Structure-Based Classification and Anti-Cancer Effects of Plant Metabolites


 and
and
Abstract
:1. Introduction
2. Phytochemicals as Bioactive Metabolites
2.1. Phenolic Compounds
2.1.1. Flavonoids
2.1.2. Stilbenes
2.1.3. Phenolic Acids
2.2. Terpenoids
2.2.1. Monoterpenoids
2.2.2. Diterpenoids
2.2.3. Tetraterpenoids (Carotenoids)
2.3. Nitrogen-Containing Alkaloids and Sulfur-Containing Compounds
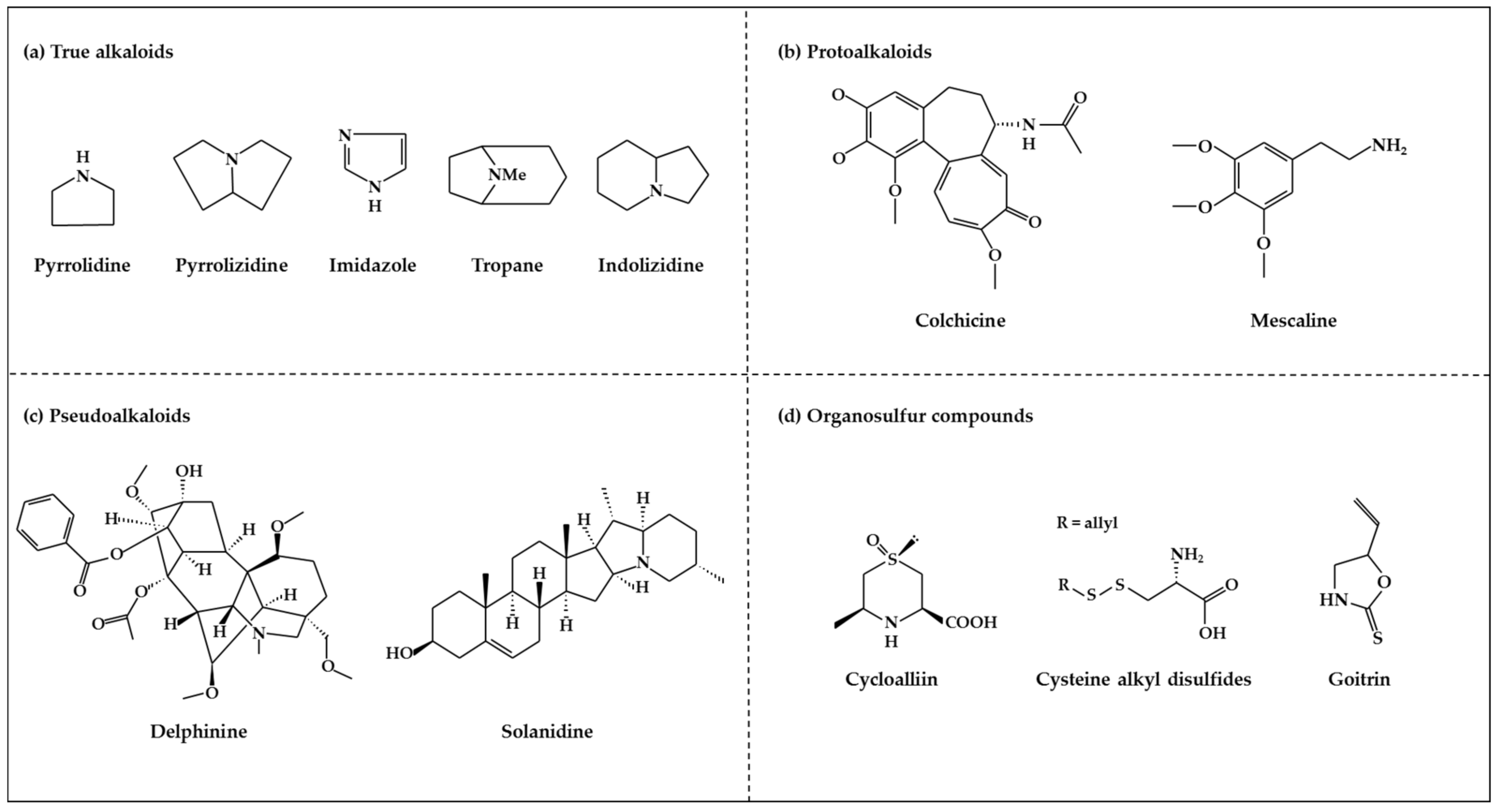
2.3.1. Alkaloids
2.3.2. Organosulfur Compounds
3. Anti-Tumor Activity of Plant Metabolites in Various Malignant Cancers
3.1. Colorectal Cancer
3.2. Gastric Cancer
3.3. Lung Cancer
3.4. Breast Cancer
3.5. Prostate Cancer
3.6. Hematologic Cancer
3.7. Skin Cancer
3.8. Head and Neck Cancer
4. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OSC | Organosulfur compounds |
| GOFA | 4′-geranyloxyferulic acid |
| GAP | 3-(4′-geranyloxy-3′;-methoxyphenyl)-l-alanyl-l-proline |
| AOM | Azoxymethane |
| DSS | Dextran sodium sulfate |
| β-CD | β-cyclodextrin |
| AUR | Auraptene |
| SIRT1 | Sirtuin 1 |
| ITCs | Isothiocyanates |
| SFN | Sulforaphane |
| BITC | Benzyl isothiocyanate |
| PEITC | Phenethyl isothiocyanate |
| MMPs | Matrix metalloproteinases |
| LVD | Lymphatic vessel density |
| EGCG | Epigallocatechin gallate |
| miRNA | micro RNA |
| HIF-1α | Hypoxia-inducible factor 1alpha |
| TGF-β | Transforming growth factor β |
| EMT | Epithelial-to-mesenchymal transition |
| ERK1/2 | Extracellular signal–regulated kinases 1/2 |
| NSCLC | Nonsmall cell lung cancer |
| SCLC | Small cell lung cancer |
| MAPK | Mitogen-activated protein kinase |
| B(a)P | Benzo(a)pyrene |
| TRAIL | Tumor necrosis factor-related apoptosis-inducing ligand |
| I3C | Indole-3-carbinol |
| VC | Vinyl carbamate |
| EGFR | Epidermal growth factor receptor |
| NF-κB | Nuclear factor kappaB |
| mTOR | mammalian target of rapamycin |
| u-PA | urokinase-type plasminogen activator |
| CPA | Cyclophosphamide |
| PC | Punicalagin |
| PFE | Pomegranate fruit extracts |
| MTA1 | Metastasis-associated protein1 |
| AMPK | AMP-activated protein kinase |
| STAT3 | Signal transducer and activator of transcription 3 |
| ROS | Reactive oxygen species |
| ERα | Estrogen receptor α |
| AhR | Aryl hydrocarbon receptor |
| ATF-3 | cyclic AMP dependent transcription factor |
| IKK | Inhibitor against IκB kinase |
| BA | Betulinic acid |
| GR | Glucocorticoid receptor |
| AP1 | Activator protein 1 |
| JNK | c-Jun N-terminal Kinase |
| AMR | Amooranin |
| AECHL-1 | Ailanthus excelsa chloroform extract-1 |
| BPA | Bisphenol A |
| miRNA-19 | microRNA-19 |
| AGN | Angelica gigas Nakai |
| TRAMP | Transgenic adenocarcinoma of mouse prostate |
| MAP | Microtubule associated proteins |
| NudC | Nuclear migration protein |
| CAF | Cancer associated fibroblast |
| MAOA | Monoamine oxidase A |
| MTX | Mitoxantrone |
| DAS | Diallyl sulfide |
| DADS | Diallyl disulfide |
| DATS | Diallyl trisulfide |
| DMBA | 7,12-dimethylbenz[a]anthracene |
| ESCC | Esophageal squamous cell carcinoma |
| HNSCC | Head and neck squamous cell carcinoma |
References
- Lee, C.S.; Baek, J.; Han, S.Y. The role of kinase modulators in cellular senescence for use in cancer treatment. Molecules 2017, 22, 1411. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.R.; Kucuk, O.; Khuri, F.R.; Shin, D.M. Perspectives for cancer prevention with natural compounds. J. Clin. Oncol. 2009, 27, 2712–2725. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, M.; Kapulnik, Y.; Koltai, H. Plant derived substances with anti-cancer activity: From folklore to practice. Front. Plant Sci. 2015, 6, 799. [Google Scholar] [CrossRef] [PubMed]
- Gali-Muhtasib, H.; Hmadi, R.; Kareh, M.; Tohme, R.; Darwiche, N. Cell death mechanisms of plant-derived anticancer drugs: Beyond apoptosis. Apoptosis 2015, 20, 1531–1562. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, K.M.; Yuraszeck, T.M.; Li, C.C.; Zhang, Y.; Kasichayanula, S. Immunotherapy and novel combinations in oncology: Current landscape, challenges, and opportunities. Clin. Transl. Sci. 2016, 9, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.; Kostyuk, V. Biotechnologically produced secondary plant metabolites for cancer treatment and prevention. Curr. Pharm. Biotechnol. 2012, 13, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.M.; Ribnicky, D.M.; Lipsky, P.E.; Raskin, I. Revisiting the ancient concept of botanical therapeutics. Nat. Chem. Biol. 2007, 3, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Gordaliza, M. Natural products as leads to anticancer drugs. Clin. Transl. Oncol. 2007, 9, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Ijaz, S.; Akhtar, N.; Khan, M.S.; Hameed, A.; Irfan, M.; Arshad, M.A.; Ali, S.; Asrar, M. Plant derived anticancer agents: A green approach towards skin cancers. Biomed. Pharmacother. 2018, 103, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Molyneux, R.J.; Lee, S.T.; Gardner, D.R.; Panter, K.E.; James, L.F. Phytochemicals: The good, the bad and the ugly? Phytochemistry 2007, 68, 2973–2985. [Google Scholar] [CrossRef] [PubMed]
- Santhi, K.S.; Sengottuvel, R. Qualitative and quantitative phytochemical analysis of moringa concanensis nimmo. Int. J. Curr. Microbiol. App. Sci. 2016, 5, 633–640. [Google Scholar] [CrossRef]
- Pichersky, E.; Gang, D.R. Genetics and biochemistry of secondary metabolites in plants: An evolutionary perspective. Trends. Plant. Sci. 2000, 5, 439–445. [Google Scholar] [CrossRef]
- Baxter, H.; Harborne, J.B.; Moss, G.P. Phytochemical Dictionary: A Handbook of Bioactive Compounds from Plants; CRC Press: New York, NY, USA, 1998. [Google Scholar]
- Kumar, A.; Irchhaiya, R.; Yadav, A.; Gupta, N.; Kumar, S.; Gupta, N.; Kumar, S.; Yadav, V.; Prakash, A.; Gurjar, H. Metabolites in plants and its classification. World J. Pharm. Pharm. Sci. 2015, 4, 287–305. [Google Scholar]
- Mocanu, M.M.; Nagy, P.; Szollosi, J. Chemoprevention of breast cancer by dietary polyphenols. Molecules 2015, 20, 22578–22620. [Google Scholar] [CrossRef] [PubMed]
- Varoni, E.M.; Lodi, G.; Sardella, A.; Carrassi, A.; Iriti, M. Plant polyphenols and oral health: Old phytochemicals for new fields. Curr. Med. Chem. 2012, 19, 1706–1720. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef] [PubMed]
- Crozier, A.; Clifford, M.N.; Ashihara, H. Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; John Wiley & Sons: Oxford, UK, 2008. [Google Scholar]
- Ravishankar, D.; Rajora, A.K.; Greco, F.; Osborn, H.M. Flavonoids as prospective compounds for anti-cancer therapy. Int. J. Biochem. Cell Biol. 2013, 45, 2821–2831. [Google Scholar] [CrossRef] [PubMed]
- Beecher, G.R. Overview of dietary flavonoids: Nomenclature, occurrence and intake. J Nutr 2003, 133, 3248S–3254S. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Cao, H.; Wang, Y.; Zhao, J.; Wei, X. Glycosylation of dietary flavonoids decreases the affinities for plasma protein. J. Agric. Food Chem. 2009, 57, 6642–6648. [Google Scholar] [CrossRef] [PubMed]
- Leo, C.H.; Woodman, O.L. Flavonols in the prevention of diabetes-induced vascular dysfunction. J. Cardiovasc. Pharmacol. 2015, 65, 532–544. [Google Scholar] [CrossRef] [PubMed]
- Fantini, M.; Benvenuto, M.; Masuelli, L.; Frajese, G.V.; Tresoldi, I.; Modesti, A.; Bei, R. In vitro and in vivo antitumoral effects of combinations of polyphenols, or polyphenols and anticancer drugs: Perspectives on cancer treatment. Int. J. Mol. Sci. 2015, 16, 9236–9282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Remesy, C.; Jimenez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Lea, M.A. Flavonol regulation in tumor cells. J. Cell Biochem. 2015, 116, 1190–1194. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kaur, M.; Silakari, O. Flavones: An important scaffold for medicinal chemistry. Eur. J. Med. Chem. 2014, 84, 206–239. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Hollman, P.C.; Grune, T.; Stahl, W.; Biesalski, H.K.; Williamson, G. Protection by flavanol-rich foods against vascular dysfunction and oxidative damage: 27th hohenheim consensus conference. Adv. Nutr. 2012, 3, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Mena, P.; Dominguez-Perles, R.; Girones-Vilaplana, A.; Baenas, N.; Garcia-Viguera, C.; Villano, D. Flavan-3-ols, anthocyanins, and inflammation. IUBMB Life 2014, 66, 745–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouysegu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. Engl. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- King, A.; Young, G. Characteristics and occurrence of phenolic phytochemicals. J. Am. Diet. Assoc. 1999, 99, 213–218. [Google Scholar] [CrossRef]
- Aron, P.M.; Kennedy, J.A. Flavan-3-ols: Nature, occurrence and biological activity. Mol. Nutr. Food Res. 2008, 52, 79–104. [Google Scholar] [CrossRef] [PubMed]
- Kuhnle, G.G.C. Nutrition epidemiology of flavan-3-ols: The known unknowns. Mol. Aspects Med. 2018, 61, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.P. Isoflavones: Chemistry, analysis, functions and effects on health and cancer. Asian Pac. J. Cancer Prev. 2014, 15, 7001–7010. [Google Scholar] [CrossRef] [PubMed]
- Bircsak, K.M.; Aleksunes, L.M. Interaction of isoflavones with the bcrp/abcg2 drug transporter. Curr. Drug. Metab. 2015, 16, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Vitale, D.C.; Piazza, C.; Melilli, B.; Drago, F.; Salomone, S. Isoflavones: Estrogenic activity, biological effect and bioavailability. Eur. J. Drug Metab. Pharmacokinet. 2013, 38, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Yang, W.; Bosland, M.C. Soy isoflavones and prostate cancer: A review of molecular mechanisms. J. Steroid Biochem. Mol. Biol. 2014, 140, 116–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomás-Barberán, F.A.; Clifford, M.N. Flavanones, chalcones and dihydrochalcones–nature, occurrence and dietary burden. J. Sci. Food Agric. 2000, 80, 1073–1080. [Google Scholar] [CrossRef]
- Chanet, A.; Milenkovic, D.; Manach, C.; Mazur, A.; Morand, C. Citrus flavanones: What is their role in cardiovascular protection? J. Agric. Food. Chem. 2012, 60, 8809–8822. [Google Scholar] [CrossRef] [PubMed]
- Fang, J. Bioavailability of anthocyanins. Drug Metab. Rev. 2014, 46, 508–520. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.S.; Stoner, G.D. Anthocyanins and their role in cancer prevention. Cancer Lett. 2008, 269, 281–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, T.C.; Giusti, M.M. Anthocyanins. Adv. Nutr. 2015, 6, 620–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marzocchella, L.; Fantini, M.; Benvenuto, M.; Masuelli, L.; Tresoldi, I.; Modesti, A.; Bei, R. Dietary flavonoids: Molecular mechanisms of action as anti- inflammatory agents. Recent Pat. Inflamm. Allergy Drug Discov. 2011, 5, 200–220. [Google Scholar] [CrossRef] [PubMed]
- Bellik, Y.; Boukraa, L.; Alzahrani, H.A.; Bakhotmah, B.A.; Abdellah, F.; Hammoudi, S.M.; Iguer-Ouada, M. Molecular mechanism underlying anti-inflammatory and anti-allergic activities of phytochemicals: An update. Molecules 2012, 18, 322–353. [Google Scholar] [CrossRef] [PubMed]
- Shen, T.; Wang, X.N.; Lou, H.X. Natural stilbenes: An overview. Nat. Prod. Rep. 2009, 26, 916–935. [Google Scholar] [CrossRef] [PubMed]
- Riviere, C.; Pawlus, A.D.; Merillon, J.M. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef] [PubMed]
- Sirerol, J.A.; Rodriguez, M.L.; Mena, S.; Asensi, M.A.; Estrela, J.M.; Ortega, A.L. Role of natural stilbenes in the prevention of cancer. Oxid. Med. Cell Longev. 2016, 2016, 3128951. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, B.; Ammazzalorso, A.; Fantacuzzi, M.; Giampietro, L.; Maccallini, C.; Amoroso, R. Anticancer activity of stilbene-based derivatives. ChemMedChem. 2017, 12, 558–570. [Google Scholar] [CrossRef] [PubMed]
- Kundu, J.K.; Surh, Y.J. Cancer chemopreventive and therapeutic potential of resveratrol: Mechanistic perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A. Cancer prevention and treatment with resveratrol: From rodent studies to clinical trials. Cancer Prev. Res. (Phila) 2009, 2, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [PubMed]
- Heleno, S.A.; Martins, A.; Queiroz, M.J.; Ferreira, I.C. Bioactivity of phenolic acids: Metabolites versus parent compounds: A review. Food Chem. 2015, 173, 501–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andjelković, M.; Van Camp, J.; De Meulenaer, B.; Depaemelaere, G.; Socaciu, C.; Verloo, M.; Verhe, R. Iron-chelation properties of phenolic acids bearing catechol and galloyl groups. Food Chem. 2006, 98, 23–31. [Google Scholar] [CrossRef]
- Kaushik, P.; Andujar, I.; Vilanova, S.; Plazas, M.; Gramazio, P.; Herraiz, F.J.; Brar, N.S.; Prohens, J. Breeding vegetables with increased content in bioactive phenolic acids. Molecules 2015, 20, 18464–18481. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, S.S.; Croteau, R.B. Strategies for transgenic manipulation of monoterpene biosynthesis in plants. Trends Plant Sci. 2002, 7, 366–373. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2016. [Google Scholar] [CrossRef] [PubMed]
- McCreath, S.B.; Delgoda, R. Pharmacognosy: Fundamentals, Applications and Strategies; Academic Press: London, UK, 2017. [Google Scholar]
- Bruckingham, J. Dictionary of Natural Products on Cd-Rom; Champman and Hall: New York, NY, USA, 2000. [Google Scholar]
- Harborne, J.B. hytochemical Methods: A Guide to Modern Techniques of Plant Analysis; Chapman and Hall: London, UK, 1980. [Google Scholar]
- Bhatti, H.N.; Khan, S.S.; Khan, A.; Rani, M.; Ahmad, V.U.; Choudhary, M.I. Biotransformation of monoterpenoids and their antimicrobial activities. Phytomedicine 2014, 21, 1597–1626. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.V.; Ray, M.R.; Rao, L.G. Lycopene. Adv. Food Nutr. Res. 2006, 51, 99–164. [Google Scholar] [PubMed]
- Akaberi, M.; Mehri, S.; Iranshahi, M. Multiple pro-apoptotic targets of abietane diterpenoids from salvia species. Fitoterapia 2015, 100, 118–132. [Google Scholar] [CrossRef] [PubMed]
- Soares Nda, C.; Teodoro, A.J.; Lotsch, P.F.; Granjeiro, J.M.; Borojevic, R. Anticancer properties of carotenoids in prostate cancer. A review. Histol. Histopathol. 2015, 30, 1143–1154. [Google Scholar] [PubMed]
- Stahl, W.; Sies, H. Separation of geometrical isomers of beta-carotene and lycopene. Methods Enzymol. 1994, 234, 388–400. [Google Scholar] [PubMed]
- Bendich, A.; Olson, J.A. Biological actions of carotenoids. FASEB. J. 1989, 3, 1927–1932. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.J. The role of carotenoids in human health. Nutr. Clin. Care 2002, 5, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Chew, B.P.; Park, J.S. Carotenoid action on the immune response. J. Nutr. 2004, 134, 257S–261S. [Google Scholar] [CrossRef] [PubMed]
- Bolhassani, A. Cancer chemoprevention by natural carotenoids as an efficient strategy. Anticancer Agents Med. Chem. 2015, 15, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.P.; Or, T.C.; Ip, N.Y. Plant alkaloids as drug leads for alzheimer’s disease. Neurochem. Int. 2015, 89, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Evans, W.C. Trease and Evans’ Pharmacognosy E-Book; Elsevier Health Sciences: New York, NY, USA, 2009. [Google Scholar]
- Jiang, Q.W.; Chen, M.W.; Cheng, K.J.; Yu, P.Z.; Wei, X.; Shi, Z. Therapeutic potential of steroidal alkaloids in cancer and other diseases. Med. Res. Rev. 2016, 36, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.P.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.; McNaught, A. Iupac Compendium of Chemical Terminology, (the “Gold Book”); International Union of Pure and Applied Chemistry: Zürich, Switzerland, 1997. [Google Scholar]
- Hesse, M. Alkaloids: Nature’s Curse or Blessing? John Wiley & Sons: Zurich, Switzerland, 2002. [Google Scholar]
- Cooper, R.; Nicola, G. Natural Products Chemistry: Sources, Separations and Structures; CRC Press: New York, NY, USA, 2014. [Google Scholar]
- Gupta, R.C. Nutraceuticals: Efficacy, Safety and Toxicity; Academic Press: London, UK, 2016. [Google Scholar]
- Moriarty, R.M.; Naithani, R.; Surve, B. Organosulfur compounds in cancer chemoprevention. Mini Rev. Med. Chem. 2007, 7, 827–838. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo, S.M.; Binda, N.S.; Nogueira-Machado, J.A.; Vieira-Filho, S.A.; Caligiorne, R.B. The antioxidant properties of organosulfur compounds (sulforaphane). Recent. Pat. Endocr. Metab. Immune. Drug Discov. 2015, 9, 24–39. [Google Scholar] [CrossRef] [PubMed]
- Reddivari, L.; Vanamala, J.; Chintharlapalli, S.; Safe, S.H.; Miller, J.C., Jr. Anthocyanin fraction from potato extracts is cytotoxic to prostate cancer cells through activation of caspase-dependent and caspase-independent pathways. Carcinogenesis 2007, 28, 2227–2235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuber, T.; Holm, D.; Byrne, P.; Ducreux, L.; Taylor, M.; Kaiser, M.; Stushnoff, C. Optimization of in vitro inhibition of ht-29 colon cancer cell cultures by solanum tuberosum l. Extracts. Food Funct. 2015, 6, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Yong, Y.; Matthew, S.; Wittwer, J.; Pan, L.; Shen, Q.; Kinghorn, A.D.; Swanson, S.M.; De Blanco, E.J. Dichamanetin inhibits cancer cell growth by affecting ros-related signaling components through mitochondrial-mediated apoptosis. Anticancer Res. 2013, 33, 5349–5355. [Google Scholar] [PubMed]
- Genovese, S.; Fiorito, S.; Locatelli, M.; Carlucci, G.; Epifano, F. Analysis of biologically active oxyprenylated ferulic acid derivatives in citrus fruits. Plant Foods Hum. Nutr. 2014, 69, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Genovese, S.; Epifano, F. Recent developments in the pharmacological properties of 4′-geranyloxyferulic acid, a colon cancer chemopreventive agent of natural origin. Curr. Drug Targets 2012, 13, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Epifano, F.; Curini, M.; Genovese, S.; Kim, M.; Ishigamori-Suzuki, R.; Yasui, Y.; Sugie, S.; Tanaka, T. A novel prodrug of 4′-geranyloxy-ferulic acid suppresses colitis-related colon carcinogenesis in mice. Nutr. Cancer 2008, 60, 675–684. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; de Azevedo, M.B.; Duran, N.; Alderete, J.B.; Epifano, F.; Genovese, S.; Tanaka, M.; Tanaka, T.; Curini, M. Colorectal cancer chemoprevention by 2 beta-cyclodextrin inclusion compounds of auraptene and 4′-geranyloxyferulic acid. Int. J. Cancer 2010, 126, 830–840. [Google Scholar] [CrossRef] [PubMed]
- Epifano, F.; Genovese, S.; Miller, R.; Majumdar, A.P. Auraptene and its effects on the re-emergence of colon cancer stem cells. Phytother Res. 2013, 27, 784–786. [Google Scholar] [CrossRef] [PubMed]
- Lestari, M.L.; Indrayanto, G. Curcumin. Profiles Drug Subst. Excip. Relat. Methodol. 2014, 39, 113–204. [Google Scholar] [PubMed]
- Lee, Y.H.; Song, N.Y.; Suh, J.; Kim, D.H.; Kim, W.; Ann, J.; Lee, J.; Baek, J.H.; Na, H.K.; Surh, Y.J. Curcumin suppresses oncogenicity of human colon cancer cells by covalently modifying the cysteine 67 residue of sirt1. Cancer Lett. 2018, 431, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, C.; Russo, G.L.; Orhan, I.E.; Habtemariam, S.; Daglia, M.; Sureda, A.; Nabavi, S.F.; Devi, K.P.; Loizzo, M.R.; Tundis, R.; et al. Genistein and cancer: Current status, challenges, and future directions. Adv. Nutr. 2015, 6, 408–419. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Teng, J.; Zhu, Z.; Chen, J.; Huang, W.J. Genistein induces activation of the mitochondrial apoptosis pathway by inhibiting phosphorylation of akt in colorectal cancer cells. Pharm. Biol. 2016, 54, 74–79. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Chen, J.X.; Zhu, Z.; Teng, J.A. Genistein inhibits human colorectal cancer growth and suppresses mir-95, akt and sgk1. Cell Physiol. Biochem. 2015, 35, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Liu, Z.; Wang, R.; Wang, J.; Zhang, S.; Cai, X.; Wu, K.; Bergan, R.C.; Xu, L.; Fan, D. Genistein suppresses flt4 and inhibits human colorectal cancer metastasis. Oncotarget 2015, 6, 3225–3239. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Yogosawa, S.; Izutani, Y.; Watanabe, H.; Otsuji, E.; Sakai, T. A combination of indol-3-carbinol and genistein synergistically induces apoptosis in human colon cancer ht-29 cells by inhibiting akt phosphorylation and progression of autophagy. Mol. Cancer 2009, 8, 100. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Takano, C.; Shimizu, T.; Yokobe, S.; Abe-Kanoh, N.; Zhu, B.; Nakamura, T.; Munemasa, S.; Murata, Y.; Nakamura, Y. Inhibition of phosphatidylinositide 3-kinase ameliorates antiproliferation by benzyl isothiocyanate in human colon cancer cells. Biochem. Biophys. Res. Commun. 2017, 491, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Tafakh, M.S.; Saidijam, M.; Ranjbarnejad, T.; Malih, S.; Mirzamohammadi, S.; Najafi, R. Sulforaphane, a chemopreventive compound, inhibits cyclooxygenase-2 and microsomal prostaglandin e synthase-1 expression in human ht-29 colon cancer cells. Cells Tissues Organs 2018, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dey, M. Dietary phenethyl isothiocyanate protects mice from colitis associated colon cancer. Int. J. Mol. Sci. 2017, 18, 1908. [Google Scholar] [CrossRef]
- Yano, S.; Wu, S.; Sakao, K.; Hou, D.X. Wasabi 6-(methylsulfinyl)hexyl isothiocyanate induces apoptosis in human colorectal cancer cells through p53-independent mitochondrial dysfunction pathway. BioFactors 2018. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yuan, Y.; Jin, Z.; Xu, T.; Gao, Y.; Wei, H.; Li, C.; Hou, W.; Hua, B. Association between tumor vasculogenic mimicry and the poor prognosis of gastric cancer in china: An updated systematic review and meta-analysis. Biomed. Res. Int. 2016, 2016, 2408645. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, E.J. Review article: Exploring the link between helicobacter pylori and gastric cancer. Aliment. Pharmacol. Ther. 1999, 13 (Suppl. 1), 3–11. [Google Scholar] [CrossRef]
- Zulueta, A.; Caretti, A.; Signorelli, P.; Ghidoni, R. Resveratrol: A potential challenger against gastric cancer. World J. Gastroenterol. 2015, 21, 10636–10643. [Google Scholar] [CrossRef] [PubMed]
- Overby, A.; Zhao, C.M.; Chen, D. Plant phytochemicals: Potential anticancer agents against gastric cancer. Curr. Opin. Pharmacol. 2014, 19, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA. 2002, 99, 7610–7615. [Google Scholar] [CrossRef] [PubMed]
- Overby, A.; Zhao, C.M.; Bones, A.M.; Chen, D. Naturally occurring phenethyl isothiocyanate-induced inhibition of gastric cancer cell growth by disruption of microtubules. J. Gastroenterol. Hepatol. 2014, 29 (Suppl. 4), 99–106. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.D.; Lai, K.C.; Lai, T.Y.; Hsu, S.C.; Kuo, C.L.; Yu, C.S.; Lin, M.L.; Yang, J.S.; Kuo, H.M.; Wu, S.H.; et al. Phenethyl isothiocyanate inhibits migration and invasion of human gastric cancer ags cells through suppressing mapk and nf-kappab signal pathways. Anticancer Res. 2010, 30, 2135–2143. [Google Scholar] [PubMed]
- Ho, C.C.; Lai, K.C.; Hsu, S.C.; Kuo, C.L.; Ma, C.Y.; Lin, M.L.; Yang, J.S.; Chung, J.G. Benzyl isothiocyanate (bitc) inhibits migration and invasion of human gastric cancer ags cells via suppressing erk signal pathways. Hum. Exp. Toxicol. 2011, 30, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, K.; Song, A.; Zhang, X.; Zhang, X.; He, X. Curcumin inhibits proliferation of gastric cancer cells by impairing atp-sensitive potassium channel opening. World J. Surg. Oncol. 2014, 12, 389. [Google Scholar] [CrossRef] [PubMed]
- Da, W.; Zhu, J.; Wang, L.; Sun, Q. Curcumin suppresses lymphatic vessel density in an in vivo human gastric cancer model. Tumour. Biol. 2015, 36, 5215–5223. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhang, K.; Zhang, Q.; Mei, J.; Chen, C.J.; Feng, Z.Z.; Yu, D.H. Effects of quercetin on the apoptosis of the human gastric carcinoma cells. Toxicol. In Vitro 2012, 26, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.Y.; Lin, M.T.; Zhou, M.J.; Yi, T.; Tang, Y.N.; Tang, S.L.; Yang, Z.J.; Zhao, Z.Z.; Chen, H.B. Combinational treatment of curcumin and quercetin against gastric cancer mgc-803 cells in vitro. Molecules 2015, 20, 11524–11534. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Fang, D.; Hang, H.; Tang, Z. The mechanism in gastric cancer chemoprevention by allicin. Anticancer Agents Med. Chem. 2016, 16, 802–809. [Google Scholar] [CrossRef] [PubMed]
- Oser, M.G.; Niederst, M.J.; Sequist, L.V.; Engelman, J.A. Transformation from non-small-cell lung cancer to small-cell lung cancer: Molecular drivers and cells of origin. Lancet Oncol. 2015, 16, e165–172. [Google Scholar] [CrossRef]
- Khan, N.; Mukhtar, H. Dietary agents for prevention and treatment of lung cancer. Cancer Lett. 2015, 359, 155–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Li, C.; Xu, Y.; Wang, L.; Liu, J.; Wang, D.; Hong, C.; Jiang, Z.; Ma, Y.; Chen, Q.; et al. Epigallocatechin gallate promotes p53 accumulation and activity via the inhibition of mdm2-mediated p53 ubiquitination in human lung cancer cells. Oncol. Rep. 2013, 29, 1983–1990. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bian, S.; Yang, C.S. Green tea polyphenol egcg suppresses lung cancer cell growth through upregulating mir-210 expression caused by stabilizing hif-1alpha. Carcinogenesis 2011, 32, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Sadava, D.; Whitlock, E.; Kane, S.E. The green tea polyphenol, epigallocatechin-3-gallate inhibits telomerase and induces apoptosis in drug-resistant lung cancer cells. Biochem. Biophys. Res. Commun. 2007, 360, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Mi, L.; Xiao, Z.; Hood, B.L.; Dakshanamurthy, S.; Wang, X.; Govind, S.; Conrads, T.P.; Veenstra, T.D.; Chung, F.L. Covalent binding to tubulin by isothiocyanates. A mechanism of cell growth arrest and apoptosis. J. Biol. Chem. 2008, 283, 22136–22146. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.N.; Yan, H.Q.; Wu, X.; Pan, Z.H.; Zhu, Y.; Meng, Z.W.; Zhou, Q.H.; Xu, K. Apoptosis induced by benzyl isothiocyanate in gefitinib-resistant lung cancer cells is associated with akt/mapk pathways and generation of reactive oxygen species. Cell Biochem. Biophys. 2013, 66, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Hecht, S.S.; Kenney, P.M.; Wang, M.; Upadhyaya, P. Benzyl isothiocyanate: An effective inhibitor of polycyclic aromatic hydrocarbon tumorigenesis in a/j mouse lung. Cancer Lett. 2002, 187, 87–94. [Google Scholar] [CrossRef]
- Pawlik, A.; Szczepanski, M.A.; Klimaszewska, A.; Gackowska, L.; Zuryn, A.; Grzanka, A. Phenethyl isothiocyanate-induced cytoskeletal changes and cell death in lung cancer cells. Food Chem. Toxicol. 2012, 50, 3577–3594. [Google Scholar] [CrossRef] [PubMed]
- Kalpana Deepa Priya, D.; Gayathri, R.; Gunassekaran, G.R.; Murugan, S.; Sakthisekaran, D. Apoptotic role of natural isothiocyanate from broccoli (brassica oleracea italica) in experimental chemical lung carcinogenesis. Pharm. Biol. 2013, 51, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.Y.; Moon, D.O.; Lee, J.D.; Heo, M.S.; Choi, Y.H.; Lee, C.M.; Park, Y.M.; Kim, G.Y. Sulforaphane sensitizes tumor necrosis factor-related apoptosis-inducing ligand-mediated apoptosis through downregulation of erk and akt in lung adenocarcinoma a549 cells. Carcinogenesis 2007, 28, 1058–1066. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Melkamu, T.; Upadhyaya, P.; Kassie, F. Indole-3-carbinol inhibited tobacco smoke carcinogen-induced lung adenocarcinoma in a/j mice when administered during the post-initiation or progression phase of lung tumorigenesis. Cancer Lett. 2011, 311, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Cho, M.C.; Lee, H.G.; Yoon, D.Y. Indole-3-carbinol induces apoptosis through p53 and activation of caspase-8 pathway in lung cancer a549 cells. Food Chem. Toxicol. 2010, 48, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Dagne, A.; Melkamu, T.; Schutten, M.M.; Qian, X.; Upadhyaya, P.; Luo, X.; Kassie, F. Enhanced inhibition of lung adenocarcinoma by combinatorial treatment with indole-3-carbinol and silibinin in a/j mice. Carcinogenesis 2011, 32, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Melkamu, T.; Zhang, X.; Tan, J.; Zeng, Y.; Kassie, F. Alteration of microrna expression in vinyl carbamate-induced mouse lung tumors and modulation by the chemopreventive agent indole-3-carbinol. Carcinogenesis 2010, 31, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, J.; Li, B.; Wang, Y.; Li, M.; Ma, D.; Wang, X. Genistein exhibits anti-cancer effects via down-regulating foxm1 in h446 small-cell lung cancer cells. Tumour Biol. 2014, 35, 4137–4145. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Cheng, H.; Ren, Y.; Liu, Z.G.; Zhang, Y.F.; De Luo, B. Synergistic inhibitory effects by the combination of gefitinib and genistein on nsclc with acquired drug-resistance in vitro and in vivo. Mol. Biol. Rep. 2012, 39, 4971–4979. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ahmed, F.; Ali, S.; Philip, P.A.; Kucuk, O.; Sarkar, F.H. Inactivation of nuclear factor kappab by soy isoflavone genistein contributes to increased apoptosis induced by chemotherapeutic agents in human cancer cells. Cancer Res. 2005, 65, 6934–6942. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Sharma, A.; Sak, K.; Tuli, H.S.; Buttar, H.S.; Bishayee, A. Fisetin: A bioactive phytochemical with potential for cancer prevention and pharmacotherapy. Life Sci 2018, 194, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Afaq, F.; Khusro, F.H.; Mustafa Adhami, V.; Suh, Y.; Mukhtar, H. Dual inhibition of phosphatidylinositol 3-kinase/akt and mammalian target of rapamycin signaling in human nonsmall cell lung cancer cells by a dietary flavonoid fisetin. Int. J. Cancer 2012, 130, 1695–1705. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.C.; Shih, Y.W.; Chao, C.H.; Lee, X.Y.; Chiang, T.A. Involvement of the erk signaling pathway in fisetin reduces invasion and migration in the human lung cancer cell line a549. J. Agric. Food Chem. 2009, 57, 8933–8941. [Google Scholar] [CrossRef] [PubMed]
- Ravichandran, N.; Suresh, G.; Ramesh, B.; Manikandan, R.; Choi, Y.W.; Vijaiyan Siva, G. Fisetin modulates mitochondrial enzymes and apoptotic signals in benzo(a)pyrene-induced lung cancer. Mol. Cell Biochem. 2014, 390, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Touil, Y.S.; Seguin, J.; Scherman, D.; Chabot, G.G. Improved antiangiogenic and antitumour activity of the combination of the natural flavonoid fisetin and cyclophosphamide in lewis lung carcinoma-bearing mice. Cancer Chemother. Pharmacol. 2011, 68, 445–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turrini, E.; Ferruzzi, L.; Fimognari, C. Potential effects of pomegranate polyphenols in cancer prevention and therapy. Oxid. Med. Cell Longev. 2015, 2015, 938475. [Google Scholar] [CrossRef] [PubMed]
- Zahin, M.; Ahmad, I.; Gupta, R.C.; Aqil, F. Punicalagin and ellagic acid demonstrate antimutagenic activity and inhibition of benzo[a]pyrene induced DNA adducts. Biomed. Res. Int. 2014, 2014, 467465. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Hadi, N.; Afaq, F.; Syed, D.N.; Kweon, M.H.; Mukhtar, H. Pomegranate fruit extract inhibits prosurvival pathways in human a549 lung carcinoma cells and tumor growth in athymic nude mice. Carcinogenesis 2007, 28, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Afaq, F.; Kweon, M.H.; Kim, K.; Mukhtar, H. Oral consumption of pomegranate fruit extract inhibits growth and progression of primary lung tumors in mice. Cancer Res. 2007, 67, 3475–3482. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wei, C.; Xi, Z. Curcumin suppresses proliferation and invasion in non-small cell lung cancer by modulation of mta1-mediated wnt/beta-catenin pathway. In Vitro Cell. Dev. Biol. Anim. 2014, 50, 840–850. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Jiang, J.; Guan, C.; Dong, C.; Wang, G.; Bai, L.; Sun, J.; Hu, C.; Bai, C. Curcumin induces autophagy via activating the ampk signaling pathway in lung adenocarcinoma cells. J. Pharmacol. Sci. 2013, 123, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Liu, Z.; Zhu, F.; Fan, X.; Wu, X.; Zhao, H.; Jiang, L. Curcumin lowers erlotinib resistance in non-small cell lung carcinoma cells with mutated egf receptor. Oncol. Res. 2013, 21, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Liu, Y.Y.; Ma, Y.G.; Xue, Y.X.; Liu, D.G.; Ren, Y.; Liu, X.B.; Li, Y.; Li, Z. Curcumin blocks small cell lung cancer cells migration, invasion, angiogenesis, cell cycle and neoplasia through janus kinase-stat3 signalling pathway. PLoS ONE 2012, 7, e37960. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Ma, Y.G.; Xue, Y.X.; Liu, Y.Y.; Xie, H.; Qiu, G.R. Curcumin induces small cell lung cancer nci-h446 cell apoptosis via the reactive oxygen species-mediated mitochondrial pathway and not the cell death receptor pathway. DNA Cell Biol. 2012, 31, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Sauter, E.R. Breast cancer prevention: Current approaches and future directions. Eur. J. Breast Health 2018, 14, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Chemistry and anticarcinogenic mechanisms of glycoalkaloids produced by eggplants, potatoes, and tomatoes. J. Agric. Food Chem. 2015, 63, 3323–3337. [Google Scholar] [CrossRef] [PubMed]
- Yelken, B.O.; Balci, T.; Susluer, S.Y.; Kayabasi, C.; Avci, C.B.; Kirmizibayrak, P.B.; Gunduz, C. The effect of tomatine on metastasis related matrix metalloproteinase (mmp) activities in breast cancer cell model. Gene 2017, 627, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Caruso, J.A.; Campana, R.; Wei, C.; Su, C.H.; Hanks, A.M.; Bornmann, W.G.; Keyomarsi, K. Indole-3-carbinol and its n-alkoxy derivatives preferentially target eralpha-positive breast cancer cells. Cell Cycle 2014, 13, 2587–2599. [Google Scholar] [CrossRef] [PubMed]
- Tin, A.S.; Park, A.H.; Sundar, S.N.; Firestone, G.L. Essential role of the cancer stem/progenitor cell marker nucleostemin for indole-3-carbinol anti-proliferative responsiveness in human breast cancer cells. BMC Biol. 2014, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Bishayee, A.; Ahmed, S.; Brankov, N.; Perloff, M. Triterpenoids as potential agents for the chemoprevention and therapy of breast cancer. Front. Biosci. (Landmark Ed.) 2011, 16, 980–996. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasam, B.; Seeram, N.P.; Nair, M.G. Anticancer and antiinflammatory activities of cucurbitacins from cucurbita andreana. Cancer Lett. 2003, 189, 11–16. [Google Scholar] [CrossRef]
- Ramalhete, C.; Mansoor, T.A.; Mulhovo, S.; Molnar, J.; Ferreira, M.J. Cucurbitane-type triterpenoids from the african plant momordica balsamina. J. Nat. Prod. 2009, 72, 2009–2013. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, N.; Yin, D.; O’Kelly, J.; Haritunians, T.; Karlan, B.; Said, J.; Xing, H.; Koeffler, H.P. Cucurbitacin b has a potent antiproliferative effect on breast cancer cells in vitro and in vivo. Cancer Sci. 2008, 99, 1793–1797. [Google Scholar] [CrossRef] [PubMed]
- Kongtun, S.; Jiratchariyakul, W.; Kummalue, T.; Tan-ariya, P.; Kunnachak, S.; Frahm, A.W. Cytotoxic properties of root extract and fruit juice of trichosanthes cucumerina. Planta Med. 2009, 75, 839–842. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, N.; Vasquez, Y.; Hussein, A.A.; Coley, P.D.; Solis, P.N.; Gupta, M.P. Cytotoxic cucurbitacin constituents from sloanea zuliaensis. J. Nat. Prod. 2003, 66, 1515–1516. [Google Scholar] [CrossRef] [PubMed]
- Blaskovich, M.A.; Sun, J.; Cantor, A.; Turkson, J.; Jove, R.; Sebti, S.M. Discovery of jsi-124 (cucurbitacin i), a selective janus kinase/signal transducer and activator of transcription 3 signaling pathway inhibitor with potent antitumor activity against human and murine cancer cells in mice. Cancer Res. 2003, 63, 1270–1279. [Google Scholar] [PubMed]
- Sun, J.; Blaskovich, M.A.; Jove, R.; Livingston, S.K.; Coppola, D.; Sebti, S.M. Cucurbitacin q: A selective stat3 activation inhibitor with potent antitumor activity. Oncogene 2005, 24, 3236–3245. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, S.; Zhang, Q.; Liu, F.; Wu, P. 23,24-dihydrocucurbitacin b induces g2/m cell-cycle arrest and mitochondria-dependent apoptosis in human breast cancer cells (bcap37). Cancer Lett. 2007, 256, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Phongmaykin, J.; Kumamoto, T.; Ishikawa, T.; Suttisri, R.; Saifah, E. A new sesquiterpene and other terpenoid constituents of chisocheton penduliflorus. Arch. Pharm. Res. 2008, 31, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.C.; Chan, M.L.; Chen, W.Y.; Tsai, C.Y.; Chang, F.R.; Wu, Y.C. Pristimerin induces caspase-dependent apoptosis in mda-mb-231 cells via direct effects on mitochondria. Mol. Cancer Ther. 2005, 4, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Idris, A.I.; Libouban, H.; Nyangoga, H.; Landao-Bassonga, E.; Chappard, D.; Ralston, S.H. Pharmacologic inhibitors of ikappab kinase suppress growth and migration of mammary carcinosarcoma cells in vitro and prevent osteolytic bone metastasis in vivo. Mol. Cancer Ther. 2009, 8, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Gu, Z.M.; Chang, C.J.; Wood, K.V.; McLaughlin, J.L. Meliavolkenin, a new bioactive triterpenoid from melia volkensii (meliaceae). Bioorg. Med. Chem. 1995, 3, 383–390. [Google Scholar] [CrossRef]
- Amico, V.; Barresi, V.; Condorelli, D.; Spatafora, C.; Tringali, C. Antiproliferative terpenoids from almond hulls (prunus dulcis): Identification and structure-activity relationships. J. Agric. Food Chem. 2006, 54, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Rzeski, W.; Stepulak, A.; Szymanski, M.; Sifringer, M.; Kaczor, J.; Wejksza, K.; Zdzisinska, B.; Kandefer-Szerszen, M. Betulinic acid decreases expression of bcl-2 and cyclin d1, inhibits proliferation, migration and induces apoptosis in cancer cells. Naunyn Schmiedebergs Arch. Pharmacol. 2006, 374, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Kessler, J.H.; Mullauer, F.B.; de Roo, G.M.; Medema, J.P. Broad in vitro efficacy of plant-derived betulinic acid against cell lines derived from the most prevalent human cancer types. Cancer Lett. 2007, 251, 132–145. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, E.; Lampronti, I.; Penolazzi, L.; Khan, M.T.; Ather, A.; Giorgi, G.; Gambari, R.; Piva, R. Expression of estrogen receptor alpha gene in breast cancer cells treated with transcription factor decoy is modulated by bangladeshi natural plant extracts. Oncol. Res. 2005, 15, 69–79. [Google Scholar] [PubMed]
- Es-Saady, D.; Simon, A.; Jayat-Vignoles, C.; Chulia, A.J.; Delage, C. Mcf-7 cell cycle arrested at g1 through ursolic acid, and increased reduction of tetrazolium salts. Anticancer Res. 1996, 16, 481–486. [Google Scholar] [PubMed]
- Chen, Y.H.; Chang, F.R.; Wu, C.C.; Yen, M.H.; Liaw, C.C.; Huang, H.C.; Kuo, Y.H.; Wu, Y.C. New cytotoxic 6-oxygenated 8,9-dihydrofurocoumarins, hedyotiscone A-C, from hedyotis biflora. Planta Med. 2006, 72, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Martin-Cordero, C.; Reyes, M.; Ayuso, M.J.; Toro, M.V. Cytotoxic triterpenoids from erica andevalensis. Z. Naturforsch. C 2001, 56, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Neto, C.C.; Vaisberg, A.J.; Zhou, B.N.; Kingston, D.G.; Hammond, G.B. Cytotoxic triterpene acids from the peruvian medicinal plant polylepis racemosa. Planta Med. 2000, 66, 483–484. [Google Scholar] [CrossRef] [PubMed]
- Kassi, E.; Sourlingas, T.G.; Spiliotaki, M.; Papoutsi, Z.; Pratsinis, H.; Aligiannis, N.; Moutsatsou, P. Ursolic acid triggers apoptosis and bcl-2 downregulation in mcf-7 breast cancer cells. Cancer Investig. 2009, 27, 723–733. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.T.; Wu, C.H.; Yen, G.C. Ursolic acid, a naturally occurring triterpenoid, suppresses migration and invasion of human breast cancer cells by modulating c-jun n-terminal kinase, akt and mammalian target of rapamycin signaling. Mol. Nutr. Food Res. 2010, 54, 1285–1295. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.L.; Kuo, P.L.; Lin, L.T.; Lin, C.C. Asiatic acid, a triterpene, induces apoptosis and cell cycle arrest through activation of extracellular signal-regulated kinase and p38 mitogen-activated protein kinase pathways in human breast cancer cells. J. Pharmacol. Exp. Ther. 2005, 313, 333–344. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Jiang, T.Y.; Sheng, S.; Tianasoa-Ramamonjy, M.; Snyder, J.K. Remangilones a-c, new cytotoxic triterpenes from physena madagascariensis. J. Nat. Prod 1999, 62, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Rabi, T.; Karunagaran, D.; Krishnan Nair, M.; Bhattathiri, V.N. Cytotoxic activity of amooranin and its derivatives. Phytother. Res. 2002, 16 (Suppl. 1), S84–S86. [Google Scholar] [CrossRef]
- Rabi, T.; Ramachandran, C.; Fonseca, H.B.; Nair, R.P.; Alamo, A.; Melnick, S.J.; Escalon, E. Novel drug amooranin induces apoptosis through caspase activity in human breast carcinoma cell lines. Breast Cancer Res. Treat. 2003, 80, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Rabi, T.; Wang, L.; Banerjee, S. Novel triterpenoid 25-hydroxy-3-oxoolean-12-en-28-oic acid induces growth arrest and apoptosis in breast cancer cells. Breast Cancer Res. Treat. 2007, 101, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Chavez, I.O.; Apan, T.R.; Martinez-Vazquez, M. Cytotoxic activity and effect on nitric oxide production of tirucallane-type triterpenes. J. Pharm. Pharmacol. 2005, 57, 1087–1091. [Google Scholar] [CrossRef] [PubMed]
- Lavhale, M.S.; Kumar, S.; Mishra, S.H.; Sitasawad, S.L. A novel triterpenoid isolated from the root bark of ailanthus excelsa roxb (tree of heaven), aechl-1 as a potential anti-cancer agent. PLoS ONE 2009, 4, e5365. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, W.; Xie, C.; Huang, C.; Zhu, J.; Liang, Z.; Deng, F.; Zhu, M.; Zhu, W.; Wu, R.; et al. Curcumin modulates mir-19/pten/akt/p53 axis to suppress bisphenol a-induced mcf-7 breast cancer cell proliferation. Phytother. Res. 2014, 28, 1553–1560. [Google Scholar] [CrossRef] [PubMed]
- Miksits, M.; Wlcek, K.; Svoboda, M.; Kunert, O.; Haslinger, E.; Thalhammer, T.; Szekeres, T.; Jager, W. Antitumor activity of resveratrol and its sulfated metabolites against human breast cancer cells. Planta Med. 2009, 75, 1227–1230. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lu, C.K.; Tu, M.C.; Chang, J.H.; Chen, Y.J.; Tu, Y.H.; Huang, H.C. Polyphenol-rich avicennia marina leaf extracts induce apoptosis in human breast and liver cancer cells and in a nude mouse xenograft model. Oncotarget 2016, 7, 35874–35893. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Maycotte, P.; Acosta-Casique, A.; Rodriguez-Rodriguez, S.; Moreno, D.A.; Ferreres, F.; Flores-Alonso, J.C.; Delgado-Lopez, M.G.; Perez-Santos, M.; Anaya-Ruiz, M. Bursera copallifera extracts have cytotoxic and migration-inhibitory effects in breast cancer cell lines. Integr. Cancer Ther. 2018. [Google Scholar] [CrossRef] [PubMed]
- G, W.W.; L, M.B.; D, E.W.; R, H.D.; Ho, E. Phytochemicals from cruciferous vegetables, epigenetics, and prostate cancer prevention. AAPS J. 2013, 15, 951–961. [Google Scholar]
- Wu, W.; Tang, S.N.; Zhang, Y.; Puppala, M.; Cooper, T.K.; Xing, C.; Jiang, C.; Lu, J. Prostate cancer xenograft inhibitory activity and pharmacokinetics of decursinol, a metabolite of angelica gigas pyranocoumarins, in mouse models. Am. J. Chin. Med. 2017, 45, 1773–1792. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.N.; Zhang, J.; Wu, W.; Jiang, P.; Puppala, M.; Zhang, Y.; Xing, C.; Kim, S.H.; Jiang, C.; Lu, J. Chemopreventive effects of korean angelica versus its major pyranocoumarins on two lineages of transgenic adenocarcinoma of mouse prostate carcinogenesis. Cancer Prev. Res. (Phila) 2015, 8, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Empl, M.T.; Albers, M.; Wang, S.; Steinberg, P. The resveratrol tetramer r-viniferin induces a cell cycle arrest followed by apoptosis in the prostate cancer cell line lncap. Phytother. Res. 2015, 29, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, E.; Adhami, V.M.; Sechi, M.; Mukhtar, H. Dietary flavonoid fisetin binds to beta-tubulin and disrupts microtubule dynamics in prostate cancer cells. Cancer Lett. 2015, 367, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Pavese, J.M.; Krishna, S.N.; Bergan, R.C. Genistein inhibits human prostate cancer cell detachment, invasion, and metastasis. Am. J. Clin. Nutr. 2014, 100 (Suppl. 1), 431S–436S. [Google Scholar] [CrossRef]
- Du, Y.; Long, Q.; Zhang, L.; Shi, Y.; Liu, X.; Li, X.; Guan, B.; Tian, Y.; Wang, X.; Li, L.; et al. Curcumin inhibits cancer-associated fibroblast-driven prostate cancer invasion through maoa/mtor/hif-1alpha signaling. Int. J. Oncol. 2015, 47, 2064–2072. [Google Scholar] [CrossRef] [PubMed]
- Dorai, T.; Diouri, J.; O’Shea, O.; Doty, S.B. Curcumin inhibits prostate cancer bone metastasis by up-regulating bone morphogenic protein-7 in vivo. J. Cancer Ther. 2014, 5, 369–386. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Ding, N.; Du, Z.Y.; Cui, X.X.; Wang, H.; Wei, X.C.; Conney, A.H.; Zhang, K.; Zheng, X. Curcumin analogues with high activity for inhibiting human prostate cancer cell growth and androgen receptor activation. Mol. Med. Rep. 2014, 10, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.; Xiao, W.; Abdel-Wahab, O. Diagnosis and classification of hematologic malignancies on the basis of genetics. Blood 2017, 130, 410–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jendzelovska, Z.; Jendzelovsky, R.; Hilovska, L.; Koval, J.; Mikes, J.; Fedorocko, P. Single pre-treatment with hypericin, a st. John’s wort secondary metabolite, attenuates cisplatin- and mitoxantrone-induced cell death in a2780, a2780cis and hl-60 cells. Toxicol. In Vitro 2014, 28, 1259–1273. [Google Scholar] [CrossRef] [PubMed]
- Azmi, A.S.; Bhat, S.H.; Hanif, S.; Hadi, S.M. Plant polyphenols mobilize endogenous copper in human peripheral lymphocytes leading to oxidative DNA breakage: A putative mechanism for anticancer properties. FEBS Lett. 2006, 580, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Estrov, Z.; Shishodia, S.; Faderl, S.; Harris, D.; Van, Q.; Kantarjian, H.M.; Talpaz, M.; Aggarwal, B.B. Resveratrol blocks interleukin-1beta-induced activation of the nuclear transcription factor nf-kappab, inhibits proliferation, causes s-phase arrest, and induces apoptosis of acute myeloid leukemia cells. Blood 2003, 102, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Noda, C.; He, J.; Takano, T.; Tanaka, C.; Kondo, T.; Tohyama, K.; Yamamura, H.; Tohyama, Y. Induction of apoptosis by epigallocatechin-3-gallate in human lymphoblastoid b cells. Biochem. Biophys. Res. Commun. 2007, 362, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Safa, M.; Tavasoli, B.; Manafi, R.; Kiani, F.; Kashiri, M.; Ebrahimi, S.; Kazemi, A. Indole-3-carbinol suppresses nf-kappab activity and stimulates the p53 pathway in pre-b acute lymphoblastic leukemia cells. Tumour Biol. 2015, 36, 3919–3930. [Google Scholar] [CrossRef] [PubMed]
- Linares, M.A.; Zakaria, A.; Nizran, P. Skin cancer. Prim. Care 2015, 42, 645–659. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Pao, J.; Lin, S.Y.; Sheen, L.Y. Molecular mechanisms of garlic-derived allyl sulfides in the inhibition of skin cancer progression. Ann. N. Y. Acad. Sci. 2012, 1271, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Nigam, N.; Shukla, Y. Preventive effects of diallyl sulfide on 7,12-dimethylbenz[a]anthracene induced DNA alkylation damage in mouse skin. Mol. Nutr. Food Res. 2007, 51, 1324–1328. [Google Scholar] [CrossRef] [PubMed]
- Arora, A.; Shukla, Y. Induction of apoptosis by diallyl sulfide in dmba-induced mouse skin tumors. Nutr. Cancer 2002, 44, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Kalra, N.; Arora, A.; Shukla, Y. Involvement of multiple signaling pathways in diallyl sulfide mediated apoptosis in mouse skin tumors. Asian Pac. J. Cancer Prev. 2006, 7, 556–562. [Google Scholar] [PubMed]
- Chang, H.P.; Sheen, L.Y.; Lei, Y.P. The protective role of carotenoids and polyphenols in patients with head and neck cancer. J. Chin. Med. Assoc. 2015, 78, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Beta-carotene. In Drugs and Lactation Database (LactMed). Bethesda (MD). Available online: www.ncbi.nlm.nih.gov/books/NBK501922/ (accessed on 22 July 2018).
- Zhang, Y.; Zhu, X.; Huang, T.; Chen, L.; Liu, Y.; Li, Q.; Song, J.; Ma, S.; Zhang, K.; Yang, B.; et al. Beta-carotene synergistically enhances the anti-tumor effect of 5-fluorouracil on esophageal squamous cell carcinoma in vivo and in vitro. Toxicol. Lett. 2016, 261, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Suzui, M.; Weinstein, I.B. Effects of epigallocatechin-3-gallate on growth, epidermal growth factor receptor signaling pathways, gene expression, and chemosensitivity in human head and neck squamous cell carcinoma cell lines. Clin. Cancer Res. 2001, 7, 4220–4229. [Google Scholar] [PubMed]
- Amin, A.R.; Khuri, F.R.; Chen, Z.G.; Shin, D.M. Synergistic growth inhibition of squamous cell carcinoma of the head and neck by erlotinib and epigallocatechin-3-gallate: The role of p53-dependent inhibition of nuclear factor-kappab. Cancer Prev. Res. (Phila) 2009, 2, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.C.; Yang, S.F.; Peng, C.Y.; Chou, M.Y.; Chang, Y.C. Epigallocatechin-3-gallate inhibits the invasion of human oral cancer cells and decreases the productions of matrix metalloproteinases and urokinase-plasminogen activator. J. Oral. Pathol. Med. 2007, 36, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Class | Active Metabolite | Structure |
|---|---|---|
| Phenolic compounds | Curcumin |  |
| Decursin |  | |
| Decursinol |  | |
| Decursinol angelate |  | |
| Dichamanetin |  | |
| Epigallocatechin gallate (EGCG) |  | |
| Fisetin |  | |
| Genistein |  | |
| Hydroxycinnamic acid |  | |
| Hypericin |  | |
| Quercetin |  | |
| Phenolic compounds | Resveratrol |  |
| Punicalagin (PC) |  | |
| γ-viniferin |  | |
| Terpenoids | Asiatic acid |  |
| Ailanthus excelsa chloroform extract-1 (AECHL-1) |  | |
| Amooranin (AMR) |  | |
| Auraptene (AUR) |  | |
| Betulinic acid (BA) |  | |
| Celastrol |  | |
| Curcubitane-type triterpenoids (Balsaminapentaol) |  | |
| Dammarane triterpenoid (Cabraleadiol) |  | |
| Lupeol |  | |
| Meliavolkenin |  | |
| Pomolic acid |  | |
| Pristimerin |  | |
| Remangilones A |  | |
| Remangilones C |  | |
| Tirucallane-type triterpenoids |  | |
| Ursolic acid |  | |
| β-carotene |  | |
| Nitrogen-containing alkaloids & sulfur-containing compounds | Allicin |  |
| Benzyl isothiocyanate (BITC) |  | |
| Diallyl sulfide (DAS) |  | |
| Indole-3-carbinol (I3C) |  | |
| Phenethyl isothiocyanate (PEITC) |  | |
| Sulforaphane (SFN) |  | |
| Tomatine |  | |
| 6-MSITC |  |
| Type of Cancer | Active Metabolites | In Vitro or In Vivo Effects | IC50 & Effective Concentration (EC) (μM). | Ref. |
|---|---|---|---|---|
| Colorectal cancer | Dichamanetin | Induction of ROS and cell cycle arrest in HT-29 colon cancer cells | IC50: 13.8 | [80] |
| GAP | Suppression of colon carcinogenesis in DSS mice | EC: 0.01 % or 0.05 % in diet | [83] | |
| Auraptene | Inhibition of the growth of colon cancer cells and suppression of colonosphere formation | EC: 10 | [85] | |
| Curcumin | Suppression of the oncogenicity of human colon cancer cells and the growth of HCT-116 tumor xenografts | EC: 10 | [87] | |
| Genistein | Inhibition of cell proliferation and induction of apoptosis in HCT 116 and LoVo cells | EC: 135 | [89,90,91] | |
| Inhibition of metastasis in colorectal cancer cell implanted nude mice | ||||
| I3C+Genistein | Induction of apoptosis in HT 29 colon cancer cells | I3C EC: 300 Genistein EC: 40 | [92] | |
| BITC | Suppression of viability in HCT 116 colon cancer cells | EC: 5–20 | [93] | |
| SFN | Induction of apoptosis and inhibition of proliferation in HT 29 colon cancer cells | EC: 5–20 | [94] | |
| PEITC | Reduction of colon carcinogenesis in AOM/DSS induced mice | EC: 0.12 % in diet | [95] | |
| 6-MSITC | Induction of apoptosis in HCT 116 colon cancer cells | IC50: 0.92–10.01 | [96] | |
| Gastric cancer | Resveratrol | Inhibition of proliferation in gastric cancer cells | EC: 50–200 | [99] |
| SFN | Prevention of benzo[a]pyrene-induced stomach tumors in mice | EC: 1.33 mg per mouse | [101] | |
| PEITC | Induction of cell cycle arrest and apoptosis in gastric cancer cells MKN74 and Kato-III | EC: 17.8 | [102,103] | |
| Inhibition of migration and invasion in AGS gastric cancer cells | ||||
| BITC | Inhibition of migration and invasion in AGS gastric cancer cells | EC: 0.25–0.5 | [104] | |
| Curcumin | Inhibition of proliferation in SGC-7901 gastric cancer cells | EC: 15–60 | [105,106] | |
| Reduction of xenograft tumor growth in mice | ||||
| Reduction of LVD in gastric cancer bearing nude mice | ||||
| Quercetin | Induction of apoptosis in BGC-823 gastric cancer cells | EC: 30–120 | [107] | |
| Allicin | Inhibition of gastric cancer cell growth | EC: 184.88 | [109] | |
| Lung cancer | EGCG | Induction of cell cycle arrest and apoptosis in lung cancer cells Reduction of proliferation and growth in lung cancer cells Inhibition of TGF-β-induced cell migration, invasion, and EMT in NSCLC cells | IC50: 70 | [112,113,114] |
| BITC | Inhibition of growth in A549 lung cancer cells | EC: 10 | [115,116,117] | |
| Inhibition of tumorigenesis in PAH-induced A/J mice | ||||
| PEITC | Induction of apoptosis in NSCLC cells | EC: 12.5–20 | [113,118] | |
| SFN | Induction of apoptosis in NSCLC cells | EC: 10 | [119,120] | |
| Alleviation of carcinogenic lung in B(a)P induced lung cancer bearing mice | ||||
| I3C | Induction of apoptosis in A549 lung adenocarcinoma cells in combination with TRAIL | EC: 100–500 | [121,122,123,124] | |
| Inhibition of progression of tobacco carcinogen induced lung adenocarcinoma progression | ||||
| Induction of cell cycle arrest and apoptosis in A549 lung cancer cells | ||||
| Inhibition of NNK-induced lung tumors in combination with silibinin in mice | ||||
| Genistein | Inhibition of carcinogenesis in mice with VC-induced lung cancer Inhibition of cell proliferation and induction of apoptosis in H446 SCLC cells | IC50: 81 | [125,126,127] | |
| Inhibition of cell proliferation and induction of apoptosis in combination with gefitinib in H1975 NSCLC cells | ||||
| Fisetin | Inhibition of cell growth and induction of apoptosis in combination with chemotherapeutic agents in H460 NSCLC cells | IC50: 59 | [129,130,131,132] | |
| Inhibition of cell viability and colony-forming activity in A549 NSCLC cells | ||||
| Inhibition of the invasion and migration of A549 NSCLC cells | ||||
| Inhibition of lung carcinogenesis in B(a)P-induced mice | ||||
| Inhibition of angiogenesis and tumor growth in Lewis lung carcinoma bearing mice | ||||
| Punicalagin | Anti-proliferative effects on A549 and H1299 NSCLC cells Inhibition of tumor growth in mice with xenografts of A549 NSCLC cells Inhibition of B(a)P-induced tumorigenesis in A/J mice | EC: 11.52–184.3 | [134,135,136] | |
| Curcumin | Inhibition of cell growth and invasion in NSCLC cells Lowering the resistance of NSCLC cells against erlotinib Suppression of cell proliferation, the cell cycle, migration, invasion, and angiogenesis in SCLC cells Induction of apoptosis in SCLC cells | EC: 30 | [137,138,139,140,141] | |
| Breast cancer | Tomatine | Induction of cell cytotoxicity and apoptosis in MCF-7 breast cancer cells | IC50: 7.07 | [144] |
| I3C | Increasing apoptotic cell death and decreasing the proliferation of the ERα-positive breast cancer cells Disruption of in vitro 10AT-Her2 cell tumorsphere formation and in vivo tumor xenograft growth | IC50: 204 | [145,146] | |
| Curcubitane-type triterpenoids | Inhibition of cell growth and induction of apoptosis in human breast cancer cells | EC: 0.5–35.7 | [148,149,150,151,152,153,154,155] | |
| Dammarane triterpenoids | Cytotoxicity against breast cancer cells | EC: 20.97 | [156] | |
| Pristimerin | Induction of apoptosis in MDA-MB-231 breast cancer cells | EC: 1–3 | [157] | |
| Celastrol | Inhibition of cell growth and invasion and induction of apoptosis in W256 breast cancer cells | EC: 1 | [158] | |
| Meliavolkenin | Cytotoxicity against MCF7 breast cancer cells | EC: 6.05 | [159] | |
| Betulinic acid | Induction of anti-proliferation in MCF7 and T47D breast cancer cells | IC50: 2.4 | [160,161,162] | |
| Lupeol | Inhibition of MDA-MB-231 ERα-negative cell proliferation | EC: 1–30 | [163] | |
| Ursolic acid | Inhibition of proliferation and induction of apoptosis in MCF7 cells Suppression of migration and invasion in MDA-MB-231 cells | IC50: 3.26 | [164,165,166,167,168,169] | |
| Asiatic acid | Inhibition of cell growth and induction of apoptosis in MCF7 and MDA-MB 231 cells | IC50: 5.95–8.12 | [170] | |
| Remangilones A and C | Cytotoxicity against MDA-MB-231 and MDA-MB-435 cells | RemangilonesA IC50: 6.6–8.5 RemangilonesC IC50: 1.6–2.0 | [171] | |
| Amooranin | Induction of apoptosis and suppression of cell growth in MDA-468 and MCF7 cells | IC50: 3.82-7.22 | [172,173,174] | |
| Tirucallane-type triterpenoids | Cytotoxicity against MCF7 cells | IC50: 41.33–86.14 | [175] | |
| AECHL-1 | Regression of MCF7 xenograft tumors in nude mice | EC: 5–100 | [176] | |
| Curcumin | Anti-proliferation of BPA-induced MCF7 cells | EC: 1 | [177] | |
| Resveratrol | Reduction of cell viability in breast cancer cells (MCF-7, ZR-75-1, and MDA-MB-231) | IC50: 67.6–82.2 | [178] | |
| Hydroxycinnamic acid | Inhibition of migration in MCF-7 and MDA-MB-231 cells | IC50: 75.71 | [180] | |
| Dichamanetin | Induction of ROS and cell cycle arrest in MDA-MB-231 cells | EC: 8.7 | [80] | |
| Prostate cancer | Decursinol | Suppression of tumor growth in mice with xenografted DU145 and PC3 prostate cancers | EC: 4.5 mg per mouse | [182] |
| Decursin & Decursinol angelate | Inhibition of prostate epithelium growth in the TRAMP model | EC: 3 mg per mouse | [183] | |
| Resveratrol & γ-viniferin | Inhibition of the growth of LNCaP prostate cancer cell | Resveratrol IC50: 10.23-228.3 γ-viniferin IC50: 8.93–90.1 | [184] | |
| Fisetin | Inhibition of cell growth and proliferation in PU3 and DU145 cells | EC: 20–80 | [185] | |
| Genistein | Inhibition of cellular invasion in in vitro prostate cancer and in vivo metastasis formation in mice with xenografts of PC3-M prostate cancer | EC: 10 | [186] | |
| Curcumin | Inhibition of CAF-induced EMT and invasion in PC3 cells Induction of cell cycle arrest and apoptosis in in vitro prostate cancer cells and the in vivo TRAMP model | EC: 25 | [187,188,189] | |
| SFN and I3C | Induction of cell cycle arrest and apoptosis of PC3, LNCaP, and DU145 cells in vitro | SFN EC: 40 I3C EC: 30–100 | [181] | |
| Hematologic cancer | Hypericin | Attenuation of MTX cytotoxicity in HL-60 promyelocytic leukemia cells | EC: 0.1–0.5 | [191] |
| Resveratrol | Induction of DNA breakage in human peripheral lymphocytes Induction of apoptosis in OCI-ANK3 and OCIM2 acute myeloid leukemia cells | EC: 10–75 | [192,193] | |
| EGCG | Induction of apoptotic death in Ramos B lymphoblastoid cells | EC: 60–100 | [194] | |
| I3C | Inhibition of cell growth and induction of apoptosis in pre-B acute lymphoblastic leukemia cells | EC: 60 | [195] | |
| Skin cancer | Diallyl sulfide | Reduction of DNA strand breaks in DMBA induced mouse skin Induction of apoptosis in DMBA-induced mouse skin tumors | EC: 25 | [197,198,199,200] |
| Pomolic acid | Cytotoxic effects against M-14 melanoma cells | EC: 14.6 | [167] | |
| Head and neck cancer | β-carotene | Inhibition of tumor growth in nude mice with xenografts of Eca109 ESCC cell xenografts | EC: 30 | [203] |
| EGCG | Induction of cell cycle arrest and apoptosis in YCU-N861 and YCU-H891 HNSCC cells Inhibition of cell growth in combination with erlotinib in HNSCC cells Inhibition of the invasion and migration in oral cancer cell OC2 | EC: 30–60 | [204,205,206] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, S.-A.; Moon, S.Y.; Kim, W.-Y.; Paek, S.-M.; Park, H.H.; Lee, C.S. Structure-Based Classification and Anti-Cancer Effects of Plant Metabolites. Int. J. Mol. Sci. 2018, 19, 2651. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092651
Shin S-A, Moon SY, Kim W-Y, Paek S-M, Park HH, Lee CS. Structure-Based Classification and Anti-Cancer Effects of Plant Metabolites. International Journal of Molecular Sciences. 2018; 19(9):2651. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092651
Chicago/Turabian StyleShin, Seong-Ah, Sun Young Moon, Woe-Yeon Kim, Seung-Mann Paek, Hyun Ho Park, and Chang Sup Lee. 2018. "Structure-Based Classification and Anti-Cancer Effects of Plant Metabolites" International Journal of Molecular Sciences 19, no. 9: 2651. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092651