to display immunogenic and inflammatory related roles [147,169]. These data strongly point to the possibility that released forms of GRP94 can perform immunogenic functions even under non-pathologic conditions. However, this notion requires further investigation [47].
6.3. TRAP1
6.3.1. Role of TRAP1 in Mitochondria
The major function of TRAP1 is to maintain mitochondrial integrity and protect against mitochondrial apoptosis [55]. Oxidative stress resulting from increased production of oxygen free radicals or reactive oxygen species (ROS) is the main cause of mitochondrial dysfunction and cell death. Various diseases such as neurodegenerative, cardiac, neoplastic maladies and stroke [170] are potential consequences of excessive ROS production. It has been shown that TRAP1 can protect against cell death induced by over production of ROS [171,172]. Over-expressed TRAP1 could reduce ROS accumulation while its knock-down increased ROS formation [173,174]. Furthermore, TRAP1 counteracts protein aggregation inside the mitochondria and support protein folding [175] leading to healthy, intact mitochondria.
6.3.2. How Does TRAP1 Protect Against Oxidative Stress?
Accumulated ROS causes opening the permeability transition (PT) pore of the inner mitochondrial membrane leading to disturbed membrane potential [176]. As a result, the mitochondria are swollen and the outer membrane can be ruptured resulting in cell mortality [177]. Studies on PT pore showed that the voltage-dependent anion channel (VDAC), located in the outer mitochondrial membrane, and the adenine nucleotide translator (ANT), located in the inner mitochondrial membrane, are not the lone PT pore components [178,179,180]. Critically, the mitochondrial matrix protein, cyclophilin D (Cyp-D), was found to play a key role in the pore formation of the mitochondrial inner membrane [181]. Being a peptidyl-prolyl cis-trans isomerase located in the mitochondrial matrix, Cyp-D supports protein folding of the PT pore proteins inside the mitochondria. In addition, increased ROS, calcium, and diminished ATP following stress conditions cause activation of Cyp-D which, in turn, modulate the conformation of PT pore proteins and consequently generate non-selective pores through the membrane [177,182,183]. TRAP1 is a major interacting protein with Cyp-D, hence, greatly influences its function. For instance, TRAP1 can hinder the Cyp-D pore opening activity in various cell types including tumour cells allowing cell survival [184,185]. Although the ATPase activity of TRAP1 has been thought to be essential in its inhibiting ability of Cyp-D, treatment of cells with TRAP1 inhibitors did not affect its function regarding Cyp-D inhibition [185]. These findings indicate that the chaperone activity rather than the interaction issue is the mainstay in Cyp-D inhibition by TRAP1 [60].
6.3.3. Extra-Mitochondrial Roles
A cohort of the mitochondrial TRAP1 molecules can be trans-located outside the mitochondria and bind extra-mitochondrial proteins. TRAP1 has been reported to interact with TNF receptor [56] retinoblastoma protein [74], and tumour suppressor EXT proteins [76]. Additionally, during mitosis, TRAP1 interacts with the retinoblastoma protein to help its folding [74]. Similar to TRAP1, the tendency to perform extra-mitochondrial tasks is a common feature of other mitochondrial chaperones, like HSP60 and mortalin, especially in cancer cells [186]. Although the role of TRAP1 in maintaining mitochondrial integrity, defending against oxidative stress, and inhibiting cell death have been addressed in many reports, how it is exported to the cytoplasm and what it exactly does there is still beyond our knowledge [60].
7. Role of HSP90 Isoforms in Pathological Conditions
Disease, in general, induces a kind of stress on the cellular machinery. Therefore, variant types of HSPs including HSP90 are involved in diverse pathological conditions like cancer, neurodegenerative diseases, infectious diseases, and others. Here, we discuss the major common diseases that have been linked to members of HSP90 family.
7.1. Cytosolic HSP90
7.1.1. Cancer
Normal mammalian cells are equipped with defined molecular machinery sufficient to regulate their proliferation, growth, differentiation, and death. On the other hand, cancer cells are characterized by persistent uncontrolled proliferation and lacks the native regulatory mechanisms responsible for normal cell proliferation and homeostasis [187]. Several cancer proteins depend on HSP90 machinery and chaperones for their folding and maturation [188]. For instance; signaling kinases, steroid hormone receptors and transcription factors, which are highly required in cancer development and progression, represent a group of HSP90 client proteins (Table 4). Therefore, pronounced expression of HSP90 has been detected in almost all classes of cancers. A new theory named “addiction to chaperones” has been adapted to explain the great demand for HSPs including HSP90 during cancer growth [22]. According to this hypothesis, overproduction of mutant proteins in cancerous cells act as a fuel for HSPs induction, increased translational processes, and expression of proteins leading to transformation [22].
HSP90 as a Target for Anti-Cancer Drugs
Due to the higher sensitivity of cancer cells to HSP90 inhibitors than healthy cells, the cytosolic molecular chaperone HSP90 has attracted a great deal of interest in the therapeutic field of cancer. Natural products like geldanamycin (GA) that was derived from Streptomyces hygroscopicus and the antibiotic radicicol (RD), isolated from Monosporium bonorden exhibited prominent suppression of tumor proliferation and growth [189,190]. Having similar structures to ATP, these HSP90 inhibitors bind the ATP binding pocket of the NTD within the HSP90 structure, thus interfering with ATP binding and hydrolysis, consequently leading to the depletion of HSP90 oncogenic clients [98,191]. Although the anti-cancer efficacy of GA has been tested in many in vivo and in vitro studies, it has not yet been approved for clinical uses due to its structural instability and hepatotoxicity. However, GA is still commonly used for in vitro studies concerning HSP90 inhibition [192,193]. Similarly, RD usage was limited due to its instability as well [194]. Further chemical modification of GA specifically by substituting its C17 with an allylamino group, producing 17-AAG (17-allyl-17-demethoxygeldanamycin) greatly enhanced its toxicity profile [195]. Therefore, 17-AAG have been used in phase I/II clinical trials [190,196]. Other strategies suggest a combination therapies including 17-AAG and other chemotherapeutic drugs or radiotherapy as a successful approach for cancer therapy [15]. Recently, many other derivatives have been developed from GA and RD, including second-generation HSP90 inhibitors, along with other drugs, which target the middle and C-terminal domain of HSP90 (see [190,197] for reviews).
7.1.2. Neurodegenerative Diseases
HSP90 plays an additional role in protecting neuronal proteins with aberrant aggregation tendency from accumulation and forming toxic aggregates [8]. HSP90 counteracts protein aggregation accompanying neurodegenerative diseases like Alzheimer disease and Parkinson disease [63]. Intriguingly, in Alzheimer disease HSP90 appeared to promote the hyper phosphorylation of tau protein (tau-p) via activation of kinases leading to the formation of neurofibrillary tangles [63,198]. Targeting HSP90 by inhibitors led to reduction in p-tau levels through a mechanism mediated by the tau ubiquitin ligase; carboxy terminus of Hsp70–interacting protein (CHIP) [198]. In Parkinson disease, in vitro experiments revealed that HSP90, as well as HSP70/40 could suppress Aβ amyloid self-assembly suggesting that induction of chaperones’ overexpression by pharmacological substances has potential therapeutic value for neurodegenerative diseases [110]. In agreement with this hypothesis, overexpression of HSP70, following treatment with drugs inducing its expression as geldanamycin, could reduce α-synuclein induced neurotoxicity in human H4 neuroglioma cells, as well as a fly model of Parkinson disease [199,200,201].
7.1.3. Infectious Diseases
Viral Diseases
Increasing reports suggest the involvement of HSP90 in viral protein homeostasis [9]. Numerous viral proteins have been shown to utilize HSP90 for folding, assembly and maturation. Viral polymerases represent the major subset of viral proteins requiring HSP90 machinery for their processing. For instance, reverse transcriptase (RT) of duck hepatitis B virus (DHBV) needs HSP90 together with other chaperones, including HSP70/HSP40, and a co-chaperone, like HOP and p23, as substrate release factors and supporting incorporation of the pgRNA into nucleocapsids [202,203,204]. Additionally, the genome replication of influenza virus A needs HSP90 for its replication [9]. A detailed list of viral proteins requiring HSP90 has been reported previously [9]. It seems that the demand for HSP90 chaperone in viral infections comes from the increased translational rates of viral proteins, high incidence of mutations, as well as high conformational and proteolytic events. These stressful processes necessitate the presence of HSP90 as a buffering system [63].
Parasitic Diseases
Interestingly, the malarial parasite Plasmodium falciparum requires the HSP90 machinery in the human host to withstand the surrounding environmental stresses including pH and periodic elevated temperature fluctuations in the patient [205]. Similarly, Leishmania donovani, the causative agent of leshmaniasis, requires HSP90 during its life cycle and pathogenicity [206]. Therefore, it is not surprising that Hsp90 forms about 3% of total protein content in the promastigote stage of L. donovani [207]. Moreover, there are 17 homologues of HSP90 genes in Leishmania major presumably functioning in its pathogenesis [15,208]. In other nematodes, HSP90 has been reported to affect the morphological features by canalizing the mouth-form plasticity in Pristionchus pacificus [209].
7.1.4. Other Diseases
HSP90 is implicated in the pathogenesis of other serious diseases, like cystic fibrosis (CFTR), atherosclerosis, and diabetes. The deletion mutation (ΔF508), of CFTR, seriously affect the protein folding and causes its degradation [63]. HSP90 was found to interact with the wild-type, as well as mutant, CFTR. As a part of the HSP90-protein complex, AHA1 appeared to play a key function in stabilizing the mutant CFTR within the HSP90 chaperone complex. Knockdown of AHA1 leads to a high number of mutant CFTR channels transferred to the plasma membrane, suggesting that the AHA1 co-chaperone mediates processing and degradation of mutant CFTR by blocking its escape from the chaperone cycle [63,210].
In patients with atherosclerosis, expression of HSP90 is increased in atherosclerotic plaques and has been associated with plaque instability [211]. Interestingly, HSP90-inhibiting drugs could diminish plaque formation by suppressing the migration and proliferation of vascular smooth muscle cells (VSMCs) [211]. Other studies reported that the high HSP90 expression is not restricted to the atherosclerotic plaques. The serum of atherosclerotic patients contained elevated HSP90 levels presumably inducing immune response, thus, HSP90 have been considered as a possible target auto-antigen in the pathogenesis of carotid atherosclerosis [212].
Recent studies reported elevated HSP90 levels in newly-diagnosed type I diabetic patients suggesting HSP90 as a potential biomarker for this serious disease [213]. Additionally, experiments in a mouse model for diabetes-accelerated atherosclerosis showed that inhibition of HSP90 was found to reduce both inflammation and oxidative stress in addition to limiting vascular and renal damage [214].
7.2. GRP94
7.2.1. Cancer
As a molecular chaperone implicated in the quality control of the ER, it is not surprising that GRP94 chaperones several proteins essential in carcinogenesis [215], including TLRs (with the exception of TLR3) [156], Wnt co-receptor LRP6 [216], GARP [217], GPIb [158], and Insulin-like growth factor [218], as well as most of the α and β integrin subunits [219,220]. GRP94 has been known as an active tumor rejection antigen that can immunize recipients against tumor transplants [221]. Complexes of GRP94 and its chaperoned tumor peptides rather than GRP94 alone seemed to be responsible for GRP94 immunogenicity. Overexpression of GRP94 has been reported in multiple types of neoplasms where high GRP94 expression was clinically indicative for advanced cancer stage and poor prognosis [215], like head and neck cancer [222], gallbladder cancer [223], and breast cancer [224]. Other studies showed that overexpressed GRP94 encouraged the growth and metastasis of certain cancers, including hepatocellular carcinoma [225], multiple myeloma [226], ovarian cancer, and inflammatory colon carcinomas [227]. Localization studies revealed that GRP94 favors cell surface expression in cancerous cells suggesting excess immunogenic roles [228,229]. These immunological functions include increased antigen presentation and enhanced T-cell activation [229]. Therefore, it can be concluded that GRP94 exhibits double roles in carcinogenesis; (1) supporting cancer progression, in terms of assisting the folding of cancer related proteins; and (2) induction of immune response against cancer through cross-presentation of tumor antigenic peptides and T-cell activation [215].
GRP94 as a Target for Anti-Cancer Drugs
It has been shown that cancerous cells exhibit high surface expression of GRP94, although it is ordinarily localized in the ER [221]. Other studies demonstrated that increased cell surface expression of GRP94 is associated with tumor immunogenicity [228]. Further studies proved that GRP94 could present antigenic peptides which have anti-tumor activity [230] highlighting the critical role of GRP94 in induction of immune response against tumors [215]. Theses previous studies were the basis for GRP94-based antitumor therapies. GRP94-based antitumor vaccines could protect against methylcholanthrene-induced fibrosarcomas (Meth A) tumors [231]. Specific monoclonal antibodies against GRP94 were used as powerful tools to inhibit cancer growth and development in tumor tissues. For instance, the monoclonal antibody (W9 mAb) was recently used and precisely targets the extracellular epitope of gp96 which is barely found in normal cells [232]. Another study used GRP94 or gp96 monoclonal antibody (gp96 mAb) to interrupt the HER2 dimerization and phosphorylation in breast cancer, which lead ultimately to inhibiting their growth and proliferation both in vitro and in vivo [215].
Chemicals targeting GRP94 have also been used as promising anti-cancer remedies. The main concern in this strategy was the pan- inhibition of other HSP90 family members in normal cells as well which may lead to disruption of vital biological process relying on them [233]. This underlines the need for developing more specific and selective anti-cancer therapies. Documented examples of GRP94 selective drugs include 5′-N-ethylcarboxamidoadenosine (NECA), Compound 2, and Radamide [234,235]. Other strategies developed selective GRP94 compounds based on conformational investigations and screening of purine-scaffold compounds. These studies produced gp96 inhibitory ligands as PU-H54, PU-WSI3, and PU-H39, which had distinct structural features in the ATP-binding pocket [236]. In summary, there are promising beneficial clinical impact for developing gp96-selective inhibitors rather than using drugs with pan-inhibition effect on the HSP90 machinery [215].
7.2.2. Neurodegenerative Diseases
Among the major consequences of neurodegenerative diseases is the accumulation and aggregation of mis-folded proteins within almost all cellular compartments. Cellular responses including ER stress, unfolded protein response (UPR) and over expression of HSPs serve as protective mechanisms to counteract these protein aggregations [237,238]. The ER chaperone GRP94 is a key player in the quality control of the ER helping refolding or disposal of mis-folded and aggregated proteins [161]. Increased secretion of GRP94 and lower intracellular levels have been associated with amyloid-β (Aβ) peptide-induced neurotoxicity in rat neuronal-like PC12 cells [239]. Recently, a vaccine composed of combination of α-synuclein (α-Syn) and Grp94 chaperone could provide a marked long-lasting immune profile in the peripheral system, which has an influence on the CNS by inhibiting chronic microglial activation in chronic mouse 1-methyl-4-phenyl-1,2,3,6,-tetrahydropyridine (MPTP)-model of Parkinson’s disease (PD) [240].
7.2.3. Infectious Diseases
Viral Diseases
GRP94 or GP96 has been reported to be involved in viral infections. GRP94 interacts with and supports protein folding and export of Toll like receptors (TLRs) [241]. Other studies confirmed the role of TLRs as key sensors of innate immunity to viruses by identifying their components like nucleic acids and envelope glycoproteins [242]. Following viral infections, activated TLRs, trigger several signaling events resulting in production of type I interferons, inflammatory cytokines, and chemokines ultimately leading to eliminating an invading pathogen [243]. In addition to its role in the functionality of TLRs, GRP94 was found to be essential in Vesicular stomatitis virus infection [244]. Overexpression of GRP94 has been reported in Rota virus-infected cells [245]. Patients with Hepatitis B virus (HBV) displayed high expression levels of GRP94 which has been associated with poor prognosis [246,247]. GRP94 appeared to be a key regulator in the stabilization and regulation of HBV RNA polymerase [248,249]. Studies with Japanese encephalitis virus revealed that unlike HSP90 α which was retained inside the nucleus, GRP94 was released into the secretion medium upon JEV infection [250].
Parasitic Diseases
A single copy of GRP94 has been characterized in Leshmania major where it has been linked to its virulence [208]. In addition, three genes of GRP94 has been found in the Chagas’ disease causing parasite, Trypanosoma cruzi. Notably, the HSP90 inhibitor geldanamycine hindered the parasite development and seemed to interfere with proteins involved in the epimastigote differentiation [15,208]. Importantly, the secreted forms of GRP94 which chaperones plasmodium falciparum (Pf) AMA1 and PfCSP sporozoite proteins have been recently exploited in developing a novel vaccine against malaria [251].
7.2.4. Other Diseases
The involvement of GRP94 in several diseases comes from its important role in the ER quality control through folding of newly synthesized proteins and proteostasis [49,161]. For instance, mutation-causing diseases or diseases causing ER stress, including atherosclerosis, obesity, and diabetes, upregulate GRP94 and other members of the ER chaperone network to deal with the resultant ER stress. In agreement with these facts, GRP94 has been found critical for regulation and development of pancreatic β-Cells providing new insights for preservation enhancement of β-cell function. [252].
7.3. TRAP1
7.3.1. Cancer
TRAP1 participates in the carcinogenesis of many tumors [253]; (1) it interferes with aerobic glycolysis by inhibiting succinate dehydrogenase; (2) supports cell survival through hindering ROS accumulation and counteracting toxic cancer drugs and oxidants; and (3) maintains protein homeostasis by regulation of de novo protein synthesis and protein degradation. Additionally, in cancerous tissues, contrasting reports about TRAP1 expression patterns have evolved [254]. High expression of TRAP1 has been reported in colon cancer [255,256], breast cancer [257,258], prostatic cancer [259,260], lung cancers [261], and glioblastoma [262]. On the other hand, reduced expression has been shown in particular tumors, such as ovarian [263,264], bladder, and renal cancers [265]. Interestingly, high TRAP1 expression has been linked to drug resistance while its downregulation has been associated with poor prognosis. The controversial expression data of TRAP1 in cancer suggested a potential dual role as an oncogene or onco-suppressor according to cancer type [254].
7.3.2. Neurodegenerative Diseases
In addition to TRAP1 function in keeping mitochondrial integrity under normal physiology, it has been shown to contribute in correcting against mitophagy and mitochondrial degeneration in many neurodegenerative diseases. As a substrate for Parkinson’s disease kinase (PINK1), phosphorylated TRAP1 sustains healthy mitochondria by suppressing cytochrome C release from mitochondria. Impaired phosphorylation of TRAP1, due to mutated PINK1, leads to Parkinson’s disease [68,253]. Moreover, neuronal cancer cell lines that lacked TRAP1 such as neuroblastoma and glioma showed irregular mitochondrial morphology with diminished levels of dynamin-related protein 1 (Drp1) and mitochondrial fission factor (Mff) [266]. In fact, Drosophila has been extensively used as a model for PD. Interestingly, human TRAP1 could rescue PINK1 loss-of-function phenotypes in Drosophila and act against mitochondrial fragmentation and dysfunction in human neuronal SH-SY5Y cells [267]. In other studies, Drosophila TRAP1 null mutants revealed impaired mitochondrial function and hypersensitivity to stress suggesting a down-stream role of TRAP1 following PINK1 in mitochondrial metabolic pathways [268]. In vitro experiments revealed that A53T mutated α-Synuclein disrupts mitochondrial functionality in terms of insufficient ATP synthesis, mitochondrial fragmentation and elevated oxidative stress. However, overexpression of human TRAP1 was able to relieve these deleterious effects [269]. Recent studies revealed that TRAP1 is a novel interactor of protease HTRA2 and mutations in both proteins cause PD [270]. The TRAP1 mutation R47X caused increased mitochondrial respiration and ATP production [271]. These consequences were similar to TRAP1 silencing in ovarian cancer which led to resistance to chemotherapy due to increased oxidative phosphorylation [264]. Interestingly, the diabetes drug Metformin could alleviate the TRAP1 mutation associated disturbances of energy metabolism in PD through mild inhibition of mitochondrial respiration [271].
7.3.3. Kidney Diseases
TRAP1 have been reported to be involved in renal diseases. Overexpression of TRAP1 in glomerulonephritis was accompanied by DNaseI shutdown, which happens in end-stage disease, suggesting transcriptional interference between TRAP1 and DNaseI genes [272]. Moreover, using deep sequencing, revealed common recessive mutations of TRAP1 among patients with congenital anomalies of the kidney and urinary tract (CAKUT) [253,273].
8. Conclusions and Perspectives
The 90 kDa HSPs constitute a class of highly conserved proteins implicated in a plethora of physiological, as well as pathological, processes. Due to their abundance and distinct cellular distribution, the function of HSP90 members attracted researchers to extensively explore their complexity. Although we present here the most significant knowledge about HSP90 family members, it only represents a fraction of the HSP90 literature. In this review we shed light on the major diverse functions of HSP90 family members in terms of structure, regulation, and function in health and disease conditions. The use of HSP90 inhibitors is of critical clinical importance especially in cancer and neurodegenerative diseases. However, the challenge remains to identify and establish the most effective and least toxic drug.
Author Contributions
A.H., M.E.E.-S., and H.Y.N. conceived and wrote the manuscript.
Funding
We would like to thank the German Academic Exchange Service (DAAD), Bonn, Germany for supporting A.H. This work has been supported by intramural funds from the University of Veterinary Medicine Hannover, Germany and the American University of Beirut, Lebanon. This publication was supported by Deutsche Forschungsgemeinschaft and University of Veterinary Medicine Hannover, Foundation within the funding programme Open Access Publishing.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| Aha1 | Activator of Hsp90 ATPase protein 1 |
| Apaf-1 | Apoptotic protease-activating factor 1 |
| ARNT | Aryl hydrocarbon receptor nuclear translocator |
| ARRB1 | Beta-arrestin-1 |
| Cdc37 | Hsp90 co-chaperone Cdc37, cell division cycle 37 |
| CDK4 | Cyclin-dependent kinase 4 |
| CHIP | Carboxyl terminus of Hsp70-interacting protein |
| CLAVATA | Receptor-like protein CLAVATA |
| c-MET | Proto-oncogene c-Met, hepatocyte growth factor receptor |
| CYP-D | Cyclophilin or Peptidyl-prolyl cis-trans isomerase F, mitochondrial |
| ERBB2 | Receptor tyrosine-protein kinase erbB-2 |
| FANCA | Fanconi anemia group A protein |
| FKBP51 and FKBP52 | Peptidyl-prolyl cis-trans isomerase or 51 kDa FK506-binding protein |
| Flt-3 | Receptor-type tyrosine-protein kinase FLT3 |
| HIF1α | Hypoxia-inducible factor 1-alpha |
| HIP | Hsc70-interacting protein |
| HMG | High mobility group protein |
| HOP | Co-chaperone Hsp70/Hsp90-organizing protein |
| HSP | Heat shock protein |
| IRAK | Interleukin-1 receptor-associated kinase |
| MAFG | Transcription factor MafG |
| MHC class II | Major Histocompatibility Complex Class II |
| MMP2 | Matrix metalloproteinase-2 |
| Myt1 | Membrane-associated tyrosine- and threonine-specific cdc2-inhibitory kinase |
| NEK8 | Serine/threonine-protein kinase Nek8, Never in mitosis A-related kinase 8 |
| NF-κB | Nuclear factor NF-kappa |
| p23 | Co-chaperone protein 23 |
| P53 | Cellular tumor antigen p53 |
| PI3K | Phosphatidylinositol 3-kinase |
| Plk | Serine/threonine-protein kinase PLK1 |
| PP5 | Serine/threonine-protein phosphatase 5 |
| RTK | Receptor Tyrosine Kinase |
| Sgt1 | Protein SGT1 homolog, Suppressor of G2 allele of SKP1 homolog |
| SREBF | Sterol regulatory element-binding protein |
| SSDF-1 | Stromal cell derived factor -1 |
| TLR | Toll-like receptor |
| TNF | Tumor Necrosis Factor |
| Tom70 | Mitochondrial import receptor subunit TOM70 |
| VEGFR | Vascular endothelial growth factor receptor |
| Wee1 | Wee1-like protein kinase |
| WISp39 | Waf1 Cip1 stabilizing protein 39 |
References
- Ritossa, F. A new puffing pattern induced by temperature shock and DNP in drosophila. Experientia 1962, 18, 571–573. [Google Scholar] [CrossRef]
- Malyshev, I. A General Description of HSPs, The Molecular Structure of HSP70 and The HSP70 Cycle. In Immunity, Tumors and Aging: The Role of HSP70; Springer: Dordrecht, The Netherlands, 2013; pp. 1–13. ISBN 978-94-007-5943-5. [Google Scholar]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef] [PubMed]
- Pirkkala, L.; Sistonen, L. Heat Shock Proteins (HSPs): Structure, Function and Genetics. Encycl. Life Sci. 2006, 1–7. [Google Scholar] [CrossRef]
- Kampinga, H.H.; Hageman, J.; Vos, M.J.; Kubota, H.; Tanguay, R.M.; Bruford, E.A.; Cheetham, M.E.; Chen, B.; Hightower, L.E. Guidelines for the nomenclature of the human heat shock proteins. Cell Stress Chaperones 2009, 14, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhong, D.; Monteiro, A. Comparative genomics and evolution of the HSP90 family of genes across all kingdoms of organisms. BMC Genom. 2006, 7, 156. [Google Scholar] [CrossRef]
- Whitesell, L.; Lindquist, S.L. HSP90 and the chaperoning of cancer. Nat. Rev. Cancer 2005, 5, 761–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Sun, W.; Taldone, T.; Rodina, A.; Chiosis, G. Heat shock protein 90 in neurodegenerative diseases. Mol. Neurodegener. 2010, 5, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Geller, R.; Taguwa, S.; Frydman, J. Broad action of Hsp90 as a host chaperone required for viral replication. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Buchner, J. Structure, function and regulation of the hsp90 machinery. Biomed. J. 2012, 36, 106–117. [Google Scholar] [CrossRef]
- Langer, T.; Rosmus, S.; Fasold, H. Intracellular localization of the 90 kDA heat shock protein (HSP90α) determined by expression of a EGFP-HSP90α-fusion protein in unstressed and heat stressed 3T3 cells. Cell Biol. Int. 2003, 27, 47–52. [Google Scholar] [CrossRef]
- Prodromou, C. Mechanisms of Hsp90 regulation. Biochem. J. 2016, 473, 2439–2452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Soroka, J.; Buchner, J. The Hsp90 chaperone machinery: Conformational dynamics and regulation by co-chaperones. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Didenko, T.; Duarte, A.M.S.; Karagöz, G.E.; Rüdiger, S.G.D. Hsp90 structure and function studied by NMR spectroscopy. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Jackson, S.E. Hsp90: Structure and function. Top. Curr. Chem. 2013, 328, 155–240. [Google Scholar] [CrossRef] [PubMed]
- Lavery, L.A.; Partridge, J.R.; Ramelot, T.A.; Elnatan, D.; Kennedy, M.A.; Agard, D.A. Structural Asymmetry in the Closed State of Mitochondrial Hsp90 (TRAP1) Supports a Two-Step ATP Hydrolysis Mechanism. Mol. Cell 2014, 53, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Sreedhar, A.S.; Kalmár, É.; Csermely, P.; Shen, Y.F. Hsp90 isoforms: Functions, expression and clinical importance. FEBS Lett. 2004, 562, 11–15. [Google Scholar] [CrossRef]
- Csermely, P.; Schnaider, T.; Soti, C.; Prohászka, Z.; Nardai, G. The 90-kDa Molecular Chaperone Family: Structure, Function, and Clinical Applications. A Comprehensive Review. Pharmacol. Ther. 1998, 79, 129–168. [Google Scholar] [CrossRef]
- Tsutsumi, S.; Mollapour, M.; Prodromou, C.; Lee, C.-T.; Panaretou, B.; Yoshida, S.; Mayer, M.P.; Neckers, L.M. Charged linker sequence modulates eukaryotic heat shock protein 90 (Hsp90) chaperone activity. Proc. Natl. Acad. Sci. USA 2012, 109, 2937–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.S. Phylogenetic analysis of the 90 kD heat shock family of protein sequences and an examination of the relationship among animals, plants, and fungi species. Mol. Biol. Evol. 1995, 12, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.L. Evolution and function of diverse Hsp90 homologs and cochaperone proteins. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Ciocca, D.R.; Arrigo, A.P.; Calderwood, S.K. Heat shock proteins and heat shock factor 1 in carcinogenesis and tumor development: An update. Arch. Toxicol. 2013, 87, 19–48. [Google Scholar] [CrossRef] [PubMed]
- Welch, W.J.; Feramisco, J.R. Purification of the major mammalian heat shock proteins. J. Biol. Chem. 1982, 257, 14949–14959. [Google Scholar] [PubMed]
- Rose, D.W.; Wettenhall, R.E.H.; Kudlicki, W.; Kramer, G.; Hardesty, B. The 90-kilodalton peptide of the heme-regulated eIF-2 alpha kinase has sequence similarity with the 90-kilodalton heat shock protein. Biochemistry 1987, 26, 6583–6587. [Google Scholar] [CrossRef] [PubMed]
- Koyasu, S.; Nishida, E.; Kadowaki, T.; Matsuzaki, F.; Iida, K.; Harada, F.; Kasuga, M.; Sakai, H.; Yahara, I. Two mammalian heat shock proteins, HSP90 and HSP100, are actin-binding proteins. Proc. Natl. Acad. Sci. USA 1986, 83, 8054–8058. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, M.; Takahashi, Y.; Inano, K.; Horigome, T.; Sugano, H. Characterization of the hydrophobic region of heat shock protein 90. J. Biochem. 1991, 110, 141–145. [Google Scholar] [CrossRef] [PubMed]
- Nardai, G.; Schnaider, T.; Söti, C.; Ryan, M.T.; Hoj, P.B.; Somogyi, J.; Csermely, P. Characterization of the 90 kDa heat shock protein (HSP90)-associated ATP/GTPase. J. Biosci. 1996, 21, 179–190. [Google Scholar] [CrossRef]
- Prodromou, C.; Roe, S.M.; Piper, P.W.; Pearl, L.H. A molecular clamp in the crystal structure of the N-terminal domain of the yeast Hsp90 chaperone. Nat. Struct. Biol. 1997, 4, 477–482. [Google Scholar] [CrossRef] [PubMed]
- Dutta, R.; Inouye, M. GHKL, an emergent ATPase/kinase superfamily. Trends Biochem. Sci. 2000, 25, 24–28. [Google Scholar] [CrossRef]
- Panaretou, B.; Prodromou, C.; Roe, S.M.; O’Brien, R.; Ladbury, J.E.; Piper, P.W.; Pearl, L.H. ATP binding and hydrolysis are essential to the function of the Hsp90 molecular chaperone in vivo. EMBO J. 1998, 17, 4829–4836. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, S.; Mollapour, M.; Graf, C.; Lee, C.T.; Scroggins, B.T.; Xu, W.; Haslerova, L.; Hessling, M.; Konstantinova, A.A.; Trepel, J.B.; et al. Hsp90 charged-linker truncation reverses the functional consequences of weakened hydrophobic contacts in the N domain. Nat. Struct. Mol. Biol. 2009, 16, 1141–1147. [Google Scholar] [CrossRef] [PubMed]
- Shiau, A.K.; Harris, S.F.; Southworth, D.R.; Agard, D.A. Structural Analysis of E. coli hsp90 Reveals Dramatic Nucleotide-Dependent Conformational Rearrangements. Cell 2006, 127, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Scheibel, T.; Siegmund, H.I.; Jaenicke, R.; Ganz, P.; Lilie, H.; Buchner, J. The charged region of Hsp90 modulates the function of the N-terminal domain. Proc. Natl. Acad. Sci. USA 1999, 96, 1297–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louvion, J.F.; Warth, R.; Picard, D. Two eukaryote-specific regions of Hsp82 are dispensable for its viability and signal transduction functions in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 13937–13942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyer, P.; Prodromou, C.; Hu, B.; Vaughan, C.; Roe, S.M.; Panaretou, B.; Piper, P.W.; Pearl, L.H. Structural and functional analysis of the middle segment of hsp90: Implications for ATP hydrolysis and client protein and cochaperone interactions. Mol. Cell 2003, 11, 647–658. [Google Scholar] [CrossRef]
- Huai, Q.; Wang, H.; Liu, Y.; Kim, H.Y.; Toft, D.; Ke, H. Structures of the N-terminal and middle domains of E. coli Hsp90 and conformation changes upon ADP binding. Structure 2005, 13, 579–590. [Google Scholar] [CrossRef] [PubMed]
- Minami, Y.; Kawasaki, H.; Suzuki, K.; Yahara, I. The calmodulin-binding domain of the mouse 90-kDa heat shock protein. J. Biol. Chem. 1993, 268, 9604–9610. [Google Scholar] [PubMed]
- Toft, D.; Meng, X.; Devin, J.; Sullivan, W.; Baulieu, E.; Catelli, M. Mutational analysis of Hsp90 alpha dimerization and subcellular localization: Dimer disruption does not impede “in vivo’’ interaction with estrogen receptor. J. Cell Sci. 1996, 109, 1677–1687. [Google Scholar]
- Soti, C.; Vermes, Á.; Haystead, T.A.J.; Csermely, P. Comparative analysis of the ATP-binding sites of Hsp90 by nucleotide affinity cleavage: A distinct nucleotide specificity of the C-terminal ATP-binding site. Eur. J. Biochem. 2003, 270, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Marcu, M.G.; Schulte, T.W.; Neckers, L. Novobiocin and Related Coumarins and Depletion of Heat Shock Protein 90-Dependent Signaling Proteins. JNCI J. Natl. Cancer Inst. 2000, 92, 242–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Z.; Henry, E.C.; Gasiewicz, T.A. (−)-Epigallocatechin-3-gallate is a novel Hsp90 inhibitor. Biochemistry 2009, 48, 336–345. [Google Scholar] [CrossRef] [PubMed]
- Young, J.C.; Schneider, C.; Hartl, F.U. In vitro evidence that hsp90 contains two independent chaperone sites. FEBS Lett. 1997, 418, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, W.P.; Toft, D.O. Mutational analysis of hsp90 binding to the progesterone receptor. J. Biol. Chem. 1993, 268, 20373–20379. [Google Scholar] [PubMed]
- Garg, G.; Khandelwal, A.; Blagg, B.S.J. Chapter Three—Anticancer Inhibitors of Hsp90 Function: Beyond the Usual Suspects. In Hsp90 in Cancer: Beyond the Usual Suspects; Isaacs, J., Whitesell, L., Eds.; Advances in Cancer Research; Academic Press: Cambridge, MA, USA, 2016; Volume 129, pp. 51–88. [Google Scholar]
- Yang, Y.; Li, Z. Roles of heat shock protein gp96 in the ER quality control: Redundant or unique function? Mol. Cells 2005, 20, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Koch, G.; Smith, M.; Macer, D.; Webster, P.; Mortara, R. Endoplasmic reticulum contains a common, abundant calcium-binding glycoprotein, endoplasmin. J. Cell Sci. 1986, 86, 217–232. [Google Scholar] [PubMed]
- Hoter, A.; Amiri, M.; Warda, M.; Naim, H.Y. Molecular cloning, cellular expression and characterization of Arabian camel (Camelus dromedarius) endoplasmin. Int. J. Biol. Macromol. 2018, 117, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Soldano, K.L.; Jivan, A.; Nicchitta, C.V.; Gewirth, D.T. Structure of the N-terminal domain of GRP94. Basis for ligand specificity and regulation. J. Biol. Chem. 2003, 278, 48330–48338. [Google Scholar] [CrossRef] [PubMed]
- Marzec, M.; Eletto, D.; Argon, Y. GRP94: An HSP90-like protein specialized for protein folding and quality control in the endoplasmic reticulum. Biochim. Biophys. Acta 2012, 1823, 774–787. [Google Scholar] [CrossRef] [PubMed]
- Immormino, R.M.; Dollins, D.E.; Shaffer, P.L.; Soldano, K.L.; Walker, M.A.; Gewirth, D.T. Ligand-induced conformational shift in the N-terminal domain of GRP94, an Hsp90 chaperone. J. Biol. Chem. 2004, 279, 46162–46171. [Google Scholar] [CrossRef] [PubMed]
- Dollins, D.E.; Immormino, R.M.; Gewirth, D.T. Structure of unliganded GRP94, the endoplasmic reticulum Hsp90: Basis for nucleotide-induced conformational change. J. Biol. Chem. 2005, 280, 30438–30447. [Google Scholar] [CrossRef] [PubMed]
- Hainzl, O.; Lapina, M.C.; Buchner, J.; Richter, K. The charged linker region is an important regulator of Hsp90 function. J. Biol. Chem. 2009, 284, 22559–22567. [Google Scholar] [CrossRef] [PubMed]
- Biswas, C.; Ostrovsky, O.; Makarewich, C.A.; Wanderling, S.; Gidalevitz, T.; Argon, Y. The peptide-binding activity of GRP94 is regulated by calcium. Biochem. J. 2007, 405, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Altieri, D.C. Hsp90 regulation of mitochondrial protein folding: From organelle integrity to cellular homeostasis. Cell. Mol. Life Sci. 2013, 70, 2463–2472. [Google Scholar] [CrossRef] [PubMed]
- Altieri, D.C.; Stein, G.S.; Lian, J.B.; Languino, L.R. TRAP-1, the mitochondrial Hsp90. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 767–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.Y.; Dunbar, J.D.; Zhang, Y.X.; Guo, D.; Donner, D.B. Identification of a protein with homology to hsp90 that binds the type 1 tumor necrosis factor receptor. J. Biol. Chem. 1995, 270, 3574–3581. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Piel, W.H.; Gui, L.; Bruford, E.; Monteiro, A. The HSP90 family of genes in the human genome: Insights into their divergence and evolution. Genomics 2005, 86, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Leskovar, A.; Wegele, H.; Werbeck, N.D.; Buchner, J.; Reinstein, J. The ATPase cycle of the mitochondrial Hsp90 analog trap1. J. Biol. Chem. 2008, 283, 11677–11688. [Google Scholar] [CrossRef] [PubMed]
- Elnatan, D.; Agard, D.A. Modulation of mitochondrial Hsp90 (TRAP1) ATPase activity by calcium and magnesium. bioRxiv 2018, 90. [Google Scholar] [CrossRef]
- Kang, B.H. TRAP1 regulation of mitochondrial life or death decision in cancer cells and mitochondria-targeted TRAP1 inhibitors. BMB Rep. 2012, 45, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masgras, I.; Sanchez-Martin, C.; Colombo, G.; Rasola, A. The Chaperone TRAP1 As a Modulator of the Mitochondrial Adaptations in Cancer Cells. Front. Oncol. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Felts, S.J.; Owen, B.A.; Nguyen, P.; Trepel, J.; Donner, D.B.; Toft, D.O. The hsp90-related protein TRAP1 is a mitochondrial protein with distinct functional properties. J. Biol. Chem. 2000, 275, 3305–3312. [Google Scholar] [CrossRef] [PubMed]
- Schopf, F.H.; Biebl, M.M.; Buchner, J. The HSP90 chaperone machinery. Nat. Rev. Mol. Cell Biol. 2017, 18, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Voellmy, R.; Boellmann, F. Chaperone regulation of the heat shock protein response. Adv. Exp. Med. Biol. 2007, 594, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Hickey, E.; Brandon, S.E.; Smale, G.; Lloyd, D.; Weber, L.A. Sequence and regulation of a gene encoding a human 89-kilodalton heat shock protein. Mol. Cell. Biol. 1989, 9, 2615–2626. [Google Scholar] [CrossRef] [PubMed]
- Mollapour, M.; Neckers, L. Post-translational modifications of Hsp90 and their contributions to chaperone regulation. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, A.D.; Beebe, K.; Neckers, L.; Prince, T. Regulation and function of the human HSP90AA1 gene. Gene 2015, 570, 8–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pridgeon, J.W.; Olzmann, J.A.; Chin, L.S.; Li, L. PINK1 protects against oxidative stress by phosphorylating mitochondrial chaperone TRAP1. PLoS Biol. 2007, 5, 1494–1503. [Google Scholar] [CrossRef] [PubMed]
- Zuehlke, A.; Johnson, J.L. Hsp90 and co-chaperones twist the functions of diverse client proteins. Biopolymers 2010, 93, 211–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasola, A.; Neckers, L.; Picard, D. Mitochondrial oxidative phosphorylation TRAP(1)ped in tumor cells. Trends Cell Biol. 2014, 24, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Rosenbaum, M.; Andreani, V.; Kapoor, T.; Herp, S.; Flach, H.; Duchniewicz, M.; Grosschedl, R. MZB1 is a GRP94 cochaperone that enables proper immunoglobulin heavy chain biosynthesis upon ER stress. Genes Dev. 2014, 28, 1165–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaiff, W.T.; Hruska, K.A.; McCourt, D.W.; Green, M.; Schwartz, B.D. HLA-DR associates with specific stress proteins and is retained in the endoplasmic reticulum in invariant chain negative cells. J. Exp. Med. 1992, 176, 657–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavany, C.; Mimnaugh, E.; Miller, P.; Bitton, R.; Nguyen, P.; Trepel, J.; Whitesell, L.; Schnur, R.; Moyer, J.; Neckers, L. p185erbB2 binds to GRP94 in vivo. Dissociation of the p185erbB2/GRP94 heterocomplex by benzoquinone ansamycins precedes depletion of p185erbB2. J. Biol. Chem. 1996, 271, 4974–4977. [Google Scholar] [PubMed]
- Chen, C.F.; Chen, Y.; Dai, K.; Chen, P.L.; Riley, D.J.; Lee, W.H. A new member of the hsp90 family of molecular chaperones interacts with the retinoblastoma protein during mitosis and after heat shock. Mol. Cell. Biol. 1996, 16, 4691–4699. [Google Scholar] [CrossRef] [PubMed]
- Ostrovsky, O.; Ahmed, N.T.; Argon, Y. The chaperone activity of GRP94 toward insulin-like growth factor II is necessary for the stress response to serum deprivation. Mol. Biol. Cell 2009, 20, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Simmons, A.D.; Musy, M.M.; Lopes, C.S.; Hwang, L.Y.; Yang, Y.P.; Lovett, M. A direct interaction between EXT proteins and glycosyltransferases is defective in hereditary multiple exostoses. Hum. Mol. Genet. 1999, 8, 2155–2164. [Google Scholar] [CrossRef] [PubMed]
- Pennell, C.A. Heat shock proteins in immune response in the Fall of 2004. Immunology 2005, 114, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Ishiguro, S.; Watanabe, Y.; Ito, N.; Nonaka, H.; Takeda, N.; Sakai, T.; Kanaya, H.; Okada, K. SHEPHERD is the Arabidopsis GRP94 responsible for the formation of functional CLAVATA proteins. EMBO J. 2002, 21, 898–908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landriscina, M.; Laudiero, G.; Maddalena, F.; Amoroso, M.R.; Piscazzi, A.; Cozzolino, F.; Monti, M.; Garbi, C.; Fersini, A.; Pucci, P.; et al. Mitochondrial chaperone trap1 and the calcium binding protein sorcin interact and protect cells against apoptosis induced by antiblastic agents. Cancer Res. 2010, 70, 6577–6586. [Google Scholar] [CrossRef] [PubMed]
- Nganga, A.; Bruneau, N.; Sbarra, V.; Lombardo, D.; Le Petit-Thevenin, J. Control of pancreatic bile-salt-dependent-lipase secretion by the glucose-regulated protein of 94 kDa (Grp94). Biochem. J. 2000, 352, 865–874. [Google Scholar] [PubMed]
- Amoroso, M.R.; Matassa, D.S.; Laudiero, G.; Egorova, A.V.; Polishchuk, R.S.; Maddalena, F.; Piscazzi, A.; Paladino, S.; Sarnataro, D.; Garbi, C.; et al. TRAP1 and the proteasome regulatory particle TBP7/Rpt3 interact in the endoplasmic reticulum and control cellular ubiquitination of specific mitochondrial proteins. Cell Death Differ. 2012, 19, 592–602. [Google Scholar] [CrossRef] [PubMed]
- Brunati, A.M.; Contri, A.; Muenchbach, M.; James, P.; Marin, O.; Pinna, L.A. GRP94 (endoplasmin) co-purifies with and is phosphorylated by Golgi apparatus casein kinase. FEBS Lett. 2000, 471, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Hecht, J.T.; Hayes, E.; Snuggs, M.; Decker, G.; Montufar-Solis, D.; Doege, K.; Mwalle, F.; Poole, R.; Stevens, J.; Duke, P.J. Calreticulin, PDI, Grp94 and BiP chaperone proteins are associated with retained COMP in pseudoachondroplasia chondrocytes. Matrix Biol. 2001, 20, 251–262. [Google Scholar] [CrossRef]
- Schmidt, B.Z.; Perlmutter, D.H. Grp78, Grp94, and Grp170 interact with alpha1-antitrypsin mutants that are retained in the endoplasmic reticulum. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G444–G455. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, G.; Chen, L.B.; Nigam, S.K. Several endoplasmic reticulum stress proteins, including ERp72, interact with thyroglobulin during its maturation. J. Biol. Chem. 1994, 269, 22990–22995. [Google Scholar] [PubMed]
- Ramakrishnan, M.; Tugizov, S.; Pereira, L.; Lee, A.S. Conformation-defective herpes simplex virus 1 glycoprotein B activates the promoter of the grp94 gene that codes for the 94-kD stress protein in the endoplasmic reticulum. DNA Cell Biol. 1995, 14, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Linnik, K.M.; Herscovitz, H. Multiple molecular chaperones interact with apolipoprotein B during its maturation. The network of endoplasmic reticulum-resident chaperones (ERp72, GRP94, calreticulin, and BiP) interacts with apolipoprotein b regardless of its lipidation state. J. Biol. Chem. 1998, 273, 21368–21373. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.R.; Norris, K.; Smith, T.; Hebert, C.; Sauk, J.J. Association of Hsp47, Grp78, and Grp94 with procollagen supports the successive or coupled action of molecular chaperones. J. Cell. Biochem. 1994, 56, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Katsumi, A.; Senda, T.; Yamashita, Y.; Yamazaki, T.; Hamaguchi, M.; Kojima, T.; Kobayashi, S.; Saito, H. Protein C Nagoya, an elongated mutant of protein C, is retained within the endoplasmic reticulum and is associated with GRP78 and GRP94. Blood 1996, 87, 4164–4175. [Google Scholar] [PubMed]
- Randow, F.; Seed, B. Endoplasmic reticulum chaperone gp96 is required for innate immunity but not cell viability. Nat. Cell Biol. 2001, 3, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Melnick, J.; Aviel, S.; Argon, Y. The endoplasmic reticulum stress protein GRP94, in addition to BiP, associates with unassembled immunoglobulin chains. J. Biol. Chem. 1992, 267, 21303–21306. [Google Scholar] [PubMed]
- Foy, S.P.; Matsuuchi, L. Association of B lymphocyte antigen receptor polypeptides with multiple chaperone proteins. Immunol. Lett. 2001, 78, 149–160. [Google Scholar] [CrossRef]
- Kuznetsov, G.; Chen, L.B.; Nigam, S.K. Multiple molecular chaperones complex with misfolded large oligomeric glycoproteins in the endoplasmic reticulum. J. Biol. Chem. 1997, 272, 3057–3063. [Google Scholar] [CrossRef] [PubMed]
- Rowlands, M.; McAndrew, C.; Prodromou, C.; Pearl, L.; Kalusa, A.; Jones, K.; Workman, P.; Aherne, W. Detection of the ATPase Activity of the Molecular Chaperones Hsp90 and Hsp72 Using the TranscreenerTM ADP Assay Kit. J. Biomol. Screen. 2010, 15, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wandinger, S.K.; Richter, K.; Buchner, J. The Hsp90 chaperone machinery. J. Biol. Chem. 2008, 283, 18473–18477. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.M.U.; Mark Roe, S.; Vaughan, C.K.; Meyer, P.; Panaretou, B.; Piper, P.W.; Prodromou, C.; Pearl, L.H. Crystal structure of an Hsp90-nucleotide-p23/Sba1 closed chaperone complex. Nature 2006, 440, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- Scheibel, T.; Weikl, T.; Buchner, J. Two chaperone sites in Hsp90 differing in substrate specificity and ATP dependence. Proc. Natl. Acad. Sci. USA 1998, 95, 1495–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roe, S.M.; Prodromou, C.; O’Brien, R.; Ladbury, J.E.; Piper, P.W.; Pearl, L.H. Structural basis for inhibition of the Hsp90 molecular chaperone by the antitumor antibiotics radicicol and geldanamycin. J. Med. Chem. 1999, 42, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Sharp, S.; Workman, P. Inhibitors of the HSP90 Molecular Chaperone: Current Status. Adv. Cancer Res. 2006, 95, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Blagg, B.S.J.; Kerr, T.D. Hsp90 inhibitors: Small molecules that transform the Hsp90 protein folding machinery into a catalyst for protein degradation. Med. Res. Rev. 2006, 26, 310–338. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, A.; Blagg, B. Novobiocin and Additional Inhibitors of the Hsp90 C-Terminal Nucleotide-binding Pocket. Curr. Med. Chem. 2008, 15, 2702–2717. [Google Scholar] [CrossRef] [PubMed]
- Walter, S.; Buchner, J. Molecular Chaperones—Cellular Machines for Protein Folding. Angew. Chem. Int. Ed. 2002, 41, 1098–1113. [Google Scholar] [CrossRef]
- Chaudhury, S.; Welch, T.R.; Blagg, B.S.J. Hsp90 as a Target for Drug Development. ChemMedChem 2006, 1, 1331–1340. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.J.M.; Kanelakis, K.C.; Galigniana, M.D.; Morishima, Y.; Pratt, W.B. Stoichiometry, Abundance, and Functional Significance of the hsp90/hsp70-based Multiprotein Chaperone Machinery in Reticulocyte Lysate. J. Biol. Chem. 2001, 276, 30092–30098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caplan, A.J.; Mandal, A.K.; Theodoraki, M.A. Molecular chaperones and protein kinase quality control. Trends Cell Biol. 2007, 17, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Kosano, H.; Stensgard, B.; Charlesworth, M.C.; McMahon, N.; Toft, D. The assembly of progesterone receptor-hsp90 complexes using purified proteins. J. Biol. Chem. 1998, 273, 32973–32979. [Google Scholar] [CrossRef] [PubMed]
- Prodromou, C.; Panaretou, B.; Chohan, S.; Siligardi, G.; O’Brien, R.; Ladbury, J.E.; Roe, S.M.; Piper, P.W.; Pearl, L.H. The ATPase cycle of Hsp90 drives a molecular “clamp” via transient dimerization of the N-terminal domains. EMBO J. 2000, 19, 4383–4392. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Yahara, I. The 90-kDa heat shock protein, HSP90, binds and protects casein kinase II from self-aggregation and enhances its kinase activity. J. Biol. Chem. 1992, 267, 7042–7047. [Google Scholar] [PubMed]
- Daturpalli, S.; Waudby, C.A.; Meehan, S.; Jackson, S.E. Hsp90 inhibits α-synuclein aggregation by interacting with soluble oligomers. J. Mol. Biol. 2013, 425, 4614–4628. [Google Scholar] [CrossRef] [PubMed]
- Evans, C.G.; Wisén, S.; Gestwicki, J.E. Heat shock proteins 70 and 90 inhibit early stages of amyloid β-(1-42) aggregation in vitro. J. Biol. Chem. 2006, 281, 33182–33191. [Google Scholar] [CrossRef] [PubMed]
- Jakob, U.; Lilie, H.; Meyer, I.; Buchner, J. Transient interaction of Hsp90 with early unfolding intermediates of citrate synthase: Implications for heat shock in vivo. J. Biol. Chem. 1995, 270, 7288–7294. [Google Scholar] [CrossRef] [PubMed]
- Wiech, H.; Buchner, J.; Zimmermann, R.; Jakob, U. Hsp90 chaperones protein folding in vitro. Nature 1992, 358, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Yahara, I. Interaction between Casein Kinase II and the 90-kDa Stress Protein, HSP90. Biochemistry 1995, 34, 8123–8129. [Google Scholar] [CrossRef] [PubMed]
- Yonehara, M.; Minami, Y.; Kawata, Y.; Nagai, J.; Yahara, I. Heat-induced chaperone activity of HSP90. J. Biol. Chem. 1996, 271, 2641–2645. [Google Scholar] [CrossRef] [PubMed]
- Jakob, U.; Buchner, J. Assisting spontaneity: The role of Hsp90 and small Hsps as molecular chaperones. Trends Biochem. Sci. 1994, 19, 205–211. [Google Scholar] [CrossRef]
- Buchner, J. Supervising the fold: Functional principles of molecular chaperones. FASEB J. 1996, 10, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Kazlauskas, A.; Sundström, S.; Poellinger, L.; Pongratz, I. The hsp90 chaperone complex regulates intracellular localization of the dioxin receptor. Mol. Cell. Biol. 2001, 21, 2594–2607. [Google Scholar] [CrossRef] [PubMed]
- Holley, S.J.; Yamamoto, K.R. A role for Hsp90 in retinoid receptor signal transduction. Mol. Biol. Cell 1995, 6, 1833–1842. [Google Scholar] [CrossRef] [PubMed]
- Boczek, E.E.; Reefschläger, L.G.; Dehling, M.; Struller, T.J.; Häusler, E.; Seidl, A.; Kaila, V.R.I.; Buchner, J. Conformational processing of oncogenic v-Src kinase by the molecular chaperone Hsp90. Proc. Natl. Acad. Sci. USA 2015, 112, E3189–E3198. [Google Scholar] [CrossRef] [PubMed]
- Brugge, J.S.; Erikson, E.; Erikson, R.L. The specific interaction of the Rous sarcoma virus transforming protein, pp60src, with two cellular proteins. Cell 1981, 25, 363–372. [Google Scholar] [CrossRef]
- Schulte, T.W.; Blagosklonny, M.V.; Ingui, C.; Neckers, L. Disruption of the Raf-1-Hsp90 molecular complex results in destabilization of Raf-1 and loss of Raf-1-Ras association. J. Biol. Chem. 1995, 270, 24585–24588. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Fujita, N.; Tsuruo, T. Modulation of Akt kinase activity by binding to Hsp90. Proc. Natl. Acad. Sci. USA 2000, 97, 10832–10837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stancato, L.F.; Silverstein, A.M.; Owens-Grillo, J.K.; Chow, Y.H.; Jove, R.; Pratt, W.B. The hsp90-binding antibiotic geldanamycin decreases Raf levels and epidermal growth factor signaling without disrupting formation of signaling complexes or reducing the specific enzymatic activity of Raf kinase. J. Biol. Chem. 1997, 272, 4013–4020. [Google Scholar] [CrossRef] [PubMed]
- Theodoraki, M.A.; Caplan, A.J. Quality control and fate determination of Hsp90 client proteins. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Akner, G.; Mossberg, K.; Sundqvist, K.G.; Gustafsson, J.A.; Wikström, A.C. Evidence for reversible, non-microtubule and non-microfilament-dependent nuclear translocation of hsp90 after heat shock in human fibroblasts. Eur. J. Cell Biol. 1992, 58, 356–364. [Google Scholar] [PubMed]
- Fostinis, Y.; Theodoropoulos, P.A.; Gravanis, A.; Stournaras, C. Heat shock protein HSP90 and its association with the cytoskeleton: A morphological study. Biochem. Cell Biol. 1992, 70, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Suetsugu, S.; Sagara, H.; Takenawa, T. HSP90 cross-links branched actin filaments induced by N-WASP and the Arp2/3 complex. Genes Cells 2007, 12, 611–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weis, F.; Moullintraffort, L.; Heichette, C.; Chrétien, D.; Garnier, C. The 90-kDa heat shock protein Hsp90 protects tubulin against thermal denaturation. J. Biol. Chem. 2010, 285, 9525–9534. [Google Scholar] [CrossRef] [PubMed]
- Krtková, J.; Zimmermann, A.; Schwarzerová, K.; Nick, P. Hsp90 binds microtubules and is involved in the reorganization of the microtubular network in angiosperms. J. Plant Physiol. 2012, 169, 1329–1339. [Google Scholar] [CrossRef] [PubMed]
- Etard, C.; Roostalu, U.; Strähle, U. Shuttling of the chaperones Unc45b and Hsp90a between the A band and the Z line of the myofibril. J. Cell Biol. 2008, 180, 1163–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrigo, A.P.; Fakan, S.; Tissières, A. Localization of the heat shock-induced proteins in Drosophila melanogaster tissue culture cells. Dev. Biol. 1980, 78, 86–103. [Google Scholar] [CrossRef]
- Carbajal, M.E.; Valet, J.P.; Charest, P.M.; Tanguay, R.M. Purification of Drosophila hsp 83 and immunoelectron microscopic localization. Eur. J. Cell Biol. 1990, 52, 147–156. [Google Scholar] [PubMed]
- Galigniana, M.D.; Echeverría, P.C.; Erlejman, A.G.; Piwien-Pilipuk, G. Role of molecular chaperones and TPR-domain proteins in the cytoplasmic transport of steroid receptors and their passage through the nuclear pore. Nucleus 2010, 1, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambraud, B.; Berry, M.; Redeuilh, G.; Chambon, P.; Baulieu, E.E. Several regions of human estrogen receptor are involved in the formation of receptor-heat shock protein 90 complexes. J. Biol. Chem. 1990, 265, 20686–20691. [Google Scholar] [PubMed]
- Mikolajczyk, M.; Nelson, M.A. Regulation of stability of cyclin-dependent kinase CDK11p110 and a caspase-processed form, CDK11p46, by Hsp90. Biochem. J. 2004, 384, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Aligue, R.; Akhavan-Niak, H.; Russell, P. A role for Hsp90 in cell cycle control: Wee1 tyrosine kinase activity requires interaction with Hsp90. EMBO J. 1994, 13, 6099–6106. [Google Scholar] [CrossRef] [PubMed]
- Jérôme, V.; Vourc’h, C.; Baulieu, E.E.; Catelli, M.G. Cell cycle regulation of the chicken hsp90 alpha expression. Exp. Cell Res. 1993, 205, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Szent-Gyorgyi, C. A bipartite operator interacts with a heat shock element to mediate early meiotic induction of Saccharomyces cerevisiae HSP82. Mol. Cell. Biol. 1995, 15, 6754–6769. [Google Scholar] [CrossRef] [PubMed]
- Marcussen, M.; Larsen, P.J. Cell Cycle-Dependent Regulation of Cellular ATP Concentration, and Depolymerization of the Interphase Microtubular Network Induced by Elevated Cellular ATP Concentration in Whole Fibroblasts. Cell Motil. Cytoskelet. 1996, 35, 94–99. [Google Scholar] [CrossRef]
- Kohda, T.; Kondo, K.; Oishi, M. Cellular HSP90 (HSP86) mRNA level and in vitro differentiation of mouse embryonal carcinoma (F9) cells. FEBS Lett. 1991, 290, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Shakoori, A.R.; Oberdorf, A.M.; Owen, T.A.; Weber, L.A.; Hickey, E.; Stein, J.L.; Lian, J.B.; Stein, G.S. Expression of heat shock genes during differentiation of mammalian osteoblasts and promyelocytic leukemia cells. J. Cell. Biochem. 1992, 48, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Duncan, C.B.; Duncan, S.A. A small-molecule screen reveals that HSP90β promotes the conversion of induced pluripotent stem cell-derived endoderm to a hepatic fate and regulates HNF4A turnover. Development 2017, 144, 1764–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, M.T.; Csermely, P.; Sodblacti, C. Hsp90 chaperones PPARγ and regulates differentiation and survival of 3T3-L1 adipocytes. Cell Death Differ. 2013, 20, 1654–1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Echeverría, P.C.; Briand, P.-A.; Picard, D. A Remodeled Hsp90 Molecular Chaperone Ensemble with the Novel Cochaperone Aarsd1 Is Required for Muscle Differentiation. Mol. Cell. Biol. 2016, 36, 1310–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hightower, L.E.; Noonan, E.J. Discovery of the Cellular Secretion of Cell Stress Proteins. In Cellular Trafficking of Cell Stress Proteins in Health and Disease; Henderson, B., Pockley, A.G., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–11. ISBN 978-94-007-4740-1. [Google Scholar]
- Hightower, L.E.; Guidon, P.T. Selective release from cultured mammalian cells of heat-shock (stress) proteins that resemble glia-axon transfer proteins. J. Cell. Physiol. 1989, 138, 257–266. [Google Scholar] [CrossRef] [PubMed]
- Calderwood, S.K.; Gong, J.; Murshid, A. Extracellular HSPs: The complicated roles of extracellular HSPs in immunity. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sahu, D.; Tsen, F. Secreted heat shock protein-90 (Hsp90) in wound healing and cancer. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.-F.; Fan, J.; Fedesco, M.; Guan, S.; Li, Y.; Bandyopadhyay, B.; Bright, A.M.; Yerushalmi, D.; Liang, M.; Chen, M.; et al. Transforming Growth Factor (TGF)-Stimulated Secretion of HSP90: Using the Receptor LRP-1/CD91 To Promote Human Skin Cell Migration against a TGF—Rich Environment during Wound Healing. Mol. Cell. Biol. 2008, 28, 3344–3358. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, Y.; Guan, S.; Fan, J.; Cheng, C.F.; Bright, A.M.; Chinn, C.; Chen, M.; Woodley, D.T. Extracellular heat shock protein-90α: Linking hypoxia to skin cell motility and wound healing. EMBO J. 2007, 26, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Woodley, D.T.; Fan, J.; Cheng, C.-F.; Li, Y.; Chen, M.; Bu, G.; Li, W. Participation of the lipoprotein receptor LRP1 in hypoxia-HSP90 autocrine signaling to promote keratinocyte migration. J. Cell Sci. 2009, 122, 1495–1498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.; Sahu, D.; Tsen, F.; Zhao, Z.; Fan, J.; Wang, X.; Kim, R.; Kuang, Y.; Chen, M.; Woodley, D.; et al. A fragment of secreted Hsp90alpha carries unique properties to accelerate acute and chronic wound healing. J. Clin. Investig. 2011, 121. [Google Scholar] [CrossRef] [PubMed]
- Gidalevitz, T.; Stevens, F.; Argon, Y. Orchestration of secretory protein folding by ER chaperones. Biochim. Biophys. Acta 2013, 1833, 2410–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruneau, N.; Lombardo, D.; Bendayan, M. Participation of GRP94-related protein in secretion of pancreatic bile salt-dependent lipase and in its internalization by the intestinal epithelium. J. Cell Sci. 1998, 111, 2665–2679. [Google Scholar] [PubMed]
- Bruneau, N.; Lombardo, D. Chaperone function of a Grp 94-related protein for folding and transport of the pancreatic bile salt-dependent lipase. J. Biol. Chem. 1995, 270, 13524–13533. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Liu, B.; Dai, J.; Srivastava, P.K.; Zammit, D.J.; Lefrançois, L.; Li, Z. Heat Shock Protein gp96 Is a Master Chaperone for Toll-like Receptors and Is Important in the Innate Function of Macrophages. Immunity 2007, 26, 215–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, C.; Wu, S.; Yang, Y.; Hao, B.; Li, Z. Drosophila glycoprotein 93 Is an ortholog of mammalian heat shock protein gp96 (grp94, HSP90b1, HSPC4) and retains disulfide bond-independent chaperone function for TLRs and integrins. J. Immunol. 2009, 183, 5121–5128. [Google Scholar] [CrossRef] [PubMed]
- Staron, M.; Wu, S.; Hong, F.; Stojanovic, A.; Du, X.; Bona, R.; Liu, B.; Li, Z. Heat-shock protein gp96/grp94 is an essential chaperone for the platelet glycoprotein Ib-IX-V complex. Blood 2011, 117, 7136–7144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen Van, P.; Peter, F.; Soling, H.D. Four intracisternal calcium-binding glycoproteins from rat liver microsomes with high affinity for calcium. No indication for calsequestrin-like proteins in inositol 1,4,5-trisphosphate-sensitive calcium sequestering rat liver vesicles. J. Biol. Chem. 1989, 264, 17494–17501. [Google Scholar]
- Barton, E.R.; Park, S.; James, J.K.; Makarewich, C.A.; Philippou, A.; Eletto, D.; Lei, H.; Brisson, B.; Ostrovsky, O.; Li, Z.; et al. Deletion of muscle GRP94 impairs both muscle and body growth by inhibiting local IGF production. FASEB J. 2012, 26, 3691–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eletto, D.; Dersh, D.; Argon, Y. GRP94 in ER quality control and stress responses. Semin. Cell Dev. Biol. 2010, 21, 479–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiest, D.L.; Burkhardt, J.K.; Hester, S.; Hortsch, M.; Meyer, D.I.; Argon, Y. Membrane biogenesis during B cell differentiation: Most endoplasmic reticulum proteins are expressed coordinately. J. Cell Biol. 1990, 110, 1501–1511. [Google Scholar] [CrossRef] [PubMed]
- Basu, S. Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappaB pathway. Int. Immunol. 2000, 12, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Berwin, B.; Reed, R.C.; Nicchitta, C.V. Virally Induced Lytic Cell Death Elicits the Release of Immunogenic GRP94/gp96. J. Biol. Chem. 2001, 276, 21083–21088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evdokimovskaya, Y.; Skarga, Y.; Vrublevskaya, V.; Morenkov, O. Release of the glucose-regulated protein 94 by baby hamster kidney cells. Cell Biochem. Funct. 2012, 30, 558–562. [Google Scholar] [CrossRef] [PubMed]
- Spee, P.; Neefjes, J. TAP-translocated peptides specifically bind proteins in the endoplasmic reticulum, including gp96, protein disulfide isomerase and calreticulin. Eur. J. Immunol. 1997, 27, 2441–2449. [Google Scholar] [CrossRef] [PubMed]
- Lammert, E.; Arnold, D.; Nijenhuis, M.; Momburg, F.; Hämmerling, G.J.; Brunner, J.; Stevanović, S.; Rammensee, H.G.; Schild, H. The endoplasmic reticulum-resident stress protein gp96 binds peptides translocated by TAP. Eur. J. Immunol. 1997, 27, 923–927. [Google Scholar] [CrossRef] [PubMed]
- Altmeyer, A.; Maki, R.G.; Feldweg, A.M.; Heike, M.; Protopopov, V.P.; Masur, S.K.; Srivastava, P.K. Tumor-specific cell surface expression of the-KDEL containing, endoplasmic reticular heat shock protein gp96. Int. J. Cancer 1996, 69, 340–349. [Google Scholar] [CrossRef]
- Srivastava, P. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Patten, D.A.; Germain, M.; Kelly, M.A.; Slack, R.S. Reactive oxygen species: Stuck in the middle of neurodegeneration. J. Alzheimers Dis. 2010, 20. [Google Scholar] [CrossRef] [PubMed]
- Gesualdi, N.M.; Chirico, G.; Pirozzi, G.; Costantino, E.; Landriscina, M.; Esposito, F. Tumor necrosis factor-associated protein 1 (TRAP-1) protects cells from oxidative stress and apoptosis. Stress 2007, 10, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Shima, G.; Aiuchi, T.; Horie, M.; Hori, K.; Nakajo, S.; Kajimoto, S.; Shibayama-Imazu, T.; Nakaya, K. Involvement of tumor necrosis factor receptor-associated protein 1 (TRAP1) in apoptosis induced by beta-hydroxyisovalerylshikonin. J. Biol. Chem. 2004, 279, 42503–42515. [Google Scholar] [CrossRef] [PubMed]
- Hua, G.; Zhang, Q.; Fan, Z. Heat shock protein 75 (TRAP1) antagonizes reactive oxygen species generation and protects cells from granzyme M-mediated apoptosis. J. Biol. Chem. 2007, 282, 20553–20560. [Google Scholar] [CrossRef] [PubMed]
- Im, C.-N.; Lee, J.-S.; Zheng, Y.; Seo, J.-S. Iron chelation study in a normal human hepatocyte cell line suggests that tumor necrosis factor receptor-associated protein 1 (TRAP1) regulates production of reactive oxygen species. J. Cell. Biochem. 2007, 100, 474–486. [Google Scholar] [CrossRef] [PubMed]
- Siegelin, M.D.; Dohi, T.; Raskett, C.M.; Orlowski, G.M.; Powers, C.M.; Gilbert, C.A.; Ross, A.H.; Plescia, J.; Altieri, D.C. Exploiting the mitochondrial unfolded protein response for cancer therapy in mice and human cells. J. Clin. Investig. 2011, 121, 1349–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattson, M.P.; Kroemer, G. Mitochondria in cell death: Novel targets for neuroprotection and cardioprotection. Trends Mol. Med. 2003, 9, 196–205. [Google Scholar] [CrossRef]
- Kroemer, G.; Galluzzi, L.; Brenner, C. Mitochondrial Membrane Permeabilization in Cell Death. Physiol. Rev. 2007, 99–163. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, Y.; Nakagawa, T.; Shimizu, S. Mitochondrial membrane permeability transition and cell death. Biochim. Biophys. Acta 2006, 1757, 1297–1300. [Google Scholar] [CrossRef] [PubMed]
- Baines, C.P.; Kaiser, R.A.; Sheiko, T.; Craigen, W.J.; Molkentin, J.D. Voltage-dependent anion channels are dispensable for mitochondrial-dependent cell death. Nat. Cell Biol. 2007, 9, 550–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokoszka, J.E.; Waymire, K.G.; Levy, S.E.; Sligh, J.E.; Cai, J.; Jones, D.P.; MacGregor, G.R.; Douglas, C.W. The ADP/ATP translocator is not essential for the mitochondrial permeability transition pore. Nature 2004, 427, 461–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Shimizu, S.; Watanabe, T.; Yamaguchi, O.; Otsu, K.; Yamagata, H.; Inohara, H.; Kubo, T.; Tsujimoto, Y. Cyclophilin D-dependent mitochondrial permeability transition regulates some necrotic but not apoptotic cell death. Nature 2005, 434, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Basso, E.; Fante, L.; Fowlkes, J.; Petronilli, V.; Forte, M.A.; Bernardi, P. Properties of the permeability transition pore in mitochondria devoid of cyclophilin D. J. Biol. Chem. 2005, 280, 18558–18561. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.; Khan, A.; Virji, S.; Ward, J.M.; Crompton, M. Import and processing of heart mitochondrial cyclophilin D. Eur. J. Biochem. 1999, 263, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.H.; Altieri, D.C. Compartmentalized cancer drug discovery targeting mitochondrial Hsp90 chaperones. Oncogene 2009, 28, 3681–3688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, B.H.; Plescia, J.; Dohi, T.; Rosa, J.; Doxsey, S.J.; Altieri, D.C. Regulation of Tumor Cell Mitochondrial Homeostasis by an Organelle-Specific Hsp90 Chaperone Network. Cell 2007, 131, 257–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deocaris, C.C.; Kaul, S.C.; Wadhwa, R. On the brotherhood of the mitochondrial chaperones mortalin and heat shock protein 60. Cell Stress Chaperones 2006, 11, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Miyata, Y.; Nakamoto, H.; Neckers, L. The therapeutic target Hsp90 and cancer hallmarks. Curr. Pharm. Des. 2013, 19, 347–365. [Google Scholar] [CrossRef] [PubMed]
- Vartholomaiou, E.; Echeverría, P.C.; Picard, D. Chapter One—Unusual Suspects in the Twilight Zone Between the Hsp90 Interactome and Carcinogenesis. In Hsp90 in Cancer: Beyond the Usual Suspects; Isaacs, J., Whitesell, L., Eds.; Advances in Cancer Research; Academic Press: Cambridge, MA, USA, 2016; Volume 129, pp. 1–30. [Google Scholar]
- Sidera, K.; Patsavoudi, E. HSP90 inhibitors: Current development and potential in cancer therapy. Recent Pat. Anticancer Drug Discov. 2014, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, S.; Burns, T.F. Targeting heat shock proteins in cancer: A promising therapeutic approach. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Grenert, J.P.; Sullivan, W.P.; Fadden, P.; Haystead, T.A.; Clark, J.; Mimnaugh, E.; Krutzsch, H.; Ochel, H.J.; Schulte, T.W.; Sausville, E.; et al. The amino-terminal domain of heat shock protein 90 (hsp90) that binds geldanamycin is an ATP/ADP switch domain that regulates hsp90 conformation. J. Biol. Chem. 1997, 272, 23843–23850. [Google Scholar] [CrossRef] [PubMed]
- Castagnola, P.; Bellese, G.; Birocchi, F.; Gagliani, M.C.; Tacchetti, C.; Cortese, K. Identification of an HSP90 modulated multi-step process for ERBB2 degradation in breast cancer cells. Oncotarget 2016, 7, 85411–85429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, C.D.; De Mazière, A.M.; Pisacane, P.I.; van Dijk, S.M.; Eigenbrot, C.; Sliwkowski, M.X.; Klumperman, J.; Scheller, R.H. Endocytosis and sorting of ErbB2 and the site of action of cancer therapeutics trastuzumab and geldanamycin. Mol. Biol. Cell 2004, 15, 5268–5282. [Google Scholar] [CrossRef] [PubMed]
- Soga, S.; Shiotsu, Y.; Akinaga, S.; Sharma, S.V. Development of radicicol analogues. Curr. Cancer Drug Targets 2003, 3, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Schulte, T.W.; Neckers, L.M. The benzoquinone ansamycin 17-allylamino-17-demethoxygeldanamycin binds to HSP90 and shares important biologic activities with geldanamycin. Cancer Chemother. Pharmacol. 1998, 42, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Neckers, L.; Workman, P. Hsp90 molecular chaperone inhibitors: Are we there yet? Clin. Cancer Res. 2012, 18, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Woodford, M.R.; Dunn, D.; Miller, J.B.; Jamal, S.; Neckers, L.; Mollapour, M. Chapter Two—Impact of Posttranslational Modifications on the Anticancer Activity of Hsp90 Inhibitors. In Hsp90 in Cancer: Beyond the Usual Suspects; Isaacs, J., Whitesell, L., Eds.; Advances in Cancer Research; Academic Press: Cambridge, MA, USA, 2016; Volume 129, pp. 31–50. [Google Scholar]
- Dickey, C.A.; Kamal, A.; Lundgren, K.; Klosak, N.; Bailey, R.M.; Dunmore, J.; Ash, P.; Shoraka, S.; Zlatkovic, J.; Eckman, C.B.; et al. The high-affinity HSP90-CHIP complex recognizes and selectively degrades phosphorylated tau client proteins. J. Clin. Investig. 2007, 117, 648–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klucken, J.; Shin, Y.; Masliah, E.; Hyman, B.T.; McLean, P.J. Hsp70 reduces α-synuclein aggregation and toxicity. J. Biol. Chem. 2004, 279, 25497–25502. [Google Scholar] [CrossRef] [PubMed]
- McLean, P.J.; Kawamata, H.; Shariff, S.; Hewett, J.; Sharma, N.; Ueda, K.; Breakefield, X.O.; Hyman, B.T. TorsinA and heat shock proteins act as molecular chaperones: Suppression of a-synuclein aggregation. J. Neurochem. 2002, 83, 846–854. [Google Scholar] [CrossRef] [PubMed]
- McLean, P.J.; Klucken, J.; Shin, Y.; Hyman, B.T. Geldanamycin induces Hsp70 and prevents α-synuclein aggregation and toxicity in vitro. Biochem. Biophys. Res. Commun. 2004, 321, 665–669. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Anselmo, D. In vitro reconstitution of a functional duck hepatitis B virus reverse transcriptase: Posttranslational activation by Hsp90. J. Virol. 2000, 74, 11447–11455. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Toft, D.; Anselmo, D.; Wang, X. In vitro reconstitution of functional hepadnavirus reverse transcriptase with cellular chaperone proteins. J. Virol. 2002, 76, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Toft, D.O.; Seeger, C. Hepadnavirus assembly and reverse transcription require a multi-component chaperone complex which is incorporated into nucleocapsids. EMBO J. 1997, 16, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, N.; Nageshan, R.K.; Ranade, S.; Tatu, U. Heat shock protein 90 from neglected protozoan parasites. Biochim. Biophys. Acta 2012, 1823, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Shonhai, A.; Maier, A.G.; Przyborski, J.M.; Blatch, G.L. Intracellular protozoan parasites of humans: The role of molecular chaperones in development and pathogenesis. Protein Pept. Lett. 2011, 18, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Graefe, S.E.B.; Wiesgigl, M.; Gaworski, I.; Macdonald, A.; Clos, J. Inhibition of HSP90 in Trypanosoma cruzi induces a stress response but no stage differentiation. Eukaryot. Cell 2002, 1, 936–943. [Google Scholar] [CrossRef] [PubMed]
- Bentel, M.; Harder, S.; Wiesgigl, M.; Heukeshoven, J.; Gelhaus, C.; Krause, E.; Clos, J.; Bruchhaus, I. Developmentally induced changes of the proteome in the protozoan parasite Leishmania donovani. Proteomics 2003, 3, 1811–1829. [Google Scholar] [CrossRef] [PubMed]
- Sieriebriennikov, B.; Markov, G.V.; Witte, H.; Sommer, R.J. The Role of DAF-21/Hsp90 in Mouth-Form Plasticity in Pristionchus pacificus. Mol. Biol. Evol. 2017, 34, 1644–1653. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Venable, J.; LaPointe, P.; Hutt, D.M.; Koulov, A.V.; Coppinger, J.; Gurkan, C.; Kellner, W.; Matteson, J.; Plutner, H.; et al. Hsp90 Cochaperone Aha1 Downregulation Rescues Misfolding of CFTR in Cystic Fibrosis. Cell 2006, 127, 803–815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Jang, S.-W.; Park, E.; Oh, M.; Park, S.; Ko, J. The role of heat shock protein 90 in migration and proliferation of vascular smooth muscle cells in the development of atherosclerosis. J. Mol. Cell. Cardiol. 2014, 72, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Businaro, R.; Profumo, E.; Tagliani, A.; Buttari, B.; Leone, S.; D’Amati, G.; Ippoliti, F.; Leopizzi, M.; D’Arcangelo, D.; Capoano, R.; et al. Heat-shock protein 90: A novel autoantigen in human carotid atherosclerosis. Atherosclerosis 2009, 207, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Gardiner, G.; Guindon, L.; Perez, L.; Evans-Molina, C.; Blum, J. Heat shock protein 90, a potential biomarker for type I diabetes, is secreted by human pancreatic beta cells in response to cytokine stress (BA7P.161). J. Immunol. 2015, 194, 115.21. [Google Scholar]
- Lazaro, I.; Oguiza, A.; Recio, C.; Mallavia, B.; Madrigal-Matute, J.; Blanco, J.; Egido, J.; Martin-Ventura, J.L.; Gomez-Guerrero, C. Targeting HSP90 ameliorates nephropathy and atherosclerosis through suppression of NF-κB and STAT signaling pathways in diabetic mice. Diabetes 2015, 64, 3600–3613. [Google Scholar] [CrossRef] [PubMed]
- Ansa-Addo, E.A.; Thaxton, J.; Hong, F.; Wu, B.X.; Zhang, Y.; Fugle, C.W.; Metelli, A.; Riesenberg, B.; Williams, K.; Gewirth, D.T.; et al. Clients and Oncogenic Roles of Molecular Chaperone gp96/grp94. Curr. Top. Med. Chem. 2016, 16, 2765–2778. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Staron, M.; Hong, F.; Wu, B.X.; Sun, S.; Morales, C.; Crosson, C.E.; Tomlinson, S.; Kim, I.; Wu, D.; et al. Essential roles of grp94 in gut homeostasis via chaperoning canonical Wnt pathway. Proc. Natl. Acad. Sci. USA 2013, 110, 6877–6882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wu, B.X.; Metelli, A.; Thaxton, J.E.; Hong, F.; Rachidi, S.; Ansa-Addo, E.; Sun, S.; Vasu, C.; Yang, Y.; et al. GP96 is a GARP chaperone and controls regulatory T cell functions. J. Clin. Investig. 2015, 125, 859–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wanderling, S.; Simen, B.B.; Ostrovsky, O.; Ahmed, N.T.; Vogen, S.M.; Gidalevitz, T.; Argon, Y. GRP94 is essential for mesoderm induction and muscle development because it regulates insulin-like growth factor secretion. Mol. Biol. Cell 2007, 18, 3764–3775. [Google Scholar] [CrossRef] [PubMed]
- Staron, M.; Yang, Y.; Liu, B.; Li, J.; Shen, Y.; Zúñiga-Pflücker, J.C.; Aguila, H.L.; Goldschneider, I.; Li, Z. gp96, an endoplasmic reticulum master chaperone for integrins and Toll-like receptors, selectively regulates early T and B lymphopoiesis. Blood 2010, 115, 2380–2390. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.; Liu, B.; Chiosis, G.; Gewirth, D.T.; Li, Z. α7 helix region of αI domain is crucial for integrin binding to endoplasmic reticulum chaperone gp96: A potential therapeutic target for cancer metastasis. J. Biol. Chem. 2013, 288, 18243–18248. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.K.; DeLeo, A.B.; Old, L.J. Tumor rejection antigens of chemically induced sarcomas of inbred mice. Proc. Natl. Acad. Sci. USA 1986, 83, 3407–3411. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.C.; Lin, C.Y.; Lee, L.Y.; Chen, Y.J.; Lu, Y.C.; Wang, H.M.; Liao, C.T.; Chang, J.T.C.; Cheng, A.J. Molecular chaperones as a common set of proteins that regulate the invasion phenotype of head and neck cancer. Clin. Cancer Res. 2011, 17, 4629–4641. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chen, C.; Ma, C.; Sun, S.; Zhang, J.; Sun, Y. Expression of heat-shock protein gp96 in gallbladder cancer and its prognostic clinical significance. Int. J. Clin. Exp. Pathol. 2015, 8, 1946–1953. [Google Scholar] [PubMed]
- Haverty, A.A.; Harmey, J.H.; Redmond, H.P.; Bouchier-Hayes, D.J. Interleukin-6 upregulates GP96 expression in breast cancer. J. Surg. Res. 1997, 69, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Rachidi, S.; Sun, S.; Wu, B.X.; Jones, E.; Drake, R.R.; Ogretmen, B.; Cowart, L.A.; Clarke, C.J.; Hannun, Y.A.; Chiosis, G.; et al. Endoplasmic reticulum heat shock protein gp96 maintains liver homeostasis and promotes hepatocellular carcinogenesis. J. Hepatol. 2015, 62, 879–888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.; White-Gilbertson, S.; Kellner, J.; Rachidi, S.; Usmani, S.Z.; Chiosis, G.; Depinho, R.; Li, Z.; Liu, B. Molecular chaperone gp96 is a novel therapeutic target of multiple myeloma. Clin. Cancer Res. 2013, 19, 6242–6251. [Google Scholar] [CrossRef] [PubMed]
- Morales, C.; Rachidi, S.; Hong, F.; Sun, S.; Ouyang, X.; Wallace, C.; Zhang, Y.; Garret-Mayer, E.; Wu, J.; Liu, B.; et al. Immune chaperone gp96 drives the contributions of macrophages to inflammatory colon tumorigenesis. Cancer Res. 2014, 74, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Robert, J.; Ménoret, A.; Cohen, N. Cell surface expression of the endoplasmic reticular heat shock protein gp96 is phylogenetically conserved. J. Immunol. 1999, 163, 4133–4139. [Google Scholar] [PubMed]
- Dai, J.; Liu, B.; Caudill, M.M.; Zheng, H.; Qiao, Y.; Podack, E.R.; Li, Z. Cell surface expression of heat shock protein gp96 enhances cross-presentation of cellular antigens and the generation of tumor-specific T cell memory. Cancer Immun. 2003, 3, 1–11. [Google Scholar] [PubMed]
- Linderoth, N.A.; Popowicz, A.; Sastry, S. Identification of the peptide-binding site in the heat shock chaperone/tumor rejection antigen gp96 (Grp94). J. Biol. Chem. 2000, 275, 5472–5477. [Google Scholar] [CrossRef] [PubMed]
- Kovalchin, J.T.; Murthy, A.S.; Horattas, M.C.; Guyton, D.P.; Chandawarkar, R.Y. Determinants of efficacy of immunotherapy with tumor-derived heat shock protein gp96. Cancer Immun. Arch. 2001, 1, 7. [Google Scholar]
- Sabbatino, F.; Favoino, E.; Wang, Y.; Wang, X.; Villani, V.; Cai, L.; Yang, L.; Ferrone, S.; Ferrone, C. Grp94-specific monoclonal antibody to counteract BRAF inhibitor resistance in BRAFV600E melanoma. J. Transl. Med. 2015, 13, K12. [Google Scholar] [CrossRef] [Green Version]
- Taldone, T.; Ochiana, S.O.; Patel, P.D.; Chiosis, G. Selective targeting of the stress chaperome as a therapeutic strategy. Trends Pharmacol. Sci. 2014, 35, 592–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duerfeldt, A.S.; Peterson, L.B.; Maynard, J.C.; Ng, C.L.; Eletto, D.; Ostrovsky, O.; Shinogle, H.E.; Moore, D.S.; Argon, Y.; Nicchitta, C.V.; et al. Development of a Grp94 inhibitor. J. Am. Chem. Soc. 2012, 134, 9796–9804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Immormino, R.M.; Metzger, L.E.; Reardon, P.N.; Dollins, D.E.; Blagg, B.S.J.; Gewirth, D.T. Different poses for ligand and chaperone in inhibitor-bound Hsp90 and GRP94: Implications for paralog-specific drug design. J. Mol. Biol. 2009, 388, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.D.; Yan, P.; Seidler, P.M.; Patel, H.J.; Sun, W.; Yang, C.; Que, N.S.; Taldone, T.; Finotti, P.; Stephani, R.A.; et al. Paralog-selective Hsp90 inhibitors define tumor-specific regulation of HER2. Nat. Chem. Biol. 2013, 9, 677–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozcan, L.; Tabas, I. Role of endoplasmic reticulum stress in metabolic disease and other disorders. Annu. Rev. Med. 2012, 63, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Xiang, C.; Wang, Y.; Zhang, H.; Han, F. The role of endoplasmic reticulum stress in neurodegenerative disease. Apoptosis 2017, 22, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Viana, R.J.S.; Steer, C.J.; Rodrigues, C.M.P. Amyloid-β peptide-induced secretion of endoplasmic reticulum chaperone glycoprotein GRP94. J. Alzheimers Dis. 2011, 27, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Villadiego, J.; Labrador-Garrido, A.; Franco, J.M.; Leal-Lasarte, M.; De Genst, E.J.; Dobson, C.M.; Pozo, D.; Toledo-Aral, J.J.; Roodveldt, C. Immunization with α-synuclein/Grp94 reshapes peripheral immunity and suppresses microgliosis in a chronic Parkinsonism model. Glia 2018, 66, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hong, F.; Gewirth, D.; Guo, B.; Liu, B.; Li, Z. The molecular chaperone gp96/GRP94 interacts with toll-like receptors and integrins via its C-terminal hydrophobic domain. J. Biol. Chem. 2012, 287, 6735–6742. [Google Scholar] [CrossRef] [PubMed]
- Xagorari, A.; Chlichlia, K. Toll-Like Receptors and Viruses: Induction of Innate Antiviral Immune Responses. Open Microbiol. J. 2008, 2, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Innate immune recognition of viral infection. Nat. Immunol. 2006, 7, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Bloor, S.; Maelfait, J.; Krumbach, R.; Beyaert, R.; Randow, F. Endoplasmic reticulum chaperone gp96 is essential for infection with vesicular stomatitis virus. Proc. Natl. Acad. Sci. USA 2010, 107, 6970–6975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, A.; Bellamy, R.; Taylor, J. BiP (GRP78) and endoplasmin (GRP94) are induced following rotavirus infection and bind transiently to an endoplasmic reticulum-localized virion component. J. Virol. 1998, 72, 9865–9872. [Google Scholar] [PubMed]
- Fan, H.; Yan, X.; Zhang, Y.; Zhang, X.; Gao, Y.; Xu, Y.; Wang, F.; Meng, S. Increased Expression of Gp96 by HBx-Induced NF-κB Activation Feedback Enhances Hepatitis B Virus Production. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.D.; Li, C.L.; Lang, Z.W.; Gao, G.F.; Tien, P. Significant correlation between expression level of HSP gp96 and progression of hepatitis B virus induced diseases. World J. Gastroenterol. 2004, 10, 1141–1145. [Google Scholar] [CrossRef] [PubMed]
- Xiao, A.; Wong, J.; Luo, H. Viral interaction with molecular chaperones: Role in regulating viral infection. Arch. Virol. 2010, 155, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.S.; Shin, H.J.; Cho, Y.H.; Rho, H.M. Expression of stable hepatitis B viral polymerase associated with GRP94 in E. coli. Arch. Virol. 2000, 145, 1305–1320. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.Y.; Tsai, M.C.; Wu, Y.P.; Wang, R.Y.L. Identification of heat-shock protein 90 beta in Japanese encephalitis virus-induced secretion proteins. J. Gen. Virol. 2011, 92, 2803–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strbo, N.; Romero, L.; Podack, E.R.; Villasante, E.F.; Edgel, K.A.; Strbo, N. Secreted heat shock protein gp96-Ig vaccine for malaria prophylaxis. J. Immunol. 2016, 196, 146.10. [Google Scholar]
- Kim, D.; Song, L.; Wang, J.; Wu, H.; Gu, G.; Sugi, Y.; Li, Z.; Wang, H. GRP94 Is an Essential Regulator of Pancreatic β-Cell Development, Mass, and Function in Male Mice. Endocrinology 2018, 159, 1062–1073. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, M.R.; Matassa, D.S.; Sisinni, L.; Lettini, G.; Landriscina, M.; Esposito, F. TRAP1 revisited: Novel localizations and functions of a “next-generation” biomarker (Review). Int. J. Oncol. 2014, 45, 969–977. [Google Scholar] [CrossRef] [PubMed]
- Matassa, D.S.; Agliarulo, I.; Avolio, R.; Landriscina, M.; Esposito, F. TRAP1 Regulation of Cancer Metabolism: Dual Role as Oncogene or Tumor Suppressor. Genes 2018, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Costantino, E.; Maddalena, F.; Calise, S.; Piscazzi, A.; Tirino, V.; Fersini, A.; Ambrosi, A.; Neri, V.; Esposito, F.; Landriscina, M. TRAP1, a novel mitochondrial chaperone responsible for multi-drug resistance and protection from apoptotis in human colorectal carcinoma cells. Cancer Lett. 2009, 279, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.G.; Koh, H.J.; Roh, M.S. Clinicopathologic significance of TRAP1 expression in colorectal cancer: A large scale study of human colorectal adenocarcinoma tissues. Diagn. Pathol. 2017, 12, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Maddalena, F.; Sisinni, L.; Lettini, G.; Condelli, V.; Matassa, D.S.; Piscazzi, A.; Amoroso, M.R.; La Torre, G.; Esposito, F.; Landriscina, M. Resistance to paclitxel in breast carcinoma cells requires a quality control of mitochondrial antiapoptotic proteins by TRAP1. Mol. Oncol. 2013, 7, 895–906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wang, J.; Huang, Z.; Wei, P.; Liu, Y.; Hao, J.; Zhao, L.; Zhang, F.; Tu, Y.; Wei, T. Aberrantly upregulated TRAP1 is required for tumorigenesis of breast cancer. Oncotarget 2015, 6, 44495–44508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leav, I.; Plescia, J.; Goel, H.L.; Li, J.; Jiang, Z.; Cohen, R.J.; Languino, L.R.; Altieri, D.C. Cytoprotective mitochondrial chaperone TRAP-1 as a novel molecular target in localized and metastatic prostate cancer. Am. J. Pathol. 2010, 176, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Lisanti, S.; Garlick, D.S.; Bryant, K.G.; Tavecchio, M.; Mills, G.B.; Lu, Y.; Kossenkov, A.V.; Showe, L.C.; Languino, L.R.; Altieri, D.C. Transgenic expression of the mitochondrial chaperone TNFR-associated protein 1 (TRAP1) accelerates prostate cancer development. J. Biol. Chem. 2016, 291, 25247–25254. [Google Scholar] [CrossRef] [PubMed]
- Im, C.N.; Seo, J.S. Overexpression of tumor necrosis factor receptor-associated protein 1 (TRAP1), leads to mitochondrial aberrations in mouse fibroblast NIH/3T3 cells. BMB Rep. 2014, 47, 280–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegelin, M.D.; Plescia, J.; Raskett, C.M.; Gilbert, C.A.; Ross, A.H.; Altieri, D.C. Global Targeting of Subcellular Heat Shock Protein-90 Networks for Therapy of Glioblastoma. Mol. Cancer Ther. 2010, 9, 1638–1646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aust, S.; Bachmayr-Heyda, A.; Pateisky, P.; Tong, D.; Darb-Esfahani, S.; Denkert, C.; Chekerov, R.; Sehouli, J.; Mahner, S.; Van Gorp, T.; et al. Role of TRAP1 and estrogen receptor alpha in patients with ovarian cancer—A study of the OVCAD consortium. Mol. Cancer 2012, 11. [Google Scholar] [CrossRef] [PubMed]
- Matassa, D.S.; Amoroso, M.R.; Lu, H.; Avolio, R.; Arzeni, D.; Procaccini, C.; Faicchia, D.; Maddalena, F.; Simeon, V.; Agliarulo, I.; et al. Oxidative metabolism drives inflammation-induced platinum resistance in human ovarian cancer. Cell Death Differ. 2016, 23, 1542–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Tsutsumi, S.; Muhlebach, G.; Sourbier, C.; Lee, M.-J.; Lee, S.; Vartholomaiou, E.; Tatokoro, M.; Beebe, K.; Miyajima, N.; et al. Molecular chaperone TRAP1 regulates a metabolic switch between mitochondrial respiration and aerobic glycolysis. Proc. Natl. Acad. Sci. USA 2013, 110, E1604–E1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takamura, H.; Koyama, Y.; Matsuzaki, S.; Yamada, K.; Hattori, T.; Miyata, S.; Takemoto, K.; Tohyama, M.; Katayama, T. TRAP1 Controls Mitochondrial Fusion/Fission Balance through Drp1 and Mff Expression. PLoS ONE 2012, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Karsten, P.; Hamm, S.; Pogson, J.H.; Müller-Rischart, A.K.; Exner, N.; Haass, C.; Whitworth, A.J.; Winklhofer, K.F.; Schulz, J.B.; et al. TRAP1 rescues PINK1 loss-of-function phenotypes. Hum. Mol. Genet. 2013, 22, 2829–2841. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.C.; Loh, S.H.; Martins, L.M. Drosophila Trap1 protects against mitochondrial dysfunction in a PINK1/parkin model of Parkinson’s disease. Cell Death Dis. 2013, 4, e467. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.K.; Voigt, A.; Lutz, A.K.; Toegel, J.P.; Gerhardt, E.; Karsten, P.; Falkenburger, B.; Reinartz, A.; Winklhofer, K.F.; Schulz, J.B. The mitochondrial chaperone protein TRAP1 mitigates α-synuclein toxicity. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadei, N.; Sood, P.; Ulrich, T.; Fallier-Becker, P.; Kieper, N.; Helling, S.; May, C.; Glaab, E.; Chen, J.; Nuber, S.; et al. Mitochondrial defects and neurodegeneration in mice overexpressing wild-type or G399S mutant HtrA2. Hum. Mol. Genet. 2016, 25, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, J.C.; Zimprich, A.; Berrio, D.A.C.; Schindler, K.M.; Maurer, B.; Schulte, C.; Bus, C.; Hauser, A.K.; Kübler, M.; Lewin, R.; et al. Metformin reverses TRAP1 mutation-associated alterations in mitochondrial function in Parkinson’s disease. Brain 2017, 140, 2444–2459. [Google Scholar] [CrossRef] [PubMed]
- Fismen, S.; Thiyagarajan, D.; Seredkina, N.; Nielsen, H.; Jacobsen, S.; Elung-Jensen, T.; Kamper, A.L.; Johansen, S.D.; Mortensen, E.S.; Rekvig, O.P. Impact of the tumor necrosis factor receptor-associated protein 1 (Trap1) on renal DNaseI shutdown and on progression of murine and human lupus nephritis. Am. J. Pathol. 2013, 182, 688–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saisawat, P.; Kohl, S.; Hilger, A.C.; Hwang, D.Y.; Yung Gee, H.; Dworschak, G.C.; Tasic, V.; Pennimpede, T.; Natarajan, S.; Sperry, E.; et al. Whole-exome resequencing reveals recessive mutations in TRAP1 in individuals with CAKUT and VACTERL association. Kidney Int. 2014, 85, 1310–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Domain structure of HSP90 family members in humans. The numbering 1–900 indicates the amino acid sequence. The lengths of HSP90α, HSP90β, GRP94, and TRAP1 are 732, 724, 704, and 803 amino acids, respectively. Below is shown a schematic representation of the domain structure of HSP90 isoforms together with the biological function of each domain.
Figure 1.
Domain structure of HSP90 family members in humans. The numbering 1–900 indicates the amino acid sequence. The lengths of HSP90α, HSP90β, GRP94, and TRAP1 are 732, 724, 704, and 803 amino acids, respectively. Below is shown a schematic representation of the domain structure of HSP90 isoforms together with the biological function of each domain.

Figure 2.
Schematic representation of the ATPase cycle of HSP90. The open and closed conformations of HSP90 are depicted.
Figure 2.
Schematic representation of the ATPase cycle of HSP90. The open and closed conformations of HSP90 are depicted.

Figure 3.
The mechanism of protein folding by HSP90 [44]. (I) The client protein either mis-folded protein or nascent polypeptide is bound by Hsp70/Hsp40/ADP complex to be protected from potential aggregation [102]; (II) The protein complex formed is further stabilized by Hsp70-interacting protein (HIP) or Bcl2 which support the change of ADP to ATP [103]; (III) HSP90 binds the client protein harboured by the Hsp70/Hsp40 protein complex to start its action; (IV) The interaction between HSP90 and HSP70 is facilitated by the adaptor protein, HOP/Sti1 (Hsp90–Hsp70 organizing protein) [104]. Importantly, additional co-chaperone known as Cdc37 (cell-division-cycle 37 homologue) or p50 is needed for loading client kinases onto HSP90 [105]; (V) As the client protein is loaded, other co-chaperones, immunophilins like (FKBP51, FKBP52) are added to the HSP90 homodimer forming an activated heteroprotein complex while the HSP70, HIP and HOP are released [106]; (VI) Binding of ATP to the NTD of HSP90 in the heteroprotein complex switches the HSP90 from the “open state” to the “closed state” [107]. Various inhibitors of HSP90 can act at this stage by competing with ATP for the NTD binding site consequently resulting in protein degradation by proteasome [101]; (VII) If no inhibitors interfere with the folding cycle, association of other co-chaperones occurs including p23 and Aha1 (activator of Hsp90 ATPase homologue 1); (VIII) Binding of Aha1, not shown in the diagram, to the MD of HSP90 induces ATP hydrolysis and supports folding of the bound client, as well as enhances the release of immunophilins and co-chaperones [96].
Figure 3.
The mechanism of protein folding by HSP90 [44]. (I) The client protein either mis-folded protein or nascent polypeptide is bound by Hsp70/Hsp40/ADP complex to be protected from potential aggregation [102]; (II) The protein complex formed is further stabilized by Hsp70-interacting protein (HIP) or Bcl2 which support the change of ADP to ATP [103]; (III) HSP90 binds the client protein harboured by the Hsp70/Hsp40 protein complex to start its action; (IV) The interaction between HSP90 and HSP70 is facilitated by the adaptor protein, HOP/Sti1 (Hsp90–Hsp70 organizing protein) [104]. Importantly, additional co-chaperone known as Cdc37 (cell-division-cycle 37 homologue) or p50 is needed for loading client kinases onto HSP90 [105]; (V) As the client protein is loaded, other co-chaperones, immunophilins like (FKBP51, FKBP52) are added to the HSP90 homodimer forming an activated heteroprotein complex while the HSP70, HIP and HOP are released [106]; (VI) Binding of ATP to the NTD of HSP90 in the heteroprotein complex switches the HSP90 from the “open state” to the “closed state” [107]. Various inhibitors of HSP90 can act at this stage by competing with ATP for the NTD binding site consequently resulting in protein degradation by proteasome [101]; (VII) If no inhibitors interfere with the folding cycle, association of other co-chaperones occurs including p23 and Aha1 (activator of Hsp90 ATPase homologue 1); (VIII) Binding of Aha1, not shown in the diagram, to the MD of HSP90 induces ATP hydrolysis and supports folding of the bound client, as well as enhances the release of immunophilins and co-chaperones [96].


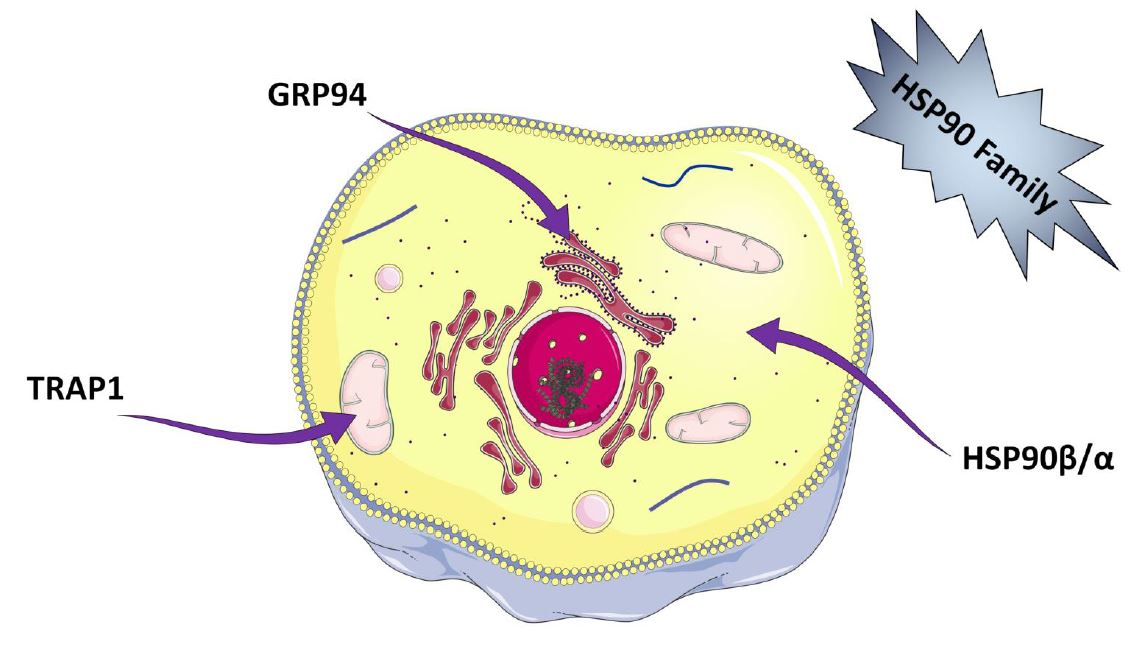{kind=link}
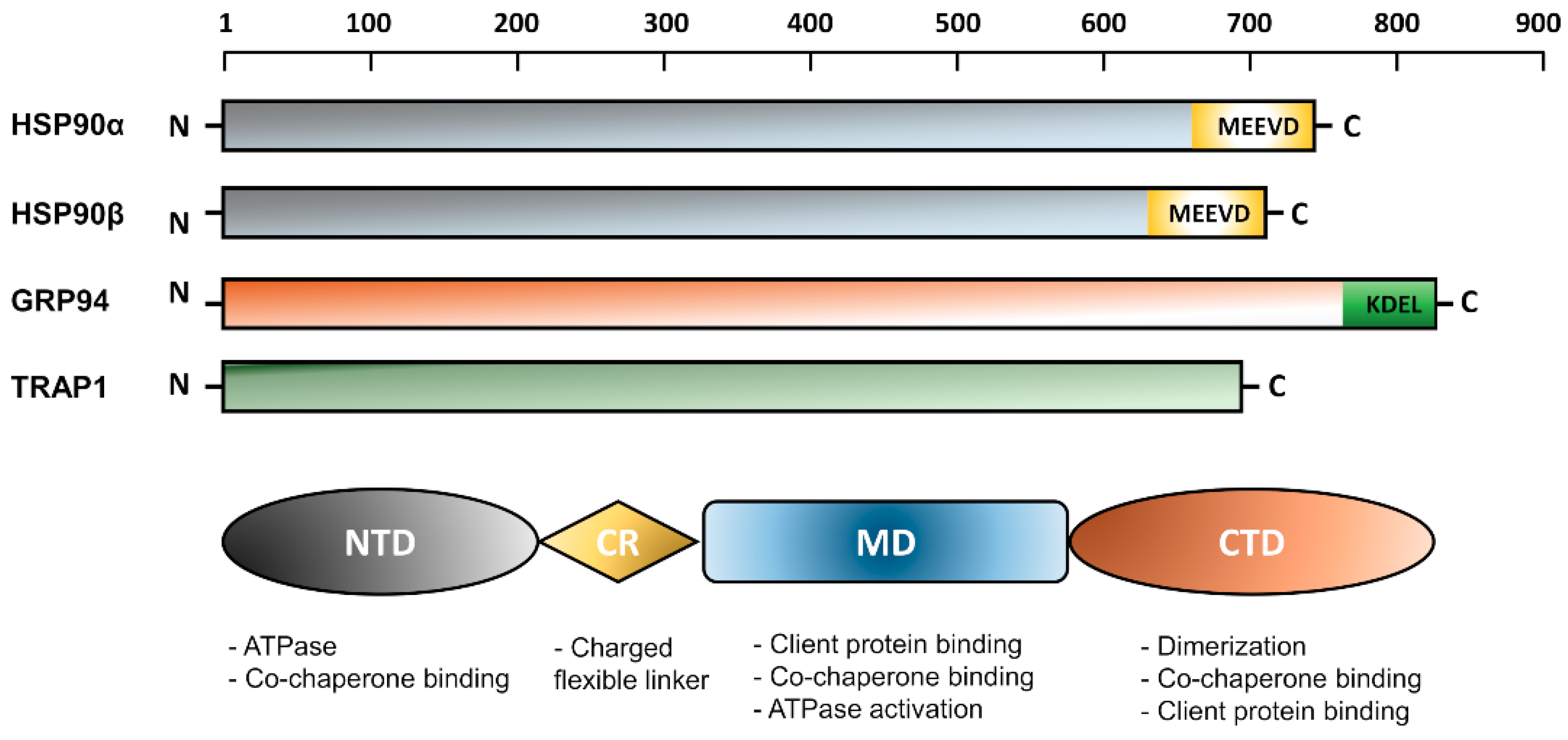{kind=link}
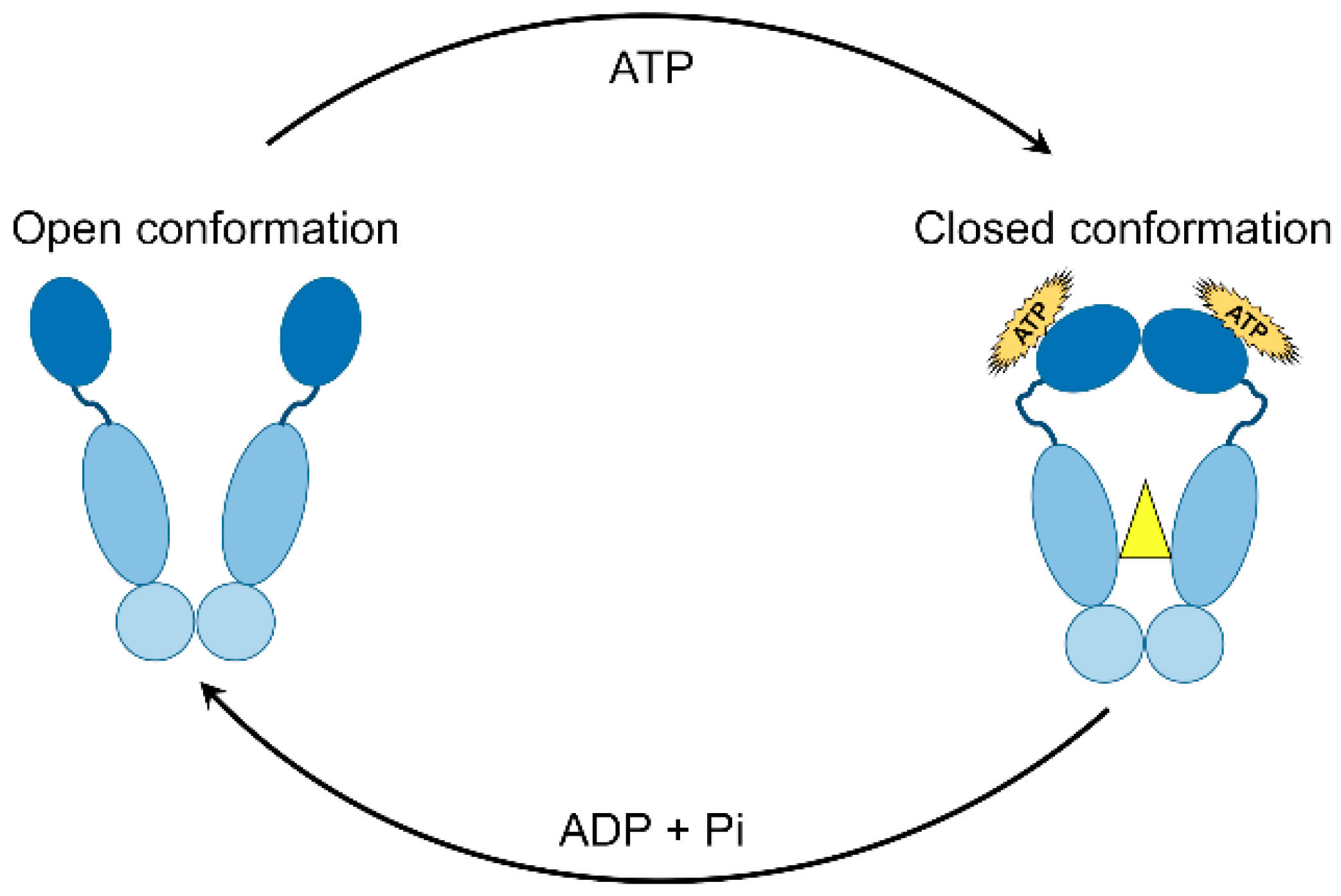{kind=link}
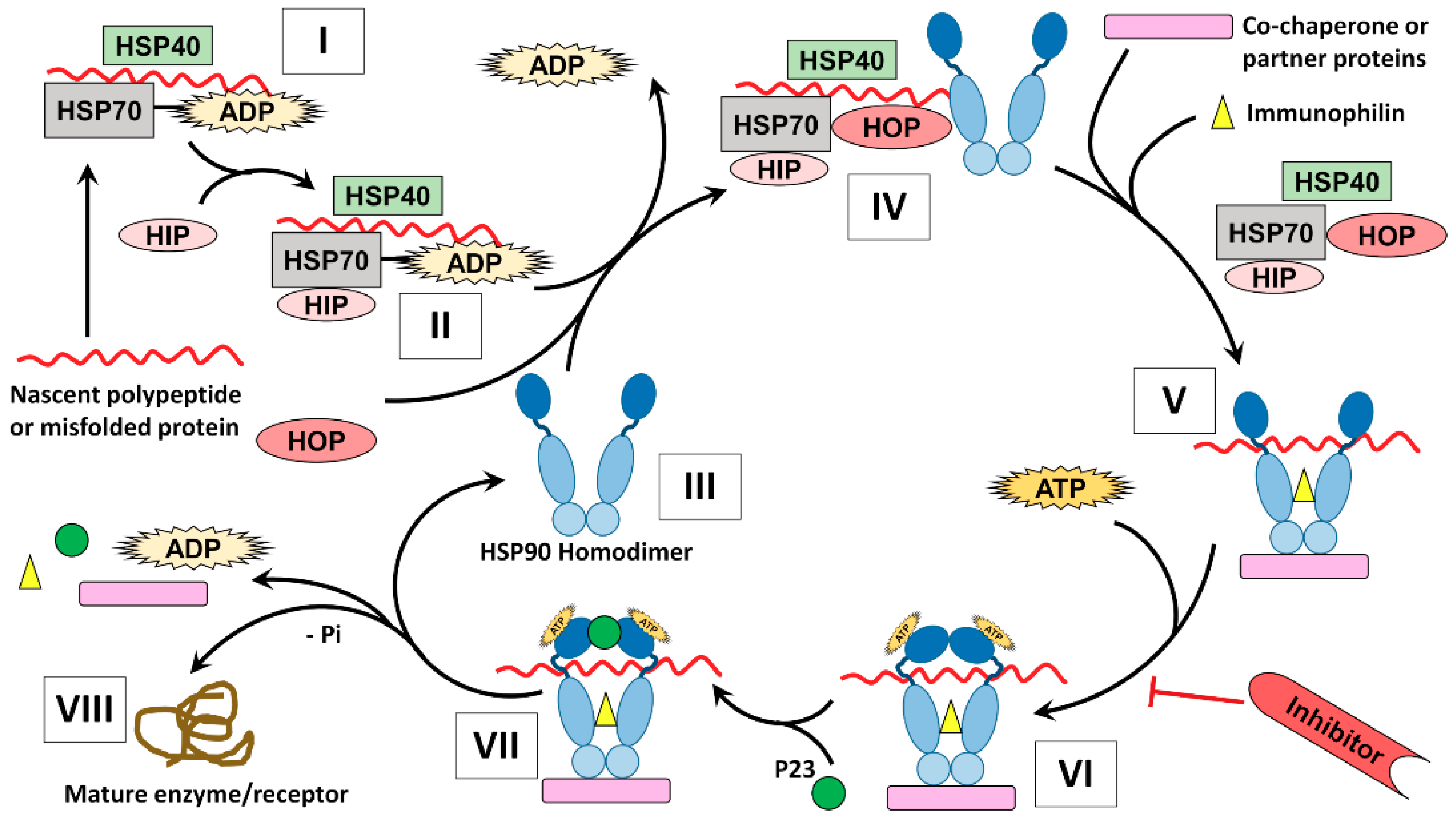{kind=link}
Table 1.
Different members of human HSPC (HSP90) family [5].
Table 1.
Different members of human HSPC (HSP90) family [5].
| Gene Name | Protein Name | Alternative Name | Human Gene ID |
|---|---|---|---|
| HSPC1 | HSPC1 | HSP90AA1; HSPN; LAP2; HSP86; HSPC1; HSPCA; HSP89; HSP90; HSP90A; HSP90N; HSPCAL1; HSPCAL4; FLJ31884 | 3320 |
| HSPC2 | HSPC2 | HSP90AA2; HSPCA; HSPCAL3; HSP90ALPHA | 3324 |
| HSPC3 | HSPC3 | HSP90AB1; HSPC2; HSPCB; D6S182; HSP90B; FLJ26984; HSP90-BETA | 3326 |
| HSPC4 | HSPC4 | HSP90B1; ECGP; GP96; TRA1; GRP94; endoplasmin | 7184 |
| HSPC5 | HSPC5 | TRAP1; HSP75; HSP90L | 10131 |
| Co-Chaperone or Protein Partner | Role in Carcinogenesis |
|---|---|
| Aha1 | Stimulates ATPase activity |
| Cdc37 | Mediates activation of protein kinase substrates |
| CHIP | Involved in degradation of unfolded client proteins |
| Cyclophilin-40 | Peptidyl propyl isomerase |
| FKBP51 and 52 | Peptidyl propyl isomerase |
| HOP | Mediates interaction between Hsp90 and Hsp70 |
| Hsp40 | Stabilizes and delivers client proteins to Hsp90 complex |
| Hsp70 | Stabilizes and delivers client proteins to Hsp90 |
| p23 | Stabilizes closed, clamped substrate bound |
| HIP | Inhibits ATPase activity of Hsp70 |
| PP5 | Protein phosphatase 5 |
| Tom70 | Facilitates translocation of pre-proteins into mitochondrial matrix |
| Sgt1 | Client adaptor, involved in client recruitment |
| WISp39 | Regulates p21 stability |
Table 3.
Protein clients of GERP94 and TRAP1.
| GRP94 Client Protein | Citation | TRAP1 Client Protein | Citation |
|---|---|---|---|
| MHC class II | [72] | TNF receptor | [56] |
| ErbB2 | [73] | retinoblastoma protein | [74] |
| Insulin-like growth factor -II | [75] | Tumour suppressor EXT proteins | [76] |
| P43 | [77] | CYP-D | [60] |
| CLAVATA complex | [78] | Calcium-binding protein, Sorcin | [79] |
| Bile salt dependent lipase | [80] | Proteasome regulatory particle TBP7 | [81] |
| Golgi apparatus casein kinase | [82] | ||
| Cartilage oligomeric matrix protein (COMP) | [83] | ||
| α1-antitrypsin 1 | [84] | ||
| Thyroglobulin | [85] | ||
| HSV-1 glycoprotein B 1 | [86] | ||
| Apolipoprotein B | [87] | ||
| Collagen | [88] | ||
| Protein C 1 | [89] | ||
| Protein apparatus casein Kinase | [82] | ||
| TLR 1,2,4 | [90] | ||
| Ig heavy chain and light chain | [91] | ||
| Ig α chain | [92] | ||
| Integrins | [90] | ||
| Thrombospondin | [93] |
1 Indicates mutant forms of the protein.
| Protein Client | Role in Carcinogenesis |
|---|---|
| Receptor tyrosine kinases, serine/threonine kinases, steroid hormone receptors | Uncontrolled proliferation |
| Telomerases | Immortalization |
| AKT, NF-κB, p53, c-MET, Apaf-1, Survivin | Anti-apoptotic |
| HIF1α, VEGFR, PI3K/AKT, RTKs, flt-3 | Angiogenesis |
| IRAK3 | Escaping immune destruction |
| ARNT, ARRB1, HIF-1α, HMG1, SREBF1 | Modification of cellular energetics |
| FANCA, MAFG, NEK8, NEK9, NEK11 | Genome instability and mutation |
| IL-6, IL-8, IRAK1, IRAK2, IRAK3 | Tumor-promoting inflammation |
| Plk, Wee1, Myc1, CDK4, CDK6, Myt1 | Evading growth suppressors |
| MMP2,c-MET, SSDF-1 | Invasion and metastasis |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hoter, A.; El-Sabban, M.E.; Naim, H.Y. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. Int. J. Mol. Sci. 2018, 19, 2560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092560
AMA Style
Hoter A, El-Sabban ME, Naim HY. The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease. International Journal of Molecular Sciences. 2018; 19(9):2560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092560
Chicago/Turabian StyleHoter, Abdullah, Marwan E. El-Sabban, and Hassan Y. Naim. 2018. "The HSP90 Family: Structure, Regulation, Function, and Implications in Health and Disease" International Journal of Molecular Sciences 19, no. 9: 2560. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092560
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.