1. Introduction
Granulocyte colony-stimulating factor (G-CSF)-mobilized peripheral blood stem cell grafts are used for allogeneic stem cell transplantation (ALLO-SCT) [
1]. This G-CSF therapy has several immediate effects on the donor immune system but does not seem to have any long-term consequences [
2]. It increases levels of various anti-inflammatory cytokines while simultaneously decreasing the production of several proinflammatory cytokines [
3,
4], inhibits T cell responsiveness and shifts their differentiation towards Th2 responses [
5,
6], induces IL-10 producing allo-inhibitory regulatory T cells [
7], promotes the development of myeloid-derived tolerogenic dendritic cells [
8], and reduces serum levels of the chemotactic C-X-C motif ligand 8 (CXCL8) and C-X-C motif ligand 12 (CXCL12) chemokines [
9]. Thus, anti-inflammatory effects are common [
8].
The increased risk of graft-versus-host disease (GVHD) for patients receiving G-CSF-mobilized stem cells has been explained by the increased number of donor T cells in these grafts [
10]. However, the effects of G-CSF therapy in healthy individuals are complex as illustrated both by the frequent reversible side effects (e.g., musculoskeletal pain) and uncommon but more severe toxicity (e.g., splenic rupture and pulmonary toxicity), including progression of arthritis as an example of a proinflammatory effect [
11,
12]. A recent study also described metabolic effects of G-CSF therapy in healthy stem cell donors, and these effects may influence immunoregulation [
13]. Furthermore, the systemic level of the proinflammatory acute phase stimulant interleukin-6 (IL-6) is also increased for a subset of such donors [
9], but it is not known which cells are responsible for this IL-6 response [
6,
14].
Optimal selection of the healthy stem cell donor is essential for outcome after allogeneic stem cell transplantation, and among the well-characterized donor risk factors are major histocompatibility complex mismatches, female donor for male patient, donor age, and Killing Immunoglobulin-like Receptor genotype [
15]. As described in a recent article, several studies have now described associations between graft compositions and outcome after ALLO-SCT [
16], and the first study of individualized GVHD prophylaxis based on graft composition has already been published [
17]. However, several studies have demonstrated that the immunomodulatory effects of G-CSF-induced stem cell mobilization differ between healthy donors [
18]. Firstly, the effects of G-CSF on serum levels of a wide range of both pro- and anti-inflammatory cytokines, as well as soluble adhesion molecules and extracellular proteases, differ between healthy donors [
9,
19]; Secondly, the effect of G-CSF on immunoregulatory metabolites also varies [
13]; Thirdly, the numbers of different immunocompetent cell subsets vary between grafts derived from different donors [
20]; Finally, a recent study suggests that the responsiveness of immunocompetent cells to G-CSF administration differs between healthy donors, i.e., there are qualitative differences, and not only quantitative differences, between grafts derived from different donors [
21]. An important question is therefore whether the G-CSF induced immunomodulation is heterogeneous and whether such differences between donors have an impact on outcome after allotransplantation [
16]. The aims of our present study were therefore to investigate whether IL-6 or other IL-6 family cytokines/receptors are influenced by G-CSF therapy and thereby contribute to the heterogeneity of healthy allogeneic stem cell donors, to examine whether this heterogeneity is important for outcome after allogeneic stem cell transplantation, and to elucidate whether G-CSF will alter the release of IL-6 by in vitro cultured monocytes and/or fibroblasts.
IL-6 depends on gp130 for transmembrane signaling, and C-reactive protein (CRP) production is mainly driven by classical IL-6 signaling (dependent on membrane-bound IL-6 receptors) whereas trans-signaling (dependent on soluble IL-6 receptors) seems less important [
22]. G-CSF increases IL-6 levels and would therefore be expected to increase the acute phase reaction (including CRP). However, one should emphasize that the final effect of G-CSF on CRP levels depends on the biological context and G-CSF can reduce the acute phase responses after tissue injury [
23]. Other cytokines that depend on gp130 for signal transduction (e.g., other IL-6 family members) may then induce an acute phase response in the absence of IL-6 [
24,
25]. Taken together, these observations suggest that the balance between pro- and anti-inflammatory effects of G-CSF and IL-6 depends on the clinical context. This is also supported by previous studies of post-transplant G-CSF therapy in allotransplant recipients; whether G-CSF therapy will influence post-transplant survival depends on the conditioning therapy and the type of stem cell graft [
26]. We have previously reviewed the scientific evidence for a role of IL-6 in the development of immune-mediated complications after allotransplantation [
27], and previous studies have also shown that IL-6 serum levels are altered during G-CSF mobilization for a large subsets of healthy stem cell donors [
9,
19,
28]. Even though risk-adapted GVHD prophylaxis based on variations in graft composition is already considered, a better understanding of the mechanisms behind, and the consequences of, donor and graft heterogeneity is needed, including the possible roles of the IL-6 family and the contribution of G-CSF to the heterogeneity. In our present study; we therefore investigated effects of G-CSF on systemic levels of CRP and IL-6 cytokine family members in healthy stem cell donors.
3. Discussion
Previous studies suggest that healthy stem cell donors are heterogeneous with regard to the effects of G-CSF on donor immunoregulation and the number, as well as the functional status, of immunocompetent graft cells [
9,
13,
15,
18,
19,
20,
21]. One of these studies even suggests that G-CSF induced donor heterogeneity is important for outcome after allotransplantation [
21]. We have previously reviewed and discussed the available evidence for a role of IL-6 in the development of immune-mediated complications after allotransplantation [
27]. Previous studies have also shown that systemic IL-6 levels in healthy stem cell donors can be altered by G-CSF therapy; these effects are divergent, and although increased levels are seen for most donors, a minority of them show decreased systemic IL-6 levels in response to G-CSF [
9,
19,
28]. In our present study we observed that healthy donors undergoing G-CSF induced stem cell mobilization and harvesting by leukapheresis are heterogeneous, both with regard to the G-CSF induced acute phase reaction and effects of G-CSF on systemic levels of various IL-6 cytokine/receptor family members.
Several recent studies have described associations between graft composition and post-transplant outcome, e.g., high CD8
+ graft cells associated with decreased relapse risk [
32] and increased regulatory T cells associated with decreased nonrelapse mortality [
33]. The first study investigating individualized risk-adapted prophylaxis against immune-mediated complications based on graft composition has already been published [
17]. However, a better understanding of the molecular mechanisms behind, and the consequences for, the recipients of differences in graft composition is needed as a scientific basis for further studies of possible interventions, e.g., in vivo graft manipulation, ex vivo graft manipulation, risk-adapted individualized prophylaxis, or early therapeutic intervention based on biomarker evaluation before clinical signs of complications [
16].
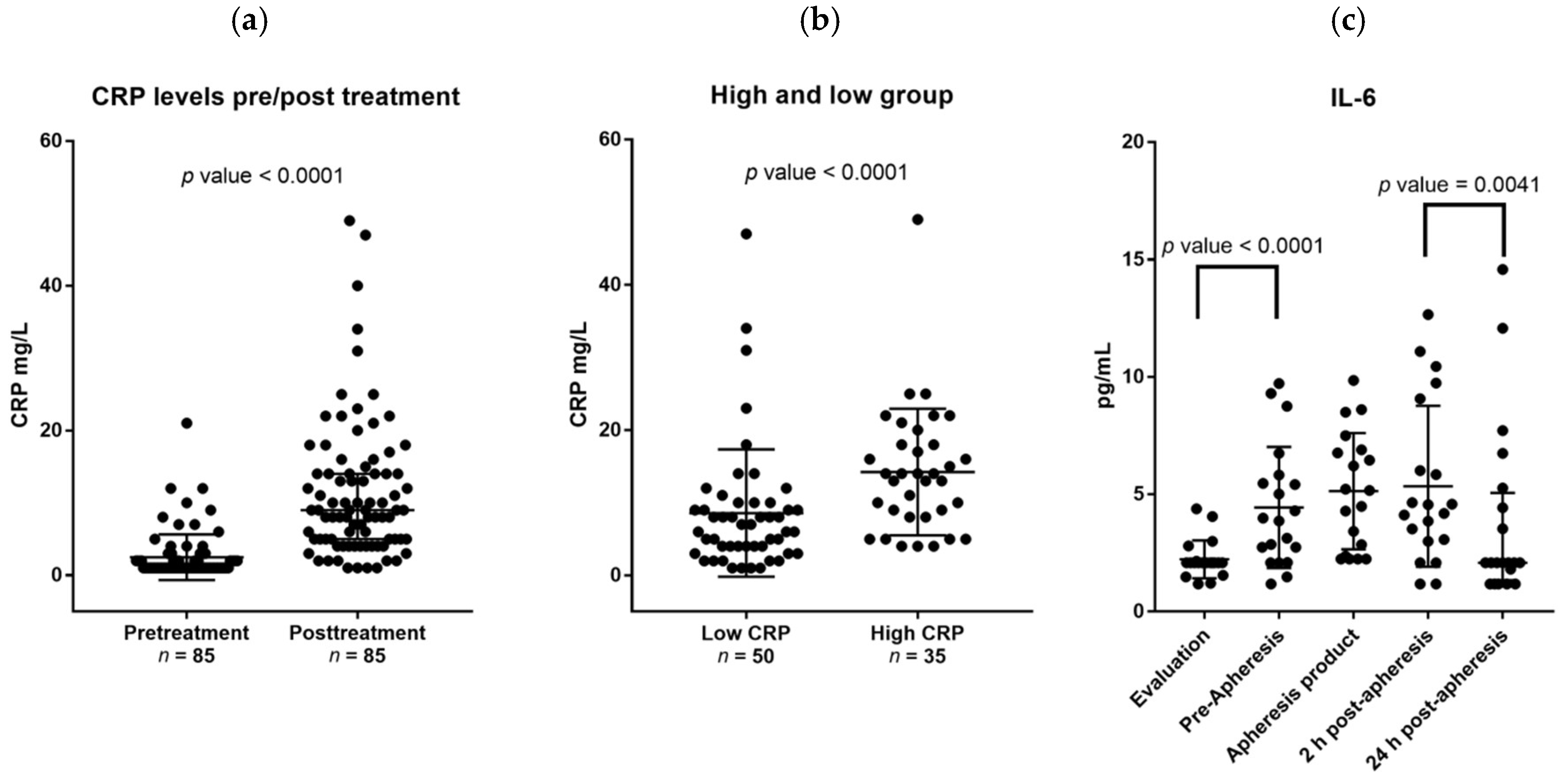
Previous studies suggest that G-CSF-induced stem cell mobilization in healthy individuals has a clinically negligible effect on CRP levels with most donors still having CRP levels below 2 mg/L after G-CSF administration [
34,
35]. In contrast, we observed an increase of at least 9.5 mg/L for a large subset of donors, especially elderly donors. The only other factor predicting this CRP increase was the pretreatment CRP levels, implicating that signs of pretreatment inflammation potentiates the effects of G-CSF on the acute phase reaction.
IL-6 and CRP levels are usually highly correlated [
36]; this was also seen for the pretreatment levels for our stem cell donors. However, we did not detect any significant association between CRP and IL-6 levels during G-CSF treatment, but CRP levels were significantly correlated with OSM levels even though the OSM levels did not increase in response to G-CSF. G-CSF itself is not able to induce CRP production in hepatocytes [
37]. Taken together, these observations suggest that G-CSF induced CRP release is independent of the IL-6 response and rather caused by a G-CSF induced modulation of OSM effects. Even though tumor necrosis factor alpha (TNF-α) or IL-1 can induce CRP release [
38], these two cytokines are less likely to contribute because G-CSF decreases their systemic levels [
8]. Finally, the ratio between serum levels of sIL-6 receptor and sgp130 is termed the IL-6 buffer; this buffer regulates the proinflammatory effects of IL-6, including its effects on the acute phase response/CRP levels [
39,
40]. However, the IL-6 buffer was not altered during G-CSF therapy and therefore is unlikely to be responsible for the increased CRP levels during G-CSF therapy [
24,
25].
We observed an association between the G-CSF induced acute phase reaction and OSM levels. OSM is released by various immunocompetent cells; it can initiate acute phase reactions and is also involved in tissue repair [
41,
42]. The OSM receptor uses gp130 as the signaling subunit of the receptor complex; this is similar to the other IL-6 family cytokines, but OSM can also utilize the LIF receptors for signal transduction [
27]. OSM seems to have the broadest downstream signaling profile among the IL-6 family members and activates Janus kinase (Jak)/ Signal transducer and activator of transcription (STAT) signaling, the extracellular signal–regulated kinases (ERK) and c-Jun N-terminal kinase, phosphatidyl-inositole-3-kinase/ Protein Kinase B (Akt) signaling, as well as protein kinase C delta [
41,
43].
OSM is also regarded as a disruptor of epithelial barrier functions, it is a biomarker for active inflammation in rheumatoid arthritis and increased levels are also reported in allergic rhinitis, psoriasis, and asthma [
42]. It seems to have a very complex role in the regulation of inflammation by enhancing the maturation of dendritic cells and thereby increasing their IL-12 release, increasing T cell proliferation, and increasing the release of Interferon-γ [
44]. However, it also seems to skew monocyte differentiation into the anti-inflammatory M2 phenotype and does not stimulate development of dendritic cells from monocytes. In vivo studies suggest that OSM has anti-inflammatory effects mediated by inhibition of IL-1 and TNF-α responses, and it seems to suppress inflammation in animal models of autoimmune diseases [
45]. OSM does not seem to have direct effects on Th17 cells and regulatory T cells [
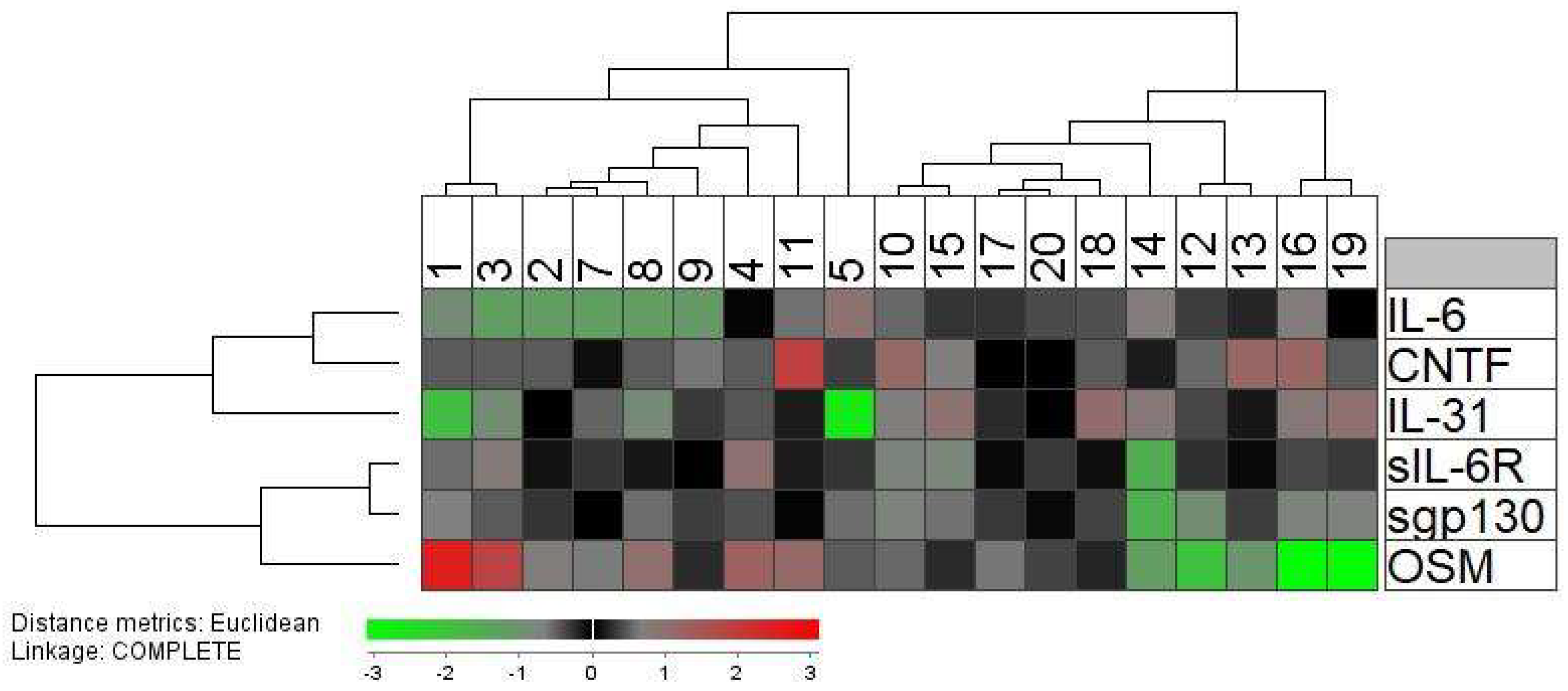
44]. Taken together, these observations suggest that the predominant effects of OSM depend on the biological context. Our present results suggest that its proinflammatory effects (i.e., the effects on the acute phase reaction) vary between, and thereby contributes to, the heterogeneity of healthy stem cell donors (
Table 2,
Figure 2), and this variation during G-CSF therapy and in graft supernatants suggests that OSM can alter the functional status of at least certain subsets of graft immunocompetent cells. Even though the possible role of OSM in allotransplant recipients has not been addressed previously, our knowledge about OSM from other studies suggests that it may contribute to the post-transplant outcome (e.g., development of immune-mediated toxicity) in allotransplant recipients through the acute phase reaction, immunoregulatory and proinflammatory effects, modulation of inflammatory resolution and tissue repair after inflammation, or effects on epithelial barrier functions.
The peripheral blood levels and the corresponding graft amounts of immunocompetent cells showed a wide variation between healthy donors (
Table 3), and the widest variation in peripheral blood levels was seen for NK cells. The NK cells seem important for outcome after stem cell transplantation [
46]. Previous studies have also demonstrated that NK cells show a transient functional alteration following G-CSF mobilization with decreased proliferative capacity; this effect also varies between patients [
47,
48]. Thus, healthy stem cell donors show both a quantitative and qualitative NK cell heterogeneity after G-CSF mobilization.
The levels of several circulating immunocompetent cell subsets showed an association with the systemic levels of sgp130 that serves as an important modulator of IL-6 signaling through its binding to soluble IL-6R [
27]. This observation suggests that IL-6 family cytokines, and especially IL-6, are important for immunocompetent cell mobilization and may contribute to the donor heterogeneity observed during G-CSF therapy. These associations were not detected for the allografts, probably because graft levels also depend on factors related to the apheresis and graft preparation and not only on the G-CSF mobilization [
28].
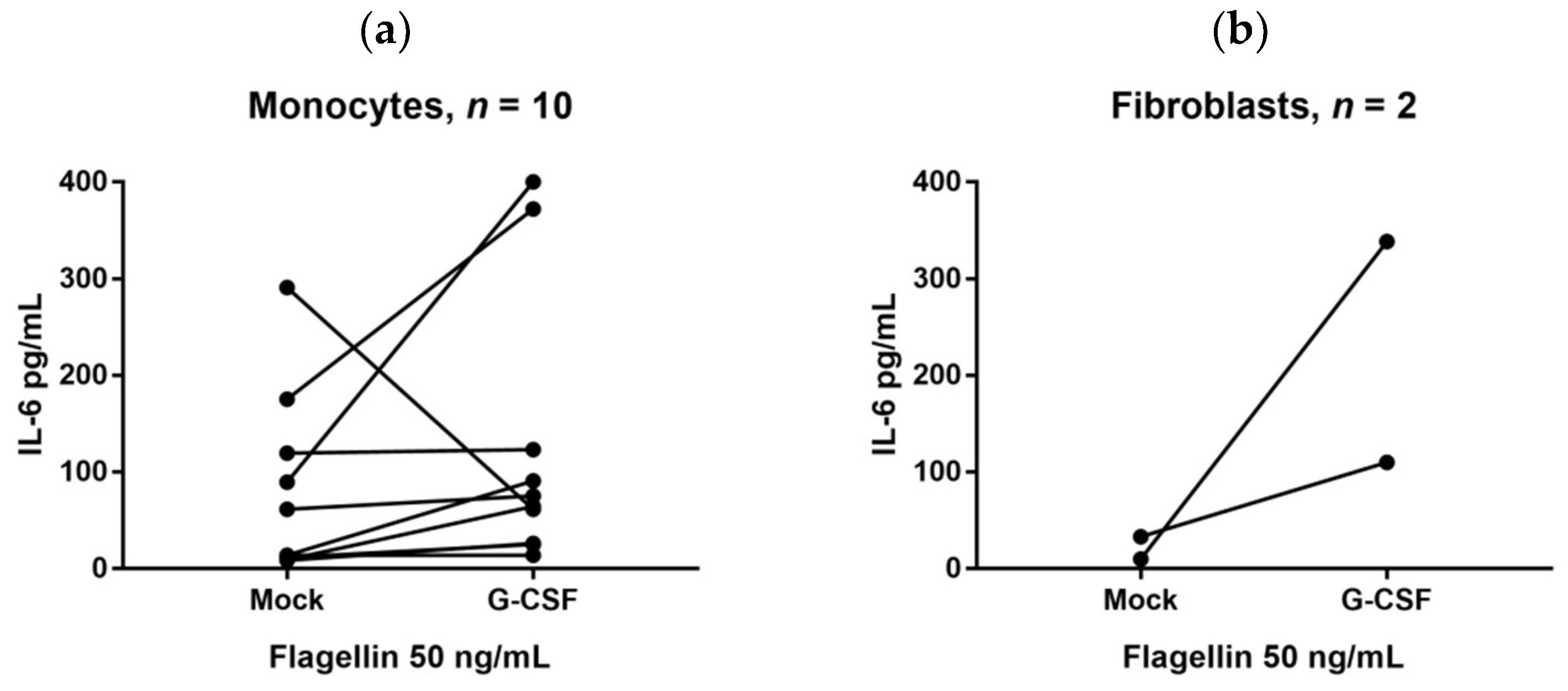
We also investigated whether G-CSF could increase IL-6 release by in vitro cultured cells. IL-6 can be released by several immunocompetent as well as mesenchymal cells [
27], and in our present study we included only monocytes together with fibroblasts and normal mesenchymal stem cells. We then used an in vitro model where monocytes and mesenchymal cells were cultured in the presence of TLR agonists; in our opinion this is a more physiological model than culture in medium alone because a wide range of endogenous TLR ligands have now been identified and are expected to be present in the in vivo microenvironments of these cells [
49]. A strong/significant alteration of the IL-6 release in the presence of G-CSF was defined as a two-fold alteration. It can be seen from
Figure 3 that the in vitro G-CSF effects on the monocyte release of IL-6 differed between healthy individuals (although increased IL-6 levels were most common). Previous in vivo studies also suggest that the effects of G-CSF on IL-6 vary between individuals, i.e., the effect of G-CSF therapy on systemic IL-6 levels of healthy stem cell donors differs and both increased, unaltered, and decreased systemic levels can be seen [
9,
19].
We investigated monocyte/fibroblast/mesenchymal stem cell release of IL-6 in an experimental model based on serum-free (i.e., possibly suboptimal) culture medium; this was use to minimize the risk of having TLR ligands in the medium. Our model is thus based on the presence of one ligand, whereas we would expect several endogenous TLR ligands to be present during physiological conditions. For these reasons we would emphasize that these results should be interpreted with great care and additional studies in other experimental models are needed to characterize, in greater detail, the effect of G-CSF IL-6 release by such cells.
Fibroblasts express a wide range of TLRs, and we also observed increased IL-6 release for both fibroblast cell lines in the presence of various TLR agonists. The constitutive IL-6 release by MSC was also increased by G-CSF. Taken together these observations suggest that various cells contribute to the IL-6 response during G-CSF therapy. This is similar to the IL-6/CRP responses during infections where both immunocompetent and mesenchymal cells contribute to these responses [
49].
Several observations suggest that immunoregulatory events early after stem cell transplantation are important for the outcome after ALLO-SCT, especially the risk of GVHD, for example, the need for early initiation of GVHD prophylaxis and the association between pretransplant conditioning, post-transplant G-CSF therapy, and risk of post-transplant outcome [
26]. Furthermore, IL-6 seems important in the development of immune-mediated complications after ALLO-SCT and is regarded a possible therapeutic target in GVHD [
27]. However, only future clinical studies can clarify whether G-CSF induced donor heterogeneity, including differences in acute phase reactions and IL-6 family cytokine levels, has any impact on the outcome for the allotransplant recipients.
4. Material and Methods
4.1. Patient Studies and Donor Samples
All studies were approved by the Regional Ethics Committee III, University of Bergen, Norway (REK VEST 2013/634 30 April 2013 and REK VEST 2015/1410, 02 July 2015). Only matched related donors (median age 49 years, range 18–77 years) mobilized with G-CSF 5 μg/kg twice daily were included. The donor and patient characteristics are given in
Table 5. These recipients/donors represent an unselected cohort. The routine GVHD prophylaxis was ciclosporin A plus methotrexate. All donors were selected according to the generally accepted suitability criteria [
50]. They were all healthy and without any signs of intercurrent disease at the times of evaluation, G-CSF therapy, and stem cell harvesting. Unless otherwise stated samples were collected between 8:00 am and 11:00 am in the morning. Twenty unselected donors were included in the cytokine studies (median age 51 years, range 25–73 years).
Stem cell collection was commenced after four days of G-CSF if the number of circulation CD34+ cells was sufficient. Samples were collected before and after 4 days of G-CSF therapy, immediately after leukapheresis, and approximately 24 h after start of leukapheresis. Graft supernatants were also collected. Samples were centrifuged at 1310× g, transferred onto cryotubes within 2 h after sampling, and stored at −70 °C until analyzed. Bio-Plex kits were used to analyze the levels of soluble mediators (Bio-Rad, Hercules, CA, USA), using the Luminex®200™ Bio-Rad platform. CRP was analyzed immediately after sampling by an immunoturbidimetric method (Roche; Basel, Switzerland); the lower detection limit being 1 mg/L.
4.2. Flow Cytometric Analysis
Peripheral blood and graft levels of immunocompetent cells were analyzed by flow cytometry. Briefly, peripheral blood mononuclear cells and graft cells were cryopreserved in DMSO and stored in liquid nitrogen until analyzed [
21,
51]. The cells were thawed and the near-IR fluorescent reactive dye (LIVE/DEAD Fixable Dead Cell Stain Kits, Molecular Probes, Eugene, OR, USA) was used for identification of viable cells. Cells were thereafter stained with CD3-PE-Cy7 (SK7), CD4-PerCP-Cy5.5 (RPA-T4), CD8-V500 (RPA-T8), CD16-Ax647 (3G8), CD19-PerCP-Cy5.5 (SJ25C1), and CD56-PE (B159) (all from Becton Dickinson Biosciences; BD Pharmingen, San Diego, CA, USA). We determined the numbers of CD3
+ T cells, CD4
+ and CD8
+ T cell subsets, B cells (CD19
+), and NK cells (CD16
+CD56
+) by using a FACS Canto II flow cytometer (Becton Dickinson Biosciences-Immunocytometry Systems; San Jose, CA, USA). The data were analyzed using FlowJo software version 10.2 (FlowJo LLC, Ashland, OR, USA). The monocyte levels were determined by multi-angle polarized scatter separation (MAPSS) optical flow cytometry (Cell-Dyn Sapphire analyzer; Abbot Diagnostics, Santa Clara, CA, USA).
4.3. In Vitro Culture of Monocytes and Fibroblasts
Samples were collected from healthy blood donors at Haukeland University Hospital. Monocytes from healthy donors were isolated from gradient-separated peripheral blood mononuclear cells (PBMCs) by negative selection using the human Monocyte Isolation Kit II (Miltenyi; Bergisch Gladbach, Germany). The isolation was performed according to the manufacturer’s instructions. Flow cytometric analysis verified that the purity was ≥95%. The Hs27 skin fibroblasts (ATCC CRL1634; Manassas, VA, USA) and HFL1 fetal lung fibroblasts (ATCC CRL153) were also examined together with mesenchymal stem cells (MSC) derived from a healthy individual (Cambrex BioScience; Walkersville, MD, USA).
Cells were cultured with each of the TLR agonists Pam3CSK4 (TLR1/2 agonist; tested at 1 and 5 ng/mL), lipopolysaccharide (LPS) (TLR4 agonist; 0.1, 0.5, 5, and 10 ng/mL), Flagellin (TLR5 agonist; 10 and 50 ng/mL), R837 (TLR7 > TLR8 agonist; 0.5 and 1.0 mg/mL), and R848 (TLR7/8 agonist; 50 and 100 ng/mL) (Invitrogen; San Diego, CA, USA), with and without G-CSF 50 ng/mL (Peprotech; Rocky Hill, NJ, USA). Monocytes (50,000 cells/mL, 1 mL/well; Multiwell™ 48 well culture plates, Falcon, Franklin, NJ, USA) were incubated in RPMI 1640 (Sigma-Aldrich; St. Louis, MO, USA) with TLR-agonists ± G-CSF for 24 h before harvesting of supernatants. Fibroblasts (10,000 cells/mL, 2 mL/well; Nunclon Delta Surface Thermofisher 6-well culture plates; Roskilde, Denmark) were incubated in Dulbecco’s Modified Eagle’s Medium (Sigma) for 24 h before TLR agonists/G-CSF were added and supernatants harvested 24 h later. MSCs (5000 cells/mL, 2 mL/well; Nunclon Delta Thermo-Fischer 6-well culture plates) were also incubated for 24 h in mesenchymal stem cell medium alone (MSCGM™; Lonza; Basel, Switzerland) for 24 h before TLR-agonists/G-CSF were added and supernatants harvested 24 h later. Cultures were incubated at 37 °C in a humidified atmosphere of 5% CO2. Supernatants were stored at −80 °C until IL-6 analysis (Quantikine ELISA kits; R&D Systems Minneapolis, MN, USA). These mediator analyses were performed in duplicates, and the variation between duplicates was generally less than 10%.
4.4. Statistical Analyses
Statistical analyses of clinical variables were performed using Stata Version 14 (StataCorp. 2009; Stata Statistical Software, College Station, TX, USA) and Graphpad Prism (GraphPad Software, Inc., La Jolla, CA, USA). Differences were regarded as statistically significant when p-values < 0.05.





{kind=link}
{kind=link}
{kind=link}