Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes




Abstract
:1. Introduction
2. Results and Discussion
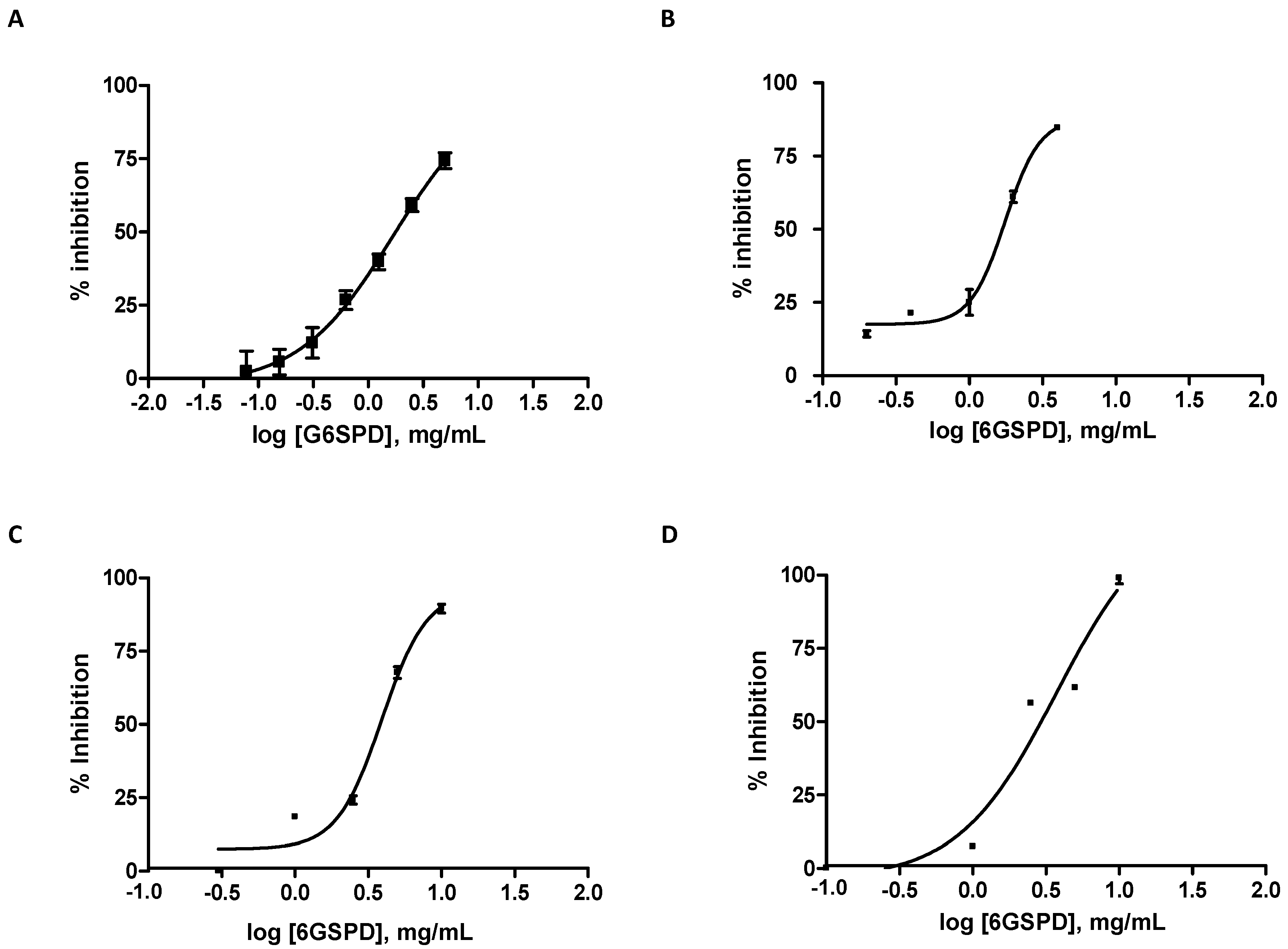
2.1. In Vitro Anti-Diabetic Activity of Germinated Soybean Protein Hydrolyzed Under Simulated Gastrointestinal Conditions
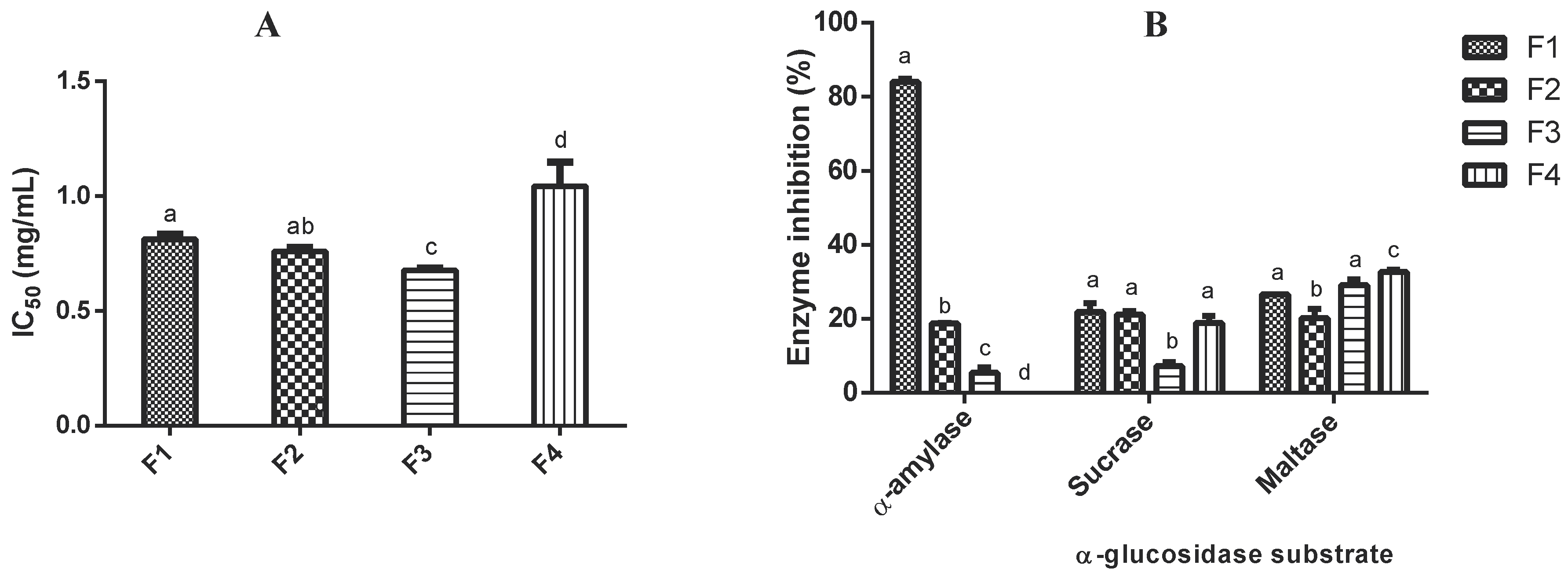
2.2. Identification of Potential Anti-Diabetic Peptides
3. Materials and Methods
3.1. Materials
3.2. Preparation of Germinated Soybean Protein Isolate
3.3. Simulation of Gastrointestinal Digestion
3.4. Measurement of the Enzymatic Inhibitory Activity
3.4.1. DPP-IV Inhibitory Activity
3.4.2. α-Amylase Inhibitory Activity
3.4.3. α-Glucosidase Inhibitory
3.5. Separation of DPP-IV Inhibitory Peptides by Semi-Preparative RP-HPLC
3.6. Identification of Anti-Diabetic Peptides by RP-HPLC-MS/MS
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| 6GSPD | 6-day germinated soybean protein digest |
| DM | diabetes mellitus |
| DPP-IV | dipeptidyl peptidase IV |
| MGAM | maltase-glucoamylase |
| RP-HPLC-MS/MS | RP-HPLC coupled to tandem mass spectrometry |
| SI | sucrase-isomaltase |
| T2DM TFA | type 2 diabetes mellitus trifluoroacetic acid |
References
- Habtamu, A.; Gasmalla, M.A.A.; Yang, R.J.; Zhang, W. Bioactive Peptides Derived from Seaweed Protein and Their Health Benefits: Antihypertensive, Antioxidant, and Antidiabetic Properties. J. Food Sci. 2018, 83, 6–16. [Google Scholar]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.I.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Primers 2015, 1, 15019. [Google Scholar] [CrossRef] [PubMed]
- Silveira, S.T.; Martínez-Maqueda, D.; Recio, I.; Hernández-Ledesma, B. Dipeptidyl peptidase-IV inhibitory peptides generated by tryptic hydrolysis of a whey protein concentrate rich in β-lactoglobulin. Food Chem. 2013, 141, 1072–1077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deacon, C.F. Peptide degradation and the role of DPP-4 inhibitors in the treatment of type 2 diabetes. Peptides 2018, 100, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E. Dipeptidyl peptidase inhibitor therapy in type 2 diabetes: Control of the incretin axis and regulation of postprandial glucose and lipid metabolism. Peptides 2018, 100, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Fitzgerald, R.J. An in silico model to predict the potential of dietary proteins as sources of dipeptidyl peptidase IV (DPP-IV) inhibitory peptides. Food Chem. 2014, 165, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of dipeptidyl peptidase IV, α-amylase and α-glucosidase inhibitory peptides from quinoa (Chenopodium quinoa Willd.) during in vitro simulated gastrointestinal digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Q.; Li, L.; Kwong, J.S.W.; Jia, P.; Zhao, P.; Wang, W.; Zhou, X.; Zhang, M.; Sun, X. α-glucosidase inhibitors and hepatotoxicity in type 2 diabetes: a systematic review and meta-analysis. Sci. Rep. 2016, 6, 32649. [Google Scholar] [CrossRef] [PubMed]
- González-Montoya, M.; Hernández-Ledesma, B.; Silván, J.M.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Peptides derived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells proliferation and inflammation. Food Chem. 2018, 242 (Suppl. C), 75–82. [Google Scholar] [CrossRef] [PubMed]
- Paucar-Menacho, L.M.; Berhow, M.A.; Mandarino, J.M.G.; de Mejia, E.G.; Chang, Y.K. Optimisation of germination time and temperature on the concentration of bioactive compounds in Brazilian soybean cultivar BRS 133 using response surface methodology. Food Chem. 2010, 119, 636–642. [Google Scholar] [CrossRef]
- Bouglé, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Marcela, G.M.; Eva, R.G.; del Carmen, R.R.M.; Rosalva, M.E. Evaluation of the Antioxidant and Antiproliferative Effects of Three Peptide Fractions of Germinated Soybeans on Breast and Cervical Cancer Cell Lines. Plant Foods Human Nutr. 2016, 71, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; Dellafiora, L.; Paolella, S.; Galaverna, G.; Cozzini, P.; FitzGerald, R.J. In Silico Approaches Applied to the Study of Peptide Analogs of Ile-Pro-Ile in Relation to Their Dipeptidyl Peptidase IV Inhibitory Properties. Front. Endocrinol. 2018, 9, 329. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yu, G.; Zhang, Y.; Zhang, B.; Fan, J. Dipeptidyl Peptidase IV Inhibitory Peptides Derived from Oat (Avena sativa L.), Buckwheat (Fagopyrum esculentum), and Highland Barley (Hordeum vulgare trifurcatum (L.) Trofim) Proteins. J. Agric. Food Chem. 2015, 63, 9543–9549. [Google Scholar] [CrossRef] [PubMed]
- Nongonierma, A.B.; FitzGerald, R.J. Investigation of the Potential of Hemp, Pea, Rice and Soy Protein Hydrolysates as a Source of Dipeptidyl Peptidase IV (DPP-IV) Inhibitory Peptides. Food Dig. 2015, 6, 19–29. [Google Scholar]
- Lacroix, I.M.E.; Li-Chan, E.C.Y. Dipeptidyl peptidase-IV inhibitory activity of dairy protein hydrolysates. Int. Dairy J. 2012, 25, 97–102. [Google Scholar] [CrossRef]
- Konrad, B.; Anna, D.; Marek, S.; Marta, P.; Aleksandra, Z.; Józefa, C. The Evaluation of Dipeptidyl Peptidase (DPP)-IV, α-Glucosidase and Angiotensin Converting Enzyme (ACE) Inhibitory Activities of Whey Proteins Hydrolyzed with Serine Protease Isolated from Asian Pumpkin (Cucurbita ficifolia). Int J. Pept Res. Ther. 2014, 20, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-L.; Jao, C.-L.; Ho, K.-P.; Hsu, K.-C. Dipeptidyl-peptidase IV inhibitory activity of peptides derived from tuna cooking juice hydrolysates. Peptides 2012, 35, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. Possible effects of dietary polyphenols on sugar absorption and digestion. Mol. Nutr. Food Res. 2013, 57, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Gericke, B.; Schecker, N.; Amiri, M.; Naim, H.Y. Structure-function analysis of human sucrase-isomaltase identifies key residues required for catalytic activity. J. Biol. Chem. 2017, 292, 11070–11078. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, E.; Oliviero, T.; Udenigwe, C.C. Functional significance and structure–activity relationship of food-derived α-glucosidase inhibitors. Curr. Opin. Food Sci. 2018, 20, 7–12. [Google Scholar] [CrossRef]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Liang, L.; Fang, H.; Xu, F.; Li, W.; Shen, L.; Wang, X.; Xu, C.; Bian, F.; Mu, Y. Efficacy and safety of saxagliptin compared with acarbose in Chinese patients with type 2 diabetes mellitus uncontrolled on metformin monotherapy: Results of a Phase IV open-label randomized controlled study (the SMART study). Diabetes Obes. Metab. 2017, 19, 1513–1520. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Gan, C.Y. Identification of Pinto bean peptides with inhibitory effects on α-amylase and angiotensin converting enzyme (ACE) activities using an integrated bioinformatics-assisted approach. Food Chem. 2018, 267, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Tye, G.J.; Gan, C.Y. The investigation of α-amylase inhibitory activity of selected Pinto bean peptides via preclinical study using AR42J cell. J. Funct. Foods 2017, 35, 641–647. [Google Scholar] [CrossRef]
- Ren, Y.; Liang, K.; Jin, Y.; Zhang, M.; Chen, Y.; Wu, H.; Lai, F. Identification and characterization of two novel α-glucosidase inhibitory oligopeptides from hemp (Cannabis sativa L.) seed protein. J. Funct. Foods 2016, 26, 439–450. [Google Scholar] [CrossRef]
- Simsek, M.; Quezada-Calvillo, R.; Ferruzzi, M.G.; Nichols, B.L.; Hamaker, B.R. Dietary Phenolic Compounds Selectively Inhibit the Individual Subunits of Maltase-Glucoamylase and Sucrase-Isomaltase with the Potential of Modulating Glucose Release. J. Agric. Food Chem. 2015, 63, 3873–3879. [Google Scholar] [CrossRef] [PubMed]
- Uraipong, C.; Zhao, J. In vitro digestion of rice bran proteins produces peptides with potent inhibitory effects on α-glucosidase and angiotensin I converting enzyme. J. Sci. Food Agric. 2018, 98, 758–766. [Google Scholar] [CrossRef] [PubMed]
- Oseguera-Toledo, M.E.; Gonzalez de Mejia, E.; Amaya-Llano, S.L. Hard-to-cook bean (Phaseolus vulgaris L.) proteins hydrolyzed by alcalase and bromelain produced bioactive peptide fractions that inhibit targets of type-2 diabetes and oxidative stress. Food Res. Int. 2015, 76, 839–851. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Bautista-Expósito, S.; Peñas, E.; Dueñas, M.; Silván, J.M.; Frias, J.; Martínez-Villaluenga, C. Individual contributions of Savinase and Lactobacillus plantarum to lentil functionalization during alkaline pH-controlled fermentation. Food Chem. 2018, 257, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [PubMed]
- Ojeda-Montes, M.J.; Gimeno, A.; Tomas-Hernández, S.; Cereto-Massagué, A.; Beltrán-Debón, R.; Valls, C.; Mulero, M.; Pujadas, G.; Garcia-Vallvé, S. Activity and selectivity cliffs for DPP-IV inhibitors: Lessons we can learn from SAR studies and their application to virtual screening. Med. Res. Rev. 2018. [Google Scholar] [CrossRef] [PubMed]
- Power, O.; Nongonierma, A.B.; Jakeman, P.; Fitzgerald, R.J. Food protein hydrolysates as a source of dipeptidyl peptidase IV inhibitory peptides for the management of type 2 diabetes. Proc. Nutr. Soc. 2014, 73, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Lammi, C.; Zanoni, C.; Arnoldi, A.; Vistoli, G. Peptides Derived from Soy and Lupin Protein as Dipeptidyl-Peptidase IV Inhibitors: In Vitro Biochemical Screening and in Silico Molecular Modeling Study. J. Agric. Food Chem. 2016, 64, 9601–9606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammi, C.; Bollati, C.; Ferruzza, S.; Ranaldi, G.; Sambuy, Y.; Arnoldi, A. Soybean-and lupin-derived peptides inhibit DPP-IV activity on in situ human intestinal Caco-2 cells and ex vivo human serum. Nutrients 2018, 1082. [Google Scholar] [CrossRef] [PubMed]
- Ramasubbu, N.; Paloth, V.; Luo, Y.; Brayer, G.D.; Levine, M.J. Structure of human salivary α-amylase at 1.6 Å resolution: Implications for its role in the oral cavity. Acta Crystallogr. D Biol. Crystallogr. 1996, 52, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Ngoh, Y.Y.; Lim, T.S.; Gan, C.Y. Screening and identification of five peptides from pinto bean with inhibitory activities against α-amylase using phage display technique. Enzyme Microb. Technol. 2016, 89, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim Mohammed, A.; Bester Megan, J.; Neitz Albert, W.H.; Gaspar Anabella, R.M. Structural properties of bioactive peptides with α-glucosidase inhibitory activity. Chem. Biol. Drug Des. 2017, 91, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Mojica, L.; De Mejía, E.G. Optimization of enzymatic production of anti-diabetic peptides from black bean (Phaseolus vulgaris L.) proteins, their characterization and biological potential. Food Funct. 2016, 7, 713–727. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kaur, A. Antidiabetic potential of a peptide isolated from an endophytic Aspergillus awamori. J. Appl. Microbiol. 2016, 120, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Mora-Escobedo, R.; Robles-Ramírez, M.d.C.; Ramón-Gallegos, E.; Reza-Alemán, R. Effect of Protein Hydrolysates from Germinated Soybean on Cancerous Cells of the Human Cervix: An In Vitro Study. Plant Foods Hum. Nutr. 2009, 64, 271–278. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Enzymatic Inhibitory Activity (IC50, mg/mL) * | ||||
|---|---|---|---|---|
| Samples | DPP-IV | α-Amylase | α-Glucosidase | |
| Maltase Activity | Sucrase Activity | |||
| Positive control 1 | 0.003 ± 0.000 a | 0.16 ± 0.01 a | 0.07 ± 0.01 a | 0.03 ± 0.00 a |
| 6GSPD | 1.49 ± 0.14 c | 1.70 ± 0.18 b | 3.73 ± 0.06 c | 2.90 ± 0.07 d |
| >10 kDa | 1.18 ± 0.15 b | 4.80 ± 0.87 c | >10.00 d | 5.27 ± 0.16 e |
| 5–10 kDa | 0.91 ± 0.17 b | >10.00 | 3.56 ± 0.14 c | 2.20 ± 0.40 c |
| <5 kDa | 2.21 ± 0.15 d | 8.30 ± 1.65 d | 2.56 ± 0.03 b | 1.23 ± 0.19 b |
| Peptide Fraction | Mass | Protein Fragment | Peptide Sequence | Net Charge c | Hydrophobicity (kcal/mol) c | DPP-IV Inhibitory Peptides a | Antioxidant Peptides a | ACE Inhibitory Peptides a |
|---|---|---|---|---|---|---|---|---|
| F1 b | 834.3 | β-conglycinin α, α’and β-chain f(270–276) | NNDDRDS | −2 | +22.79 | DR, ND, NN | --- | --- |
| 1013.4 | β-conglycinin α and α’-chain f(295–303) | VVNPDNNEN | −2 | +17.79 | DN, NE, NN, NP, VN, VV | NEN | VNP | |
| 949.4 | β-conglycinin α and β-chain f(324–332) | LSSTEAQQS | −1 | +13.95 | QQ, QS, TE | --- | EA, ST, TE | |
| 958.4 | β-conglycinin α, α’and β-chain f(530–537) | NAENNQRN | 0 | +18.01 | AE, NA, NN, NQ, RN | --- | --- | |
| 777.4 | β-conglycinin α’-chain f(604–610) | IKSQSES | 0 | +15.36 | ES, KS, QS | --- | --- | |
| 981.4 | Glycinin G1 subunit f(112–119) | EEPQQPQQ | −2 | +18.52 | EP, QP, PQ, QQ | --- | PQ | |
| 874.4 | Glycinin G1 subunit f(121–128) | GQSSRPQD | 0 | +17.10 | GQ, PQ, QD, QS, RP | --- | GQ, PQ, RP | |
| 887.4 | Glycinin G2 and G1 subunit f(181–188) | LAGNQEQE | −2 | +17.95 | AG, LA, NQ, QE | --- | AG, LA | |
| 787.4 | Glycinin G1 subunit f(465–471) | NLKSQQA | +1 | +12.80 | KS, NL, QA, QQ | LK | --- | |
| 972.4 | Glycinin G2 subunit f(109–116) | QEPQESQQ | −2 | +18.84 | EP, ES, PQ, QE, QQ | --- | PQ | |
| 1150.6 | Glycinin G2 subunit f(120–128) | SQRPQDRHQ | +1 | +20.40 | DR, PQ, QD, RH, RP | RHQ | PQ, RP | |
| 1202.5 | Glycinin G2 subunit f(193–203) | QQQQQGGSQSQ | 0 | +16.50 | GG, QG, QQ, QS | --- | GG, GS, QG | |
| 1387.7 | Glycinin G2 subunit f(193–205) | QQQQQGGSQSQKG | +1 | +20.46 | GG, KG, QG, QQ, QS | --- | GG, GS, KG, QG, QK | |
| 1244.6 | Glycinin G2 and G5 subunit f(198–207) | PETMQQQQQQ | −1 | +15.87 | ET, MQ, QQ, TM | --- | --- | |
| 1311.5 | P34 Probable thiol protease f(250–261) | SDESTESETEQA | −5 | +29.21 | ES, ET, QA, TE | --- | ST, TE | |
| F2 | 1546.7 | Glycinin G2 subunit f(238–251) | RNLQGENEEEDSGA | −4 | +32.34 | GA, GE, NE, NL, QG, RN | --- | GA, GE, LQ, QG, SG |
| 843.5 | Glycinin G4 subunit f(451–458) | VTRGQGKV | +2 | +14.89 | GQM KVM QG, RG, TR, VT | --- | GK, GQ, QG, RG, GKV, VTR | |
| 644.4 | P34 Probable thiol protease f(143–148) | KKGVIT | +2 | +13.32 | GV, KG, KK, VI | --- | GV, KG | |
| 1555.6 | P34 Probable thiol protease f(248–261) | IMSDESTESETEQA | −5 | +27.42 | ES, ET, IM, QA, TE | --- | ST, TE | |
| F3 | 1497.7 | Glycinin G1 subunit f(37–50) | NALKPDNRIESEGG | −1 | +26.14 | AL, DN, EG, ES, GG, KP, NA, NR, RI | LK, KP, LKP | EG, GG, IE, KP, LKP |
| 1433.7 | Glycinin G1 subunit f(329–342) | SSPDIYNPQAGSVT | −1 | +14.43 | AG, NP, PQ, QA, SP, SV, VT, YN | IY | AG, GS, IY, PQ, YN, AGS | |
| 1497.7 | Glycinin G2 subunit f(34–47) | NALKPDNRIESEGG | −1 | +26.14 | AL, DN, EG, ES, GG, KP, NA, NR, RI | LK, KP, LKP | EG, GG, IE, KP, LKP | |
| 1956.9 | Glycinin G2 subunit f(312–329) | RQNIGQNSSPDIYNPQAG | 0 | +19.57 | AG, GQ, PQ, QA, QN, SP, YN | IY | AG, GQ, IG, IY, PQ, YN | |
| 1497.7 | Glycinin G3 subunit f(37–50) | NALKPDNRIESEGG | −1 | +26.14 | AL, DN, EG, ES, GG, KP, NA, NR, RI | LK, KP, LKP | EG, GG, IE, KP, LKP | |
| 1261.6 | Glycinin G4 subunit f(486–497) | VVAEQAGEQGFE | −3 | +21.00 | AE, AG, GE, GF, QA, QG, VA, VV | --- | AG, GE, GF, QG | |
| 882.5 | β-conglycinin α-chain f(190–196)β-conglycinin α’-chain f(206–212) | HKNKNPF | +2 | +15.96 | NP, PF | --- | HK, NK |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Montoya, M.; Hernández-Ledesma, B.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes. Int. J. Mol. Sci. 2018, 19, 2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102883
González-Montoya M, Hernández-Ledesma B, Mora-Escobedo R, Martínez-Villaluenga C. Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes. International Journal of Molecular Sciences. 2018; 19(10):2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102883
Chicago/Turabian StyleGonzález-Montoya, Marcela, Blanca Hernández-Ledesma, Rosalva Mora-Escobedo, and Cristina Martínez-Villaluenga. 2018. "Bioactive Peptides from Germinated Soybean with Anti-Diabetic Potential by Inhibition of Dipeptidyl Peptidase-IV, α-Amylase, and α-Glucosidase Enzymes" International Journal of Molecular Sciences 19, no. 10: 2883. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19102883