Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1

 , ,
, ,  and
and 
Abstract
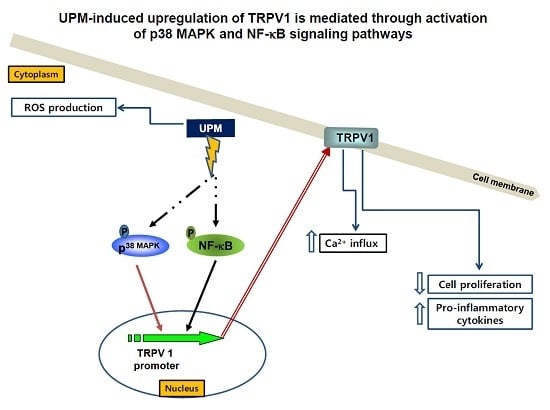
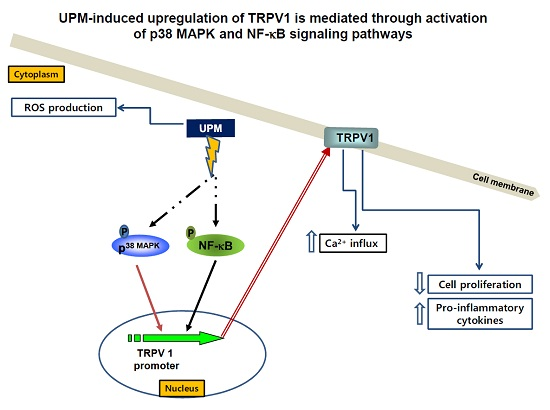
:
1. Introduction
2. Results
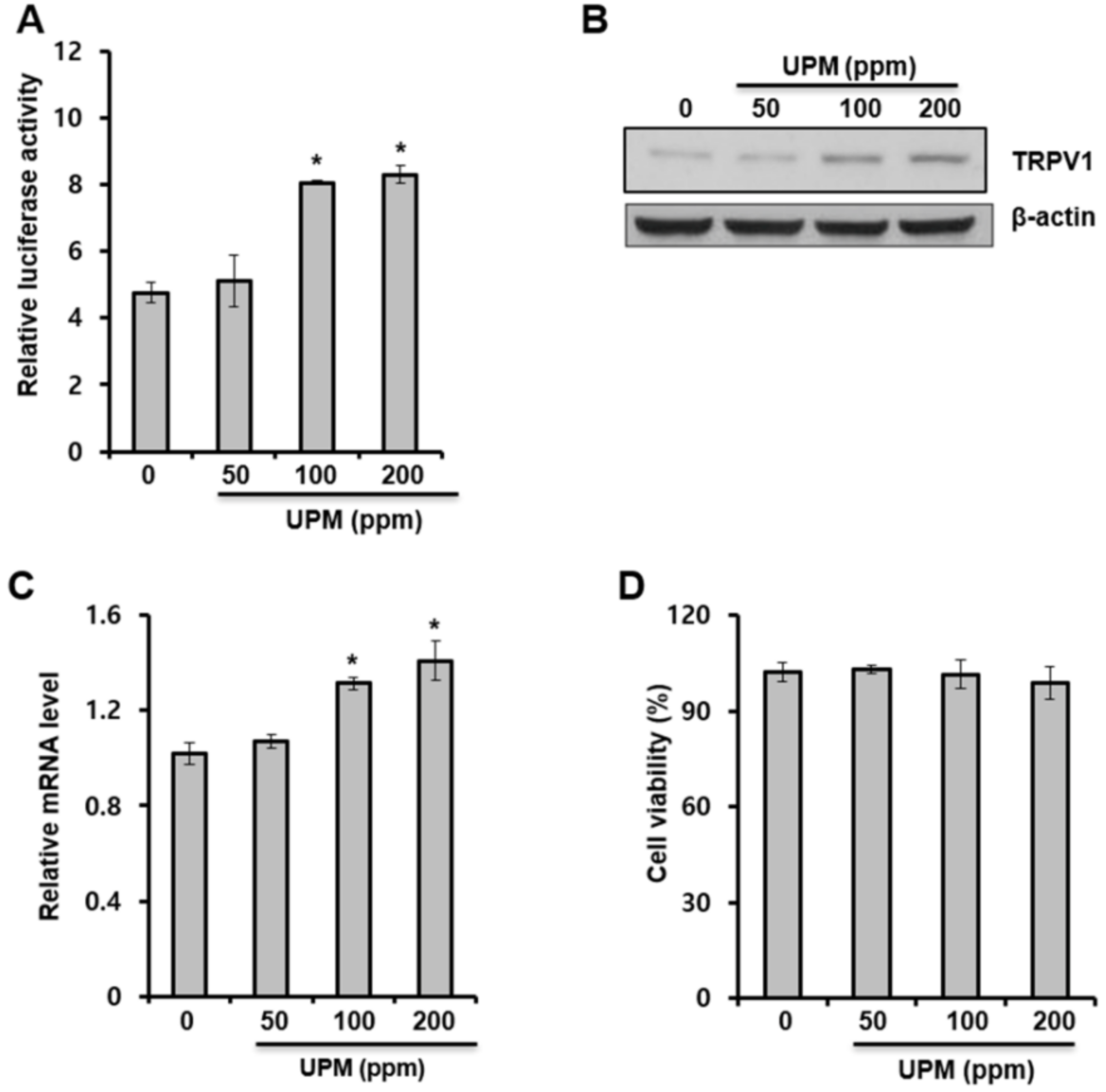
2.1. Urban Particulate Matter Upregulates TRPV 1 Gene Expression in Human Keratinocytes
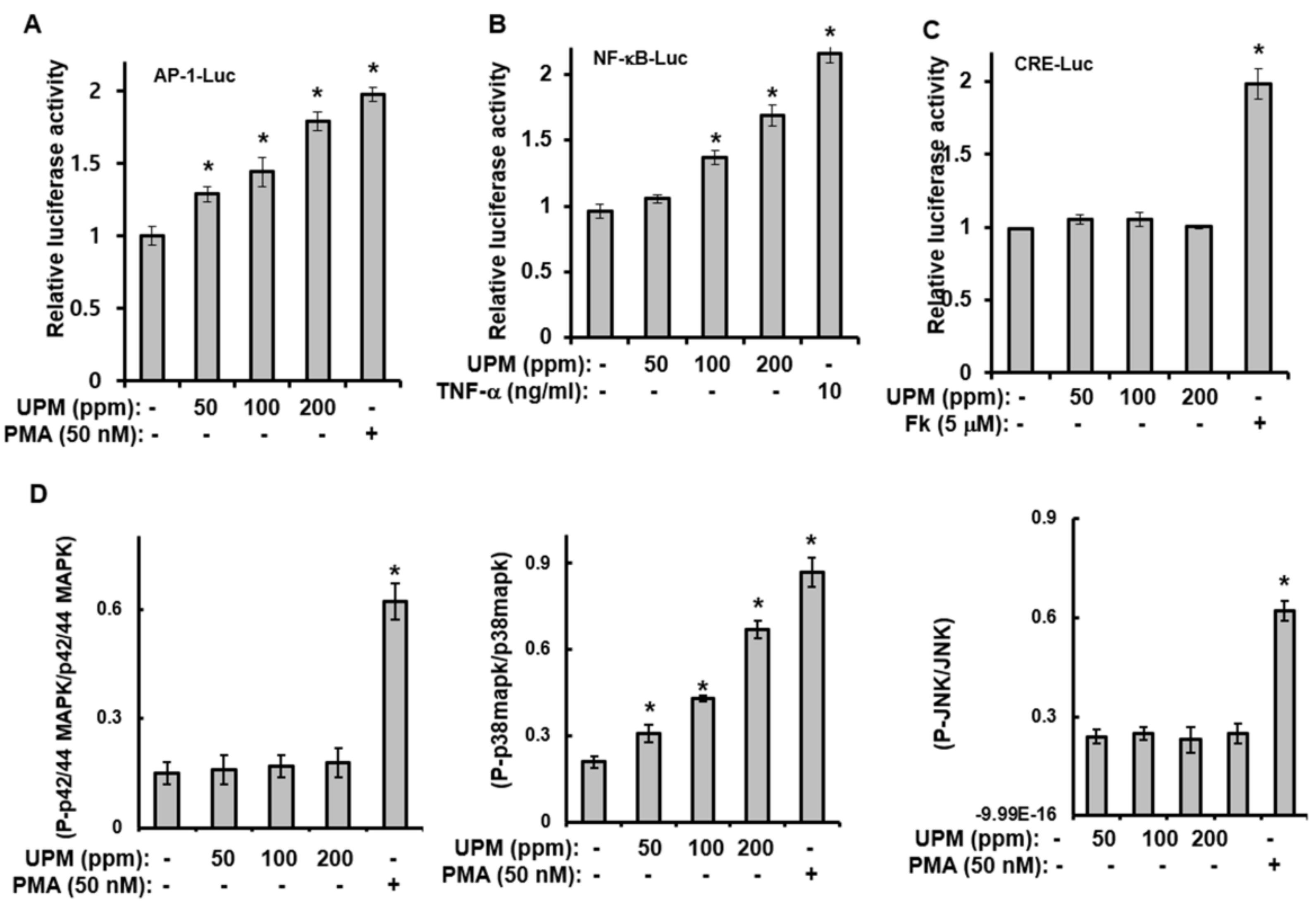
2.2. Urban Particulate Matter Activates AP-1-, CRE-, and NF-κB-Signaling
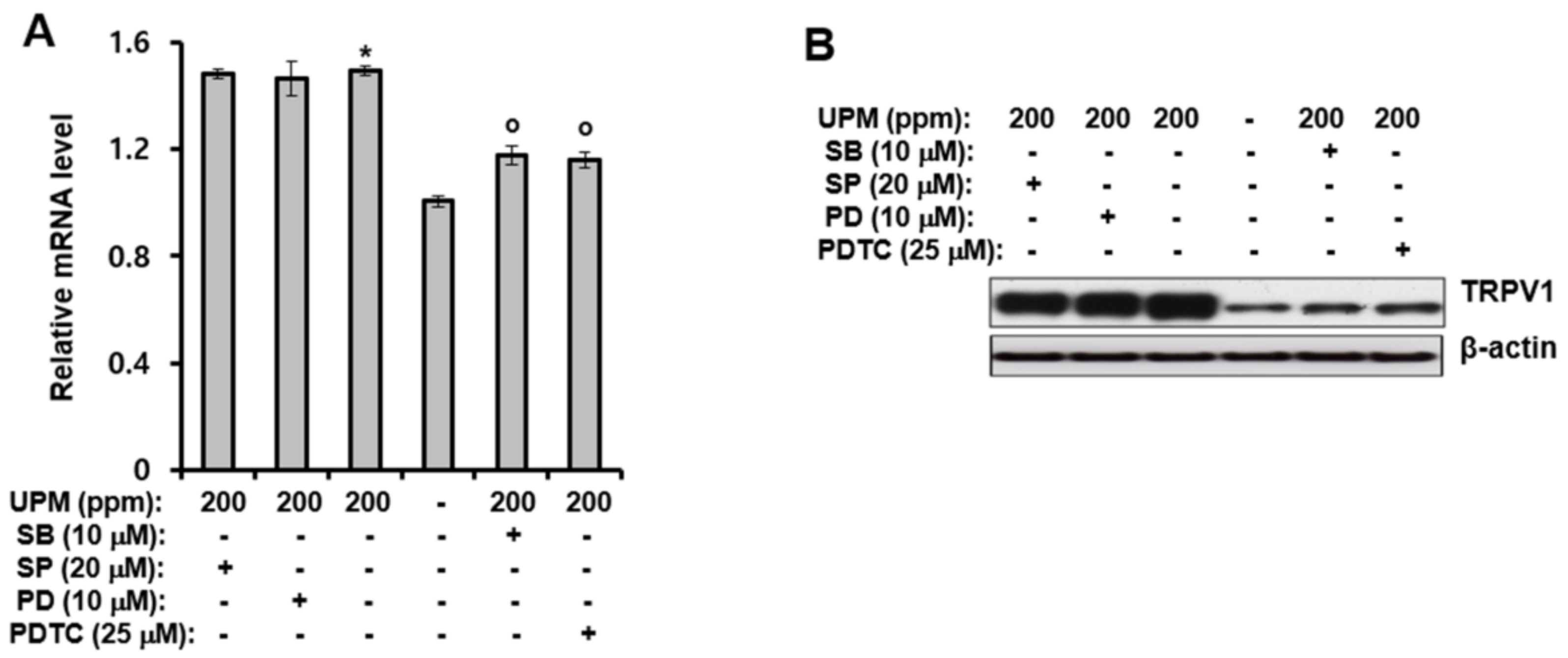
2.3. UPM-induced Expression of TRPV 1 Is Mediated through Activation of p38 MAPK and NF-κB
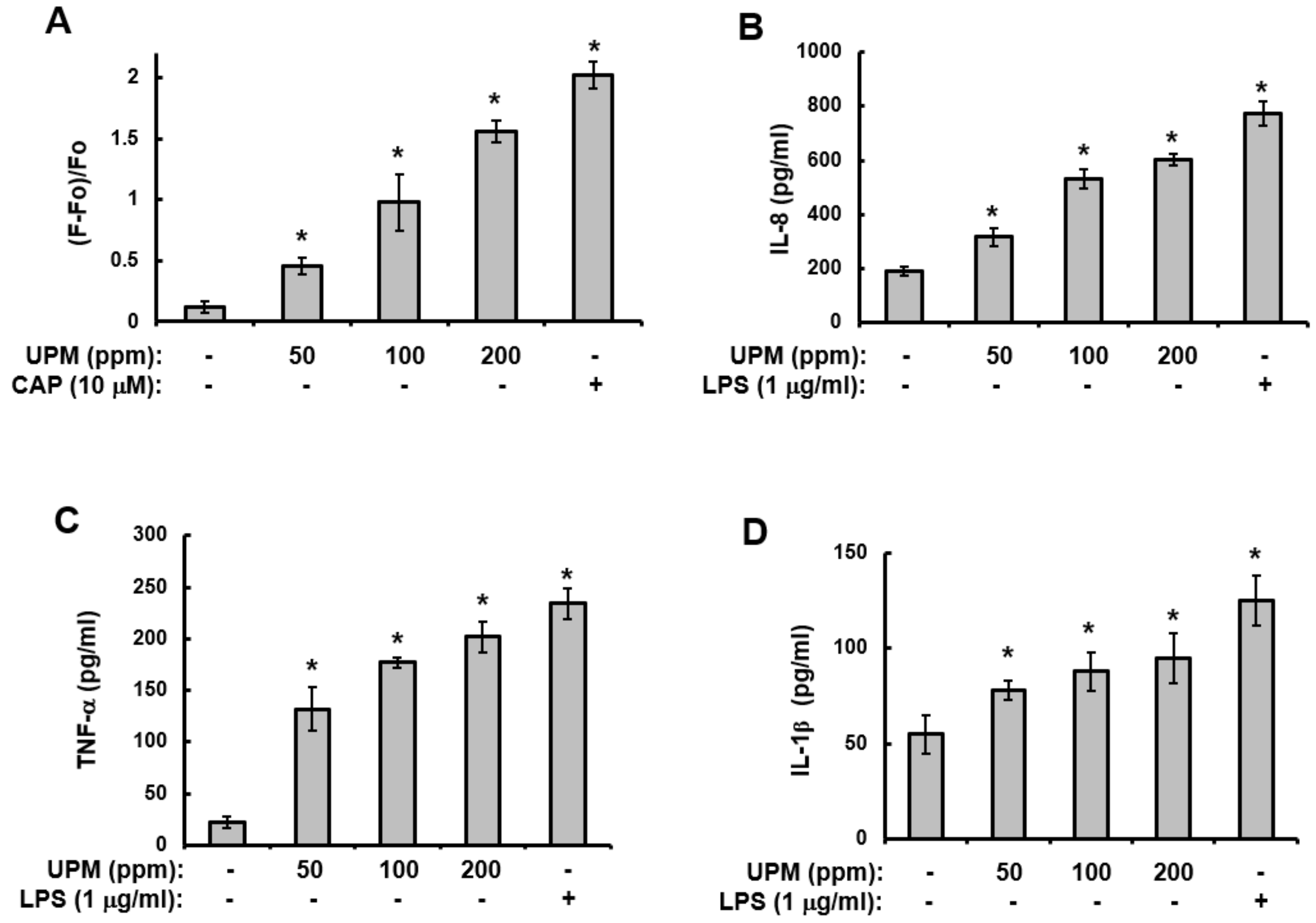
2.4. UPM Increases Ca2+Influx and The Production of Pro-inflammatory Cytokines
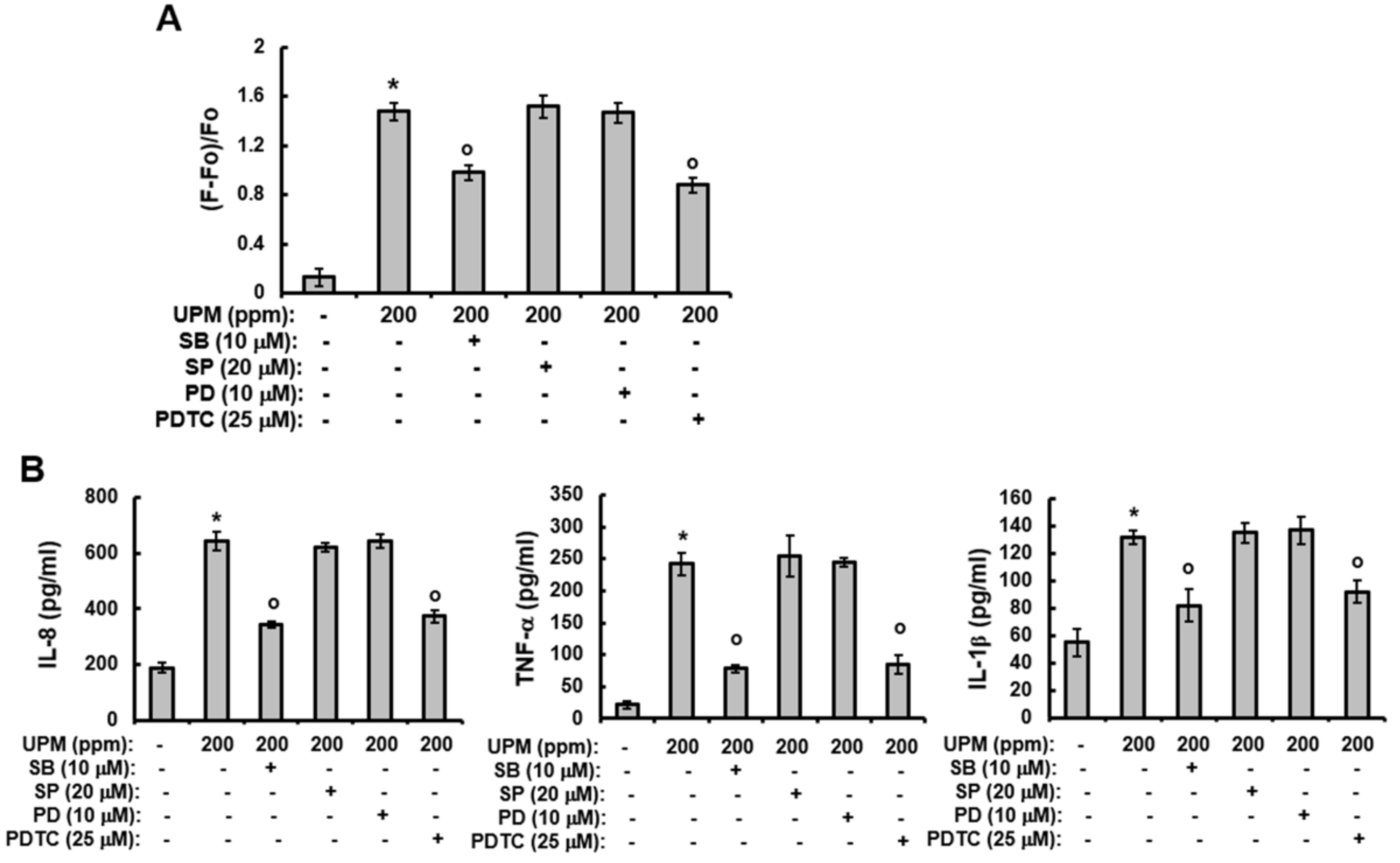
2.5. UPM-induced Increases in Ca2+Influx and The Production of Pro-inflammatory Cytokines are Mediated Through Activation of p38 MAPK and NF-κB
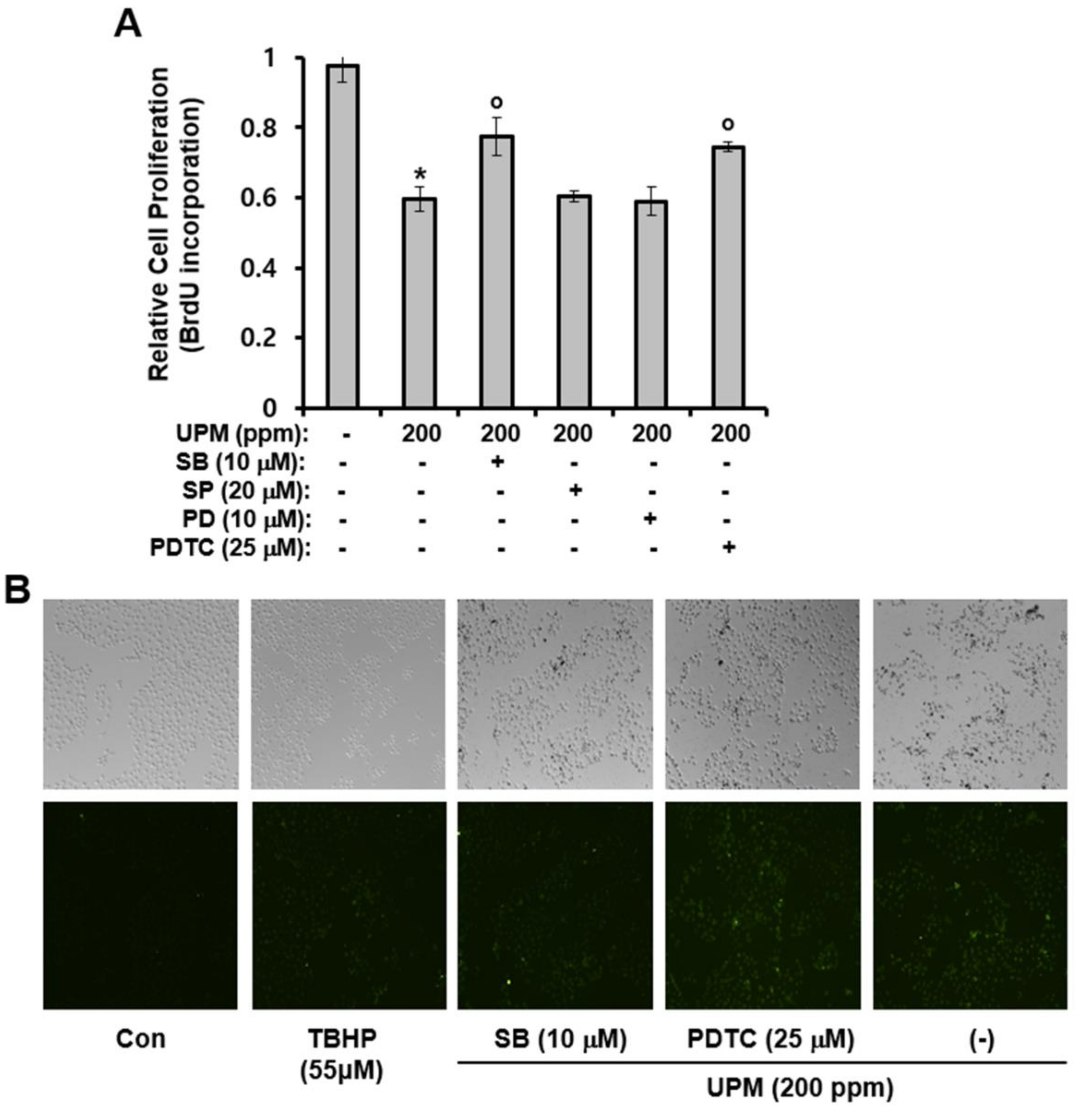
2.6. Effects of UPM On Cell Proliferation and ROS Production
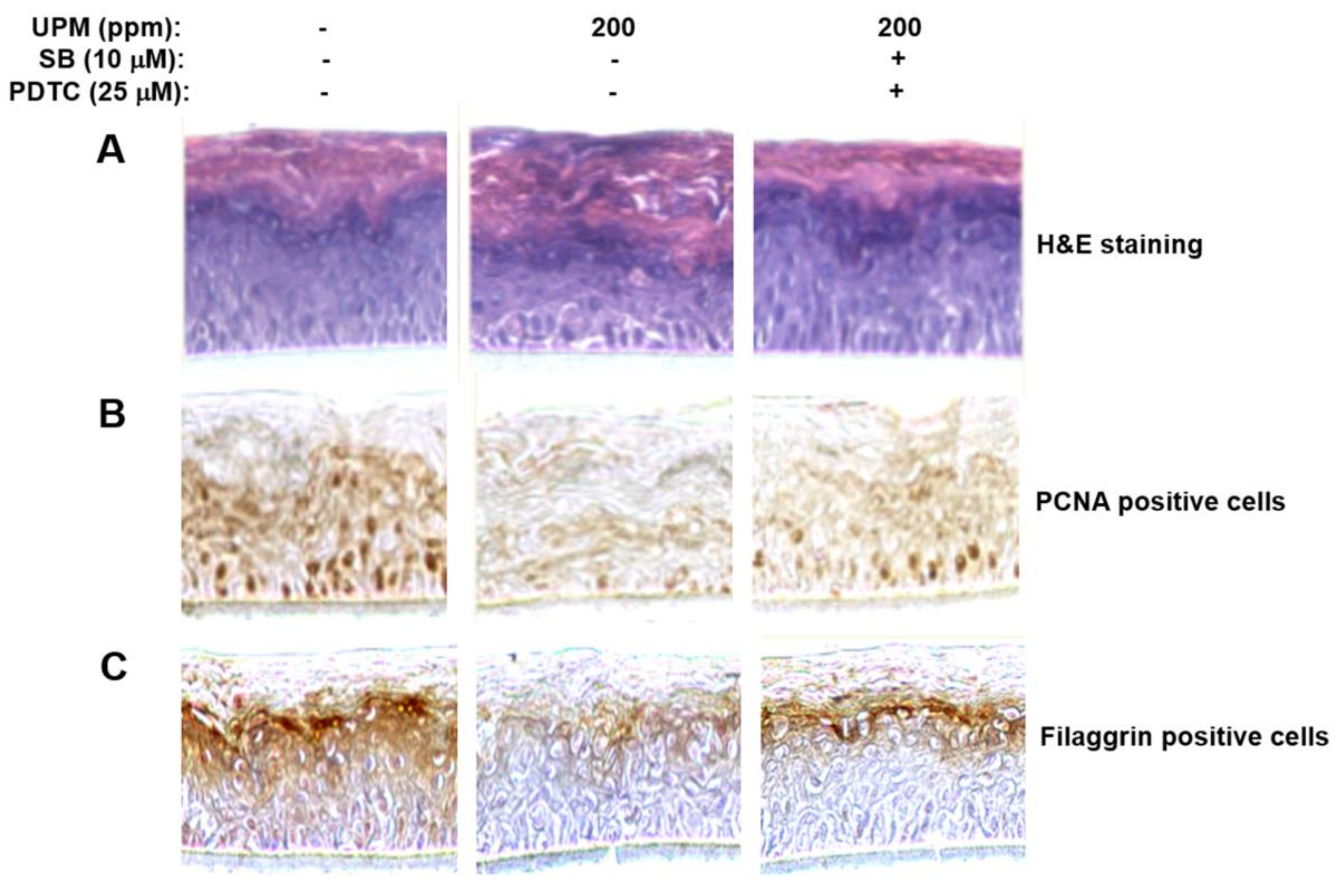
2.7. UPM Affected Morphology of the Reconstructed Epidermis and Reduced Expression of Proliferating Cell Nuclear Antigen (PCNA) Gene and Filaggrin Gene
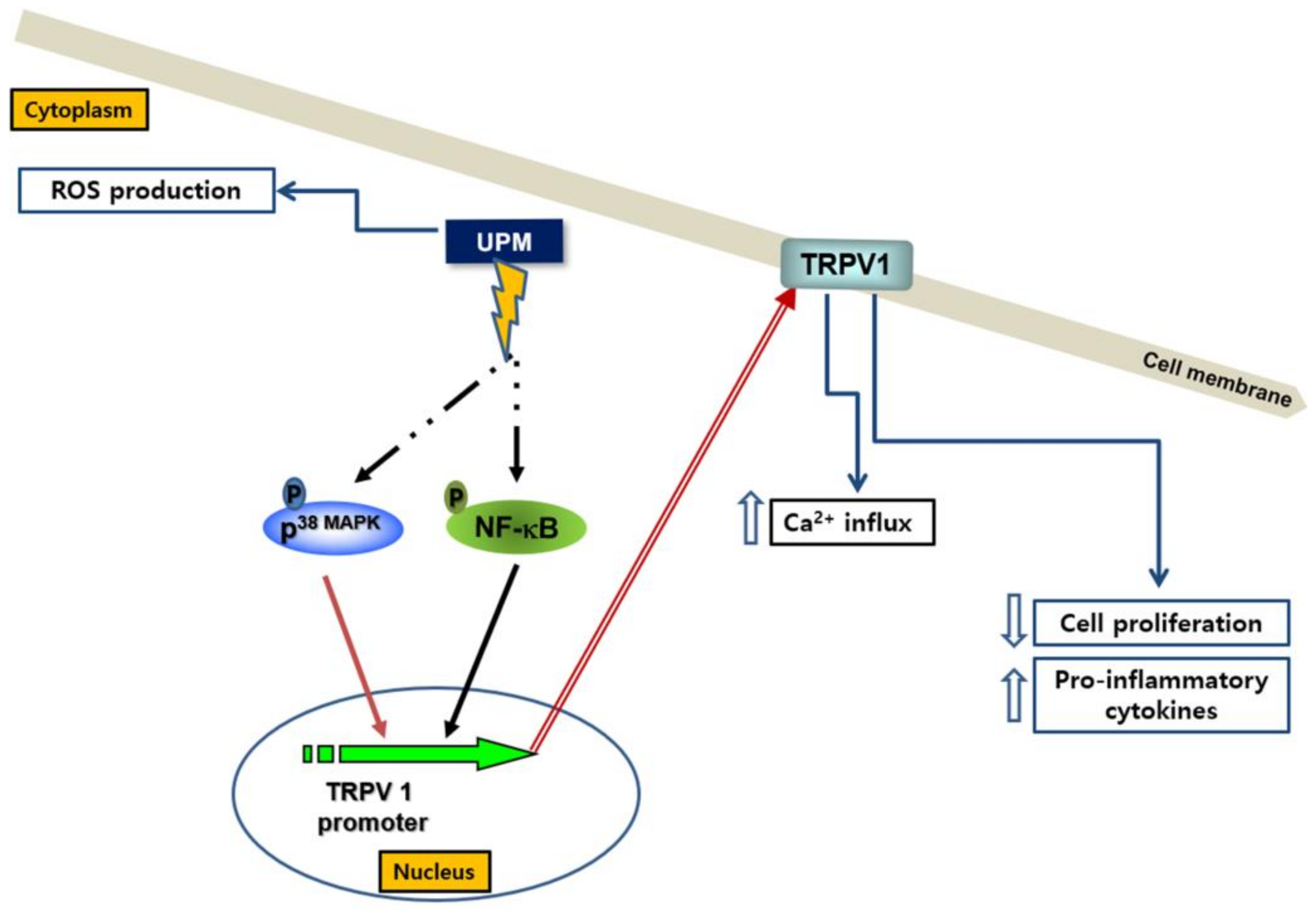
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Culture of HaCaT cells
4.3. Assay for Cytotoxicity of UPM
4.4. BrdU-Incorporation Assay
4.5. Reverse Transcription
4.6. Quantitative Real-time-PCR (qRT-PCR) Analysis
4.7. Measurement of Intracellular Level of Ca2+
4.8. Assay for Luciferase-Reporter Activity
4.9. Immunoblotting
4.10. Analysisof MAPK-Phosphorylation
4.11. ELISA
4.12. Immunohistochemical Staining
4.13. Measurement of ROS Level
4.14. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| UPM | Urban particulate matter |
| TRPV1 | transient receptor potential vanilloid 1 |
| PDTC | Ammonium pyrrolidinedithiocarbamate |
| AP-1 | activator protein-1 |
| NF-κB | nuclear factor kappa B |
| TNF-α | tumor necrosis factor-α |
| IL-8 | interleukin-8 |
References
- Newby, D.E.; Mannucci, P.M.; Tell, G.S.; Baccarelli, A.A.; Brook, R.D.; Donaldson, K.; Forastiere, F.; Franchini, M.; Franco, O.H.; Graham, I.; et al. Expert positionpaper on air pollution and cardiovascular disease. Eur. Heart J. 2015, 36, 83b–93b. [Google Scholar] [CrossRef] [PubMed]
- Franchini, M.; Mannucci, P.M. Short-term effects of air pollution on cardiovascular diseases: Outcomes and mechanisms. J. Thromb. Haemost. 2007, 5, 2169–2174. [Google Scholar] [CrossRef] [PubMed]
- Langrish, J.P.; Bosson, J.; Unosson, J.; Muala, A.; Newby, D.E.; Mills, N.L.; Blomberg, A.; Sandström, T. Cardiovascular effects of particulate air pollution exposure: Time course and underlying mechanisms. Intern. Med. J. 2012, 272, 224–239. [Google Scholar] [CrossRef] [PubMed]
- KimK, H.; Kabir, E.; Kabir, S. A review on the human health impact of airborne particulate matter. Environ. Int. 2015, 74, 136–143. [Google Scholar]
- Palleschi, S.; Rossi, B.; Armiento, G.; Montereali, M.R.; Nardi, E.; MazziottiTagliani, S.; Inglessis, M.; Gianfagna, A.; Silvestroni, L. Toxicity of the readily leachable fraction of urban PM2.5 to human lung epithelial cells: Role of soluble metals. Chemosphere 2018, 196, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.H.; Song, S.H.; Guo, M.; Zhou, J.; Liu, F.; Peng, L.; Fu, Z.R. Long-term exposure to PM2.5 lowers influenza virus resistance via down-regulating pulmonary macrophage Kdm6a and mediates histones modification in IL-6 and IFN-β promoter regions. Biochem. Biophys. Res. Commun. 2017, 493, 1122–1128. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.; Han, S.; Jeon, K.J.; Kwon, S. Effects of collected road dusts on cell viability, inflammatory response, and oxidative stress in cultured human corneal epithelial cells. Toxicol. Lett. 2018, 284, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Guarnieri, M.; Balmes, J.R. Outdoor air pollution and asthma. Lancet 2014, 383, 1581–1592. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Wu, J.; Song, J.; Liang, P.; Zheng, K.; Xiao, G.; Liu, L.; Zouboulis, C.C.; Lei, T. Particulate matter 2.5 regulates lipid synthesis and inflammatory cytokine production in human SZ95 sebocytes. Int. J. Mol. Med. 2017, 40, 1029–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weichenthal, S.A.; Lavigne, E.; Evans, G.J.; Godri Pollitt, K.J.; Burnett, R.T. Fine particulate matter and emergency room visits for respiratory illness effect modification by oxidative potential. Am. J. Respir. Crit. Care Med. 2016, 194, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Fajersztajn, L.; Veras, M.; Barrozo, L.V.; Saldiva, P. Air pollution: A potentially modifiable risk factor for lung cancer. Nat. Rev. Cancer 2013, 13, 674–678. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Sun, Q. Fine particulate matter air pollution and atherosclerosis: Mechanistic insights. Biochim. Biophys. Acta 2016, 1860, 2863–2868. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.W.; Bang, O.Y.; Ahn, K.; Park, S.S.; Park, T.H.; Kim, J.G.; Ko, Y.; Lee, S.; Lee, K.B.; Lee, J.; et al. Air pollution is associated with ischemic stroke via cardiogenic embolism. Stroke 2017, 48, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.S.; Langrish, J.P.; Nair, H.; McAllister, D.A.; Hunter, A.L.; Donaldson, K.; Newby, D.E.; Mills, N.L. Global association of air pollution and heart failure: A systematic review and meta-analysis. Lancet 2013, 382, 1039–1048. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Whitsel, E.A.; Quibrera, P.M.; Smith, R.L.; Liao, D.; Anderson, G.L.; Prineas, R.J. Ambient fine particulate matter exposure and myocardial ischemia in the Environmental Epidemiology of Arrhythmogenesis in the Women’s Health Initiative (EEAWHI) study. Environ. Health Perspect. 2009, 117, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Girguis, M.S.; Strickland, M.J.; Hu, X.; Liu, Y.; Bartell, S.M.; Vieira, V.M. Maternal exposure to traffic-related air pollution and birth defects in Massachusetts. Environ. Res. 2016, 146, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y. The functional regulation of TRPV1 and its role in pain sensitization. Neurochem. Res. 2008, 33, 2008–2012. [Google Scholar] [CrossRef] [PubMed]
- Bodo, E.; Kovacs, I.; Telek, A.; Varga, A.; Paus, R.; Kovacs, L.; Bíró, T. Vanilloidreceptor-1 (VR1) is widely expressed on various epithelial and mesenchymal cell types of human skin. J. Investig. Dermatol. 2004, 123, 410–413. [Google Scholar] [CrossRef] [PubMed]
- Stander, S.; Moormann, C.; Schumacher, M.; Buddenkotte, J.; Artuc, M.; Shpacovitch, V.; Brzoska, T.; Lippert, U.; Henz, B.M.; Luger, T.A.; et al. Expression of vanilloid receptor subtype 1in cutaneous sensory nerve fibers, mast cells, and epithelial cells of appendage structures. Exp. Dermatol. 2004, 13, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Xing, B.M.; Yang, Y.R.; Du, J.X.; Chen, H.J.; Qi, C.; Huang, Z.H.; Zhang, Y.; Wang, Y. Cyclin-dependent kinase 5 controls TRPV1 membrane trafficking and the heat sensitivityof nociceptors through KIF13B. J. Neurosci. 2012, 32, 14709–14721. [Google Scholar] [CrossRef] [PubMed]
- Laı´nez, S.; Valente, P.; Ontoria-Oviedo, I.; Estévez-Herrera, J.; Camprubí-Robles, M.; Ferrer-Montiel, A.; Planells-Cases, R. GABAA receptor associated protein (GABARAP) modulates TRPV1 expression and channel function and desensitization. FASEB J. 2010, 24, 1958–1970. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Du, J.; Yang, Y.; Wang, Y. Phosphorylation of TRPV1 by cyclin-dependent kinase 5 promotes TRPV1 surface localization, leading to inflammatory thermal hyperalgesia. Exp. Neurol. 2015, 273, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Valacchi, G.; Sticozzi, C.; Pecorelli, A.; Cervellati, F.; Cervellati, C.; Maioli, E. Cutaneous responses to environmental stressors. Ann. N. Y. Acad. Sci. 2012, 1271, 75–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ejaz, S.; Ashraf, M.; Nawaz, M.; Lim, C.W. Total particulate matter and wound healing: An in vivo study with histological insights. Biomed. Environ. Sci. 2009, 22, 278–287. [Google Scholar] [CrossRef]
- Vierkotter, A.; Schikowski, T.; Ranfit, U.; Sugiri, D.; Matsui, M.; Kramer, U.; Krutmann, J. Airborne particle exposure and extrinsic skin aging. J. Investig. Dermatol. 2010, 130, 2719–2726. [Google Scholar] [CrossRef] [PubMed]
- Otani, S.; Onishi, K.; Mu, H.; Yokoyama, Y.; Hosoda, T.; Okamoto, M.; Kurozawa, Y. The relationship between skin symptoms and allergic reactions to Asian dust. Int. J. Environ. Res. Public Health 2012, 9, 4606–4614. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Kang, S.M.; Chung, J.H. The role of TRPV 1 channel in aged human skin. J. Dermatol. Sci. 2012, 65, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Pope, C.A., 3rd; Ezzati, M.; Dockery, D.W. Fine-particulate air pollution and life expectancy in the United States. N. Engl. J. Med. 2009, 360, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Rosenlund, M.; Picciotto, S.; Forastiere, F.; Stafoggia, M.; Perucci, C.A. Traffic-related air pollution in relation to incidence and prognosis of coronary heart disease. Epidemiology 2008, 19, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Zhang, X.; Wang, J.; Dandekar, A.; Kim, H.; Qiu, Y.; Xu, X.; Cui, Y.; Wang, A.; Chen, L.C.; et al. Exposure to fine airborne particulate matters induces hepatic fibrosis in murine models. J. Hepatol. 2015, 63, 1397–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinin, A.E.; Kajava, A.V.; Steinert, P.M. Epithelial barrier function: Assembly and structural features of the cornified cell envelope. Bioessays 2002, 24, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, A.V.; Harding, C.R. Moisturization and skin barrier function. Dermatol. Ther. 2004, 17, 43–48. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Concentration | Component | Concentration | |
|---|---|---|---|---|
| Elements | Aluminum (Al) | 3.43 ± 0.13 (%) | Iron (Fe) | 3.92 ± 0.21 (%) |
| Bromine (Br) | 502 ± 10 (mg/kg) | Manganese (Mn) | 790 ± 44 (mg/kg) | |
| Calcium (Ca) | 5.84 ± 0.19 (%) | Sodium (Na) | 4240 ± 60 (mg/kg) | |
| Chlorine (Cl) | 4543 ± 47 (mg/kg) | Sulfur (S) | 5.51 ± 0.36 (%) | |
| Chromium (Cr) | 402 ± 13 (mg/kg) | Titanium (Ti) | 4021 ± 86 (mg/kg) | |
| Copper (Cu) | 610 ± 70 (mg/kg) | Zinc (Zn) | 4800 ± 270 (mg/kg) | |
| Polycyclic aromatic hydrocarbons (PAHs) | Phenanthrene | 4.86 ± 0.17 (mg/kg) | Chrysene | 6.12 ± 0.06 (mg/kg) |
| Fluoranthene | 8.07 ± 0.14 (mg/kg) | Benzo[e]pyrene | 4.85 ± 0.07 (mg/kg) | |
| Pyrene | 5.88 ± 0.07 (mg/kg) | Benzo[ghi] perylene | 5.00 ± 0.18 (mg/kg) | |
| Benz[a] anthracene | 2.71 ± 0.15 (mg/kg) | Indeno[1,2,3-cd] pyrene | 4.17 ± 0.17 (mg/kg) | |
| Polychlorinated biphenyl(PCB) | 2,3,3′,4,4′- Pentachloro- biphenyl | 19.6 ± 2.3 (μg/kg) | 2,2′,4,4′,5,5′- Hexachloro- biphenyl | C40.0 ± 4.9 (μg/kg) |
| 2,3,3′,4′,6- Pentachloro -biphenyl | 25.4 ± 1.9 (μg/kg) | 2,2′,3,4′,5,5′,6- Heptachloro- biphenyl | 17.1 ± 1.4 (μg/kg) | |
| 2,2′,3,4′,5′,6- Hexachloro- biphenyl | 38.9 ± 2.6 (μg/kg) | 2,2′,3,3′,4,4′,5,5′- Octachloro- biphenyl | 19.1 ± 2.2 (μg/kg) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kwon, K.; Park, S.-H.; Han, B.S.; Oh, S.W.; Lee, S.E.; Yoo, J.A.; Park, S.J.; Kim, J.; Kim, J.W.; Cho, J.Y.; et al. Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1. Int. J. Mol. Sci. 2018, 19, 2660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092660
Kwon K, Park S-H, Han BS, Oh SW, Lee SE, Yoo JA, Park SJ, Kim J, Kim JW, Cho JY, et al. Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1. International Journal of Molecular Sciences. 2018; 19(9):2660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092660
Chicago/Turabian StyleKwon, Kitae, See-Hyoung Park, Byung Seok Han, Sae Woong Oh, Seung Eun Lee, Ju Ah Yoo, Se Jung Park, Jangsoon Kim, Ji Woong Kim, Jae Youl Cho, and et al. 2018. "Negative Cellular Effects of Urban Particulate Matter on Human Keratinocytes Are Mediated by P38 MAPK and NF-κB-dependent Expression of TRPV 1" International Journal of Molecular Sciences 19, no. 9: 2660. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19092660