Supplementary Light Source Affects the Growth and Development of Codonopsis lanceolata Seedlings



Abstract
:1. Introduction
2. Results
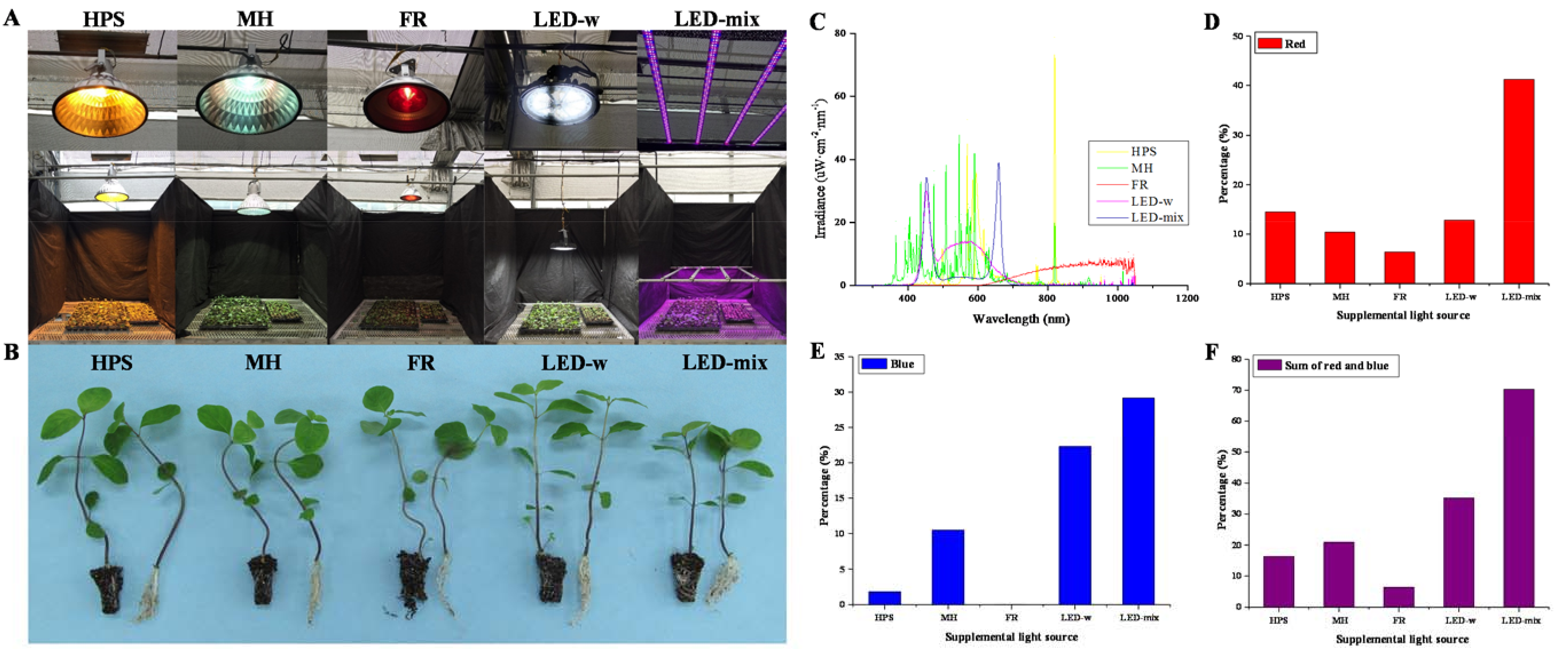
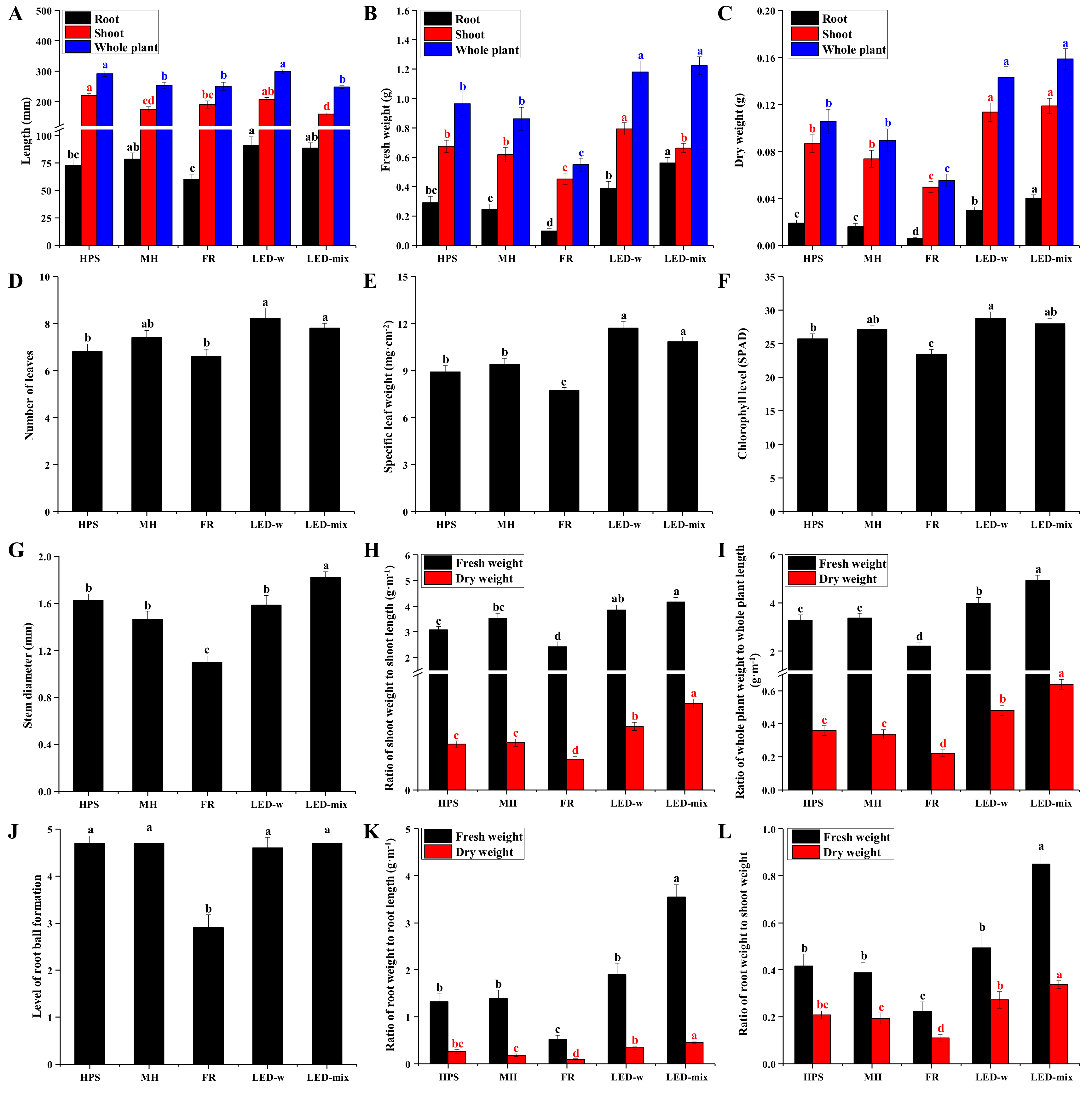
2.1. Morphology and Growth Parameters Analysis
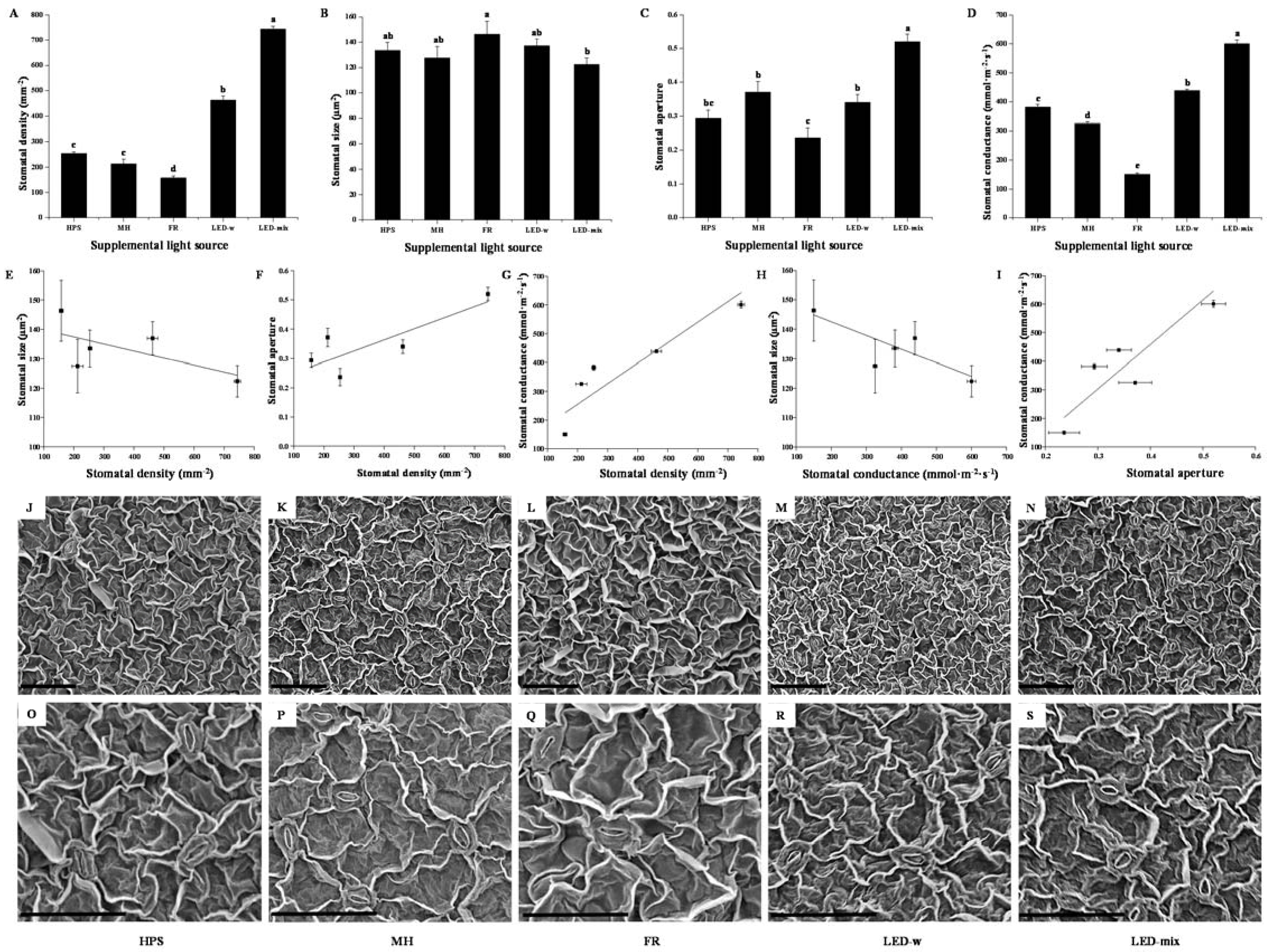
2.2. Stomatal Properties Analysis and Micrographs
2.3. Primary and Secondary Metabolites Analysis
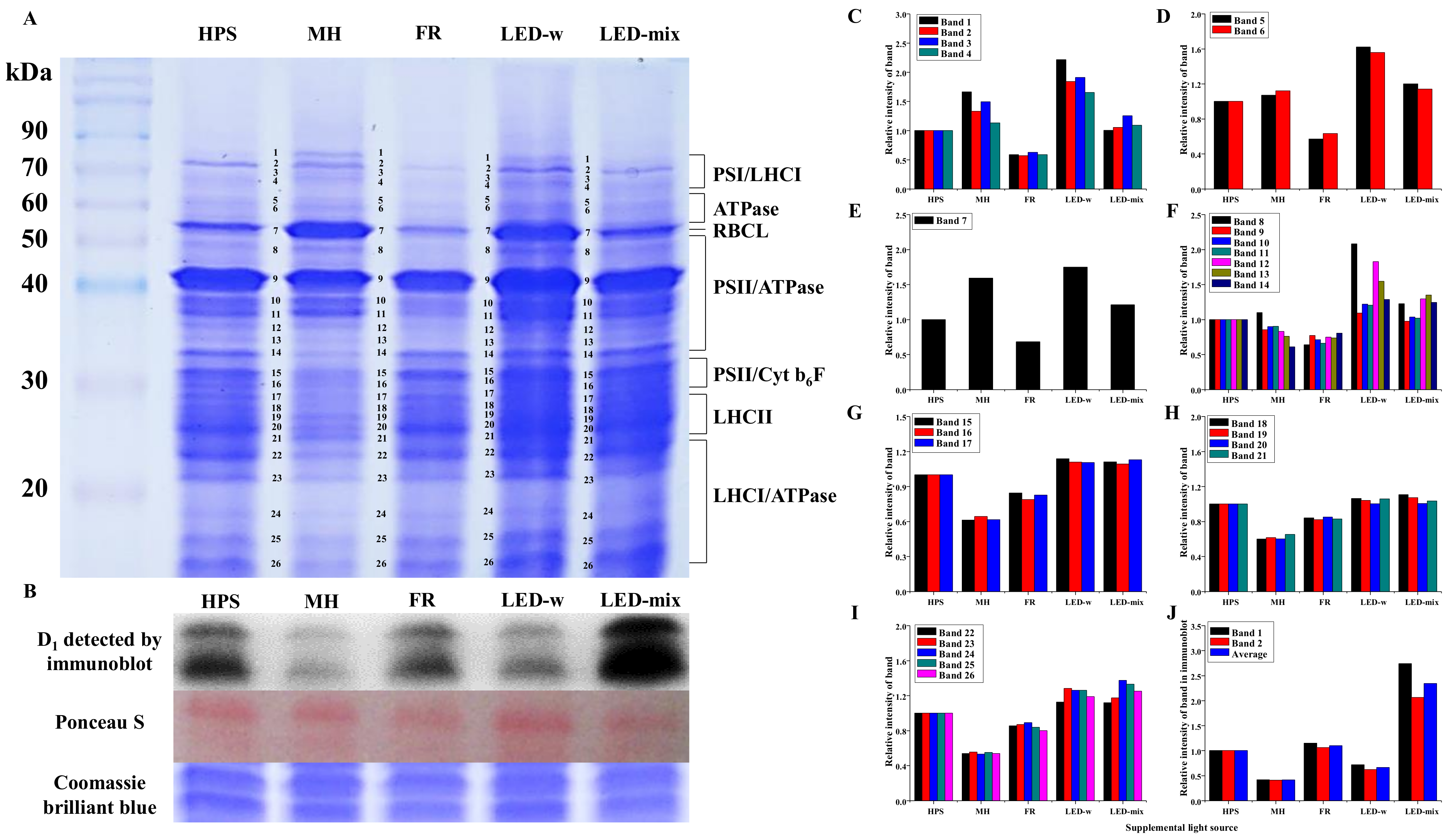
2.4. Chloroplast Protein Profiles and Immunoblot Analysis of D1 Protein (Anti-PsbA, PSII)
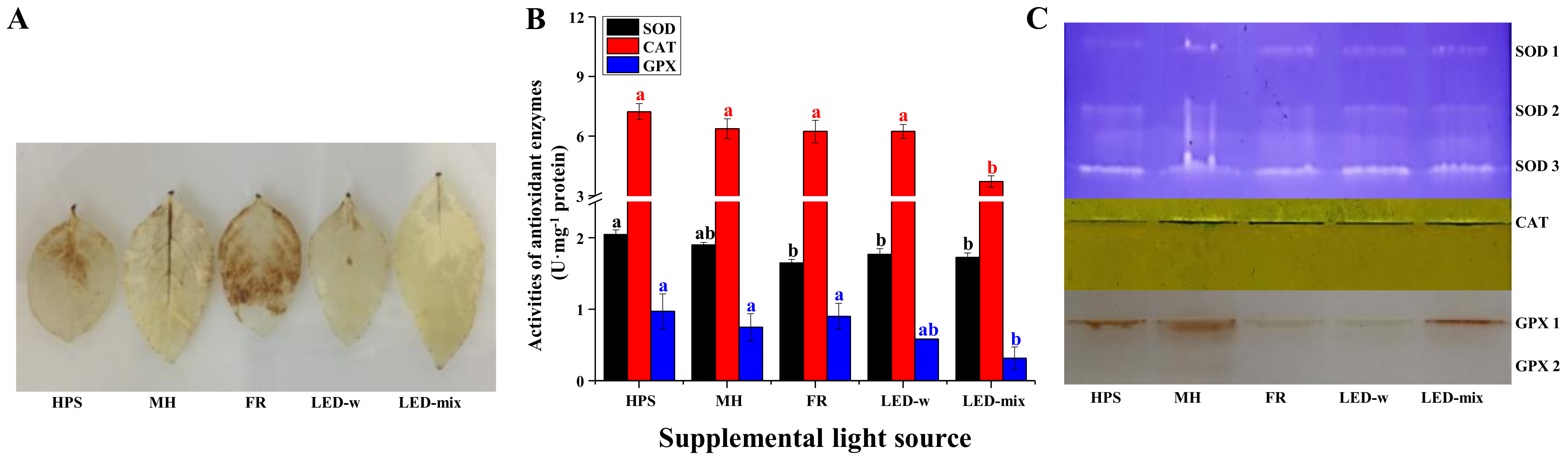
2.5. Analysis of H2O2 Localization and Antioxidant Enzyme Activities
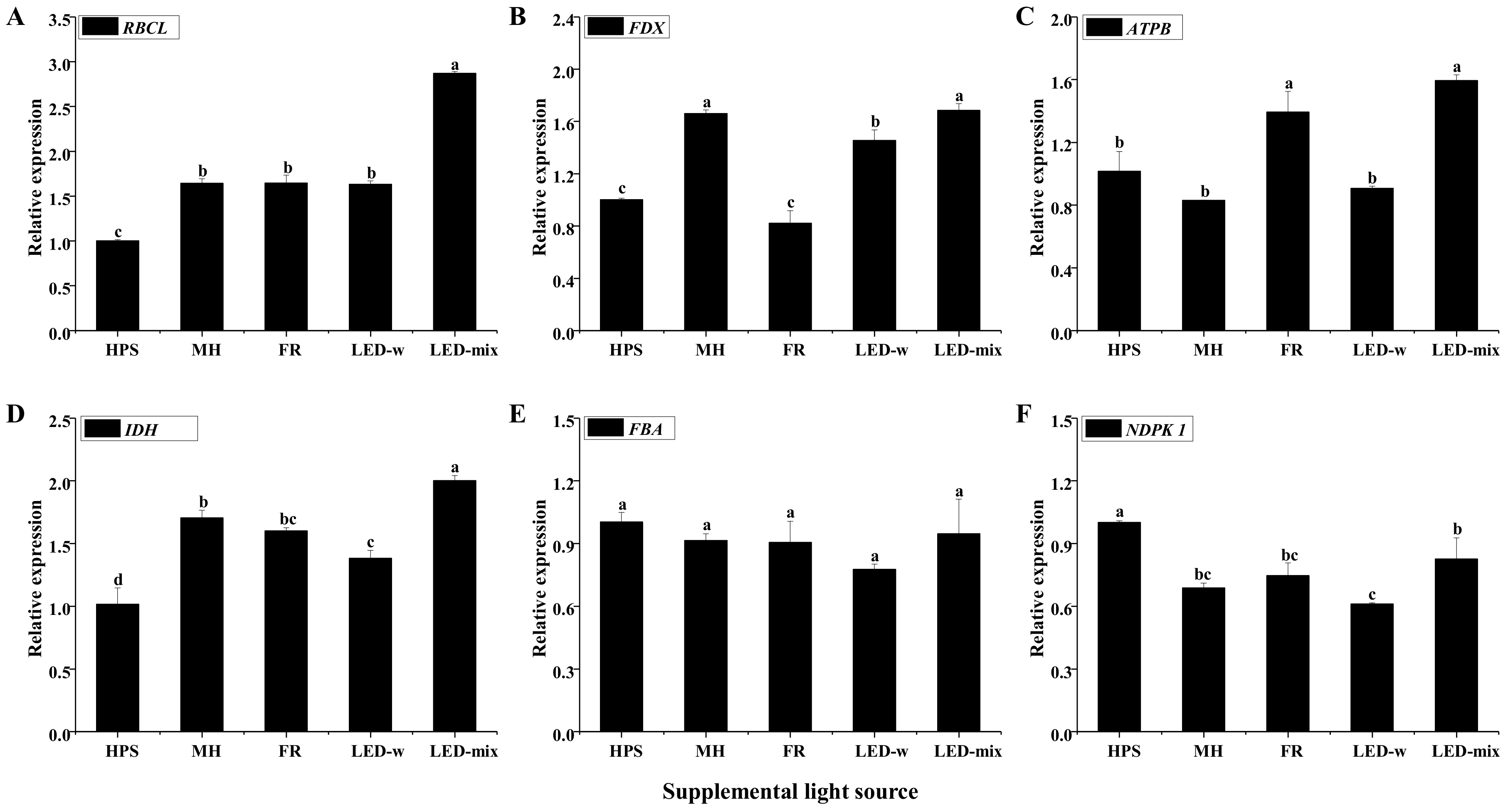
2.6. Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Culture Conditions and Supplementary Light Treatments
4.2. Scanning Electron Microscopic (SEM) Analysis of the Stomata
4.3. Chlorophyll, Carbohydrate, and Soluble Protein Analyses
4.4. Total Phenol and Flavonoid Analyses
4.5. Localization of Hydrogen Peroxide (H2O2)
4.6. Activities of Antioxidant Enzymes and Native Polyacrylamide Gel Electrophoresis (Native-PAGE) Analysis
4.7. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) Analysis and Immunoblot Assay
4.8. Primer Design and qRT-PCR
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Guo, W.L.; Gong, L.; Ding, Z.F.; Li, Y.D.; Li, F.X.; Zhao, S.P.; Liu, B. Genomic instability in phenotypically normal regenerants of medicinal plant Codonopsis lanceolata Benth. et Hook. f., as revealed by ISSR and RAPD markers. Plant Cell Rep. 2006, 25, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Piao, S. The Function and Utilization of Korean Traditional Vegetables; The Encyclopedia Press: Seoul, Korea, 1991. [Google Scholar]
- Jahangir, H.M.; Mi-Yeon, K.; Jong-Hoon, K.; Youl, C.J. Codonopsis lanceolata: A review of its therapeutic potentials. Phytother. Rep. 2016, 30, 347–356. [Google Scholar]
- Johkan, M.; Shoji, K.; Goto, F.; Hahida, S.; Yoshihara, T. Effect of green light wavelength and intensity on photomorphogenesis and photosynthesis in Lactuca sativa. Environ. Exp. Bot. 2012, 75, 128–133. [Google Scholar] [CrossRef]
- Haliapas, S.; Yupsanis, T.A.; Syros, T.D.; Kofidis, G.; Economou, A.S. Petunia × hybrida during transition to flowering as affected by light intensity and quality treatments. Acta Physiol. Plant. 2008, 30, 807. [Google Scholar] [CrossRef]
- Wang, H.; Gu, M.; Cui, J.; Shi, K.; Zhou, Y.; Yu, J. Effects of light quality on CO2 assimilation, chlorophyll-fluorescence quenching, expression of Calvin cycle genes and carbohydrate accumulation in Cucumis sativus. J. Photochem. Photobiol. B 2009, 96, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, J.; Qu, L.; Hager, J.; Chen, Z.; Zhao, H.; Deng, X.W. Light control of Arabidopsis development entails coordinated regulation of genome expression and cellular pathways. Plant Cell 2001, 13, 2589–2607. [Google Scholar] [CrossRef] [PubMed]
- Hogewoning, S.W.; Trouwborst, G.; Maljaars, H.; Poorter, H.; van Ieperen, W.; Harbinson, J. Blue light dose–responses of leaf photosynthesis, morphology, and chemical composition of Cucumis sativus grown under different combinations of red and blue light. J. Exp. Bot. 2010, 61, 3107–3117. [Google Scholar] [CrossRef] [PubMed]
- Macedo, A.F.; Leal-Costa, M.V.; Tavares, E.S.; Lage, C.L.S.; Esquibel, M.A. The effect of light quality on leaf production and development of in vitro-cultured plants of Alternanthera rasiliana Kuntze. Environ. Exp. Bot 2011, 70, 43–50. [Google Scholar] [CrossRef]
- Roni, M.Z.K.; Islam, M.S.; Shimasaki, K. Response of Eustoma leaf phenotype and photosynthetic performance to LED light quality. Horticulturae 2017, 3, 50. [Google Scholar] [CrossRef]
- Do Nascimento Vieira, L.; de Freitas Fraga, H.P.; dos Anjos, K.G.; Puttkammer, C.C.; Scherer, R.F.; da Silva, D.A.; Guerra, M.P. Light-emitting diodes (LED) increase the stomata formation and chlorophyll content in Musa acuminata (AAA) ‘nanicão corupá’ in vitro plantlets. Theor. Exp. Plant Phys. 2015, 27, 91–98. [Google Scholar] [CrossRef]
- Muneer, S.; Park, Y.G.; Jeong, B.R. Red and blue light emitting diodes (LEDs) participate in mitigation of hyperhydricity in in vitro-grown carnation genotypes (Dianthus caryophyllus). J. Plant Growth Regul. 2018, 37, 370–379. [Google Scholar] [CrossRef]
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Runkle, E.S. Far-red radiation promotes growth of seedlings by increasing leaf expansion and whole-plant net assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Walton, L.J.; Reid, D.M. Interaction of red to far red light ratio and ethylene in regulating stem elongation of Helianthus annuus. Plant Growth Regul. 2007, 51, 53–61. [Google Scholar] [CrossRef]
- Tran, L.; Jung, S. Effects of light-emitting diode irradiation on growth characteristics and regulation of porphyrin biosynthesis in rice seedlings. Int. J. Mol. Sci. 2017, 18, 641. [Google Scholar] [CrossRef] [PubMed]
- Yorio, N.C.; Goins, G.D.; Kagie, H.R.; Wheeler, R.M.; Sager, J.C. Improving spinach, radish, and lettuce growth under red light-emitting diodes (LEDs) with blue light supplementation. HortScience 2001, 36, 380–383. [Google Scholar] [PubMed]
- Lian, M.L.; Murthy, H.N.; Paek, K.Y. Effects of light emitting diodes (LEDs) on the in vitro induction and growth of bulblets of Lilium oriental hybrid ‘pesaro’. Sci. Hortic. 2002, 94, 365–370. [Google Scholar] [CrossRef]
- Nhut, D.T.; Takamura, T.; Watanabe, H.; Okamoto, K.; Tanaka, M. Responses of strawberry plantlets cultured in vitro under superbright red and blue light-emitting diodes (LEDs). Plant Cell Tissue Organ Cult. 2003, 73, 43–52. [Google Scholar] [CrossRef]
- Lee, S.H.; Tewari, R.K.; Hahn, E.J.; Paek, K.Y. Photon flux density and light quality induce changes in growth, stomatal development, photosynthesis and transpiration of Withania somnifera (L.) Dunal. plantlets. Plant Cell Tissue Organ Cult. 2007, 90, 141–151. [Google Scholar] [CrossRef]
- Shin, K.S.; Murthy, H.N.; Heo, J.W.; Hahn, E.J.; Paek, K.Y. The effect of light quality on the growth and development of in vitro cultured Doritaenopsis plants. Acta Physiol. Plant. 2008, 30, 339–343. [Google Scholar] [CrossRef]
- Wheeler, R.M. A historical background of plant lighting: An introduction to the workshop. HortScience 2008, 43, 1942–1943. [Google Scholar]
- Samuolienė, G.; Sirtautas, R.; Brazaitytė, A.; Duchovskis, P. LED lighting and seasonality effects antioxidant properties of baby leaf lettuce. Food Chem. 2012, 134, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food. Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Folta, K.M.; Koss, L.L.; McMorrow, R.; Kim, H.-H.; Kenitz, J.D.; Wheeler, R.; Sager, J.C. Design and fabrication of adjustable red-green-blue LED light arrays for plant research. BMC Plant Biol. 2005, 5, 17. [Google Scholar]
- Lin, K.-H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Elvidge, C.; Keith, D.; T Tuttle, B.; Baugh, K. Spectral identification of lighting type and character. Sensors 2010, 10, 3961–3988. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Park, Y.; Manivannan, A.; Soundararajan, P.; Jeong, B. Physiological and proteomic analysis in chloroplasts of Solanum lycopersicum L. Under silicon efficiency and salinity stress. Int. J. Mol. Sci. 2014, 15, 21803. [Google Scholar] [CrossRef] [PubMed]
- Muneer, S.; Kim, E.; Park, J.; Lee, J. Influence of green, red and blue light emitting diodes on multiprotein complex proteins and photosynthetic activity under different light intensities in lettuce leaves (Lactuca sativa L.). Int. J. Mol. Sci. 2014, 15, 4657. [Google Scholar] [CrossRef] [PubMed]
- Kügler, M.; Jänsch, L.; Kruft, V.; Schmitz, U.K.; Braun, H.P. Analysis of the chloroplast protein complexes by blue-native polyacrylamide gel electrophoresis (BN-PAGE). Photosynth. Res. 1997, 53, 35–44. [Google Scholar] [CrossRef]
- Apostol, K.G.; Dumroese, R.K.; Pinto, J.R.; Davis, A.S. Response of conifer species from three latitudinal populations to light spectra generated by light-emitting diodes and high-pressure sodium lamps. Can. J. For. Res. 2015, 45, 1711–1719. [Google Scholar] [CrossRef]
- Chen, C.C.; Huang, M.Y.; Lin, K.H.; Wong, S.L.; Huang, W.D.; Yang, C.M. Effects of light quality on the growth, development and metabolism of rice seedlings (Oryza sativa L.). Res. J. Biotechnol. 2014, 9, 15–24. [Google Scholar]
- Kasajima, S.Y.; Inoue, N.; Mahmud, R.; Kato, M. Developmental responses of wheat cv. Norin 61 to fluence rate of green light. Plant Prod. Sci. 2008, 11, 76–81. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Wu, M.C.; Hou, C.Y.; Jiang, C.M.; Wang, Y.T.; Wang, C.Y.; Chen, H.H.; Chang, H.M. A novel approach of LED light radiation improves the antioxidant activity of pea seedlings. Food Chem. 2007, 101, 1753–1758. [Google Scholar] [CrossRef]
- Cope, K.R.; Bugbee, B. Spectral effects of three types of white light-emitting diodes on plant growth and development: Absolute versus relative amounts of blue light. HortScience 2013, 48, 504–509. [Google Scholar]
- Islam, M.A.; Kuwar, G.; Clarke, J.L.; Blystad, D.R.; Gislerød, H.R.; Olsen, J.E.; Torre, S. Artificial light from light emitting diodes (LEDs) with a high portion of blue light results in shorter poinsettias compared to high pressure sodium (HPS) lamps. Sci. Hortic. 2012, 147, 136–143. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; Carlos, E.J. Phenolic compounds and related enzymes as determinants of quality in fruits and vegetables. J. Sci. Food. Agric. 2001, 81, 853–876. [Google Scholar] [CrossRef]
- Pala, Ç.U.; Toklucu, A.K. Effect of UV-C light on anthocyanin content and other quality parameters of pomegranate juice. J. Food Compos. Anal. 2011, 24, 790–795. [Google Scholar] [CrossRef]
- Connor, A.M.; Stephens, M.J.; Hall, H.K.; Alspach, P.A. Variation and heritabilities of antioxidant activity and total phenolic content estimated from a red raspberry factorial experiment. J. Am. Soc. Hortic. Sci. 2005, 130, 403–411. [Google Scholar]
- Nehir, S.; El, S.N.; Taş, A.A. Antioxidant activity of some foods containing phenolic compounds. Int. J. Food Sci. Nutr. 2001, 52, 501–508. [Google Scholar]
- Andrea, L.; Judit, H. Antioxidant properties of commercial alcoholic and nonalcoholic beverages. Food Nahrung 2003, 47, 79–86. [Google Scholar]
- Surh, Y.J. Molecular mechanisms of chemopreventive effects of selected dietary and medicinal phenolic substances. Mutat. Res. 1999, 428, 305–327. [Google Scholar] [CrossRef]
- Bliznikas, Z.; Žukauskas, A.; Samuolienė, G.; Viršile, A.; Brazaityte, A.; Jankauskiene, J.; Duchovskis, P.; Novičkovas, A. Effect of supplementary pre-harvest LED lighting on the antioxidant and nutritional properties of green vegetables. Acta Hortic. 2012, 939, 85–91. [Google Scholar] [CrossRef]
- Luthria, D.L.; Mukhopadhyay, S.; Krizek, D.T. Content of total phenolics and phenolic acids in tomato (Lycopersicon esculentum Mill.) fruits as influenced by cultivar and solar UV radiation. J. Food Compos. Anal. 2006, 19, 771–777. [Google Scholar] [CrossRef]
- García-Macías, P.; Ordidge, M.; Vysini, E.; Waroonphan, S.; Battey, N.H.; Gordon, M.H.; Hadley, P.; John, P.; Lovegrove, J.A.; Wagstaffe, A. Changes in the flavonoid and phenolic acid contents and antioxidant activity of red leaf lettuce (Lollo rosso) due to cultivation under plastic films varying in ultraviolet transparency. J. Agric. Food Chem. 2007, 55, 10168–10172. [Google Scholar] [CrossRef] [PubMed]
- Bukhov, N.G. Dynamic light regulation of photosynthesis (a review). Russ. J. Plant Physl. 2004, 51, 742–753. [Google Scholar] [CrossRef]
- Sack, L.; Buckley, T.N. The developmental basis of stomatal density and flux. Plant Physiol. 2016, 171, 2358–2363. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Hahn, E.J.; Heo, J.W.; Paek, K.Y. Effects of LEDs on net photosynthetic rate, growth and leaf stomata of chrysanthemum plantlets in vitro. Sci. Hortic. 2004, 101, 143–151. [Google Scholar] [CrossRef]
- Heo, J.; Lee, C.; Chakrabarty, D.; Paek, K. Growth responses of marigold and salvia bedding plants as affected by monochromic or mixture radiation provided by a light-emitting diode (LED). Plant Growth Regul. 2002, 38, 225–230. [Google Scholar] [CrossRef]
- Savvides, A.; Fanourakis, D.; Ieperen, W. Co-ordination of hydraulic and stomatal conductances across light qualities in cucumber leaves. J. Exp. Bot. 2011, 63, 1135–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shimazaki, K.-I.; Doi, M.; Assmann, S.M.; Kinoshita, T. Light regulation of stomatal movement. Annu. Rev. Plant Biol. 2007, 58, 219–247. [Google Scholar] [CrossRef] [PubMed]
- Talbott, L.D.; Zhu, J.; Han, S.W.; Zeiger, E. Phytochrome and blue light-mediated stomatal opening in the orchid, paphiopedilum. Plant Cell Physiol. 2002, 43, 639–646. [Google Scholar] [CrossRef] [PubMed]
- Franks, P.J.; Beerling, D.J. Maximum leaf conductance driven by CO2 effects on stomatal size and density over geologic time. Proc. Natl. Acad. Sci. USA 2009, 106, 10343–10347. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, P.; Buchanan, B.B. The ferredoxin/thioredoxin system of oxygenic photosynthesis. Antioxid. Redox Sign. 2008, 10, 1235–1274. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Shimizu, H.; Shikanai, T. The chloroplast NAD(P)H dehydrogenase complex interacts with photosystem I in Arabidopsis. J. Biol. Chem. 2008, 283, 34873–34879. [Google Scholar] [CrossRef] [PubMed]
- Sirpiö, S.; Allahverdiyeva, Y.; Holmström, M.; Khrouchtchova, A.; Haldrup, A.; Battchikova, N.; Aro, E.-M. Novel nuclear-encoded subunits of the chloroplast NAD(P)H dehydrogenase complex. J. Biol. Chem. 2009, 284, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Droux, M.; Jacquot, J.P.; Miginac-Maslow, M.; Gadal, P.; Huet, J.C.; Crawford, N.A.; Yee, B.C.; Buchanan, B.B. Ferredoxin-thioredoxin reductase, an iron-sulfur enzyme linking light to enzyme regulation in oxygenic photosynthesis: Purification and properties of the enzyme from C3, C4, and cyanobacterial species. Arch. Biochem. Biophys. 1987, 252, 426–439. [Google Scholar] [CrossRef]
- Falciatore, A.; Bowler, C. The evolution and function of blue and red light photoreceptors. Curr. Top. Dev. Biol. 2005, 68, 317–350. [Google Scholar] [PubMed]
- Barber, J.; Kühlbrandt, W. Photosystem II. Curr. Opin. Struct. Biol. 1999, 9, 469–475. [Google Scholar] [CrossRef]
- Diner, B.A.; Babcock, G.T. Structure, dynamics, and energy conversion efficiency in photosystem II. In Oxygenic Photosynthesis: The Light Reactions; Yocum, C.F., Heichel, I.F., Eds.; Springer: Dordrecht, The Netherlands, 1996; pp. 213–247. [Google Scholar]
- Järvi, S.; Suorsa, M.; Paakkarinen, V.; Aro, E.M. Optimized native gel systems for separation of thylakoid protein complexes: Novel super-and mega-complexes. Biochem. J. 2011, 439, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Nelson, N.; Yocum, C.F. Structure and function of photosystems I and II. Annu. Rev. Plant Biol. 2006, 57, 521–565. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.; Senior, A.E. ATP synthesis driven by proton transport in F1F0-ATP synthase. FEBS Lett. 2003, 545, 61–70. [Google Scholar] [CrossRef]
- Zhang, H.; Kurisu, G.; Smith, J.L.; Cramer, W.A. A defined protein–detergent–lipid complex for crystallization of integral membrane proteins: The cytochrome B6F complex of oxygenic photosynthesis. Proc. Natl. Acad. Sci. USA 2003, 100, 5160–5163. [Google Scholar] [CrossRef] [PubMed]
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions, and possibilities for a better enzyme. Annu. Rev. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Wu, Q.; Shen, Z.; Xia, K.; Cui, J. Effects of light quality on the chloroplastic ultrastructure and photosynthetic characteristics of cucumber seedlings. Plant Growth Regul. 2014, 73, 227–235. [Google Scholar] [CrossRef]
- Gálvez, S.; Gadal, P. On the function of the NADP-dependent isocitrate dehydrogenase isoenzymes in living organisms. Plant Sci. 1995, 105, 1–14. [Google Scholar] [CrossRef]
- Mininno, M.; Brugière, S.; Pautre, V.; Gilgen, A.; Ma, S.; Ferro, M.; Tardif, M.; Alban, C.; Ravanel, S. Characterization of chloroplastic fructose 1,6-bisphosphate aldolases as lysine-methylated proteins in plants. J. Biol. Chem. 2012, 287, 21034–21044. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Tepperman, J.M.; Hudson, M.E.; Khanna, R.; Zhu, T.; Chang, S.H.; Wang, X.; Quail, P.H. Expression profiling of phyB mutant demonstrates substantial contribution of other phytochromes to red-light-regulated gene expression during seedling de-etiolation. Plant J. 2004, 38, 725–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thum, K.E.; Shin, M.J.; Palenchar, P.M.; Kouranov, A.; Coruzzi, G.M. Genome-wide investigation of light and carbon signaling interactions in Arabidopsis. Genome Biol. 2004, 5, R10. [Google Scholar] [CrossRef] [PubMed]
- Yukio, M.; Shin-ichirou, S.; Tadahito, S. Phytochrome-mediated control of respiratory activities of mitochondria in cotyledons of dark-grown cucumber seedlings. Physiol. Plant. 1993, 89, 295–300. [Google Scholar]
- Rasmusson, A.G.; Escobar, M.A. Light and diurnal regulation of plant respiratory gene expression. Physiol. Plant. 2007, 129, 57–67. [Google Scholar] [CrossRef]
- Muneer, S.; Ko, C.H.; Wei, H.; Chen, Y.; Jeong, B.R. Physiological and proteomic investigations to study the response of tomato graft unions under temperature stress. PLoS ONE 2016, 11, e0157439. [Google Scholar] [CrossRef] [PubMed]
- Song, X.S.; Tiao, C.L.; Shi, K.; Mao, W.H.; Ogweno, J.O.; Zhou, Y.H.; Yu, J.Q. The response of antioxidant enzymes in cellular organelles in cucumber (Cucumis sativus L.) leaves to methyl viologen-induced photo-oxidative stress. Plant Growth Regul. 2006, 49, 85–93. [Google Scholar] [CrossRef]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Suzuki, N.; Miller, G.; Tognetti, V.B.; Vandepoele, K.; Gollery, M.; Shulaev, V.; van Breusegem, F. ROS signaling: The new wave? Trends Plant Sci. 2011, 16, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Ranieri, A.; Castagna, A.; Baldan, B.; Soldatini, G.F. Iron deficiency differently affects peroxidase isoforms in sunflower. J. Exp. Bot. 2001, 52, 25–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeoka, S.; Ishikawa, T.; Tamoi, M.; Miyagawa, Y.; Takeda, T.; Yabuta, Y.; Yoshimura, K. Regulation and function of ascorbate peroxidase isoenzymes. J. Exp. Bot. 2002, 53, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhang, Y.; Zhang, Y. Scavenger enzyme activities in subcellular fractions of white clover (Trifolium repens L.) under PEG-induced water stress. J. Plant Growth Regul. 2008, 27, 387–393. [Google Scholar] [CrossRef]
- Fukamatsu, Y.; Yabe, N.; Hasunuma, K. Arabidopsis NDK1 is a component of ROS signaling by interacting with three catalases. Plant Cell Physiol. 2003, 44, 982–989. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.S.; Hyun-Gyun, Y.; Hoon, K.G.; Weibiao, Z. Application of light-emitting diodes in food production, postharvest preservation, and microbiological food safety. Compr. Rev. Food Sci. Food Saf. 2015, 14, 719–740. [Google Scholar]
- Doheny-Adams, T.; Hunt, L.; Franks, P.J.; Beerling, D.J.; Gray, J.E. Genetic manipulation of stomatal density influences stomatal size, plant growth and tolerance to restricted water supply across a growth carbon dioxide gradient. Philos. Trans. R. Soc. B 2012, 367, 547–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monda, K.; Araki, H.; Kuhara, S.; Ishigaki, G.; Akashi, R.; Negi, J.; Kojima, M.; Sakakibara, H.; Takahashi, S.; Hashimoto-Sugimoto, M.; et al. Enhanced stomatal conductance by a spontaneous Arabidopsis tetraploid, Me-0, results from increased stomatal size and greater stomatal aperture. Plant Physiol. 2016, 170, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.X.; Xue, J.Q.; Wang, S.L.; Xue, Y.Q.; Zhang, P.; Jiang, H.D.; Zhang, X.X. Proteomic analysis of tree peony (Paeonia ostii ‘Feng Dan’) seed germination affected by low temperature. J. Plant Physiol. 2018, 224–225, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, A.; Soundararajan, P.; Park, Y.; Jeong, B. Chemical elicitor-induced modulation of antioxidant metabolism and enhancement of secondary metabolite accumulation in cell suspension cultures of Scrophularia kakudensis Franch. Int. J. Mol. Sci. 2016, 17, 399. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Manivannan, A.; Cho, Y.S.; Jeong, B.R. Exogenous supplementation of silicon improved the recovery of hyperhydric shoots in Dianthus caryophyllus L. by stabilizing the physiology and protein expression. Front. Plant Sci. 2017, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Khanna, R.; Carle, C.M.; Quail, P.H. Phytochrome induces rapid PIF5 phosphorylation and degradation in response to red-light activation. Plant Physiol. 2007, 145, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Beruto, M.; Xue, J.; Zhu, F.; Liu, C.; Yan, Y.; Zhang, X. Molecular cloning and potential function prediction of homologous SOC1 genes in tree peony. Plant Cell Rep. 2015, 34, 1459–1471. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2−ΔΔCt method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71–85. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Supplementary Light Source | Irradiance (μW·cm−2) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| UV (250−380 nm) | Violet (381–450 nm) | Blue (451–495 nm) | Green (496–570 nm) | Yellow (571–590 nm) | Orange (591–620 nm) | Red (621–710 nm) | FR (711–850 nm) | IR (851–1050 nm) | |
| HPS | 11.0 | 46.8 | 54.9 | 356.0 | 509.5 | 906.1 | 434.9 | 655.7 | 25.2 |
| MH | 118.5 | 703.2 | 314.1 | 788.9 | 296.6 | 258.6 | 311.6 | 147.5 | 61.0 |
| FR | 1.9 | 0.7 | 0.2 | 0.5 | 0.5 | 4.5 | 191.6 | 934.5 | 1865.6 |
| LED-w | 12.3 | 329.2 | 668.7 | 917.8 | 268.2 | 345.5 | 385.2 | 33.4 | 39.8 |
| LED-mix | 8.5 | 445.6 | 874.7 | 258.0 | 69.7 | 93.6 | 1235.6 | 8.8 | 5.6 |
| Supplementary Light Source | Guard Cell Length (µm) | Width of Guard Cell Pair (µm) | Pore Length (µm) | Pore Width (µm) |
|---|---|---|---|---|
| HPS | 16.9 ± 0.4 a | 7.9 ± 0.3 b | 11.2 ± 0.3 a | 3.3 ± 0.3 a |
| MH | 16.0 ± 0.4 a | 7.9 ± 0.4 b | 8.9 ± 0.3 b | 3.4 ± 0.3 a |
| FR | 15.8 ± 0.6 a | 8.9 ± 0.4 a | 7.8 ± 0.3 cd | 1.8 ± 0.2 b |
| LED-w | 15.7 ± 0.4 a | 8.7 ± 0.4 ab | 7.6 ± 0.3 d | 2.4 ± 0.2 b |
| LED-mix | 15.6 ± 0.4 a | 7.8 ± 0.3 b | 7.9 ± 0.3 c | 4.0 ± 0.2 a |
| Supplementary Light Source | Soluble Sugar Content (% of Fresh Mass) | Starch Content (% of Fresh Mass) | Protein Content (% of Fresh Mass) | Total Phenol Content (µg·g−1 Fresh Mass) | Total Flavonoid Content (µg·g−1 Fresh Mass) |
|---|---|---|---|---|---|
| HPS | 2.66 ± 0.08 b | 1.51 ± 0.06 ab | 0.55 ± 0.03 b | 0.53 ± 0.03 b | 0.65 ± 0.04 b |
| MH | 2.51 ± 0.08 b | 1.35 ± 0.03 b | 0.71 ± 0.01 a | 0.30 ± 0.02 c | 0.37 ± 0.02 c |
| FR | 1.69 ± 0.15 c | 1.15 ± 0.05 c | 0.74 ± 0.03 a | 0.28 ± 0.02 c | 0.35 ± 0.02 c |
| LED-w | 1.52 ± 0.10 c | 1.50 ± 0.07 ab | 0.68 ± 0.04 a | 1.10 ± 0.06 a | 1.35 ± 0.07 a |
| LED-mix | 3.28 ± 0.11 a | 1.62 ± 0.01 a | 0.70 ± 0.01 a | 0.97 ± 0.02 a | 1.18 ± 0.02 a |
| Gene | Forward Primers | Reverse Primers |
|---|---|---|
| β-Actin | 5′-CGAGAAGAGCTACGAGCTACCCGATGG-3′ | 5′-CTCGGTGCTAGGGCAGTGATCTCTTTGCT-3′ |
| ATPB | 5′-TGCCTTCTGCTGTGGGTTAT-3′ | 5′-GGTCGGTCAAATCGTCTGC-3′ |
| FDX | 5′-CTTCGGCGTTTCTTCGT-3′ | 5′-CTGCCAAACCCTTGATAACT-3′ |
| RBCL | 5′-GCTTACCCATTAGACCTTT-3′ | 5′-GGGACGACCATACTTGTT-3′ |
| FBA | 5′-ACAGGTGGGCTCTTC GTG-3′ | 5′-CCTTGGGTGGTGGTTTCA-3′ |
| IDH | 5′-TGACGGAGGTTATGTATGG-3′ | 5′-AATGCTGTTCGTGCTGGT-3′ |
| NDPK 1 | 5′-AGAGGCTTGGTTGGTGAGA-3′ | 5′-AGAGGCAGCAGGGTTTGT-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, X.; Liu, Y.; Jeong, H.K.; Jeong, B.R. Supplementary Light Source Affects the Growth and Development of Codonopsis lanceolata Seedlings. Int. J. Mol. Sci. 2018, 19, 3074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103074
Ren X, Liu Y, Jeong HK, Jeong BR. Supplementary Light Source Affects the Growth and Development of Codonopsis lanceolata Seedlings. International Journal of Molecular Sciences. 2018; 19(10):3074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103074
Chicago/Turabian StyleRen, Xiuxia, Ya Liu, Hai Kyoung Jeong, and Byoung Ryong Jeong. 2018. "Supplementary Light Source Affects the Growth and Development of Codonopsis lanceolata Seedlings" International Journal of Molecular Sciences 19, no. 10: 3074. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms19103074